tissue interaction ans d the initiation and avian...
TRANSCRIPT

/ . Embryol. exp. Morph. Vol. 58, pp. 251-264, 1980 2 5 1Printed in Great Britain © Company of Biologists Limited 1980
Tissue interactions and the initiationof osteogenesis and chondrogenesis in the neural
crest-derived mandibular skeleton of theembryonic mouse as seen in isolated murine
tissues and in recombinations of murineand avian tissues
By BRIAN K. HALL1
From the Department of Biology, Life Sciences Centre,Dalhousie University, Halifax
SUMMARYMandibular processes from 9- to 13-day-old embryonic mice formed both bone and
cartilage when grafted to the chorioallantoic membranes of host embryonic chicks. Isolatedectomesenchyme, taken from 9-day-old embryos did not form bone or cartilage, while olderectomesenchyme formed both. Recombination of the epithelial and ectomesenchymal com-ponents confirmed that the presence of the epithelium was a sufficient stimulus for theinitiation of both chondro- and osteogenesis. Recombinations between components of mouseand chick mandibular processes showed that 9-day-old mouse ectomesenchyme could re-spond to chick epithelium but that, although older murine epithelia could initiate osteo-genesis from the avian ectomesenchyme, epithelia from 9-day-old embryos could not. Theseresults indicated that an epithelial-ectomesenchymal interaction was responsible for theinitiation of both osteo- and chondrogenesis within the mandibular arch of the mouse;that the interaction began at 10 days of gestation; that the ectomesenchyme was capable ofresponding at 9 days, but that the epithelium did not acquire its ability to act on the ecto-mesenchyme until 10 days of gestation.
INTRODUCTION
Both the cartilaginous and the bony elements in the mandibular skeletons ofvertebrate embryos form from ectomesenchymal cells derived from the embryo-nic neural crest. These cells have their origin within the crests of the neural folds.During neurulation these neural crest cells migrate away from the neural folds.Those cells which originate in the cranial neural crest become preferentiallylocalized within the primordia of the cranio-facial skeleton. The ability of thesecells to form the cartilages and bones of the craniofacial skeleton depends upon
1 Author's address; Department of Biology, Life Sciences Centre, Dalhousie University,Halifax, Nova Scotia, Canada, B3H 4J1.
17 EMB 58

252 B. K. HALL
interactions which they undergo, either during or after, their migration. Thus,in the embryo of the lamprey, Lampetrafluviatilis L., contact with the endodermis required before chondrogenesis can be initiated (Damas, 1944; Newth, 1951,1956). While it is known that cranio-facial skeletal elements are of neural crestorigin in the fishes (Jollie, 1971; Berkovitz & Moore, 1974; Schaeffer, 1977), therole of tissue interactions in their genesis has not been studied. Interaction ofneural crest cells with pharyngeal endoderm is a prerequisite for the initiation ofchondrogenesis in the visceral skeleton of the urodele amphibians (Wagner, 1949;Wilde, 1955; Cusimano-Carollo, 1963, 1972; Drews, Kocher-Becker & Drews,1972; Epperlein, 1974; Corsin, 1975). However, although neural crest cells areknown to form bone in the urodeles (de Beer, 1947; Cassin & Capuron, 1977),tissue interactions have not been studied. Nor is there any information for theanuran amphibians (see Horstadius, 1950 and Hall, 1978a for a discussion).
The initiation of intramembranous osteogenesis within the scleral, maxillaryand mandibular skeletons of the domestic fowl results from inductive inter-actions between scleral, maxillary, or mandibular epithelia and post-migratoryectomesenchymal cells from the neural crest (Murray, 1943; Tyler & Hall, 1977;Tyler, 1978; Hall, 1978c, b, 1980; Bradamante & Hall, 1980; Tyler & McCobb,1980). Pre-migratory neural crest cells must interact, (a) with cranial epithelialectoderm if they are to be able to form Meckel's cartilage later in development(Hall & Tremaine, 1979; Bee & Thorogood, 1980), or (b) with the pigmentedretinal epithelium if they are to form scleral cartilage (Reinbold, 1968; Newsome,1972, 1976; Hall, 1978a).
Although the details vary, epithelial-mesenchymal interactions emerge asimportant prerequisites for the initiation of osteogenesis and chondrogenesis inthe sub-mammalian vertebrates. The possible existence of similar interactionsin the mammalian embryo has not been explored. Indeed, even the origin of themammalian cranio-facial skeleton from neural crest cells has not been substan-tiated experimentally. There is histological evidence for spurs of ectodermalcells leading into the visceral arches (Bartelmez, 1952; Mulnard, 1955; O'Rahilly,1965; Mayer, 1973); histochemical visualization of presumed neural crest cells(Milaire, 1959; 1974); indications that high doses of vitamin A both arrestmigration of cranial neural crest cells (Morriss & Thorogood, 1978) and pro-duce first arch defects such as Treacher Collins syndrome (Poswillo, 1974), andthe evidence that ectoderm or teratocarcinomas isolated from rat embryos formboth cartilage and bone (Levak-Svajger & Svajger, 1971; Damjanov, Solter& Serman, 1973) - all of which provide circumstantial evidence for contri-butions to the facial skeleton from the neural crest.
The only study known to the author which explores the role of tissue inter-actions in the development of the mammalian mandibular skeleton is that ofSvajger and Levak-Svajger (1976). These workers separated first branchial archmesenchyme of 13-day rat embryos from their epithelia, organ cultured themesenchyme, and observed the subsequent development of bone and cartilage.

Skeletogenesis in the murine mandible 253If an epithelial-mesenchymal interaction was involved, it must have taken placebefore thirteen days of gestation.
The aim of the present study was to determine whether an epithelial-mesen-chymal (ectomesenchymal) interaction was involved in the initiation ofchondro-genesis and/or osteogenesis within the mandibular skeleton of the embryonicmice. To this end, whole mandibular processes, and mandibular ectomesen-chyme, separated from the mandibular epithelium by treatment with proteo-lytic enzymes, were grafted to the chorioallantoic membranes of host embryonicchicks. (This vascularized site is preferred over organ culture when studyingosteogenesis.) The separated components were also either recombined with oneanother or combined with components isolated from the mandibular processesof embryonic chicks and grafted to chorioallantoic membranes. Histologicalanalysis was used to document the presence or absence of cartilage and bone inthe recovered grafts.
MATERIALS AND METHODSAnimals
ICR Swiss albino mice obtained from Bio-breeding Laboratories of Canada,Ottawa, Ontario, were used as the experimental animal. Mice were weaned at4 weeks of age, housed with three or four of the same sex per cage, and fedPurina lab. chow and water ad Hbidum. Matings were carried out using animalswhich were at least 8 weeks old. Each male was placed with from one to threefemales overnight (16.00 to 08.00 hours), 08.00 hours was designated as thecommencement of day zero of the pregnancy. Pregnancy was determined bypresence of a vaginal plug and by weight gain, as measured on alternate days.
EmbryosPregnant females were killed by an overdose of ether, the embryos removed by
dissection under sterile conditions and aged using the morphological series ofstages of Griineberg (1943). Embryos of 9-13 gestational days were used in thisstudy. The 9-day-old embryos had well developed mandibular processes, lackedposterior limb buds, and had unsegmented posterior somitic mesoderm. By 13days of gestation the embryos were recognizably murine, with five rows ofwhiskers containing hair follicles, footplates on the limbs and marked segmen-tation to the distal tips of the tails.
Mandibular processes
Mandibular processes were dissected from these embryos and either graftedintact, or separated into their epithelial and ectomesenchymal components andthen grafted. Separation was achieved by placing the processes into a solution oftrypsin and pancreatin in calcium- and magnesium-free Tyrode's solution(257 mg bovine pancreatic trypsin + 43 mg porcine pancreatic pancreatin/10
17-2

254 B. K. HALL
ml. both obtained from BDH Chemicals, Toronto, Ontario) for 2 h at 4° C.The mandibular processes were then placed into a solution of the syntheticculture medium BGJb (GIBCO, Grand Islands, N.Y.) and Horse Serum (1:1) tostop the enzymic digestion and the epithelia separated from the ectomesenchymeby microdissection using sharpened hypodermic needles. A similar separationwas performed on mandibular processes obtained from embryonic chicks whichhad been incubated for 4 days and attained Hamburger & Hamilton (1951)stage 22.
Grafting
Whole mandibular processes, recombined components from the mouse, orcomponents combined with epithelia or ectomesenchyme from mandibularprocesses of the chick, were grafted to the chorio-allantoic membranes of 8- or9-day-old embryonic chicks following the procedure outlined by Hall (1978 c).Briefly, the mandibular process or epithelium was placed onto a square of sterile,black, Millipore filter (0-45/*m porosity, 125-150/mi thick, Millipore FilterCorp., Montreal, Quebec), mandibular ectomesenchyme was placed in directcontact with the epithelium and the intact, or recombined processes, graftedfor 7 days.
Histology
Grafts were recovered, dissected away from the chorioallantoic membranes,fixed in neutral buffered formal saline, processed histologically, serially sectionedand stained with haematoxylin, alcian blue and chlorantine fast red (modifiedfrom Lison, 1954).
RESULTSIntact mandibular processes
The chorioallantoic membrane (CAM) has previously been shown to be agraft site which supports the initiation and maintenance of chondrogenesis andosteogenesis from avian mandibular processes (Tyler & Hall, 1977; Hall, 1978£,1980). To determine whether the CAM would support cartilage and boneformation from murine tissue, and to determine whether the initiation of eithertissue was dependent upon the age of the donor embryo, whole mandibularprocesses from mouse embryos of 9-13 gestational days were grafted to theCAMs of host embryonic chicks. Osteogenesis and chondrogenesis were initi-ated in at least 75 % of the grafts, irrespective of the ages of the mouse embryos(column I, Table 1). Typically, the cartilage consisted of one or two rods ofhypertrophic cartilage, with scant amounts of extracellular matrix and anill-defined perichondrium. The bone typically consisted of one or two ossicles oftrabecular, membrane bone, usually separated from the cartilage by inter-vening connective tissue and often resting on the Millipore Filter substrate.

Tab
le 1
. Num
ber
of g
raft
s fo
rmin
g ca
rtil
age
and/
or m
embr
ane
bone
Inta
ct m
andi
bula
rpr
oces
ses
Age
r-
embr
yo C
artil
age
Bon
e(d
ays)
IIIs
olat
ed e
cto-
mes
ench
yme
Car
tilag
e B
one
III
Rec
ombi
ned
epith
eliu
m a
ndec
tom
esen
chym
eA
Car
tilag
e B
one
IVf
Mou
se e
cto-
mes
ench
yme
+ch
ick
epith
eliu
mA
I Car
tilag
e B
one
vtC
hick
ect
omes
en-
chym
e +
mou
seep
ithel
ium
Car
tilag
e B
one
N
9 10 11 12 13
3 7 10 18 7
4 6 6 16 6
4 7 10 21 8
0 13 5 14 18
1 13 5 12 16
8 17 6 15 18
4 5 3 7 10
9 7 4 7 10
6 7 12 6 10
0 5 12 4 6
6 7 12 6 11
* n,
num
ber
of g
raft
s an
alys
ed.
t T
he c
hick
tiss
ues
wer
e ob
tain
ed f
rom
em
bryo
s of
H. H
. sta
ge 2
2 (4
day
s of
incu
batio
n). W
hen
chic
k m
andi
bula
r ec
tom
esen
chym
e w
as re
com
bine
dw
ith c
hick
epi
thel
ium
and
gra
fted
, al
l gr
afts
for
med
bot
h ca
rtila
ge a
nd b
one.

256 B. K. HALL
Fig. 1. Membrane bone (B) developed when mandibular ectomesenchyme from a10-day-old embryo was grafted to the chorioallantoic membrane of a host embryonicchick. O, osteocyte; M, Millipore filter substrate. Alcian blue, chlorantine fast redand haematoxylin.
Both the large percentage of grafts forming skeleton tissues and the advancedstate of differentiation attained by the tissues, indicated that the chorioallantoicmembrane was a suitable graft site for these mouse cells. The mandibularepithelium also continued to differentiate and formed a multi-layered squamousepithelia, with numerous buds extending into the underlying ectomesenchyme.
Isolated mandibular ectomesenchyme
Mandibular ectomesenchyme which had been dissected away from themandibular epithelium after treatment of the intact mandibular process withtrypsin and pancreatin was then grafted. Provided that the embryo was at least10 days old, cartilage and membrane bone formed in a similar percentage ofgrafts to that seen when intact mandibular processes were grafted, viz. at least75 % (see column II, Table 1, Fig. 1). The lack of murine epithelial cells in thesegrafts attested to the adequacy of the tissue separations. Although the amountof cartilage and bone present was not quantified, it was not noticeably less thanthat obtained from intact mandibular processes.
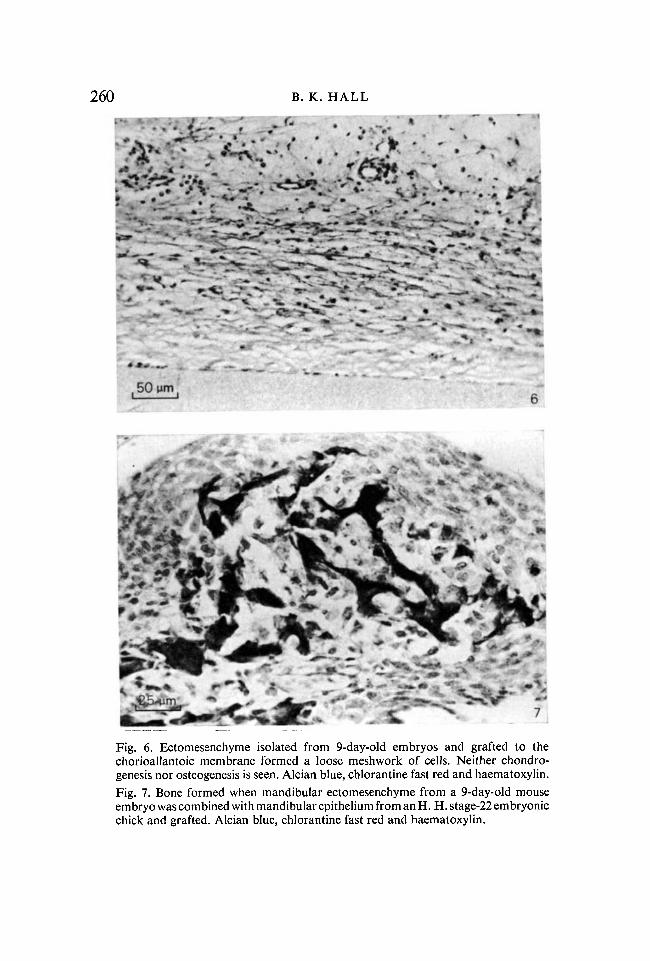
The ectomesenchyme isolated from the 9-day-old embryos behaved quitedifferently. Chondrogenesis was not initiated in any graft, and osteogenesisonly in one out of the eight grafts. The grafts consisted of a loose meshwork ofetomesenchymal cells (Fig. 6).
Thus, mandibular ectomesenchyme would only form bone and cartilage if theepithelium was present until 10 days of gestation.

Skeletogenesis in the murine mandible 257
Recombined epithelia and ectomesenchyme
To confirm that the inability of ectomesenchyme from 9-day-old embryos toform cartilage and bone was a consequence of its isolation from mandibularepithelium, these two components were recombined and grafted to the CAMs ofhost embryonic chicks. Recombinations of components from 10- to 13-day-oldembryos served as a control for the procedure. In these specimens chondrogene-sis and osteogenesis were initiated at the same, on a higher rate, than whenintact mandibular processes or isolated ectomesenchyme were grafted (columnIII, Table 1, Fig. 2).
Having confirmed that the procedure of recombination did not inhibitskeletogenesis in these older tissues, epithelia and ectomesenchyme from mandi-bular processes of 9-day-old embryos were recombined and grafted. Bothosteogenesis and chondrogenesis were initiated in such grafts (column III,Table 1), indicating that the lack of cartilage and bone in the isolated ecto-mesenchymes was attributable to the absence of the mandibular epithelia.
Heterospecific recombinations
Mandibular processes from embryonic mice and from H. H. (Hamburger &Hamilton, 1951) stage-22 embryonic chicks were separated and recombinedheterospecifica lly.
It is known that mandibular epithelium from the H. H. stage-22 embryonicchick will allow chick mandibular ectomesenchyme to initiate osteogenesis(Tyler & Hall, 1977; Hall, 1978Z?). Would chick mandibular epithelium sub-stitute for murine epithelium in initiating osteogenesis? What of the initiation ofchondrogenesis? Chondrogenesis in the chick mandible is not dependent onthe presence of the mandibular epithelium. Would the chick epithelium allowthe murine ectomesenchyme to form cartilage?
In the first recombination, mouse mandibular ectomesenchyme was recom-bined with chick mandibular epithelium and grafted. Ectomesenchymes from10- to 13-day-old mice were used as controls. Both bone and cartilage formed inthese recombinants (column IV, Table 1, Figs. 3, 4). Cartilage and bone alsoformed in four out of the nine grafts containing 9-day murine ectomesenchyme(Fig. 7). (In assessing these results all recovered grafts were counted. Given thatthe amount of epithelial-ectomesenchymal contact varied from graft to graft,the positive results reported represent a minimal estimate.) This recombinationindicated that mandibular ectomesenchyme from 9-day-old embryonic micecould respond to chick mandibular epithelium by initiating both chondro-genesis and intramembranous osteogenesis.
In the second recombination, H. H. stage-22 chick mandibular ectomesen-chyme was recombined with mouse mandibular epithelium of various ages(column V, Table 1). As noted above, the initiation of chondrogenesis in suchavian ectomesenchyme is not dependent upon the presence of mandibular

258 B. K. HALL
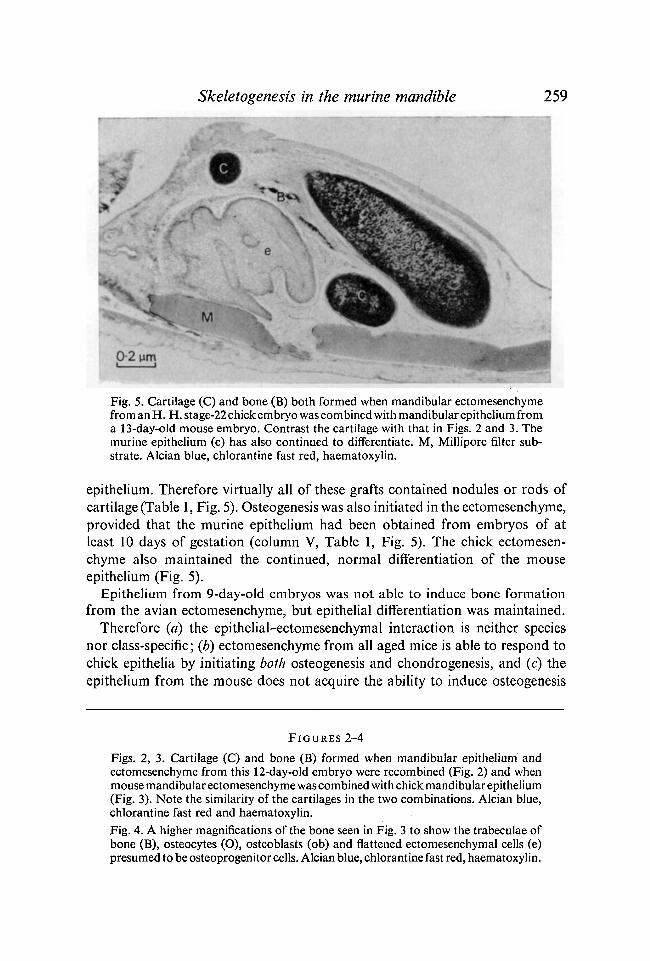
Skeletogenesis in the marine mandible 259
Fig. 5. Cartilage (C) and bone (B) both formed when mandibular ectomesenchymefrom an H. H. stage-22 chick embryo was combined with mandibular epithelium froma 13-day-old mouse embryo. Contrast the cartilage with that in Figs. 2 and 3. Themurine epithelium (e) has also continued to differentiate. M, Millipore filter sub-strate. Alcian blue, chlorantine fast red, haematoxylin.
epithelium. Therefore virtually all of these grafts contained nodules or rods ofcartilage (Table 1, Fig. 5). Osteogenesis was also initiated in the ectomesenchyme,provided that the murine epithelium had been obtained from embryos of atleast 10 days of gestation (column V, Table 1, Fig. 5). The chick ectomesen-chyme also maintained the continued, normal differentiation of the mouseepithelium (Fig. 5).
Epithelium from 9-day-old embryos was not able to induce bone formationfrom the avian ectomesenchyme, but epithelial differentiation was maintained.
Therefore (a) the epithelial-ectomesenchymal interaction is neither speciesnor class-specific; (b) ectomesenchyme from all aged mice is able to respond tochick epithelia by initiating both osteogenesis and chondrogenesis, and (c) theepithelium from the mouse does not acquire the ability to induce osteogenesis
FIGURES 2-4
Figs. 2, 3. Cartilage (C) and bone (B) formed when mandibular epithelium andectomesenchyme from this 12-day-old embryo were recombined (Fig. 2) and whenmouse mandibular ectomesenchyme was combined with chick mandibular epithelium(Fig. 3). Note the similarity of the cartilages in the two combinations. Alcian blue,chlorantine fast red and haematoxylin.Fig. 4. A higher magnifications of the bone seen in Fig. 3 to show the trabeculae ofbone (B), osteocytes (O), osteoblasts (ob) and flattened ectomesenchymal cells (e)presumed to be osteoprogenitor cells. Alcian blue, chlorantine fast red, haematoxylin.
260 B. K. HALL
Fig. 6. Ectomessnchyme isolated from 9-day-old embryos and grafted to thechorioallantoic membrane formed a loose meshwork of cells. Neither chondro-genesis nor osteogenesis is seen. Alcian blue, chlorantine fast red and haematoxylin.Fig. 7. Bone formed when mandibular ectomesenchyme from a 9-day-old mouseembryo was combined with mandibular epithelium from an H. H. stage-22 embryonicchick and grafted. Alcian blue, chlorantine fast red and haematoxylin.

Skeletogenesis in the marine mandible 261or chondrogenesis until 10 days of gestation. The lack of bone or cartilagedevelopment from ectomesenchyme isolated from 9-day-old embryos reflectedthe absence of an epithelial signal rather than the inability of the ectomesenchymeto respond to such a signal.
DISCUSSION
Although a considerable amount is known about chondrogenesis, surprisinglylittle information is available on the microenvironmental control of the initi-ation of intramembranous osteogenesis. It is known (a) that calvarial osteo-genesis is only initiated after pre-osteogenetic cells have undergone an inductiveinteraction with the brain or spinal cord (Schowing, 1968); (b) that scleral bonesonly form in the avian eye after ectomesenchymal cells have interacted withepithelial scleral papillae (Coulombre, Coulombre & Mehta, 1962); (c) that thesub-periosteal bone around the shafts of developing long bones only forms afterperichondrial cells have been in contact, either with hypertrophic chondrocytesor with their extracellular products (Shimomura, Yoneda & Suzuki, 1975); and(d) that membrane bones of the avian mandible and maxilla only form afterectomesenchymal cells have interacted with the appropriate epithelia (seeIntroduction). The dentary of the mouse may now be added to this list.
The present study demonstrates that cells of the mandibular process mustinteract with the mandibular epithelium if they are to initiate the sequence ofdifferentiative events which culminate in the differentiation of osteoblasts andosteocytes, the deposition of osteoid and the mineralization of that osteoid toform bone. That this developmental interaction had not occurred by 9 daysof gestation was shown by the fact that ectomesenchyme isolated from suchaged embryos failed to form bone (except in one case (column II, Table 1), whereepithelial cells were found to have been left on the ectomesenchyme). The re-initiation of osteogenesis in recombinants confirmed that presence of the epi-thelium was a sufficient osteogenetic stimulus for these ectomesenchymal cells.Therefore, the murine dentary resembles the membrane bones of the avianmandible (Tyler & Hall, 1977) in requiring a mandibular epithelial-ectomesen-chymal interaction for its initiation.
Unlike Meckel's cartilage in avian embryos, initiation of the murine cartilagewas dependent upon the presence of the mandibular epithelium. In the chick theectomesenchymal cells are already able to form cartilage before they reach themandibular arch (Tyler & Hall, 1977; Hall & Tremaine, 1979), apparentlybecause of prior interactions with cranial epithelial ectoderm (Bee & Thorogood,1980). In the urodele amphibians the equivalent interaction is between cellsderived from the neural crest and the pharyngeal endoderm (see Introduction).Thus, these three classes of vertebrates (Amphibia, Aves, Mammalia) shareepithelial-ectomesenchymal interactions as a common component for theinitiation of chondrogenesis in the mandibular skeleton, differing only in thenature of the signalling epithelium. Whether these interactions are mediated via

262 B. K. HALL
epithelial cell products, as is the initiation of scleral chondrogenesis (Newsome,1972, 1976), or via direct cell-to-cell contact, remains to be determined. Theosteogenetic interaction in the chick requires (a) that the epithelium be vital,and proliferating (Hall, 1980), and (b) the presence of epithelially-derivedcollagen and glycosaminoglycans (Bradamante & Hall, 1980).
It was further shown that these interactions, like those which are responsiblefor the formation of epidermal structures such as hairs, feathers and teeth(Kollar, 1972; Dhouailly, 1975), were neither species nor class specific. Mouseectomesenchyme of any age responded to chick epithelium by forming bothbone and cartilage. Chick ectomesenchyme responded to mouse epitheliumolder than 9 days by forming bone and cartilage. From these results I concludedthat mouse mandibular ectomesenchyme was capable of responding to epi-thelial influences at 9 days of gestation but that the murine epithelium was notactive until 10 days of gestation. A qualitatively similar temporal pattern hasbeen shown for inductively active epithelia from the chick (Hall, 1978 b). Ineach combination the bone and cartilage formed were typical of the speciesproviding the ectomesenchyme (cf. Fig. 4 and 5), indicating that, while theepithelium was required to initiate osteo- or chondrogenesis, it played no rolein determining the type of cartilage or bone which formed.
The identification of these tissue interactions provide a new basis for the studyof malformations affecting the mammalian facial skeleton. Failure of thesetissues to interact, interaction with fewer than normal ectomesenchymal cells,or delayed interactions, could all alter craniofacial growth and/or morpho-genesis. Mutant mice, or embryos from females given teratogens, could be usedto provide epithelia or ectomesenchyme for recombination with componentsfrom wild-type or untreated embryos to further explore the role of such tissueinteractions in development.
This research has been supported by the National Sciences and Engineering ResearchCouncil of Canada (Grant no. A5O56). The Dalhousie University Research DevelopmentFund in the Sciences provided funds to purchase the CO2 Incubator. Pilot grafts of wholemandibular process and ectomesenchyme from the older embryos were performed by P.Elliott. Sharon Brunt provided expert technical assistance.
REFERENCES
BARTELMEZ, G. W. (1952). The proliferation of the neural crest from forebrain levels in therat. Contrib. Embryol. Carnegie Inst. 37, 3-12.
BEE, J. & THOROGOOD, P. (1980). The role of tissue interactions in the skeletogenic differentia-tion of avian neural crest cells. Devi Biol. (in the press).
BERKOVITZ, B. K. B. & MOORE, M. H. (1974). A longitudinal study of replacement patterns ofteeth on the lower jaw and tongue in the rainbow trout, Salmo gairdneri. Archs oral Biol. 19,1111-1120.
BRADAMANTE, Z. & HALL, B. K. (1980). The role of epithelial collagen and proteoglycan inthe initiation of osteogenesis by avian neural crest cells. Anat. Rec. (in the press).
CASSIN, C. & CAPURON, A. (1977). Evolution de la capacite morphogenetique de la regionstomodeale chez Pembryon de Pleurodeles waltlii Michah (Amphibien Urodele). Etude par

Skeletogenesis in the murine mandible 263transplantation intrablastocelienne et par culture in vitro. W. Roux's Arch, devl Biol. 181,107-112.
CORSIN, J. (1975). Differentiation in vitro de cartilage a partir des cretes neurales cephaliqueschez Pleurodeles waltlii Michah. J. Embryol. exp. Morph. 33, 335-342.
COULOMBRE, A. J., COULOMBRE, J. L. & MEHTA, H. (1962). The skeleton of the eye. I. Con-junctival papillae and scleral ossicles. Devi Biol. 5, 382-401.
CUSIMANO-CAROLLO, T. (1963). Investigation on the ability of the neural folds to induce amouth in the Discoglossus pictus embryos. Acta Embryol. Morph. Exp. 6, 158-169.
CUSIMANO-CAROLLO, T. (1972). On the mechanism of the formation of the larval mouth inDiscoglossus pictus. Acta Embryol. Morph. exp. 4, 289-332.
DAMAS, H. (1944). Recherches sur le developpement de Lampetra fluviatilis L. Contribution al'etude de la cephalogenese des vertebres. Archs Biol. 55, 1-128.
DAMJANOV, I., SOLTER, D. & SERMAN, D. (1973). Teratocarcinoma with the capacity fordifferentiation restricted to neuro-ectodermal tissue. Virchow's Arch. Cell Pathol. 13,179-195.
DE BEER, G. R. (1947). The differentiation of neural crest cells into visceral cartilages andodontoblasts in Amblystoma, and a re-examination of the germ-layer theory. Proc. R. Soc.(Lond.) B 134, 377-398.
DHOUAILLY, D. (1975). Formation of cutaneous appendages in dermal-epidermal recom-binations between reptiles, birds and mammals. W. Roux's Arch, devl Biol. 177, 323-340.
DREWS, U., KOCHER-BECKER, U. & DREWS, U. (1972). The induction of visceral cartilagefrom cranial neural crest by pharyngeal endoderm in hanging drop cultures and thelocomotory behaviour of the neural crest cells during cartilage differentiation. W. RouxArch. EntwMech. org. 171, 17-37.
EPPERLEIN, H. H. (1974). The ectomesenchymal-endodermal interaction-system (EEIS) ofTriturus alpestris in tissue culture. 1. Observations on attachment, migration, and dif-ferentiation of neural crest cells. Differentiation 2, 151-168.
GRUNEBERG, H. (1943). The development of some external features in mouse embryos./ . Hered. 34, 89-92.
HALL, B. K. (1978a). Developmental and Cellular Skeletal Biology. New York and London:Academic Press.
HALL, B. K. (19786). Initiation of osteogenesis by mandibular mesenchyme of the embryonicchick in response to mandibular and non-mandibular epithelia. Archs oral Biol. 23, 1157-1161.
HALL, B. K. (1978 C). Grafting organs and tissues to the chorioallantoic membrane of theembryonic chick. Tissue Culture Assoc. Manual 4, 881-884.
HALL, B. K. (1980). Viability and proliferation of epithelia and the initiation of osteogenesiswithin mandibular ectomesenchyme in the embryonic chick. / . Embryol. exp. Morph.56, 71-89.
HALL, B. K. & TREMAINE, R. (1979). Ability of neural crest cells from the embryonic chick todifferentiate into cartilage before their migration away from the neural tube. Anat. Rec.194, 469-476.
HAMBURGER, V. & HAMILTON, H. L. (1951). A series of normal stages in development of thechick embryo. / . Morph. 88, 49-92.
HORSTADIUS, S. (1950). The Neural Crest; Its Properties and Derivatives in the Light of Experi-mental Research. London: Oxford University Press.
JOLLIE, M. (1971). Some developmental aspects of the head skeleton of the 35-37 mmSqualus acanthias foetus. / . Morph. 133, 17-40.
KOLLAR, E. J. (1972). The development of the integument: spatial, temporal and phylogeneticfactors. Amer. Zool. 12, 125-135.
LEVAK-SVAJGER, B. & SVAJGER, A. (1971). Differentiation of endodermal tissues in homo-grafts of primitive ectoderm from two-layered rat embryonic shields. Experientia 27,683-684.
LISON, L. (1954). Alcian blue 8G with chlorantine fast red 5B: a technique for selectivestaining of mucopolysaccharides. Stain Technol. 29, 131-138.

264 B. K. HALL
MAYER, T. C. (1973). The migratory pathway of neural cells into the skin of mouse embryos.Devi Biol. 34, 39-46.
MILAIRE, J. (1959). Predifferenciation cytochemique des divers ebauches cephaliques chezl'embryon des souris. Archs Biol. {Liege) 70, 587-730.
MILAIRE, J. (1974). Histochemical aspects of organogenesis in Vertebrates. In Handbuch d.Histochemie, vol. 8, suppl. 3. pp. 1-135. Stuttgart: Fischer-Verlag.
MORRISS, G. A. & THOROGOOD, P. V. (1978). An approach to cranial neural crest cell migra-tion and differentiation in mammalian embryos. In Development of Mammals (ed. M. H.Johnson), vol. 3, pp. 363-412. Amsterdam: North Holland.
MULNARD, J. (1955). Contribution a la connaissance des enzymes dans l'ontogenese. ArchsBiol. {Liege) 66, 525-685.
MURRAY, P. D. F. (1943). The development of the conjunctival papillae and of the scleralbones in the chick embryo. / . Anat. 11, 225-240.
NEWSOME, D. A. (1972). Cartilage induction by retinal pigmented epithelium of chick em-bryos. Devi. Biol. 27, 575-579.
NEWSOME, D. A. (1976). In vitro stimulation of cartilage in embryonic chick neural crestcells by products of retinal pigmented epithelium. Devi Biol. 49, 496-507.
NEWTH, D. R. (1951). Experiments on the neural crest of the Lamprey embryo. / . exp. Biol.28, 247-260.
NEWTH, D. R. (1956). On the neural crest of the Lamprey embryo. / . Embryol. exp. Morph. 4,358-375.
O'RAHILLY, R. (1965). The optic, vestibulocochlear and terminal vomeronasal neural crest instaged human embryos. In The Structure of the Eye (ed. J. W. Rohen), pp. 557-564.Stuttgart: F. K. Schattauer-Verlag.
POSWILLO, D. (1974). Otomandibular deformity: pathogenesis as a guide to reconstruction.J. Maxillofac. Surg. 2, 64-72.
REINBOLD, R. (1968). Role dutapetum dans la difFerenciation de la sclerotique chez l'embryonde poulet. / . Embryol. exp. Morph. 19, 43-47.
SCHAEFFER, B. (1977). The dermal skeleton in fishes. In Problems in Vertebrate Evolution (ed.S. M. Andrews, R. S. Miles and A. D. Walker), Linn, Soc. Symp. 4, pp. 25-52.
SCHOWING, J. (1968). Influence inductrice de l'encephale embryonnaire sur la developpementdu crane chez le poulet. III. Mise en evidence du role inducteur de l'encephale dansPosteogenese du crane embryonnaire du poulet. / . Embryol. exp. Morph. 19, 83-94.
SHIMOMURA, Y., YONEDA, T. & SUZUKI, F. (1975). Osteogenesis by chondrocytes fromgrowth cartilage of rat rib. Calc. Tissue Res. 19, 179-188.
SVAJGER, A. & LEVAK-SVAJGER, B. (1976). Differentiation of the first branchial arch mesen-chyme of the rat embryo in renal homografts and in organ culture. Bull. Scient. Cons.Acads RSF Yougosl. (A) 21, 139-140.
TYLER, M. S. (1978). Epithelia influences on membrane bone formation in the maxilla of theembryonic chick. Anat. Rec. 192, 225-234.
TYLER, M. S. & HALL, B. K. (1977). Epithelial influences on skeletogenesis in the mandible ofthe embryonic chick. Anat. Rec. 188, 229-240.
TYLER, M. S. & MCCOBB, D. P. (1980). The genesis of membrane bone in the embryonicchick maxilla: epithelial-mesenchymal tissue recombination studies. J. Embryol. exp.Morph. 56, 269-281.
WAGNER, G. (1949). Die Bedeutung der Neuralleiste fiir die Kopfgestaltung der Amphibien-larven, Untersuchungen an Chimaeren von Triton und Bombinator. Rev. suisse Zool. 56,519-620.
WILDE, C. E. JR. (1955). The urodele neuroepithelium. The differentiation in vitro of thecranial neural crest. J. exp. Zool. 130, 573 -591.
{Received 9 January 1980, revised 14 March 1980)