tierärztliche hochschule hannover charakterisierung eines ... · ihr genom ist eine lineare,...
TRANSCRIPT

Tierärztliche Hochschule Hannover
_______________________________________________________
Charakterisierung eines murinen Norovirus –
Auswirkungen des Virus auf die experimentelle chronisch
entzündliche Darmerkrankung bei der Interleukin-10
defizienten Maus
INAUGURAL-DISSERTATION
zur Erlangung des Grades eines
Doktors der Veterinärmedizin
-Doctor medicinae veterinariae-
(Dr. med. vet.)
vorgelegt von
Marcel Achard
Münster
Hannover 2011

Wissenschaftliche Betreuung: Univ.-Prof. Dr. Hans-J. Hedrich
Institut für Versuchstierkunde und
Zentrales Tierlaboratorium
Medizinische Hochschule Hannover
Univ.-Prof. A. Bleich, Ph.D.
Institut für Versuchstierkunde und
Zentrales Tierlaboratorium
Medizinische Hochschule Hannover
Apl.-Prof. Dr. M. Mähler
Labor für biomedizinische Diagnostik - BioDoc
Hannover
1. Gutachter: Univ.-Prof. Dr. Hans-J. Hedrich
2. Gutachter: Apl.-Prof. Dr. Ludwig Haas
Tag der mündlichen Prüfung: 24. November 2011

Meinen Eltern


Inhaltsverzeichnis
I
Inhaltsverzeichnis
Abkürzungsverzeichnis ........................................................................................... V
1 Einleitung ........................................................................................................... 1
2 Literaturübersicht .............................................................................................. 2
2.1 Caliciviridae ................................................................................................... 2
2.2 Genetik und Molekularbiologie von MNV ....................................................... 3
2.3 MNV in der Zellkultur ..................................................................................... 5
2.4 Nachweisverfahren für MNV .......................................................................... 7
2.5 MNV-Prävalenz in Versuchstierhaltungen ..................................................... 8
2.6 Erfassung und Bekämpfung von MNV in der Labortierhaltung ...................... 8
2.7 Infektionsverlauf ............................................................................................. 9
2.8 Untersuchungen zum Einfluss von MNV auf bestimmte Mausstämme und
Tiermodelle .................................................................................................... 9
2.9 Weitere Forschung mit murinen Noroviren .................................................. 12
2.10 Tiermodelle für chronisch entzündliche Darmerkrankungen (CED) ............. 12
2.11 Die Interleukin-10 defiziente Maus ............................................................... 15
2.12 Ziele dieser Arbeit ........................................................................................ 17
3 Eigene Untersuchungen ..................................................................................18
3.1 Material ........................................................................................................ 18
3.1.1 Mäuse ................................................................................................ 18
3.1.2 Herkunft ............................................................................................. 18
3.1.3 Haltung im Experiment ...................................................................... 19
3.1.4 Synthetische Oligonukleotide ............................................................ 22
3.1.5 Zelllinie .............................................................................................. 24
3.1.6 Eingesetzter Virusstamm ................................................................... 24

Inhaltsverzeichnis
II
3.1.7 Eingesetzter Bakterienstamm ............................................................ 24
3.1.8 Materialliste ....................................................................................... 24
3.2 Methoden ..................................................................................................... 31
3.2.1 Zellkultur ............................................................................................ 31
3.2.2 Virusisolation ..................................................................................... 31
3.2.3 Virus-Nachweis mittels PCR .............................................................. 32
3.2.4 Nachweis von Helicobacter hepaticus mittels PCR ........................... 35
3.2.5 Sequenzierung .................................................................................. 36
3.2.6 Virus-Anzucht für den Infektionsversuch ........................................... 37
3.2.7 Virus-Titer-Bestimmung ..................................................................... 37
3.2.8 Immunofluorescence Assay (IFA) ...................................................... 39
3.2.9 Virusaufreinigung mittels Ultrazentrifugation ..................................... 40
3.2.10 Enzyme-linked Immunosorbent Assay (ELISA) ................................. 42
3.2.11 Elektronenmikroskopische Aufnahmen.............................................. 44
3.2.12 Isolation und Anzucht von Helicobacter hepaticus für den
Doppelinfektionsversuch ................................................................... 44
3.2.13 Experimentelle Infektion der Mäuse mit MNV .................................... 45
3.2.14 Experimentelle Infektion der Mäuse mit Helicobacter hepaticus ........ 45
3.2.15 Sterile Aufarbeitung des Mesenteriallymphknotens unter einer
Werkbank .......................................................................................... 48
3.2.16 Vorbereitung und Verwendung der Zellkulturplatten für die
Bestimmung von Interferon γ............................................................. 49
3.2.17 Bestimmung von Interferon γ im Lymphozytenzellkulturüberstand mit
Hilfe eines ELISA .............................................................................. 49
3.2.18 Anfertigung histologischer Präparate ................................................. 50
3.2.19 Histologische Auswertung der Präparate ........................................... 51

Inhaltsverzeichnis
III
3.2.20 Statistische Auswertung .................................................................... 52
4 Ergebnisse ........................................................................................................53
4.1 Anzucht von MNV in der Zellkultur ............................................................... 53
4.2 Virus-Nachweis mittels PCR ........................................................................ 53
4.3 Sequenzierung des Virus ............................................................................. 56
4.4 Virusquantifizierung ..................................................................................... 58
4.5 Indirekte Antikörpernachweisverfahren ........................................................ 58
4.5.1 IFA ..................................................................................................... 58
4.5.2 Antigenaufreinigung und ELISA ......................................................... 60
4.6 Elektronenmikroskopische Aufnahmen ........................................................ 66
4.7 Serologische Untersuchung von Anzeigertieren des zentralen
Tierlaboratoriums auf MNV .......................................................................... 68
4.8 Infekionsversuche ........................................................................................ 68
4.8.1 Klinik .................................................................................................. 70
4.8.2 Sektion ............................................................................................... 70
4.8.3 Serologie ........................................................................................... 70
4.8.4 Lymphozytenstimulation .................................................................... 71
4.8.5 PCR-Nachweis von MNV ................................................................... 78
4.8.6 PCR-Nachweis von Helicobacter hepaticus ...................................... 79
4.8.7 Histologie ........................................................................................... 79
5 Diskussion ........................................................................................................84
5.1 Zellkultur und Quantifizierung ...................................................................... 84
5.2 Sequenzierung ............................................................................................. 84
5.3 Nachweisverfahren ...................................................................................... 85
5.4 Prävalenz ..................................................................................................... 89
5.5 MNV im Tierversuch .................................................................................... 90

Inhaltsverzeichnis
IV
5.6 Ausblick ....................................................................................................... 91
6 Zusammenfassung ...........................................................................................93
7 Summary ...........................................................................................................95
8 Anhang ..............................................................................................................97
9 Literaturverzeichnis .......................................................................................115
Abbildungsverzeichnis .........................................................................................126
Tabellenverzeichnis ..............................................................................................129
Erklärung ................................................................................................................131
Danksagung ...........................................................................................................132

Abkürzungsverzeichnis
V
Abkürzungsverzeichnis
% Prozent
BHI Brain-Heart-Infusion
BLAST Basic Local Alignment Search Tool
bzw. beziehungsweise
CD Cluster of differentiation
cDNA Complementary DNA
CED Chronisch entzündliche Darmerkrankung
CPE Cytopathischer Effekt
DMEM Dulbecco's Modified Eagle's Medium
DNA Desoxyribonucleic Acid
EDTA Ethylendiamintetraacetat
EL Charles-River-ELISA-Messung
ELISA Enzyme linked immune sorbent assay
FELASA Federation of Laboratory Animal Science Associations
FKS L.E. Fetales Kälberserum low endotoxin
GM-CSF Granulocyte macrophage colony-stimulating factor
h Stunde
H. hepaticus Helicobacter hepaticus
H.E. Hämatoxylin-Eosin
HPRT Hypoxanthin-Guanin-Phosphoribosyltransferase
IFA Immunofluorescence Assay
IFNγ Interferon gamma
IL Interleukin
kb Kilobasenpaare
l Liter
m männlich
MCMV Maus Cytomegalie Virus
MDA Melanoma-differentiation-associated gene
MDR Multiple drug resistance
MEZ Mitteleuropäische Zeit
MFI Multiplex fluorescent immunoassay
MHH Medizinische Hochschule Hannover
min Minute
ml Milliliter
MLK Mesenteriallymphknoten
MNV Murines Norovirus
MNV-h Murines Norovirus/ Hannover1/2007/DEU
MW Mittelwert
NCBI National Center for Biotechnology Information

Abkürzungsverzeichnis
VI
NEA Nicht essentielle Aminosäuren
ng Nanogramm
NSR Nicht spezifische Reaktion
OD Optical density
ORF Open reading frame
p.i. post infectionem
PBS Phosphate buffered saline
PBSM Phosphate buffered saline ohne Mg2+- und Ca2+-Ionen
PCR Polymerase Chain Reaction
pg Picogramm
RAG Recombination-activating gene
RNA Ribonucleic Acid
RT Raumtemperatur
scid Severe combined immunodeficiency
SD Standardabweichung
SEM Standard Error of the Mean
STAT Signal transducer and activator of transcription
TBE TRIS-Borat-EDTA-Puffer
TCID Tissue culture infectious dose
TEM Transmissionselektronenmikroskopie
Th T-Helfer
TJL The Jackson Laboratory
TLR Toll-like receptor
TNF Tumornekrosefaktor
TRIS Trishydroxymethylaminomethan
U Umdrehung
v.a. vor allem
VK Variationskoeffizient
vs. versus
w weiblich
ZTL Zentrales Tierlaboratorium

Einleitung
1
1 Einleitung
Noroviren sind unbehüllte, sehr umweltresistente RNA-Viren und gehören zur Familie
der Caliciviridae, in der sie ein eigenständiges Genus bilden. Sie wurden erstmals
1968 bei einem Ausbruch akuter Gastroenteritis an einer Schule in Norwalk/Ohio in
den USA entdeckt (DOLIN et al. 1972) und werden für ca. 90% aller nicht-bakteriell
bedingten epidemischen Gastroenteritiden beim Menschen verantwortlich gemacht.
Zu den animalen Noroviren gehören bovine, porzine und murine Noroviren. Bisher
wurde keine Übertragung dieser Viren vom Tier auf den Menschen oder umgekehrt
nachgewiesen.
Der erste Nachweis eines Norovirus bei der Maus wurde 2003 beschrieben (KARST
et al. 2003). Infektionsexperimente mit diesem Virus, dem murinen Norovirus 1
(MNV-1), zeigen, dass Infektionsdauer und Krankheitsmanifestation vom
Mausstamm beeinflusst werden (KARST et al. 2003; HSU et al. 2005; MUMPHREY
et al. 2007). Seit der Erstbeschreibung von MNV-1 wurden viele weitere MNV-
Stämme mit unterschiedlichen biologischen Eigenschaften isoliert (HSU et al. 2006;
THACKRAY et al. 2007). Untersuchungen in Nordamerika (HSU et al. 2005;
PRITCHETT-CORNING et al. 2009) und Westeuropa (MÜLLER et al. 2007;
MÄHLER u. KÖHL 2009) dokumentieren eine hohe Prävalenz von MNV-Infektionen
bei Labormäusen. In der bis dato größten Studie (PRITCHETT-CORNING et al.
2009) wiesen 32,4% von 44.876 getesteten Serumproben MNV-spezifische
Antikörper auf.
Die Bedeutung von murinen Noroviren als mögliche Einflussgröße für
Tierexperimente ist gegenwärtig unklar und Untersuchungsziel verschiedener
Forschergruppen. Ziel dieser Arbeit war es, ein murines Norovirusisolat zu
charakterisieren, direkte und indirekte Nachweisverfahren für MNV zur Erfassung der
Prävalenz in einer Labortierhaltung zu entwickeln, sowie den Einfluss des murinen
Norovirus auf die Interleukin-10 (IL10) defiziente Maus - ein Tiermodell für chronisch
entzündliche Darmerkrankungen (CED) (KÜHN et al. 1993) - zu untersuchen.

Literaturübersicht
2
2 Literaturübersicht
2.1 Caliciviridae
Die Familie der Caliciviridae besteht aus kleinen (27 bis 40 nm) unbehüllten
ikosaedrischen Viren. Ihr Genom ist eine lineare, positiv polare, einsträngige RNA.
Innerhalb der Familie Caliciviridae gibt es vier Genus: Norovirus, Sapovirus,
Vesivirus und Lagovirus. Die am häufigsten auftretenden humanmedizinschen
Krankheitserreger dieser Familie gehören zu den Noro- und Sapoviren, die eine
akute Gastroenteritis hervorrufen. Diese sind auch bekannt als „Norwalk-like“ Viren;
erstmals wurde ein Krankheitsausbruch 1968 an einer Schule in Norwalk/Ohio in den
USA beschrieben und weitergehend untersucht (DOLIN et al. 1972).
Wichtige veterinärmedizinische Erreger sind das feline Calicivirus (FCV, Vesivirus),
das Atemwegserkrankungen bei der Katze hervorruft, und das Virus der
hämorrhagischen Erkrankung beim Kaninchen, abgekürzt RHDV (rabbit hemorrhagic
disease virus), das zu den Lagoviren gehört (FIELDS et al. 2007).
Noroviren bei Labornagern
Der erste Nachweis eines Norovirus bei der Maus wurde 2003 beschrieben (KARST
et al. 2003). Weitere murine Noroviren mit der Bezeichnung MNV-2, MNV-3 und
MNV-4 wurden 2006 isoliert (HSU et al. 2006) und charakterisiert (HSU et al. 2007).
Im gleichen Jahr wurden auch in Berlin drei neu Stämme isoliert und charakterisiert
(MÜLLER et al. 2007). Thackray et al. vergleichen 2007 die Genome 21 neuer MNV-
Isolate mit 5 bekannten und nehmen eine phylogenetische Einteilung in 15 Stämme
vor (THACKRAY et al. 2007). Auch aktuell werden weitere Isolate publiziert (KIM et
al. 2010). Bisher konnten keine Noroviren bei Ratten oder anderen Nagern
nachgewiesen werden (HENDERSON 2008).

Literaturübersicht
3
2.2 Genetik und Molekularbiologie von MNV
Das murine Norovirus besitzt ein lineares, positiv polares, einsträngiges RNA-
Genom, das ungefähr 7,5 kb lang ist und drei offene Leseraster beinhaltet (ORF1,
ORF2 und ORF3; Abbildung 1) (FIELDS et al. 2007). ORF1 kodiert ein Polyprotein,
das durch eine 3C-ähnliche Protease endolytisch in mindestens sechs nicht-
strukturelle Proteine gespalten wird (BLAKENEY et al. 2003). ORF2 kodiert das
Haupt-Kapsid-Protein (Virales Protein 1). ORF 3 kodiert das virale Protein 2, das
eine Rolle bei der Stabilität vom Kapsid-Protein spielt (HSU et al. 2006). Ein viertes
offenes Leseraster, das sich mit ORF2 überlappt, wird von Thackray beschrieben
(THACKRAY et al. 2007). Die verschiedenen murinen Noroviren weisen eine
genetische Diversität von bis zu 13% auf (THACKRAY et al. 2007). Konservierte
Bereiche werden in der „junction region“ zwischen ORF1 und ORF2 beschrieben
(MÜLLER 2009).
Abbildung 1: Schema des MNV-Genoms; unterhalb der Leseraster sind die
verschiedenen Proteinprodukte mit ihrem Gewicht in Kilodalton aufgeführt; N, N
Terminus; 3A, NTPase 3A-like; P, Vpg Protease; RdRp, RNA dependent RNA
Polymerase; VP1, VP2 Hauptkapsidproteine (HENDERSON 2008).

Literaturübersicht
4
Das murine Norovirus ist unbehüllt, hat eine Größe von 28 - 35 nm und weist die
typische äußere Kelchstruktur der Caliciviridae auf (siehe Abbildung 2 und Abbildung
3) (KARST et al. 2003).
Abbildung 2: Elektronenmikroskopische Aufnahme von MNV-1 (markiert durch
schwarze Pfeile) (HSU et al. 2005).

Literaturübersicht
5
Abbildung 3: Dreidimensionale Rekonstruktion anhand cryoelektronen-
mikroskopischer Aufnahmen von humanem Norovirus (NV, virus-like particles),
murinem Norovirus (MNV) und dem Virus der hämorrhagischen Erkrankung beim
Kaninchen (RHDV, virus-like particles) im Vergleich (KATPALLY et al. 2010).
2.3 MNV in der Zellkultur
Als erstes Virus des Genus Norovirus ist eine Anzucht von MNV in der Zellkultur
möglich. Das murine Norovirus zeigt dabei einen Tropismus zu Makrophagen und
dendritischen Zellen (WOBUS et al. 2004). Die Anzucht erfolgt in der
Mausmakrophagen-Zelllinie RAW 264.7 (RASCHKE et al. 1978; WOBUS et al.
2004). Das Virus verursacht einen deutlichen cytopathischen Effekt (CPE). Eine

Literaturübersicht
6
Virusquantifizierung erfolgt daher mittels Plaque-Test (WOBUS et al. 2004). Nicht
alle isolierten MNV-Stämme weisen jedoch auszählbare Resultate im Plaque-Test
auf. In diesen Fällen wird ein „tissue culture infectious dose“ (TCID)50-Test
angewandt (MÜLLER et al. 2007; THACKRAY et al. 2007).
Durch Passagieren in der Zellkultur kann es zu einer Attenuierung von MNV kommen
(WOBUS et al. 2004). In einem Fall ließ sich diese Attenuierung auf eine
Aminosäurenveränderung auf Position 296 des Viruskapsid-Proteins zurückführen.
Wird die Veränderung rückgängig gemacht, erreicht der Erreger seine alte Virulenz
im Infektionsversuch mit signal transducer and activator of transcription (STAT) 1-
defizienten Mäusen (KARST et al. 2003; BAILEY et al. 2008). Die erfolgreiche
Anzucht ermöglicht biochemische und molekulare Untersuchungen zum Zelleintritt
und zur Replikation des murinen Norovirus. So untersuchten Perry et al. den
Mechanismus des Zelleintritts von MNV1-Virionen, der im Gegensatz zu felinen
Caliciviren keinen niedrigen pH-Wert im Endosom erfordert (PERRY et al. 2009;
PERRY u. WOBUS 2010). Ein niedriger pH-Wert löst bei felinen Caliciviren die
Freisetzung des Virusgenoms in der Zelle aus (STUART u. BROWN 2006).
Gerondopoulos et al. untersuchten weitere Abhängigkeiten der Virusendozytose bei
MNV-1 (GERONDOPOULOS et al. 2010). Elektronenmikroskopische Aufnahmen der
Virusreplikation zeigen starke Veränderungen der Zellmorphologie. Unter Verlust
eines intakten Golgi-Apparates werden intrazelluläre Membranen extensiv
umgewandelt (WOBUS et al. 2004). Hyde et al. untersuchten die Virus-Replikation
und spätere Proteinlokalisation in der Wirtszelle bei MNV-1 (HYDE et al. 2009; HYDE
u. MACKENZIE 2010). Han et al. analysierten die in vitro RNA-Synthese-Aktivität der
„RNA dependent RNA Polymerase“ (HAN et al. 2010). Die virusbedingte
Zellapoptose mit MNV infizierter Makrophagen erfolgt durch eine Aktivierung von
Kaspase-9 und Kaspase-3. Außerdem wird die Expression von Survivin
herunterreguliert (BOK et al. 2009).

Literaturübersicht
7
2.4 Nachweisverfahren für MNV
MNV kann in der Zellkultur vermehrt und anschließend mittels Ultrazentrifugation per
Cäsiumchlorid (CsCl)-Gradient aufgereinigt werden. Es ergibt sich eine Bande auf
der Höhe anderer Noroviren (Dichte 1,36 ± 0,04 g/cm³) (KARST et al. 2003). Aus der
entnommenen Bande angefertigte elektronenmikroskopische Präparate zeigen
Partikel mit der für Caliciviren typischen Struktur (KARST et al. 2003).
Das MNV-Genom kann mittels PCR nachgewiesen werden. Hierzu muss die Virus-
RNA zuerst zu cDNA umgeschrieben werden. Dann erfolgt im zweiten Schritt eine
Vervielfältigung der cDNA. Da Noroviren eine hohe Mutationsrate haben, ist ein
Nachweis aller murinen Noroviren mit einem Primerpaar bisher nicht beschrieben.
Bisher werden mehrere Primerpaare für einen diagnostischen Nachweis
herangezogen (HENDERSON 2008). Ein Nachweis ist beschrieben aus Milz, Leber,
Mesenteriallymphknoten, Duodenum, Jejunum, Zäkum und Kotproben (HSU et al.
2005; MANUEL et al. 2008; GOTO et al. 2009). Dabei gelten Kot sowie Organe des
Magen-Darm-Traktes, vor allem Zäkum und Mesenteriallymphknoten, als
zuverlässige Nachweisorte für MNV (GOTO et al. 2009). Auch das „Poolen“ von
mehreren MNV-negativen und einer MNV-positiven Kotprobe sowie das
anschließende Lagern bei Raumtemperatur für 14 Tage führten zu einem positiven
PCR-Ergebnis (MANUEL et al. 2008). Der PCR-Nachweis wird erschwert durch die
variable Ausscheidungsdauer von MNV je nach Virusstamm und infiziertem
Mausstamm (HSU et al. 2006).
Von der Maus gegen MNV gebildete Antikörper können mit serologischen Tests
nachgewiesen werden. Hsu et al. stellten eine Kreuzreaktivität bei Antikörpern gegen
MNV-1, MNV-2, MNV-3 und MNV-4 fest (HSU et al. 2006). Die Antikörper richten
sich dabei gegen das Viruskapsid (CHACHU et al. 2008). Die Kreuzreaktivität
ermöglicht die Anwendung serologischer Testverfahren wie multiplex fluorescent
immunoassay (MFI), enzyme-linked immunosorbent assay (ELISA) und
immunofluorescence assay (IFA) (HSU et al. 2005). Murine Noroviren sind

Literaturübersicht
8
Bestandteil einer einzigen Genogruppe und weisen einen Serotyp auf (THACKRAY
et al. 2007).
2.5 MNV-Prävalenz in Versuchstierhaltungen
Die Prävalenz von MNV in Labortierhaltungen in den USA und Kanada wurde 2005
anhand von 12639 Serumproben untersucht. Hierbei ergab sich eine Prävalenzrate
von 22,1% (HSU et al. 2005). Eine von Januar 2007 bis Juni 2008 durchgeführte
Erhebung bei Mäuseseren aus westeuropäischen Labortierhaltungen ergab für MNV
eine Prävalenzrate von 31,8% (MÄHLER u. KÖHL 2009). Die bisher größte Studie
ergab eine Prävalenzrate von 32,4% bei 44876 auf MNV getesteten Mäuseseren, die
aus Versuchstierhaltungen in Nordamerika und Europa stammten (PRITCHETT-
CORNING et al. 2009). Eine koreanische Studie zeigte eine 36,5 prozentige
serologische Prävalenz für MNV bei gentechnisch veränderten Mäusen (YEOM et al.
2009). Goto et al. untersuchten Maus-Zäkum-Proben von verschiedenen
Labortierhaltungen per PCR auf ein Vorhandensein von MNV und ermittelten dabei
eine Prävalenzrate von 13,1% (GOTO et al. 2009).
2.6 Erfassung und Bekämpfung von MNV in der Labortierhaltung
Der Einsatz von Anzeiger-Mäusen zur MNV-Erfassung im Experiment wird in zwei
Studien erläutert. Bei Manuel et al. waren 80% der Anzeiger-Mäuse positiv für MNV
im PCR-Nachweis, nachdem sie Kontakt zur Einstreu von mit MNV infizierten
Mäusen hatten (MANUEL et al. 2008). Eine gegensätzliche Beobachtung schildern
Compton et al. Hier kam es keineswegs immer zu einer MNV-Infektion nach Haltung
von Anzeiger-Mäusen auf der Einstreu von MNV-infizierten Mäusen (COMPTON et
al. 2010).
Die Sanierung eines Haltungsbereichs, in dem Mäuse mit MNV infiziert waren,
gelang nur durch Keulen aller Tiere und anschließende Raumdesinfektion vor der
Neubesiedlung. Eine Testung und Entfernung positiver Tiere allein war nicht
erfolgreich (KASTENMAYER et al. 2008; GOTO et al. 2009). Eine Infektion
neugeborener Mäuse von MNV-positiven Muttertieren konnte durch Transfer der

Literaturübersicht
9
Säuglinge auf MNV-freie Ammen („cross fostering“) in den ersten 24 Stunden nach
der Geburt verhindert werden (ARTWOHL et al. 2008; COMPTON 2008). Eine
andere Studie hatte weniger Erfolg bei der Sanierung von gentechnisch
manipulierten Mäusen durch „cross fostering“ (YEOM et al. 2009). Embryotransfer
und Hysterektomie wurden ebenfalls als erfolgreiche Eradikationsmethoden
beschrieben (PERDUE et al. 2007; GOTO et al. 2009).
2.7 Infektionsverlauf
Die Infektion mit MNV unterscheidet sich in Dauer und klinischer Symptomatik je
nach infiziertem Mausstamm als auch dem für die Infektion verantwortlichen
Norovirusstamm (WOBUS et al. 2004; MUMPHREY et al. 2007; THACKRAY et al.
2007; KELMENSON et al. 2009). Je nach Virusstamm und infiziertem Mäusestamm
kann es zu einer persistenten Infektion oder verlängerten Ausscheidung über den
Kot kommen (HSU et al. 2006). Bei immunkompetenten 129S6 Mäusen wurde eine
Infektionsdauer mit MNV-1 von 7-14 Tagen festgestellt. Bei immunkompetenten
Hsd:ICR Mäusen dauerte die Infektion 5 Wochen und länger. Die Infektion ging nicht
mit klinischen Symptomen einher (KARST et al. 2003; HSU et al. 2005). Bei
bestimmten immundefizienten Mausstämmen [Interferon (INF)-αβγ Rezeptor-
Defizienz und STAT1-Defizienz] konnte die Infektion jedoch zu tödlichen
systemischen Erkrankungen wie Enzephalitis, Vaskulitis, Meningitis, Hepatitis und
Pneumonie führen. Bei anderen immundefizienten Mausstämmen [Recombination-
activating gene (RAG)1- und RAG2-Defzienz] persistierte die Infektion länger als 90
Tage ohne jegliche Krankheitssymptome (KARST et al. 2003).
2.8 Untersuchungen zum Einfluss von MNV auf bestimmte Mausstämme und
Tiermodelle
- Ward et al. erforschten die Pathologie natürlich vorkommender MNV-Infektionen
bei 28 immundefizienten Mäusen verschiedener Genotypen in zwei
verschiedenen Maushaltungen. In der einen Haltung stellten sie bei Tieren mit
verschiedenen kombinierten genetischen Defekten Hepatitis, Peritonitis oder
Pneumonie anhand histologischer Veränderungen fest. Diese kombinierten

Literaturübersicht
10
genetischen Defekte betrafen das RAG1, den IFNγ-Rezeptor, OT-1, OT-2 und
STAT1. RAG2-defiziente Mäuse aus der zweiten Maushaltung waren histologisch
unauffällig. Bei ihnen wurde MNV jedoch per PCR im Mesenteriallymphknoten
nachgewiesen (WARD et al. 2006).
- Wobus et al. stellten eine STAT1-abhängige Interferon-Immunantwort im
Zellkultur- und Infektionsversuch fest (WOBUS et al. 2004). Mumphrey et al.
infizierten im Folgeversuch Wildtyp- und STAT1-defiziente Mäuse. Verwandt
wurden zwei verschiedene MNV-Isolate, eines davon war attenuiert.
Neugeborene Wildtyp-Mäuse zeigten weder Diarrhoe noch Gewichtsverlust. Bei
den STAT1-defizienten Mäusen kam es zu einem Gewichtsverlust. Außerdem
wurden milde histologische Veränderungen in Darm und Milz erfasst. Das Virus
wurde in Milz, Leber, Lunge und Lymphknoten nachgewiesen; in der Milz konnte
MNV dauerhaft bis Versuchsende nachgewiesen werden, was für eine
persistente Infektion spricht (MUMPHREY et al. 2007).
- Chachu et al. beobachteten einen längeren Verlauf einer MNV-Infektion bei B-
Zell-defizienten Mäusen. Außerdem versahen sie RAG1-defiziente Mäuse mit
Splenozyten von B-Zell defizienten Mäusen. Dies führte nicht zu einer
Virusclearance. Die Gabe von polyklonalem Anti-MNV-Serum oder monoklonalen
Antikörpern verringerte den systemischen und luminalen Virusgehalt (CHACHU et
al. 2008).
- Lencioni et al. untersuchten den Einfluss von MNV-4 auf die multiple drug
resistance 1α (MDR1α) defiziente Maus, ein Tiermodell für CED. Die Infektion mit
MNV-4 und anschließender Verabreichung von Helicobacter (H.) bilis 7 Tage
danach führte hierbei zu größerem Gewichtsverlust und höhergradigen
histologischen Veränderungen im Vergleich zur Kontrollgruppe, die nur mit H. bilis
infiziert wurde. Eine alleinige MNV-4-Infektion führte zu erhöhten Serum-
Biomarkern. Mittels Durchflusszytometrie wurden außerdem Veränderungen bei
dendritischen Zellen (CD11c+) und anderen nicht T-Zell (CD4- CD8-)

Literaturübersicht
11
Populationen ermittelt. Dendritische Zellen, gewonnen aus MNV-4-infizierten
Mäusen, sorgten für eine höhere IFNγ-Sekretion bei polyklonalen T-Zellen im in
vitro-Versuch an Tag 2 p.i. Zu späteren Messzeitpunkten wurde keine höhere
IFNγ-Sekretion gemessen. Die akute Infektion mit MNV-4 förderte die Antigen-
Präsentation dendritischer Zellen und wirkte sich immunmodulatorisch auf den
Krankheitsverlauf aus. Es wurden keine klinische Symptome bei Mäusen
beobachtet, die nur mit MNV-4 infiziert waren (LENCIONI et al. 2008).
- McCartney et al. untersuchten den Einfluss von MNV auf melanoma-
differentiation-associated gene (MDA)5- und toll-like receptor (TLR)3-defiziente
Mäuse sowie dendritische Zellen mit dem jeweiligen Defekt. Sowohl bei MDA5-
defizienten Mäusen als auch bei MDA5-defizienten dendritischen Zellen wurde
ein höherer Virustiter im Vergleich zum Wildtyp durch einen Plaquetest ermittelt.
TLR3-defiziente dendritische Zellen hatten keinen höheren Virustiter im in vitro-
Versuch, TLR3-defiziente Mäuse wiesen jedoch einen leicht erhöhten Virustiter
auf. MDA5-defiziente Mäuse eliminierten die MNV-Infektion vollständig
(MCCARTNEY et al. 2008).
- Doom et al. zeigten, dass MNV kaum Einfluss auf die experimentelle Infektion mit
dem Cytomegalievirus der Maus (MCMV) hat. Lediglich die CD8 T-Zell-Antwort
wurde in der akuten Phase der Infektion geringgradig herabgesetzt. Eine MNV-
Infektion führte nicht zu einem Wiederaufblühen einer latenten MCMV-Infektion
(DOOM et al. 2009).
- Hensley et al. stellten keinen Einfluss von MNV auf die adaptive Immunität gegen
das Vaccinia oder Influenza A Virus bei C57BL/6 Wildtyp Mäusen fest (HENSLEY
et al. 2009).
- Compton et al. erforschten den Einfluss von MNV auf die Ausscheidung von
Maus Parvovirus (MPV). Eine MNV-Infektion verlängerte die Ausscheidungsdauer
von MPV über den Kot und erhöhte den MPV-DNA-Spiegel in Organen von

Literaturübersicht
12
BALB/c-Mäusen. Die Serokonversion für MPV erfolgte bei einer Doppelinfektion
von BALB/c-Mäusen innerhalb kürzerer Zeit. Außerdem konnte bei Anzeiger-
Mäusen, die Kontakt mit der Einstreu MNV-infizierter Tiere hatten, nicht in jedem
Fall eine Serokonversion für MNV festgestellt werden (COMPTON et al. 2010).
- Paik et al. stellten keinen Einfluss von MNV auf ernährungsbedingte Fettleibigkeit
und Insulinresistenz bei C57BL/6 Wildtyp Mäusen fest. Bei diesen Mäusen war
vor der Infektion durch eine extrem fettreiche Diät eine Insulinresistenz
herbeigeführt worden (PAIK et al. 2010).
2.9 Weitere Forschung mit murinen Noroviren
Da das murine Norovirus in der Zellkultur anzüchtbar ist, wird es als Ersatz für das
humane Norovirus im Rahmen der Effektivitätsuntersuchung von Desinfektions-
techniken und -mitteln verwendet (BAERT et al. 2008; PARK et al. 2010). Auch für
die Erfassung sowie Reduktion des Eintrags von Noroviren in die Lebensmittelkette
über Muscheln, Fisch, Salat und Gemüse wird MNV als Surrogatmarker eingesetzt
(WEI et al. 2010, 2011).
2.10 Tiermodelle für chronisch entzündliche Darmerkrankungen (CED)
CED sind charakterisiert durch rezidivierende oder kontinuierliche Entzündungen des
Intestinaltraktes (BAUMGART u. CARDING 2007; BAUMGART u. SANDBORN
2007). Zu den häufigsten CED beim Menschen gehören Morbus Crohn und Colitis
ulcerosa (FARROKHYAR et al. 2001). Morbus Crohn ist durch eine transmurale
Entzündung verschiedener Segmente des Gastrointestinaltraktes mit
diskontinuierlicher Ausbreitung und episodischer Progression gekennzeichnet.
Hierbei können Strikturen, Abszesse und Fisteln als Komplikation auftreten
(BAUMGART u. SANDBORN 2007). Colitis ulcerosa weist eine Entzündung der
Mukosa und oberflächlichen Submukosa des Kolons auf, die schubweise
voranschreitet und sich kontinuierlich nach oral ausbreitet. Typische Symptome für
beide Erkrankungen sind blutige bis wässrige Durchfälle, Bauchschmerzen, Übelkeit,
Erbrechen und Fieber (BAUMGART u. SANDBORN 2007). Die Äthiopathogenese

Literaturübersicht
13
der CED ist bisher noch nicht vollständig aufgeklärt. Es gibt deutliche Hinweise, dass
durch eine Kombination von immunologischen und umweltbedingten Faktoren eine
unkontrollierte Immunantwort innerhalb des Darms ausgelöst wird, die in einem
genetisch prädisponierten Individuum zu einer chronischen Entzündung führt
(TORRES u. RIOS 2008).
Tiermodelle, zum Beispiel für CED, ermöglichen es, störende Umwelteinflüsse durch
standardisierte Haltungsbedingungen zu kontrollieren. Durch den Einsatz von
Inzuchtstämmen können Studien an genetisch homogenen Populationen
durchgeführt werden. Techniken der Transgenese und der gerichteten Mutagenese
stehen zur Verfügung, um das Genom der Tiere gentechnisch zu verändern. Seit
Anfang der 90er Jahre werden zahlreiche Tiermodelle, v.a. Mausmodelle, für CED
beschrieben, die klinische und histopathologische Analogien zum Morbus Crohn oder
zur Colitis ulcerosa aufweisen.
Einige Tiermodelle für CED
- spontan:
SAMP1/Yit-Maus (MATSUMOTO et al. 1998)
- durch Chemikalien induziert:
Dextran Natriumsulfat (OKAYASU et al. 1990)
Trinitrobenzolsulfonsäure (ELSON et al. 1996)
Oxazolon (BOIRIVANT et al. 1998)
- durch Zelltransfer:
CD4+/CD45RBhigh-Zellen in Prkdcscid-Mäuse (POWRIE et al. 1993)
Knochenmarkszellen in CD3ε transgene Mäuse (HOLLANDER et al. 1995)
- durch genetische Manipulation:
IL2-defiziente Maus (SADLACK et al. 1993)
IL10-defiziente Maus (KÜHN et al. 1993)
MDR1α-defiziente Maus (PANWALA et al. 1998)

Literaturübersicht
14
SMAD3-defiziente Maus (YANG et al. 1999)
Verschiedene Tiermodelle für CED werden durch Umweltfaktoren beeinflusst.
Besonders die Zusammensetzung der intestinalen Mikroflora sowie spezifische
Keime haben starken Einfluss (siehe Tabelle 1). Der genetische Hintergrundstamm
der Merkmalsträger kann sich ebenfalls auf den Verlauf der CED auswirken (siehe
Tabelle 2).
Tabelle 1: Interaktion verschiedener Umweltfaktoren auf das CED-Modell der IL10-
defizienten Maus
Umweltfaktor Auswirkung Referenz
Haltung unter keimfreien Bedingungen
Keine Kolitis (SELLON et al. 1998)
Haltung unter keimarmen Bedingungen bzw. nach Antibiotikagabe
Reduktion der Kolitissymptome
(KÜHN et al. 1993; MADSEN et al. 2000)
Keim Lactobacillus spp.
Protektiv (MADSEN et al. 1999; SCHULTZ u. SARTOR 2000)
Keim Enterococcus faecalis
Proinflammatorisch (BALISH u. WARNER 2002)
Keim Helicobacter spp.
Proinflammatorisch (CAHILL et al. 1997; KULLBERG et al. 1998; BURICH et al. 2001)
Tabelle 2: Ausprägung der Kolitis bei IL10-defizienten Inzuchtstämmen
Hintergrundstamm Schwere und Verlauf der Kolitis
Referenz
C57BL/6J Mild, protrahiert (BERG et al. 1996; TAKEDA et al. 1999; BRISTOL et al. 2000)
BALB/c Intermediär, progressiv (BERG et al. 1996)
NOD/Lt Intermediär, progressiv (MÄHLER u. LEITER 2002)
NOD.NON-H2nb1 Intermediär, progressiv (MÄHLER u. LEITER 2002)
129/SvEv Schwer, progressiv (TAKEDA et al. 1999)
C3H/HeJBir Schwer, beginnt sehr früh (BRISTOL et al. 2000)
C3H.SW Schwer, beginnt sehr früh (MÄHLER u. LEITER 2002)

Literaturübersicht
15
2.11 Die Interleukin-10 defiziente Maus
Die IL10-defiziente Maus (formal: Il10tm1Cgn; abgekürzt Il10-/-) wurde 1993 am Institut
für Genetik der Universität Köln durch gezielte Mutation entwickelt (KÜHN et al.
1993). Ein Teil des ersten Exons des IL10-Gens (Codon 5-55) wurde mit Hilfe
homologer Rekombination durch ein Stopcodon und ein neo-Gen (Neomycin
exprimierend) ersetzt. Zusätzlich wurde ein Stopcodon in Exon 3 des Gens
eingefügt. Mäuse mit dieser Nullmutation entwickeln nach dem Absetzen eine
spontane Form der CED. Der Entzündungsprozess kann sich über den gesamten
Intestinaltrakt erstrecken. Er tritt dabei segmental, teilweise transmural auf.
Histologisch ist diese Enterokolitis durch Entzündungszellinfiltrate in der Lamina
propria und der Submukosa, Erosionen und Ulzerationen der Mukosa,
Schleimhauthyperplasie, eine abnorme Kryptarchitektur, Becherzelldepletionen
sowie Kryptabszesse gekennzeichnet (KÜHN et al. 1993; LÖHLER et al. 1995;
BERG et al. 1996). Im Spätstadium der Erkrankung können kolorektale Tumoren
auftreten (BERG et al. 1996). Da bei IL10-defizienten Mäusen außer dem
Intestinaltrakt keine weiteren Organsysteme entzündlich verändert sind, eignet sich
dieses Tiermodell besonders für Studien bezüglich der CED. Daher wurde es auch
an der Medizinischen Hochschule Hannover (MHH) bereits häufig untersucht und für
CED-Studien eingesetzt (MOST 2001; BLEICH 2003; BÜCHLER 2010).
IL10 ist ein regulatorisches Zytokin, welches von zahlreichen Immunzellen, wie
Makrophagen, dendritischen Zellen und T-Zellen produziert wird. Innerhalb der T-
Zellen sezernieren fast alle Untergruppen IL10, inklusive Th1-, Th2- und
regulatorischen T-Zellen (MURRAY 2006). Neben Immunzellen sind auch
Epithelzellen und Keratinozyten in der Lage, IL10 freizusetzen. IL10 unterdrückt die
Effektorfunktion von Th1/Th17-Zellen, Natürlichen Killerzellen und Makrophagen. Es
moduliert die zelluläre Immunreaktion (MOORE et al. 1993). Diese modulierenden
Effekte bleiben bei IL10-defizienten Mäusen aus. Ihr Immunsystem reagiert
gegenüber luminalen Antigenen bzw. der intestinalen Bakterienflora mit einer
überschießenden Immunantwort (siehe Abbildung 4) (BERG et al. 1996). Eine
Analyse von konditionalen Knockout-Mäusen, bei denen IL10 nur in T-Zellen

Literaturübersicht
16
inaktiviert wurde, zeigte eine vergleichbare Kolitis wie bei IL10-defizienten Mäusen
(ROERS et al. 2004). Das von T-Zellen produzierte IL10 ist also essentiell für die
Verhinderung einer spontanen CED in diesem Tiermodell. Vermutlich nehmen die
IL23-Th17-Achse und die von Th17-Zellen produzierten Zytokine wie IL17, IL21,
IL22, Tumornekrosefaktor (TNF), granulocyte macrophage colony-stimulating factor
(GM-CSF) und IL6 eine wichtige Funktion bei der Entzündungsreaktion ein (YEN et
al. 2006).
Abbildung 4: Pathogenese der CED bei IL10-defizienten Mäusen, modifiziert nach
Powrie (POWRIE 1995) und Elson (ELSON 1999).

Literaturübersicht
17
2.12 Ziele dieser Arbeit
Ziel dieser Arbeit war es, das bereits isolierte murine Norovirus zu charakterisieren.
Das Virus sollte mit gebräuchlichen Methoden quantifiziert werden. Zusätzlich sollte
eine serologische Diagnostik in Form eines ELISA aufgebaut werden. Mit Hilfe
serologischer Testverfahren sollte die Prävalenz für MNV in einer Labortierhaltung -
dem Zentralen Tierlaboratorium (ZTL) der Medizinischen Hochschule Hannover
(MHH) – ermittelt werden.
Nach der Charakterisierung sollten dann Infektionsversuche durchgeführt werden,
die den Einfluss von MNV auf die IL10-defiziente Maus - ein Tiermodell für CED
(KÜHN et al. 1993) - erfassen.

Eigene Untersuchungen
18
3 Eigene Untersuchungen
3.1 Material
Die Durchführung der Versuche wurde mit den Bescheiden vom 13.03.2008 und
10.04.2008 durch das Niedersächsische Landesamt für Verbraucherschutz und
Lebensmittelsicherheit mit den Aktenzeichen 08/1443 und 08/1467 genehmigt.
3.1.1 Mäuse
In der Zeit vom 2.4.2008 bis 6.5.2009 wurden die in Tabelle 3 aufgeführten
Mäuseinzucht- und Defektmutantenstämme für den Infektionsversuch mit dem
murinen Norovirus Stamm Hannover eingesetzt. Im Weiteren werden für eine
übersichtlichere Gestaltung nur noch die entsprechenden Abkürzungen für die
Mausstämme benutzt.
Tabelle 3: Verwendete Mausstämme und ihre Abkürzung
Stammbezeichnung Knockout/ Wildtyp
Abkürzung
C3H/HeJBirZtm Wildtyp C3-wt
C57BL/6JZtm Wildtyp B6-wt
C3Bir.129P2-Il10tm1Cgn/JZtm Knockout C3-Il10-/-
B6.129P2-Il10tm1Cgn/JZtm Knockout B6-Il10-/-
3.1.2 Herkunft
Alle im Experiment eingesetzten Mäuse stammten aus Zuchten des ZTL der MHH.
Sie wurden dort regelmäßigen, mikrobiologischen Kontrolluntersuchungen
entsprechend den Empfehlungen der Federation of European Laboratory Animal
Science Associations (FELASA) unterzogen (NICKLAS et al. 2002). Die
Versuchstiere entstammten aus Haltungsbereichen, die frei von bakteriellen, viralen
(inklusive MNV) und parasitären Infektionserregern gemäß der Empfehlungsliste der
FELASA waren.

Eigene Untersuchungen
19
3.1.3 Haltung im Experiment
Die experimentell infizierten Mäuse wurden im S2-Bereich des ZTL der MHH in
einem belüfteten Käfigregalschrank (Scantainer) gehalten. Es wurden jeweils 1-5
Mäuse in Makrolon-Käfigen des Typs II oder III mit Filterdeckeln
getrenntgeschlechtlich gehalten. Die Kontrolltiere wurden bis auf 5 Tiere in der
Haltungsabteilung belassen, um eine mögliche Kontamination mit MNV zu
vermeiden. Die 5 Kontrolltiere, die gesondert mit im Käfigregalschrank saßen,
wurden daher zuerst umgesetzt und versorgt. Futter (Tabelle 4) und autoklaviertes
Trinkwasser (über Tränkeflaschen) wurden ad libitum angeboten. Die Einstreu wurde
ein- bis zweimal in der Woche erneuert. Die Reinigung der Käfige erfolgte
routinemäßig. Die Raumtemperatur betrug 22 ± 2° C, die relative Luftfeuchte lag bei
55 ± 5%. Der Hell-Dunkel-Zyklus wurde automatisch geregelt; es herrschte ein
Lichtrhythmus von 14 Stunden Helligkeit zu 10 Stunden Dunkelheit, wobei die
Hellphase von 7 bis 21 Uhr mitteleuropäischer Zeit (MEZ) dauerte.
Tabelle 4: Zusammensetzung von Altromin 1324 TPF
Inhaltsstoffe Prozent %
Rohprotein 19,0
Rohfett 4,0
Rohfaser 6,0
Rohasche 7,0
Kalzium 0,9
Phosphor 0,7
Zusatzstoffe je Kilogramm
Vitamin A 15000 I.E.
Vitamin D3 600 I.E.
Vitamin E 75 mg
Vitamin C 36 mg
Kupfer 5 mg

Eigene Untersuchungen
20
Verwendete Lösungen
Alle verwendeten Chemikalien wurden, wenn nicht anders angegeben, über die
Firma Carl Roth in Karlsruhe bezogen.
Phosphate buffered saline (PBS) (pH = 7,2)
Menge
NaCl (Natriumchlorid) 8,00 g
KCl (Kaliumchlorid) 0,20 g
Na2HPO4 (di-Natriumhydrogenphosphat) 1,15 g
KH2PO4 (Kaliumdihydrogenphosphat) 0,20 g
Aqua bidest. 1,0 l
CaCl2 (Calciumchlorid) 0,10 g
MgCl2 x 6 H2O (Magnesiumchlorid – Hexahydrat)
0,10 g
PBS Tween (pH = 7,4)
Menge
NaCl (Natriumchlorid) 8,00 g
KCl (Kaliumchlorid) 0,20 g
Na2HPO4 (di-Natriumhydrogenphosphat) 2,90 g
KH2PO4 (Kaliumdihydrogenphosphat) 0,20 g
Aqua bidest. 1,0 l
Tween 20 0,5 ml
PBSM (pH = 7,2)
Menge
NaCl (Natriumchlorid) 8,00 g
KCl (Kaliumchlorid) 0,20 g
Na2HPO4 (di-Natriumhydrogenphosphat) 1,15 g
KH2PO4 (Kaliumdihydrogenphosphat) 0,20 g
Aqua bidest. 800 ml
Evan´s Blue
Menge
Evans blue 5 mg
Glycerin 200 ml
Aqua bidest. 800 ml

Eigene Untersuchungen
21
Coating Buffer (ph = 9,6)
Menge
Na2CO3 (Natriumcarbonat, wasserfrei) 1,59 g
NaHCO3 (Natriumhydrogencarbonat) 2,93 g
Aqua bidest. 1,0 l
Konjugate
Hersteller/ Artikelnummer
Goat Anti-Mouse IgG (whole molecule)-FITC Sigma / Art. F 0257
Goat Anti-Mouse IgG (whole molecule)-Peroxidase Sigma / Art. A 4416
10x TBE-Puffer
Menge
TRIS 108 g
Borsäure 53,4 g
EDTA 7,4 g
Aqua bidest. ad 1 l
1x TBE-Puffer
Menge
10x TBE-Puffer 100 ml
Aqua bidest. 900 ml
Orange G-Ladepuffer
Menge
Orange G (Sigma) 0,05 g
Glycerin 99% 30 ml
Aqua bidest. 70 ml
RAW-Medium
Pro Flasche Menge
Dulbecco´s Modified Eagle´s medium (DMEM) 22 ml
Fetales Kälberserum (FKS) Ultra Low Endotoxin (L.E.) 2,5 ml
Penicillin-Streptomycin 0,5 ml

Eigene Untersuchungen
22
Anti-mCD3
Menge
PBS 10 ml
Anti-mCD3 (145-2C11) 1 µg/µl 50 µl
Lymphozyten-Medium
Menge
RPMI 1640 500 ml
FKS Ultra Low Endotoxin 25 ml
NEA (nicht essentielle Aminosäuren) 5 ml
L-Glutamin 5 ml
2-Mercaptoethanol 0,5 ml
Pyruvat 5 ml
Penicillin-Streptomycin 5 ml
Neutrales Formalin nach Sörensen (10%ig)
Puffer A Menge
Kaliumhydrogenphosphat 13,6 g
Aqua bidest. 1 l
Puffer B
Kaliumhydrogenphosphat 17,7 g
Aqua bidest. 1 l
Puffer A 352,8 ml
Puffer B 547,2 ml
35 – 37% Formalin 100 ml
3.1.4 Synthetische Oligonukleotide
Für die Umschreibung der MNV-RNA in cDNA wurden Oligo(dT)18 und Random
Hexamer Primer der Firma Fermentas vergleichend eingesetzt.
Die zur Detektion und Sequenzierung verwendeten Primer wurden von der Firma
Sigma Aldrich bezogen.
Tabelle 5: Primer zur Detektion von MNV
Sequenz von 5´ nach 3´
5205f AATTGACCCCTGGATCTTCC
5388r GACCAGCTGAACCTCCATGT
5469r AGTGGTGAGTGACCCTTTGG

Eigene Untersuchungen
23
Fortsetzung Tabelle 5
5473f CAGATCACATGCTTCCCAC
5659r AGACCACAAAAGACTCATCAC
5580f GGAGGAATCTATGCGCCTGG
5671r GAAGGCGGCCAGAGACCAC
4972f CACGCCACCGATCTGTTCTG
5580r GCGCTGCGCCATCACTC
5004f TTTGGAACAATGGATGCTGA
5219r ATCCAGGGGTCAATTTGGTT
5277r GGTGTTTCGAGGTGAAATGG
Tabelle 6: Primer zur Klonierung von MNV
Sequenz von 5´ nach 3´
Noro Hannover for 1 GTGAAATGAGGATGGCAACG
Noro Hannover rev 1 GTGATGACATCTGTGGCCAT
Noro Hannover for 2 ATGGCCACAGATGTCATCAC
Noro Hannover rev 2 GGAAGCATGTGATCTGAGCA
Noro Hannover for 3 TGCTCAGATCACATGCTTCC
Noro Hannover rev 3 GCCAACGACCGGGAGGCCAGCTTTTTTTTTTTTTTTV
Tabelle 7: Primer zur Sequenzierung von MNV
Sequenz von 5´ nach 3´
Norosense 1 CCGATCAAGAACCTACTGGCA
Noroantisense 1 AATCATCCCAGAGAACCACC
Norosense 2 ACTTCACCAATGCCGTTA
Noroantisense 2 CAAACTGGTCACCGACTATC
Norosense 3 TTCCTTCCAAGATGAGCTCC
Noroantisense 3 CCCTGAGTAAGACACAGCAA
Norosense 1 CCGATCAAGAACCTACTGGCA
Noroantisense 1 AATCATCCCAGAGAACCACC
Tabelle 8: Primer zur Kontrolle der RNA-Umschreibung zu cDNA
Sequenz von 5´ nach 3´
RPS9f TGACGTTGGCGGATGAGCACA
RPS9r TTTTTGACAGGGGGAGTGG
Hierbei wurde das Housekeeping Gen RPS-9 nachgewiesen.

Eigene Untersuchungen
24
Tabelle 9: Primer zur Detektion von Helicobacter (H.) hepaticus
Sequenz von 5´ nach 3´
H276f CTATGACGGGTATCCGGC
H676r ATTCCACCTACCTCTCCCA
B38 GCATTTGAAACTGTTACTCTG
B39 CTGTTTTCAAGCTCCCCGAAG
3.1.5 Zelllinie
Zur Anzucht von MNV wurden RAW 264.7 Zellen verwendet (ATCC TIB-71™). Es
handelt sich hierbei um eine Mausmakrophagenzelllinie, die vom Deutschen
Krebsforschungszentrum (DKFZ) Heidelberg zur Verfügung gestellt wurde. Die
Zelllinie wurde von der Firma QM Diagnostics (Nimwegen, Niederlande) per PCR
negativ auf Mycoplasmen getestet.
3.1.6 Eingesetzter Virusstamm
Der im Experiment eingesetzte Virusstamm wurde aus NOD-Prkdcscid-Mäusen des
ZTL der MHH isoliert und nach genetischer Charakterisierung als Murines
Norovirus/Hannover1/2007/DEU (MNV-h) bezeichnet.
3.1.7 Eingesetzter Bakterienstamm
Der im Experiment eingesetzte Bakterienstamm H. hepaticus wurde im ZTL der MHH
von Mitarbeitern des mikrobiologischen Labors im Rahmen der wissenschaftlichen
Arbeit von Frau Büchler isoliert und angezüchtet (BÜCHLER 2010).
3.1.8 Materialliste
Vorversuche
Zellkultur
Zelllinie RAW 264.7, DKFZ Heidelberg, Deutschland
Brutschrank, Heraeus, Thermo Fisher Scientific, Schwerte, Deutschland
Sicherheitswerkbank MSC 1.8, Thermo Fisher Scientific, Schwerte, Deutschland
Zellkultur-Flaschen 75 cm² Vented Cap, Straight Neck, Sarstedt, Nümbrecht,
Deutschland

Eigene Untersuchungen
25
Zellschaber Cell Scraper, Greiner Bio One, Frickenhausen, Deutschland
Medium DMEM High Glucose 4,5 g/l, L-Glutamin, 500 ml, PAA, Parching, Österreich
Antibiotikum Penicillin (5000IU/ml) Streptomycin 5000 µg/ml, MP Biomedicals,
Eschwege, Deutschland
Fetales Kälberserum Ultra Low Endotoxin, PAA, Parching, Österreich
Zentrifuge Minifuge 2, Heraeus, Thermo Fisher Scientific, Schwerte, Deutschland
Glas-Gewindeflaschen mit flachem Boden und Deckel, Omnilab, Bremen,
Deutschland
Gefrierschrank Premium No-Frost, Liebherr, Biberach an der Riss, Deutschland
-80° C Truhe 6385, GFL, Großburgwedel, Deutschland
Identifizierung, Sequenzierung
Nucleospin RNA II Kit, Macherey Nagel, Düren Deutschland
Omniscript RT-Kit, Quiagen, Hilden, Deutschland
Primer Fermentas, Sigma-Aldrich, Steinheim, Deutschland
PTC Thermocycler-200TM, MJ Research, Watertown, MA, USA
Phusion™ Hot Start High-Fidelity DNA Polymerase, Finnzymes, Maahantuonti,
Finnland
Stratagene StrataClone PCR Cloning Kit, Agilent, La Jolla, CA, USA
QIAprep Spin Miniprep Kit (50), Qiagen, Hilden, Deutschland
Sequenzierung: MWG Biotech AG, Ebersberg, Deutschland
Software zur Analyse der Sequenz:
DNA Dynamo, Blue Tractor Software, North Wales, UK
BLAST http://blast.ncbi.nlm.nih.gov/Blast.cgi
Nachweisverfahren
IFA
Wasserbad 1008, GFL, Großburgwedel, Deutschland
Objektträger Cel-Line Diagnostic Microscope Slides 8 Well 8mm, Thermo Fisher
Scientific, Schwerte, Deutschland
PBS
PBSM

Eigene Untersuchungen
26
Diluter Microlab 500, Hamilton-Robotics, Martinsried, Deutschland
Antikörper Anti Mouse IGG-FITC F0257, Sigma-Aldrich, Steinheim, Deutschland
Evans Blue, Fluka, Buchs, Schweiz
Fluoreszenzmikroskop DM2000, Leica Mikrosysteme, Wetzlar, Deutschland
Lichtquelle EL6000, Leica Mikrosysteme, Wetzlar, Deutschland
ELISA
Zentrifuge Minifuge 2, Heraeus, Thermo Fisher Scientific, Schwerte, Deutschland
Ultrazentrifuge L7, Beckman-Coulter Deutschland, Krefeld, Deutschland
Ultrazentrifugenrotor SW 28, SW 32, SW 55, Beckman-Coulter Deutschland, Krefeld,
Deutschland
Bechereinsätze Centrifuge Tubes Polyallomer 32683; 326819 (SW55), Beckman-
Coulter Deutschland, Krefeld, Deutschland
Sucrose analytical grade, Serva, Heidelberg, Deutschland
Waage LA 2200S, Sartorius, Göttingen
Waage ALJ 120-4, Kern und Sohn, Balingen-Frommern, Deutschland
PBSM
Coating Buffer
Immuno-Modules U8 Maxisorp 475078, Nunc, Thermo Fisher Scientific, Schwerte,
Deutschland
Multikanal-Pipette Eppendorf, Hamburg, Deutschland
Washer Columbus Pro, Tecan, Männedorf, Schweiz
PBS Tween
Zweitantikörper Anti Mouse IGG Peroxidase, Sigma-Aldrich, Steinheim, Deutschland
ABTS-Substrat Tablets 11 112 422 001, Roche, Grenzach-Wyhlen, Deutschland
Photometer Sunrise, Tecan, Männedorf, Schweiz
Laptop Inspiron 6000, Dell Deutschland, Frankfurt, Deutschland
Software Magellan 5.03, Tecan, Männedorf, Schweiz
PCR-Nachweis
Für die RNA-Isolierung: Nucleospin RNA II Kit, Macherey Nagel, Düren, Deutschland

Eigene Untersuchungen
27
Für die DNA-Isolierung: PSP® Spin Stool DNA Kit, Invitek, Berlin, Deutschland
Photometer Nanodrop 1000, Peqlab, Erlangen, Deutschland
Omniscript RT-Kit, Quiagen, Hilden, Deutschland
REDExtract-N-AmpTM PCR ReadyMix, Sigma-Aldrich, Steinheim, Deutschland
Taq PCR Core-Kit, Quiagen, Hilden, Deutschland
Primer, Fermentas, Thermo Fisher Scientific, Schwerte, Deutschland
Primer, Sigma-Aldrich, Steinheim, Deutschland
DNA-Leiter, Biozym Art.-Nr.: 850321 (50321), Biozym Scientific, Hessisch Oldendorf
PTC Thermocycler-200TM, MJ Research, Watertown, MA, USA
Magnetrührer L81, Kisker Biotech, Steinfurt, Deutschland
Agarose LE Agarose, Biozym Scientific, Hessisch Oldendorf
DNA-Farbstoff Gelstar Nucleic Acid Gel Stain, Lonza, Basel, Schweiz
10x TBE
Orange G Ladepuffer, Carl Roth, Karlsruhe, Deutschland
Gelschlitten, Gelkammer Eigenkonstruktion der MHH, Hannover, Deutschland
Gleichstromgerät, Omnilab, Bremen, Deutschland
UV-Transilluminator, PC+Kamera, INTAS, Göttingen, Deutschland
Virusquantifizierung
Mikroskop, Zeiss, Jena, Deutschland
Neubauer-Zählkammer 0,1mm Tiefe; 0,0025mm² Fläche, Brand, Wertheim,
Deutschland
Kristallviolett-Indikator, Carl Roth, Karlsruhe, Deutschland
Plaque-Test
Platten Multi Dish 6 Well 140675, Nunc, Thermo Fisher Scientific, Schwerte,
Deutschland
Agarose SeaPlaque, Lonza, Basel, Schweiz
TCID50-Test
96-Well Platte 655185, Greiner Bio One, Frickenhausen, Deutschland

Eigene Untersuchungen
28
Elektronenmikroskopische Darstellung
Tischultrazentrifuge Airfuge® im Institut für Virologie der MHH, Beckman-Coulter
Deutschland, Krefeld, Deutschland
Elektronenmikroskop Elektronenmikroskopie-Labor MHH, Tecnai 20, FEI Europe;
Eindhoven, Niederlande
Hauptversuche
Infektionsversuche
Sicherheitswerkbank Klasse II, Thermo Fisher Scientific, Schwerte, Deutschland
Ventilierter Haltungsschrank, Scantainer, Scanbur Technology, Karlslunde,
Dänemark
Mini-Isolator „Gnotocage“ (Deckel: Eigenkonstruktion der technischen Werkstatt der
MHH; Unterbau: Multifunktionstopf Nalgene DS5300-9212), Hannover, Deutschland
bzw. Thermo Fisher Scientific, Schwerte, Deutschland
Filterdeckelkäfige Typ II, III, UNO, Roestvaststaal BV, Niederlande
Futter, pelletiertes Altromin© 1324 total pathogen free (TPF) Haltungsfutter, Altromin,
Lage, Deutschland
Autoklaviertes Trinkwasser
Einstreu, Weichholzgranulat, Fichten- und Tannenholz, Hahn & Co, Bredenbeck-
Kronsburg, Deutschland
Spritze, Spritzenkolben, Knopfkanüle/Braunüle zur Gavage, Braun, Melsungen,
Deutschland
Histocassetten, Thermo Fisher Scientific, Schwerte, Deutschland
Eppendorf-Reaktionsgefäße 1,5 ml, Eppendorf, Hamburg, Deutschland
Lymphozytenstimulation
3-Polysterene-Reaktionsgefäße-Gefäße 15 ml, Greiner Bio One, Frickenhausen,
Deutschland
Petrischalen, Greiner Bio One, Frickenhausen, Deutschland
Gewebesieb, 40µm BD Falcon M, BD Bioscience, Heidelberg, Deutschland
Zentrifuge Eppendorf, Hamburg, Deutschland

Eigene Untersuchungen
29
Lymphozytenmedium
Zellzählgerät scil Vet ABC, scil animal care company, Viernheim, Deutschland
96-Well Platte 655185, Greiner Bio One, Frickenhausen, Deutschland
Anti-mCD3 (145-2C11) 1µg/µl
ELISA zur IFNBestimmung (extern: Arbeitsgruppe Detlef Neumann, MHH)
mIFN Antikörper eingesetzt als Fänger (an ELISA-Platte gebunden) und späterer
Detektor, Pierce Antibodies, Thermo Fisher Scientific, Schwerte, Deutschland
P96 Immuno MaxiSorb Nunc, Thermo Fisher Scientific, Schwerte, Deutschland
Beschichtungspuffer: 100 mM Carbonat/Bicarbonat-Puffer, pH 9,6
Blockpuffer: PBS mit 4 % BSA
Lymphozytenmedium
Waschpuffer: PBS mit 0,005 % Tween 20
Streptavidin-HRP 0,5 mg/ml
Substrat: 3,3’,5,5’-Tetramethyl-Benzidine (TMB) 1 mg/ml in DMSO
Substratpuffer: 1,36 g Natriumacetat x 3 H2O +
2,1 g Zitronensäuremonohydrat in 100 ml H2O (pH 4,9)
3% H2O2
Isolation und Anzucht von H. hepaticus für den Doppelinfektionsversuch (ZTL)
Brutschrank, Heraeus, Thermo Fisher Scientific, Schwerte, Deutschland
Impföse, Omnilab, Bremen, Deutschland
Wattetupfer, Omnilab, Bremen, Deutschland
Polypropylen-Röhrchen, Sarstedt, Nümbrecht, Deutschland
Brain-Heart-Infusion Medium, Merck, Darmstadt, Deutschland
0,45 µm Weißrandfilter, Schleicher & Schuell, Basel, Schweiz
Petrischale Ø 60 mm, Greiner Bio One, Frickenhausen, Deutschland
Columbiaagar, Oxoid Deutschland, Wesel, Deutschland
Amphotericin B, Biochrom, Berlin, Deutschland
Campylobacter-Selektiv-Supplement (nach Skirrow), Oxoid Deutschland, Wesel,
Deutschland

Eigene Untersuchungen
30
Pferdeblut, Oxoid Deutschland, Wesel, Deutschland
Anaerobiertopf HP0011A, Oxoid Deutschland, Wesel, Deutschland
1,5 ml Schraubdeckelgefäß Twist Top Vial und Twist Top Vial Caps, Sorenson
BioScience, Salt Lake City, UT, USA
Stereomikroskop zur histologischen Auswertung
Zur statistischen Auswertung wurden folgende Programme genutzt:
StatView D-4.5, Abacus Corporation
Graphpad Prism 5, Graphpad Software Inc., La Jolla, CA, USA
Excel 2003, 2007, Microsoft, Redmond, WA, USA

Eigene Untersuchungen
31
3.2 Methoden
In den ersten Versuchen mussten grundlegende Erkenntnisse zum Virus gesammelt
werden. Das Virus war bereits isoliert worden, jedoch nicht klassifiziert. Außerdem
mussten direkte und indirekte Nachweismethoden und Virusquantifizierungs-
verfahren für den Infektionsversuch etabliert werden. Im Anschluss erfolgten
Infektionsversuche, die den Einfluss einer MNV-Infektion auf das Tiermodell der
IL10-defizienten Maus erfassen sollten.
3.2.1 Zellkultur
Die RAW 264.7 Zellen wurden in 75 ml Filterdeckel-Zellkulturflaschen mit DMEM, 2%
Penicillin-Streptomycin und 10% low endotoxine fetalem Kälberserum bei 37° C und
5% CO2-Begasung in einem Brutschrank kultiviert. Zum Passagieren wurde der Zell-
rasen mit einem Zellschaber abgelöst. Die Zellen wurden dann in 5 ml vorbereitetem
Medium und anschließend anteilig je nach gewünschter Zelldichte in insgesamt 25
ml Medium pro Zellkulturflasche resuspendiert.
3.2.2 Virusisolation
Die Virusisolation wurde von Mitarbeitern der MHH und der Firma Biodoc (Hannover,
Deutschland) durchgeführt. Aus vorhergehenden wissenschaftlichen Veröffent-
lichungen war bekannt, dass MNV weit verbreitet in versuchstierkundlichen
Einrichtungen ist. Klinische Symptome blieben jedoch abhängig vom infizierten
Mausstamm aus. Mesenteriallymphknoten und Milz waren als Virusreservoir
beschrieben. Eine Virusvermehrung gelang in RAW 264.7 Zellen. Mehrere
genetische Sequenzen von MNV-Isolaten waren bereits beschrieben. Als potentielle
Virusträger wurden NOD-Prkdcscid-Mäuse, die als Anzeigertiere im ZTL verwendet
wurden, eingeschätzt. Von ihnen wurden im Rahmen einer diagnostischen Routine-
Sektion Mesenteriallymphknoten und Milz gewonnen. Diese wurden bei -80° C
eingefroren. Eine kleine Probe der Organe wurde für eine PCR-Untersuchung mit
dem Primerpaar 5004f/5219r verwandt. Die PCR-Untersuchung ergab für Milz und
Mesenteriallymphknoten der Mäuse einen positiven Befund. Daraufhin wurde ein
Viertel der PCR-positiven Milz mit einem Cell-Douncer in DMEM homogenisiert.

Eigene Untersuchungen
32
Nach einer Zentrifugation bei 6000 U/min für 30 min (2800 x g) wurde der Überstand
filtriert. Mit dem aus dem Milz-Isolat gewonnenen Überstand wurde eine zu 75%
besiedelte RAW-Zellkulturflasche beimpft. Nach zwei Tagen trat ein cytopathischer
Effekt >90% auf, der bei einer parallel bebrüteten Kontrollflasche nicht zu
beobachten war. Die beimpfte Zellkulturflasche wurde daraufhin bei -20° C
eingefroren, aufgetaut, erneut eingefroren (-20° C) und dann aufgetaut. Das Medium
inklusive cryolysierter RAW 264.7 Zellen sowie vermeintlicher Viruspartikel wurde bei
6000 U/min für 10 min (2800 x g) zentrifugiert. Der Überstand wurde abgenommen
und filtriert, das Pellet wurde verworfen. Das Filtrat wurde in 1 ml Portionen
konfektioniert und bei -80° C tiefgefroren.
3.2.3 Virus-Nachweis mittels PCR
3.2.3.1 RNA-Isolation
Da es sich bei Noroviren um RNA-Viren handelt, musste zuerst die Virus-RNA aus
dem Filtrat gewonnen werden. Hierzu wurde das Macherey Nagel Kit NucleoSpin
RNA II mit dem Support-Protokoll zur Isolation von Gesamt-RNA aus
Zellkulturüberstand eingesetzt (siehe Anhang S.97). Bei der RNA-Isolation aus
Organen wurde ebenfalls das vom Hersteller empfohlene Protokoll angewandt (siehe
Anhang S. 97). Zur Isolation von Gesamt-RNA aus Kot wurde erfolgreich ein
modifiziertes Protokoll angewandt, das der Hersteller auf Anfrage übersandte. Es
handelte sich hierbei um die Modifikation eines Kunden, dem es auf diese Art und
Weise gelungen war, RNA aus Kot zu isolieren (siehe Anhang S. 99).
3.2.3.2 Reverse Transkription
Um die isolierte RNA weiter zu analysieren, wurde diese mit Hilfe des Quiagen
Omniscript RT-Kits umgeschrieben. Für den Reaktionsansatz wurde der RNA-Gehalt
der Proben photometrisch bestimmt. Es erfolgte eine Einstellung auf 500 ng RNA pro
10 µl Ansatz.

Eigene Untersuchungen
33
Ansatz pro Probe
RT-Buffer 2 µl
dNTPs 5 µM 2 µl
Oligo(dT)18 20 µM bzw. Random Hexamer 1 µl
RNAse-Inhibitor 1:4 verdünnt 1 µl
Reverse Transkriptase 1 µl
Eingestellte RNA 13 µl
Summe 20 µl
Temperaturprofil für die reverse Transkription
Temperatur Dauer
37° C 60 min
3.2.3.3 PCR
Die gewünschten cDNA-Abschnitte wurden nun entweder mit dem Redextract-Kit
oder dem Quiagen-Taq PCR Core-Kit vervielfältigt, je nach verwendetem
Primerpaar.
Ansatz pro Probe
Redextract 5 µl
H2O 2,5 µl
Forward-Primer RPS 9 (13,7 µM) 0,25 µl
Reverse-Primer RPS 9 (13,7 µM) 0,25 µl
cDNA 2 µl
Summe 10 µl
Ansatz pro Probe
Q-Solution 2 µl
10x PCR-Buffer 1 µl
dNTPs 2 µM 1 µl
Forward-Primer (13,7 µM) 1,5 µl
Reverse-Primer (13,7 µM) 1,5 µl
Taq-Polymerase 0,05 µl
cDNA 2,95 µl
Summe 10 µl

Eigene Untersuchungen
34
Anhand von bereits publizierten Sequenzen verschiedener MNV-Stämme (MNV 1 -
4) wurden passende Primerpaare in einer genetisch konservierten Region entwickelt,
mit denen möglichst viele Varianten von MNV reagieren; zusätzlich wurden
Primerpaarsequenzen aus Publikationen übernommen.
Tabelle 10: Primmerpaare zum Nachweis von MNV mit Hilfe des Quiagen-Taq PCR
Core-Kit (Primersequenzen siehe S. 22f.)
Name des Primerpaars Länge des Amplikon
5205f/5388r 183 bp
5004f/5219r 215 bp
Nach Sequenzierung getestet:
5004f/5277r 273 bp
5580f/5671r modifiziert (MÜLLER et al. 2007) 91 bp
4972f/5080r (BAERT et al. 2008) 608 bp
5205f/5469r 264 bp
5473/5659r modifiziert (HSU et al. 2006) 186 bp
Die Probenansätze wurden anschließend mit nachfolgendem Temperaturprofil im
Thermocycler zur Reaktion gebracht.
Temperaturprofil für die MNV-PCR
Temperatur Dauer
95° C 4 min
94° C 1 min
55° C 1 min
72° C 2 min
39x Schritt 2-4
72° C 7 min
8° C Bis zur Entnahme
Zur Kontrolle der RNA-Umschreibung zu cDNA wurde das Housekeeping-Gen RPS-
9 mit Hilfe des Primerpaares RPS9f/RPS9r nachgewiesen. Hierbei wurde das
Redextract-Kit verwendet.

Eigene Untersuchungen
35
Temperaturprofil für die RPS-9-PCR
Temperatur Dauer
95° C 4 min
94° C 1 min
55° C 1 min
72° C 2 min
24x Schritt 2-4
72° C 7 min
8° C Bis zur Entnahme
3.2.4 Nachweis von Helicobacter hepaticus mittels PCR
Die Gesamt-DNA wurde aus den im Infektionsversuch gewonnenen Kotproben mit
Hilfe des Invitek PSP® Spin Stool DNA Kit gemäß Herstellerprotokoll isoliert. Für die
Detektion kamen die Primerpaare H276f/H676r und B38/B39 zum Einsatz. Mit dem
Primerpaar H276f/H676r lassen sich 16S rRNA Gensequenzen der Helicobacter spp.
nachweisen (RILEY et al. 1996). Alle Helicobacter spp. ergeben ein 374-
Basenpaarfragment. Das Primerpaar B38/B39 weist eine H. hepaticus spezifische
16S rRNA-Gensequenz nach (SHAMES et al. 1995). Das Basenpaarfragment ist 417
bp groß. Für beide Primerpaare wurde das Redextract-Kit verwendet.
Ansatz pro Probe
Redextract 5 µl
H2O 2,5 µl
Forward-Primer (20 µM) 0,25 µl
Reverse-Primer (20 µM) 0,25 µl
cDNA 2 µl
Summe 10 µl
Temperaturprofil für die Helicobacter spp.- / H. hepaticus - PCR
Temperatur Dauer
94° C 3 min
94° C 0,5 min
53° C 0,5 min
72° C 0,5 min
34x Schritt 2-4
72° C 10 min
8° C Bis zur Entnahme

Eigene Untersuchungen
36
3.2.4.1 Agarosegel-Elektrophorese
Für ein 1,5%iges Gel wurden 2,25 g Agarosepulver in 150 ml 1x TBE-Puffer gegeben
und in der Mikrowelle erhitzt. Unter ständigem Rühren mittels Magnetrührer wurde
die Agaroselösung auf 50° C abgekühlt. Dann wurden der Lösung 6 µl Gelstar DNA-
Farbstoff zugefügt. Die Flüssigkeit wurde dann bis zu einer Höhe von 0,5 cm in den
mit bis zu zwei 25er-Taschen-Kämmen präparierten Gelschlitten gegossen. Nach
einem Auskühlen von 30 min konnten die Kämme gezogen werden, und das Gel war
einsatzbereit.
Für die Elektrophorese wurde das Gel in die mit 1x TBE-Puffer gefüllte Gelkammer
gelegt. Danach wurden die dem Thermocycler entnommenen Proben mit 4 µl Orange
G Ladepuffer versehen, sofern der PCR-Ansatz mit dem Quiagen-Taq PCR Core-Kit
erstellt wurde. Ein mit Redextract erstellter PCR-Ansatz enthält bereits Ladepuffer.
Je nach Ansatz wurden pro Tasche 10-14 µl Probenvolumen einpipettiert. Zur
Größendifferenzierung der Basenfragmente wurde in mindestens eine Tasche pro
Kamm 6 µl einer standardisierten Basenleiter gegeben. Die anschließende Laufzeit
bei 180 V 250 mA betrug ca. 45 min. Zur Auswertung wurde das Gel abschließend
mit einem UV-Transilluminator betrachtet und digital fotografiert.
3.2.5 Sequenzierung
Die Klonierung und Vorbereitung für eine externe Sequenzierung wurde durch Herrn
Dr. Nils-Holger Zschemisch vom ZTL durchgeführt. Aus Zellkulturüberstand isolierte
MNV-RNA wurde mit dem Oligo(dT)18-Primer zu cDNA umgeschrieben. Diese cDNA
wurde als Template verwandt. Die Primerpaare für die PCR-Produkte, die kloniert
wurden, sind Tabelle 6 (S.23) zu entnehmen. Der Primer Noro Hannover rev 3
entspricht dem Primer 15T-aTAG (MÜLLER et al. 2007). Die Klonierung und
Transformation erfolgte gemäß Anweisungen und gelieferten Materialien des
StrataCloneTM PCR Cloning Kit von Agilent Technologies. Detaillierte Protokolle sind
zusätzlich in der Dissertation von Herrn Dr. Zschemisch zu finden (ZSCHEMISCH
2004).

Eigene Untersuchungen
37
Die Bakterienkulturen mit erfolgreich klonierten Abschnitten des MNV-Genoms in
Plasmidform wurden mit dem QIAprep Spin Miniprep Kit gemäß Anweisungen des
Herstellers aufgearbeitet. Abschließend erfolgte eine PCR mit den Primerpaaren, die
in Tabelle 7 (S.23) genannt werden. Die Sequenzierung der drei PCR-Produkte
wurde extern durch die Firma MWG Biotech AG durchgeführt. Eine Analyse der
erhaltenen Sequenzen erfolgte auf einem Macbook mit DNA Dynamo und
internetbasiert mit BLAST (ALTSCHUL et al. 1990).
3.2.6 Virus-Anzucht für den Infektionsversuch
Eine Portion des Virusisolats wurde zweimal mit RAW-Zellen passagiert, von dieser
Passage wurden für Infektionsversuche und weitere Virusanzucht erneut 1 ml
Portionen in sterile Glasfläschchen abgefüllt und bei -80° C tiefgefroren.
3.2.7 Virus-Titer-Bestimmung
3.2.7.1 Plaque-Test
Um den Virus-Titer zu bestimmen, wurde ein Plaque-Test durchgeführt. Hierzu
wurden RAW 264.7 Zellen auf einer 6er-Zellkultur-Platte in einer Dichte von 6 x 105
Zellen/ml ausgesät. Die Zelldichte pro ml wurde durch Auszählung mit einer
Neubauer Zellzählkammer bestimmt. Pro Vertiefung der Platte wurden 2,5 ml
aufgetragen, was einer Gesamtzellzahl von 1,5 x 106 Zellen pro Vertiefung
entsprach. Die Zellen wurden über Nacht im Brutschrank bei 5% CO2 und 37° C
inkubiert. Am nächsten Tag wurden Zehner-Verdünnungen der Virussuspension für
den Infektionsversuch angefertigt. Das ursprüngliche Zellmedium wurde von den
Platten abgesaugt. Danach wurden pro Verdünnung 2 Vertiefungen als
Doppelbestimmung mit 500 µl der angefertigten Virusverdünnungsreihe beimpft. Die
Platten wurden danach eine Stunde im Brutschrank bei 5% CO2 und 37° C bebrütet.
Währenddessen wurde eine Lösung aus 2,5% SeaPlaque Agarose und ergänztem
DMEM-Medium (10% FKS LE, 2% Penicillin-Streptomycin) hergestellt. Nach der
einstündigen Inkubation wurde die Virussuspension von den Vertiefungen abgesaugt
und durch 2,5 ml der vorbereiteten Agarose-Medium-Mischung ersetzt. Nachdem die

Eigene Untersuchungen
38
Agarose-Mediummischung bei Raumtemperatur erstarrt war, wurden die Platten bei
5% CO2 und 37° C für 48h bebrütet. Um die Auswertung zu erleichtern, wurden die
Platten mit einer 5% Kristallviolett-Lösung angefärbt.
3.2.7.2 Auswertung eines Plaque-Tests
Die Löcher im Zellrasen werden bei zwei bis drei auszählbaren Verdünnungsstufen
der Virussuspension erfasst und mit dem Kehrwert der Verdünnungsstufe
multipliziert. Hierbei erhält man die Anzahl von plaqueforming units pro inokkuliertes
Volumen der Virussuspension.
3.2.7.3 TCID50-Test
Hierzu wurden RAW 264.7 Zellen in der Konzentration von 3 x 104 Zellen/Vertiefung
auf eine Reaktionsplatte mit 96 Vertiefungen aufgetragen. Pro Vertiefung wurden 100
µl Zellsuspension aufgetragen, dies entspricht einer Zellkonzentration der
Suspension von 3 x 105 Zellen/ml. Die Vertiefungen am Plattenrand wurden mit 300
µl Medium befüllt, um einer Verdunstung im Randbereich vorzubeugen. Die Platten
wurden über Nacht bei 5% CO2 und 37° C inkubiert. Am folgenden Tag wurde eine
Verdünnungsreihe der Virussuspension in Zehnerschritten angelegt. Je 200 µl einer
Verdünnungsstufe der Virussuspension wurden pro Vertiefung auf der
Reaktionsplatte zugegeben. Je eine Reihe mit 10 Vertiefungen wurde mit einer
Verdünnungsstufe befüllt. Eine Kontrollreihe bekam statt der Virussuspension 200 µl
Medium. Dies entsprach einem Gesamtvolumen von 300 µl Flüssigkeit pro
Vertiefung. Danach erfolgte eine weitere Bebrütung bei 5% CO2 und 37° C über acht
Tage.
3.2.7.4 Auswertung eines TCID50-Tests
Zur Auswertung wurde der CPE optisch erfasst. Bereits der dem Nährmedium
zugesetzte Indikator Phenolrot ließ eine Unterscheidung zwischen toten (rosa) und
lebenden (gelb) RAW-Zellen in den Vertiefungen zu, zur Erleichterung wurde jedoch
auch hier mit einer 5% Kristallviolett-Lösung angefärbt.

Eigene Untersuchungen
39
Die Berechnung der Viruskonzentration erfolgt mit der Formel nach Spearmann und
Kärber.
Zuerst wurde die Verdünnungsstufe bestimmt, in der noch ein vollständiger CPE in
allen Vertiefungen zu ermitteln ist. Diese Verdünnungsstufe ergab die Variable a (bei
10-6 ist diese gleich -6). Für die Variable b wurden die Anzahl Vertiefungen mit CPE
der niedrigeren Verdünnungsstufen zu der Anzahl Vertiefungen der
Verdünnungsstufe mit vollständigem CPE addiert (10+9+2+1=22). Die Variable c war
gleich der Gesamtzahl an Vertiefungen pro Verdünnungsstufe (in diesem Fall c=10).
Dies entsprach 107,7 TCID50/200 µl oder 2,5 x 108 TCID50/ml.
3.2.8 Immunofluorescence Assay (IFA)
Dieser Test dient dazu, in einem Mausserum möglicherweise vorhandene Antikörper
gegen MNV nachzuweisen. Entwickelt und validiert wurde der Test bei der Firma
Biodoc. Verdünntes Maus-Serum wird hierbei auf Objektträger aufgetragen, die mit
fixierten MNV-infizierten Zellen beschichtet sind. Die Auswertung erfolgt optisch mit
Hilfe eines Fluoreszenzmikroskops.
3.2.8.1 Herstellung und Anwendung der Objektträger für die Immunfluoreszenz
Für diesen Test wurden RAW-Zellen in der Flasche kultiviert. Bei einer Zelldichte von
75% wurden diese mit 0,5 ml der eingefrorenen Virussuspension infiziert. Nach einer
flexiblen Inkubationszeit von 4 bis 8 Stunden wurde das Medium entfernt, die Zellen
abgeschabt und in 25 ml neuem Medium (DMEM, 2% Penicilin-Streptomycin, 10%
LE-FKS) pro Flasche suspendiert. Diese Zellsuspension wurde in 50 µl-Tropfen auf
teflonbeschichtete 8-Loch-Objektträger aufgetragen. Danach erfolgte eine weitere

Eigene Untersuchungen
40
Inkubation der Objektträger für 4 bis 8 Stunden im Brutschrank, bis die erwünschte
Infektionsrate von 10 - 30% infizierter Zellen im Zellrasen erreicht war. Die
Infektionsrate wurde zwischendurch durch Anfertigen von Probeobjektträgern gemäß
dem Anwendungsprotokoll (siehe weiter unten) kontrolliert. Bei Erreichen der
gewünschten Infektionsrate wurde das Medium von den Objektträgern abgesaugt.
Die Objektträger wurden an der Luft getrocknet und dann in einem Aceton-Bad für 10
min fixiert. Danach erfolgte eine abschließende Lufttrocknung. Die Objektträger
wurden bis zum Gebrauch bei -20° C gelagert.
Anwendungsprotokoll der Objektträger bei Raumtemperatur:
Es wurden 32 µl des 1:20 mit PBS verdünnten Mausserums pro Kavität aufgetragen.
Anschließend folgten 20 Minuten Inkubationszeit. Dann wurde das Serum
abgesaugt, und es wurden 50 µl PBS pro Kavität zum Spülen aufgetragen. Nach
einer Inkubationszeit von 10 Minuten wurde das PBS abgesaugt. Erneut wurden 50
µl PBS pro Kavität zum 2. Spülen aufgetragen. Nach einer Inkubationszeit von 10
Minuten wurde das PBS abgesaugt. Jetzt wurden 25 µl des in PBSM 1:3000
verdünnten Fluoreszenz Anti-Maus-IGG-Antikörpers pro Kavität aufgetragen und für
20 Minuten inkubiert. Nachdem die Antikörperlösung abgesaugt worden war, wurden
50 µl PBS pro Kavität zum Spülen aufgetragen. Nach einer Inkubationszeit von 10
Minuten wurde das PBS abgesaugt. Pro Kavität wurden erneut 50 µl PBS zum 2.
Spülen aufgetragen und nach einer Inkubationszeit von 10 Minuten abgesaugt.
Abschließend wurden 50 µl Evan´s Blue pro Kavität zum Anfärben des
Zellhintergrunds aufgetragen, die nach einer Inkubationszeit von 5 Minuten wieder
abgesaugt wurden. Der Objektträger konnte dann mit einem Fluoreszenzmikroskop
bei 320 facher Vergrößerung optisch beurteilt werden.
3.2.9 Virusaufreinigung mittels Ultrazentrifugation
MNV wurde wie bereits beschrieben angezüchtet. Nach zwei Einfrier-Auftauzyklen
wurde der Flascheninhalt bei 6000 U/min für 30 min (2800 x g) zentrifugiert. Der
Überstand wurde abgenommen, das Pellet verworfen. Die weitere Aufreinigung

Eigene Untersuchungen
41
erfolgte dann mit der Ultrazentrifuge. Zum Einsatz kam hierbei ein SW32-Rotor mit
sechs Bechern. In einem matten Ultrazentrifugenröhrchen wurden 25 ml Überstand
mit 5 ml einer 30% Sucrose-Lösung unterschichtet. Danach wurde für drei Stunden
bei 27000 U/min (89500 x g) und 4° C zentrifugiert. Nach sorgfältigem Abgießen des
Überstandes und der Sucrose-Lösung war ein hauchdünnes Pellet am Boden des
Ultrazentrifugenröhrchens zu sehen, das in je 50 µl PBSM resuspendiert wurde.
3.2.9.1 Weitergehende Aufreinigung über einen CsCl-Gradienten im SW55-
Rotor
Hierfür wurde eine CsCl-Lösung mit einer Dichte von 1,3393 g/cm³ angesetzt (5,15 g
CsCl auf 10 g H2O). In 9,5 ml dieser Lösung wurden 0,5 ml der PBSM-MNV-
Suspension gegeben. Je 5 ml wurden auf zwei klare Ultrazentrifugenröhrchen
verteilt. Die Zentrifugation im SW55-Rotor erfolgte für 18 Stunden bei 35000 U/min
(116000 x g) und 4° C. Die sichtbare Bande wurde im Gegenlicht mit einer Spritze
abgenommen und bei -20° C für eine weitere Verwendung gelagert.
3.2.9.2 Aufreinigung für elektronenmikroskopische Aufnahmen
MNV wurde wie bereits beschrieben angezüchtet. Nach zwei Auftauzyklen wurde der
Flascheninhalt bei 6000 U/min für 30 min (2800 x g) zentrifugiert. Insgesamt 50 ml
des Zellkultur-Überstands wurden abgenommen, das Pellet verworfen. Nun kam ein
anderes Aufreinigungsprotokoll zum Einsatz, da CsCl-Ionen eine elektronen-
mikroskopische Aufnahme erschweren. Je 2 ml Zellkulturüberstand wurden bei
55000 U/min für 32 min (201000 x g) in einem SW-TLS 55 Rotor zentrifugiert. Der
Großteil des Überstandes wurde verworfen. Die Pellets wurden in 20 µl
Restüberstand gelöst. Insgesamt entstanden so 500 µl aus 50 ml. Von der
Pelletsuspension wurden 125 µl mit 1,875 ml steriler Kochsalzlösung aufgefüllt.
Diese Lösung wurden erneut bei 55000 U/min für 32 min im SW-TLS 55 Rotor
zentrifugiert. Der Überstand wurde verworfen. Die hierbei entstandenen Pellets
wurden in 20 µl Restüberstand gelöst. Anschließend erfolgte ein dritter
Zentrifugationsschritt bei 55000 U/min für 32 min im SW-TLS 55 Rotor.

Eigene Untersuchungen
42
3.2.9.3 Gewinnung von RAW-Zellbestandteilen für eine Zellkontrolle
Parallel zur MNV-Anzüchtung wurden Zellkulturflaschen mit geschlossenem RAW-
Zellrasen (Zelldichte ähnlich der zu infizierenden Zellkulturflaschen) bei -20° C
eingefroren. Die weitere Behandlung entsprach den Schritten der Virusaufreinigung
mittels Ultrazentrifugation. Auch hier konnte visuell ein Pellet am Boden des
Ultrazentrifugenröhrchens ausgemacht werden, das in je 50 µl PBSM resuspendiert
wurde.
3.2.10 Enzyme-linked Immunosorbent Assay (ELISA)
Dieser Test dient dazu, in einem Mausserum möglicherweise vorhandene Antikörper
gegen MNV nachzuweisen. Verdünntes Maus-Serum wird hierbei auf mit MNV-
Antigen beschichtete Microtiter-Riegel aufgetragen. Die Auswertung der
enzymatischen Umsetzung des Substrats ABTS durch Peroxidase-markierte Anti-
Maus-IgG-Antikörper erfolgt optisch mit Hilfe eines Photometers. Bei einer
Substratumsetzung färbt sich die Lösung grün.
3.2.10.1 Beschichtung (Coating) und Anwendung der ELISA-Platten
Die durch die Aufreinigung gewonnene Virus-PBSM-Lösung wurde mit
Beschichtungspuffer (Coating Buffer) 1:100 verdünnt. Von der Coating-Lösung
wurden 150 µl pro Vertiefung der 8er-Microtiterriegel aufgetragen. Eine Platte besteht
aus 12 Riegeln mit 8 Vertiefungen. Danach wurden die ELISA-Platten für 24 Stunden
im Kühlschrank bei 8° C inkubiert. Der Inkubation folgten ein Waschen und ein
Blockvorgang mit 200 µl Roche-Blocking Reagent pro Vertiefung über 15 min bei
Raumtemperatur gemäß Gebrauchsanweisung des Herstellers. Zum Abschluss
wurden die Riegel erneut gewaschen. Die Riegel der RAW-Zellkontrolle wurden
genau wie die MNV-Riegel beschichtet und anschließend geblockt. Verdünnt in
Coating-Buffer wurde hier stattdessen das in PBSM gelöste RAW-Pellet. Hierbei
wurde die gleiche Verdünnungsstufe wie beim MNV-Coating (1:100) verwendet. Die
ELISA-Platten wurden dann bis zum Gebrauch bei -20° C gelagert.

Eigene Untersuchungen
43
Das Standardwaschprogramm des Columbus Washer, das sowohl nach dem
Coating als auch beim Elisa-Standardprotokoll zur Anwendung kam, sah 3
Waschschritte mit jeweil 250 µl PBS Tween vor. Das PBS Tween wurde direkt nach
dem Einfüllen in die Vertiefungen wieder abgesaugt.
Beim Standard-Protokoll zur Anwendung der beschichteten ELISA-Riegel wurden
100 µl des 1:50 mit PBS verdünnten Mausserums in je eine MNV- und eine RAW-
Vertiefung pipettiert. Nach einer Inkubation von 45 Minuten bei 37° C im Brutschrank
wurden die Vertiefungen mit PBS Tween im Washer gespült. Anschließend wurden
100 µl Zweitantikörper 1:1000 verdünnt in PBS Tween pro Vertiefung aufgetragen.
Es folgte eine Inkubation über 45 Minuten bei 37° C im Brutschrank. Nach einem
erneuten Waschen mit PBS Tween wurden schließlich 100 µl ABTS-Substrat pro
Vertiefung aufgetragen. Nach 45 Minuten Inkubation bei 37° C im Brutschrank wurde
die Extinktion im Photometer beim Wellenlängenpaar 405 nm vs. 492 nm gemessen.
Die Verdünnungsstufe des Antigens zur Beschichtung wurde durch eine Antigen-
Verdünnungsreihe ermittelt. Virus-PBSM-Lösung wurde in verschiedenen
Konzentrationen mit Coating Buffer verdünnt und in die Vertiefungen gemäß
Beschichtungsprotokoll gegeben. Mit einem positiven Kontrollserum-Pool wurde das
Standardprotokoll zur Anwendung der beschichteten ELISA-Riegel abgearbeitet.
Dieser positive Kontrollserumpool war zuvor aus in der IFA deutlich positiv
getesteten Mäuseseren erstellt worden. Ziel der Verdünnungsreihe war es, mit einer
möglichst hohen Verdünnung des Antigens dennoch einen hohen OD-Wert zu
erreichen.
3.2.10.2 Der korrigierte OD-Wert
Um eventuelle Kreuzreaktionen mit RAW-Zellbestandteilen auszuschließen, wurde
die Zellkontrolle eingeführt. Diese ermöglicht die Bestimmung eines korrigierten OD-
Werts:
OD MNV-Kavität – ODRAW-Kavität = ODkorrigiert

Eigene Untersuchungen
44
3.2.11 Elektronenmikroskopische Aufnahmen
Nach der bereits beschriebenen Virusaufreinigung für elektronenmikroskopische
Aufnahmen wurden durch Jutta Milzer im Institut für Virologie der MHH
elektronenmikroskopische Präparate hergestellt.
Unbeschichtete Trägerplättchen, sogenannte Grids, wurden auf einen Formvarfilm
(Formvarpulver gelöst in Chloroform) auf einer Wasserfläche aufgebracht.
Anschließend wurden sie mit einem Filterpapierstreifen abgenommen und luftfixiert.
Danach erfolgte eine Bedampfung mit Graphit. Auf die vorbereiteten Grids wurden 20
µl der konzentrierten Viruspartikel-Lösung aufgetragen. Nach sechsmaligem
Waschen mit Aqua ad iniectabilia wurden 5 µl 1% Uranylacetat (gelöst in Wasser) für
die Negativkontrastierung aufgetragen. Abschließend wurden die Grids luftfixiert.
Drei Präparate (Grids) wurden im Zentrallabor Elektronenmikroskopie der MHH mit
einem FEI Tecnai 20 Elektronenmikroskop im TEM-Modus bei 80 kV in 62000-facher
und 80000-facher Vergrößerung untersucht und digital gespeichert. Dies geschah mit
Unterstützung von Prof. Dr. Ungewickell, Gerhard Preiß und Jutta Milzer.
3.2.12 Isolation und Anzucht von Helicobacter hepaticus für den Doppel-
infektionsversuch
Der Keim wurde im mikrobiologischen Untersuchungslabor des ZTL isoliert und
identifiziert. Hierzu wurde die Maus Nummer 1240 des Stammes BC-R4-Il10-/- nach
CO2 Betäubung durch Genickbruch getötet. Nach Eröffnung der Bauchhöhle wurde
die Zäkumspitze mit sterilem Besteck entfernt und in ein 5 ml Polypropylen-Röhrchen
mit 2 ml Brain-Heart-Infusion Medium überführt. Nach Homogenisierung wurde die
Suspension durch einen 0,45 µm Weißrandfilter gegeben. Von der gefilterten Lösung
wurden jeweils 200 µl auf eine Petrischale mit Columbiaagar mit Zusatz von
Amphotericin B, Campylobacter-Selektiv-Supplement (nach Skirrow) und Pferdeblut
getropft und mit einer Impföse verteilt. Die Platten wurden unter mikroaerophilen
Bedingungen in einem Anaerobiertopf bei 37° C kultiviert. Zur Erzeugung der
mikroaerophilen Bedingungen wurde die Luft viermalig evakuiert auf -0,2 bar.

Eigene Untersuchungen
45
Danach wurde ein Gasgemisch, bestehend aus 10% CO2, 10% H2 und 80% N2, bis
0,2 bar hinzugefügt. Die Bakterien wurden alle 3 bis 4 Tage auf eine neue Platte
überimpft, indem etwas Bakterienmaterial mit einer Impföse von der alten Platte
abgenommen und auf einer neuen Platte verteilt wurde. Die vierte Passage dieses
Keimes wurde in großem Umfang als Dauerkultur bei -80° C eingefroren, da hieraus
die Bakterien zur Infektion gewonnen wurden. Zum Einfrieren der Bakterien wurde
auf jede Platte 1 ml brain-heart-infusion (BHI)-Medium gegeben. Anschließend
wurden die Bakterien mit Hilfe eines Wattetupfers in das Medium eingerieben. Von
dieser Suspension wurden 800 µl abgenommen und in ein 1,5 ml
Schraubdeckelgefäß mit 100 µl Gycerin und 100 µl FKS überführt. Mehrere gefüllte
Schraubdeckelgefäße wurden bei -80° C eingefroren.
3.2.13 Experimentelle Infektion der Mäuse mit MNV
Die Mäuse wurden anhand ihres Alters in die Infektionsgruppen (siehe Tabelle 11)
eingeteilt, um zum geplanten Sektionstermin ein möglichst gleiches Alter (12-15
Wochen) zu haben. Sie wurden im Alter von 7 bis 12 Wochen zweimal
hintereinander im Abstand von 3 Tagen mit jeweils 0,1 ml virushaltigem
Zellkulturüberstand 2,5x108 TCID50/ml per oraler Gavage und 10 µl per nasaler
Applikation experimentell infiziert. Für die Gavage wurde eine Knopfkanüle
eingesetzt. Die Infektion erfolgte unter einer sterilen Sicherheitswerkbank. Die
Kontrolltiere erhielten 0,1 ml PBS per oraler Gavage und 10 µl PBS nasal. Sie
wurden als Einzelgruppe zusammengefasst und zu einem gesonderten
Sektionstermin getötet und seziert.
3.2.14 Experimentelle Infektion der Mäuse mit Helicobacter hepaticus
Zwei Wochen nach der Inokulation mit MNV wurden Mäuse im letzten Teil
(Infektionsversuch 5) der Versuchsreihe ebenfalls per oraler Gavage mit 0,1 ml einer
H. hepaticus – Suspension (106 KBE, 7. Passage) experimentell inokuliert. Fünf Tage
später erfolgte eine zweite Inokulation. Die Kontrolltiere (infiziert mit MNV) erhielten
zweimal 0,1 ml PBS per oraler Gavage.

46
Tabelle 11: Übersichtsdiagramm der Infektionsversuche
1. Versuch
Stamm
Geschlecht (Tiere pro Sektions-termin) Sektion p.i. Kontrollgruppe Probenverwendung Besonderheit
C3-Il10-/-
m (2+3+3),w (3+4+6) 2, 4, 8 Wochen
m (4), w (2+2) Serologie, Histologie, PCR, Lymphozytenstimulation
Lymphozytenstimulation zusammengefasst pro Stamm und Geschlecht B6-Il10
-/- m (5+6+5),w (5+5+4) m (3), w (3+3)
2. Versuch
C3-Il10-/-
m (4+4),w (3+4)
2, 4 Wochen
m (3), w (2)
Serologie, Histologie, PCR, Lymphozytenstimulation
Lymphozytenstimulation pro Einzeltier für alle weiteren Versuche B6-Il10
-/- m (6+4),w (6+4) m (4), w (5)
C3-wt m (2+1(1),w (0+0) m (5), w (0)
B6-wt m (4+4),w (6+6) m (4), w (6)
3. Versuch (Ergänzung zum 2. Versuch, sowie Untersuchung des akuten Infektionsverlaufs)
C3-Il10-/-
w (3) 2 Wochen w (3)
Serologie, Histologie, PCR, Lymphozytenstimulation
B6-Il10-/-
w (3) 4 Wochen -
C3-wt m (2) 2 Wochen -
B6-wt - - -
B6-Il10-/-
m (4) 7d
m (4) Serologie, Histologie, PCR, Lymphozytenstimulation B6-wt m (4), w (4) m (4), w (4)
B6-wt w (4+4+3+3) 3, 6, 9, 12 d 0 Serologie, Histologie, PCR Verlaufsversuch, Darmlymphknoten für PCR verwendet
4. Versuch (Infektion keimfreier Tiere)
C3-Il10-/-
w (5) 4 Wochen m (4) Serologie, Histologie, PCR, Lymphozytenstimulation
keimfreie Tiere, im Micro-Isolator gehalten
5. Versuch (Doppelinfektion)
C3-Il10-/-
m (2), w (4) 8 (6) Wochen
m (2), w (4) Serologie, Histologie, PCR, Lymphozytenstimulation
Infektion mit MNV, sowie 2 Wochen später mit H. hepaticus
B6-Il10-/-
w (6) w (5) Kontrollen: Infektion nur mit H. hepaticus
m = männlich; w = weiblich; p.i. = post inoculationem; d = Tag

Eigene Untersuchungen
47
3.2.14.1 Tötung
Am Sektionstag wurden die Tiere von der Infektionsabteilung in das mikrobiologische
Untersuchungslabor des ZTL der MHH gebracht. Dort wurden sie durch
Kohlendioxidgasinhalation betäubt und anschließend durch Entbluten getötet.
3.2.14.2 Postmortale Untersuchung
Die Tierkörper wurden gewogen, auf äußerliche Veränderungen untersucht und
anschließend seziert.
3.2.14.3 Sektion
Die Sektion erfolgte mit sterilem Sektionsbesteck. Dabei wurden alle Organe auf
makroskopische Veränderungen untersucht.
3.2.14.4 Probennahme
Serum
Nach Eröffnen des Brustkorbs des Tieres wurde das Herz mit einer Spritze mit
aufgesetzter Kanüle punktiert, um möglichst viel Vollblut zu gewinnen. Das Blut
wurde in sterile Eppendorf-Gefäße gegeben und bei Raumtemperatur bis zum Eintritt
der Gerinnung stehen gelassen. Dann erfolgte eine Zentrifugation bei 6000 U/min
(2800 x g) für 7 min. Der Serumteil wurde abgenommen und für weitere
Untersuchungen bei -80° C gelagert.
PCR
Als PCR-Proben wurden bei jedem Versuch mit sterilem neuen Besteck Lunge,
Leber, Niere, Milz, Mesenteriallymphknoten (nur Versuch 3; Tag 3, 6, 9, 12 p.i.),
proximaler Dünndarm, Ileum, proximales Kolon und Kot genommen. Die Proben
wurden in sterile Eppendorf-Gefäße gefüllt und sofort in flüssigem Stickstoff gekühlt
bis zur anschließenden Lagerung bei -80° C.

Eigene Untersuchungen
48
Histologie
Für die Histologie wurden Zäkum, Kolon, proximaler Dünndarm, distaler Dünndarm,
Magen, Leber, Lunge und Milz entnommen. Die Proben wurden in
Standardeinbettkassetten gelegt und abschließend bis zur Zuschneidung in
10%igem, gepufferten Formalin nach Sörensen gelagert. Das Kolon wurde dabei als
„modifizierte Swiss-Roll“ in die Kassette gebettet (MOOLENBEEK u. RUITENBERG
1981; BLEICH et al. 2004). Hierzu wurde das proximale Kolon in die Mitte der
Kassette verbracht und das restliche Kolon sowie Rektum und Anus in Form einer
Schnecke flach ausgerollt.
3.2.14.5 Präparation des Mesenteriallymphknotens
Der Mesenteriallymphknoten (MLK) wurde mit sterilem Besteck präpariert und in mit
5 ml Lymphozyten-Medium gefüllte verschließbare 15 ml Plastik-Reaktionsgefäße
gegeben und bis zur weiteren Aufarbeitung auf Eis gelagert. In Versuch 1 wurden die
MLK pro Stamm und Geschlecht in einem Gefäß zusammengefasst, in den
folgenden Versuchen wurde jeder MLK pro Tier separat aufgearbeitet.
3.2.15 Sterile Aufarbeitung des Mesenteriallymphknotens unter einer Werkbank
Der MLK wurde mit Lymphozytenmedium auf ein steriles Zellsieb gegeben, welches
in einer sterilen Petrischale lag. Anschließend wurde das Gewebe im Sieb
gleichmäßig mit dem sterilen Stempel einer 1 ml-Spritze homogenisiert. Die
Lymphozyten wurden dabei freigesetzt und konnten durch das Sieb ins Medium
passieren. Nach mehrmaligem Auf- und Abpipettieren wurde das Medium mit den
suspendierten Lymphozyten in ein neues steriles BlueCap gegeben. Dann erfolgte
eine Pelletierung der Lymphozyten durch Zentrifugation bei 4° C und 1100 U/min
(163 x g) für 7 min. Das Medium wurde verworfen, das Pellet erneut in 5 ml Medium
gelöst und in ein neues Reaktionsgefäß übertragen. Nach einer erneuten
Zentrifugation bei 4° C und 1100 U/min (163 x g) für 7 min wurde auch hier das
Medium verworfen, und das gewaschene Pellet in 0,5 ml Medium gelöst. Zur
Bestimmung des Lymphozytengehalts pro Volumen wurde ein scil VetABC

Eigene Untersuchungen
49
eingesetzt, um die Lymphozyten-Konzentration der jeweiligen Mesenterial-
Lymphknotensuspension auf 2 x 106 Zellen/ml einzustellen.
3.2.16 Vorbereitung und Verwendung der Zellkulturplatten für die Bestimmung von
Interferon γ
Am Tag vor der Sektion wurden Reaktionsplatten mit 96 Vertiefungen mit 100
µl/Vertiefung der PBS-anti-mCD3-Lösung [5 µg anti-mCD3 (145-2C11)/ml]
beschichtet. Diese wurden über Nacht bei 4° C inkubiert. Abschließend erfolgte am
Sektionstag ein zweimaliges Waschen mit 200 µl Lymphozytenmedium/Vertiefung.
Danach waren die Platten verwendungsbereit.
Die vorbereitete Platte wurde entleert und 2 x mit Lymphozytenmedium gespült. Die
äußeren Vertiefungen wurden zum Verdunstungsschutz mit 200 µl
Lymphozytenmedium versehen. Jeweils 100 µl der Mesenteriallymphknoten-
suspensionen (2 x 105 Zellen) wurden anschließend in die freien Vertiefungen
aufgetragen, pro Suspension 3 Vertiefungen. Zusätzlich wurden 100 µl Medium
hinzugefügt. Die Platten wurden für 24 h bei 37° C, 5% CO2 und gesättigter H2O-
Atmosphäre bebrütet. Nach der Bebrütung wurden je 100 µl der Plattenüberstände
auf zwei neue unbeschichtete Platten verteilt. Diese wurden bis zur externen INFγ-
Bestimmung durch einen ELISA bei -80° C gelagert.
3.2.17 Bestimmung von Interferon γ im Lymphozytenzellkulturüberstand mit Hilfe
eines ELISA
Die Bestimmung erfolgte durch die Arbeitsgruppe von PD Dr. Detlef Neumann vom
Institut für Pharmakologie der MHH (NEUMANN et al. 2006).
Zuerst wurde der Erstantikörper in der Konzentration 0,5 µg/ml in Coating Buffer
verdünnt. Dann wurden je 100 µl der Verdünnung pro Vertiefung auf die Platten
aufgetragen. Anschließend erfolgte eine Inkubation über Nacht bei 4° C. Am
nächsten Tag wurde die Platte entleert und 3 x gewaschen. Zum Blocken wurden
250 µl Blockpuffer pro Vertiefung für eine Stunde bei Raumtemperatur aufgetragen.

Eigene Untersuchungen
50
Für die Standards wurde die 100 ng/ml Stammlösung IFNγ in Kulturmedium 1:100
auf 1000 pg/ml verdünnt. Für weitere Verdünnungen wurde 1:2 weiterverdünnt bis
zur höchsten Verdünnungsstufe von 16,5 pg/ml, insgesamt 7 Stufen. Im
Doppelansatz wurden 50 µl pro Vertiefung jeder Verdünnungsstufe sowie eine
zusätzliche Blindprobe aufgetragen.
Die Proben wurden ebenfalls mit Medium verdünnt und doppelt aufgetragen. Nach
einer einstündigen Inkubation wurden 50 µl biotinylierter Zweitantikörper in einer
Verdünnung von 1:1000 pro Vertiefung zugegeben. Nach erneuter einstündiger
Inkubation bei Raumtemperatur wurde die Platte entleert und 3mal gewaschen. Nach
Zugabe von 100 µl verdünnter Streptavidin-HPRT-Lösung wurde für 30 min bei
Raumtemperatur inkubiert. Anschließend wurde die Platte entleert und 4mal
gewaschen. Pro Vertiefung wurden 100 µl Substrat zugegeben und unter
Beobachtung bei RT ca. 20 min inkubiert. Dann wurde die Reaktion mit 50 µl 1M
Schwefelsäure gestoppt.
Die Extinktion wurde im ELISA-Reader bei 450 nm vs. 570 nm gemessen. Anhand
einer aus den Standards erstellten linearen Regression wurden danach die
Probenwerte berechnet.
3.2.18 Anfertigung histologischer Präparate
Die histologischen Gewebeschnitte wurden am ZTL von Frau Wiebe angefertigt und
mit Hämatoxilyn-Eosin (HE) gefärbt. Die Organe wurden über Nacht paraffinisiert und
anschließend per Hand eingebettet. Nach Einbettung wurden 3 µm dicke Längs-
schnitte gemacht und im Färbeautomaten HE gefärbt. Die gefärbten Schnitte wurden
anschließend mit Korbitbalsam eingedeckt.

Eigene Untersuchungen
51
3.2.19 Histologische Auswertung der Präparate
Die Schnitte wurden ohne vorher bekannte Zuordnung beurteilt. Zur Beurteilung der
Entzündungsherde in Zäkum und Kolon kam der The Jackson Laboratory-Schlüssel
zur Anwendung (BLEICH et al. 2004).
Dieser gliedert sich in:
Schweregrad
Der Schweregrad der Entzündung wurde beurteilt als: 0 = keine Entzündung;
1 = mild; 2 = mäßig; 3 = schwer. Milde Läsionen waren kleine, fokale oder weiter
voneinander entfernte multifokale Areale, in denen das Entzündungsgeschehen auf
die Lamina propria beschränkt blieb. Bei mäßigen Veränderungen handelte es sich
entweder um multifokale oder lokal extensive Entzündungsgebiete, in denen das
Entzündungsgeschehen bis in die Submukosa fortschritt. Schwere Läsionen waren
Ulzerationen größeren Ausmaßes (>1 mm der Mukosa betroffen).
Hyperplasie
Der Grad der Hyperplasie wurde beurteilt als: 0 = keine Hyperplasie; 1 = mild;
2 = mäßig; 3 = schwer. Bei einer milden Hyperplasie erschien das Epithel
morphologisch normal als einfaches, hochprismatisches Schleimhautepithel, das
aber im Vergleich zur normalen Mukosa eine mindestens zweifache
Schleimhautdicke aufwies, was durch die Länge der Krypten angezeigt wurde. Eine
mäßige Hyperplasie war durch eine zwei bis dreifache Schleimhautdicke mit
hyperchromatischen Zellen, eine verringerte Anzahl an Becherzellen und vereinzelt
verlängerte und verzweigte Krypten charakterisiert. Bei einer schweren Hyperplasie
war das Epithel deutlich verdickt (vierfach oder mehr) und zeigte eine deutliche
Hyperchromasie der Zellen, sehr wenige bis keine Becherzellen, einen hohen
Mitoseindex der Zellen innerhalb der Krypten und viele Krypten mit verlängerter und
verzweigter Struktur.

Eigene Untersuchungen
52
Ulzeration
Ulzeration wurde beurteilt als: 0 = kein Ulkus; 1 = eine bis zwei Ulzerationen, die
insgesamt maximal 20 Krypten einbezogen; 2 = eine bis vier Ulzerationen, an denen
insgesamt 20-40 Krypten beteiligt waren; 3 = jegliche Ulzeration, die über die vorher
genannten Kriterien hinaus gingen.
Fläche
Der Anteil der entzündlich veränderten Fläche wurde in 10%-Schritten auf einer
Skala von 0-100% geschätzt, bezogen auf die gesamte Fläche des jeweiligen
Darmabschnitts. Für die Analysen wurden die Prozentzahlen folgendermaßen
umgerechnet: 0 = 0%; 1 = 10-30%; 2 = 40-70%; 3 = >70%
Aus den Werten für Schweregrad, Hyperplasie, Ulzeration und Fläche wurde die
Summe gebildet und als Gesamtscore angegeben.
3.2.20 Statistische Auswertung
Die statistische Auswertung der INFγ-Bestimmung wurde mit dem Programm
GraphPad Prism auf einem PC durchgeführt. Das Programm StatView wurde für die
statistische Auswertung des MNV-ELISA auf einem Apple Powerbook G4 genutzt.
Für weitere graphische Darstellungen und Berechnungen wurde Microsoft Excel
2007 verwendet.

Ergebnisse
53
4 Ergebnisse
4.1 Anzucht von MNV in der Zellkultur
Eine Anzucht und Vermehrung von MNV war problemlos möglich. RAW-Zellen, die in
einer gleichmäßigen Zellschicht wuchsen, reagierten mit einem deutlichen CPE.
Dieser trat je nach Infektionsdosis innerhalb weniger Stunden bis zu 1,5 Tagen auf.
Die Zellen kugelten sich ab und lösten sich aus dem Zellverband von der
Flaschenoberfläche. Der CPE erfasste vollständig alle Zellen.
4.2 Virus-Nachweis mittels PCR
Zu Beginn wurden die Primerpaare 5205f/5388r und 5004f/5219r erprobt. Die
isolierte RNA wurde mit dem Reverse-Primer des jeweiligen Paares zur cDNA
umgeschrieben. Hierbei erwies sich das Paar 5004f/5219r als geeignet.
Bei den PCR-Untersuchungen des Infektionsversuches kamen Zweifel auf, ob das
Primerpaar 5004f/5219r MNV in Organproben detektieren kann. Das MNV-Isolat
wurde aus einer Milz isoliert (siehe 3.2.2 Virusisolation). Alle im Infektionsversuch
gewonnenen Milzen wurden jedoch negativ getestet. Es erfolgte eine Entwicklung
von weiteren Primerpaaren unter Berücksichtigung der Sequenzierungsergebnisse.
Zur Austestung der Primerpaare wurde RNA verwandt, die aus Zellkulturüberstand,
sowie aus Organ- und Kotproben des Infektionsversuchs isoliert worden war. Die
RNA wurde mit dem OligoDT-Primer zu cDNA umgeschrieben, um mit der gleichen
cDNA auch die RPS9-PCR durchführen zu können (siehe Abbildung 6). Letztlich
zeigte sich, dass das Primerpaar 5004f/5219r weiterhin die besten Resultate brachte,
da eine deutliche Bande auszumachen war. MNV war bei den in den
Infektionsversuchen verwendeten Mausstämmen in keiner Milz nachweisbar (siehe
Tabelle 20 S. 78).

Ergebnisse
54
Abbildung 5: Vergleich verschiedener Primerpaare im Gradienten.
Die PCR in Abbildung 5 wurde mit einer cDNA-Positivkontrolle (gewonnen aus MNV-
positivem Zellkulturüberstand) bei 4 unterschiedlichen Annealingtemperaturen (53°
C, 55,6° C, 58,4° C und 62° C) durchgeführt. Die Primerpaare 5580f/5671r und
5205f/5469r wiesen besonders kräftige Banden auf.
Abbildung 6: Vergleich der Primerpaare 5580f/5671r und 5205f/5469r mit 5
verschiedenen Proben, die aus Organen und Kot isoliert wurden.

Ergebnisse
55
Die PCR in Abbildung 6 wurde mit 3 MNV-positiven cDNA-Proben (gewonnen aus 2
MLK und einer Kotprobe), 2 MNV-negativen cDNA-Proben (gewonnen aus 2 Milzen)
sowie Negativ- und Positiv-Kontrolle durchgeführt. Die 3 Banden der MNV-positiven
Proben in der PCR mit dem Primerpaar 5580f/5671r sind schwer zu unterscheiden
von den negativen Proben. Dies wird beim Primerpaar 5205f/5469r aufgrund eines
größeren Amplikons (265bp) wesentlich erleichtert. Die RPS9-PCR wurde mit der
cDNA, die aus den 2 MNV-negativen Milzen isoliert worden war, sowie Negativ- und
Positiv-Kontrolle durchgeführt.
Abbildung 7: Vergleich der Primerpaare 5205f/5469r und 5004f/5219r.
Das Gel in Abbildung 7 zeigt 8 MNV-positive Kotproben aus einer PCR mit dem
Primerpaar 5205f/5469r, dann eine Lücke gefolgt von Positiv- und Negativkontrolle.
Nach der Basenleiter sind die gleichen 8 MNV-positiven Kotproben mit Positiv- und
Negativkontrolle mit dem Primerpaar 5004f/5219r aufgetragen. Das Primerpaar
5205f/5469r weist bei allen Proben, die aus Kot isoliert wurden, Doppelbanden auf.
Nur die Positivkontrolle, die aus MNV-positivem Zellkulturüberstand isoliert wurde,
hat eine Bande. Bei Probe 3 ist nur eine schwache Bande zu erkennen, Probe 5
zeigt keine Bande. Das Primerpaar 5004f/5219r hingegen zeigt bei jeder Probe eine
starke Bande.

Ergebnisse
56
4.3 Sequenzierung des Virus
Als Template wurde die mit dem OligoDT-Primer umgeschriebene cDNA verwendet.
Um eine reproduzierbare Sequenzierung zu ermöglichen, wurden die RNA-
Abschnitte als cDNA in Bakterienplasmide kloniert. Die Plasmid-DNA wurde per PCR
vervielfältigt, anschließend über ein Gel laufen gelassen und dann ausgeschnitten
und zur externen Sequenzierung verschickt. Die vollständige Sequenz wurde an die
NCBI-Nukleotiddatenbank übermittelt. Sie wurde unter der Zugangsnummer
EU854589 registriert. Das Isolat bekam die Bezeichnung Murine
norovirus/Hannover1/2007/DEU. Der Stammbaum in Abbildung 8 zeigt die
Verwandtschaft des Isolats mit anderen murinen Noroviren. Er wurde mit BLAST
erstellt (ALTSCHUL et al. 1990).

57
Abbildung 8: Verwandschaft vom isolierten MNV-Stamm Hannover1/2007/DEU erstellt durch BLAST (ALTSCHUL et al. 1990).

Ergebnisse
58
4.4 Virusquantifizierung
Im Plaque-Test trat ein CPE auf. Der CPE war bereits an der Verfärbung des
Mediums (Indikator Phenolrot gelb bei pH 0,9-6,4, rot bei pH >6,4) zu erkennen. Bei
den lebenden Kontrollen war das Medium gelb, bei den toten Zellen rosarot. Die
Einfärbung der Zellen mit Kristallviolett zeigte jedoch keine zählbaren Löcher im
Zellrasen, sondern eine schlierige Veränderung des Zellrasens. Dies wird für einige
MNV-Isolate beschrieben (THACKRAY et al. 2007). Daraufhin wurde ein TCID50-Test
durchgeführt. Auch hier zeigte bereits der dem Zellmedium zugesetzte Indikator den
CPE an (Abbildung 9). Abschließend wurde mit Kristallviolett angefärbt.
Abbildung 9: TCID50-Test; der Indikator des Mediums zeigt lebende Zellen (gelb) an;
zweites Bild nach der Anfärbung mit Kristallviolett (lebende Zellen stark lila).
Nach Auszählung der Vertiefungen mit lebenden/toten Zellen und Auswertung nach
Spearmann und Kärber (siehe S. 38) ergab sich für die konfektionierte Viruscharge
eine TCID50 von 2,5 x 108 TCID50/ml. Diese Charge wurde für die Infektionsversuche
verwendet.
4.5 Indirekte Antikörpernachweisverfahren
4.5.1 IFA
Die Untersuchung von Mäuseseren auf Antikörper gegen MNV per IFA wurde bereits
im Labor der Firma Biodoc etabliert und als Goldstandard für die Etablierung des
ELISA sowie zur Untersuchung aller Tiere des Infektionsversuchs genutzt.

Ergebnisse
59
Bei der Herstellung der Objektträger ist eine maximal 25% Infektionsrate des
Zellrasens angestrebt. Abbildung 10 zeigt ein MNV-positives Mausserum in der
Immunfluoreszenz. Die infizierten Zellen zeigen eine cytoplasmatische Fluoreszenz
im Randbereich mit einem „Polkappenphänomen“. Als Bewertung wurde ein
semiquantitativer Schlüssel angewendet. Dazu wurde die Serumprobe mit einem
deutlich positiven Kontrollserum verglichen.
Kennzeichnung Serumreaktion beurteilt anhand der Intensität
+++ Starke Reaktion
++ Mittelstarke Reaktion
+ Schwach positive Reaktion
- Keine Reaktion
+/- Grenzwertige Reaktion
NSR Nicht spezifische Reaktion
Abbildung 10: MNV-positives Mausserum (+++) in der Immunfluoreszenz. MNV-
infizierte Zellen sind grüngelblich markiert.

Ergebnisse
60
4.5.2 Antigenaufreinigung und ELISA
Die Aufreinigung per Ultrazentrifuge erfolgte angelehnt an die Publikation von Wobus
et al. (WOBUS et al. 2004). Ziel war es, MNV-Antigen für einen ELISA und
elektronenmikroskopische Aufnahmen zu gewinnen.
Nach der ersten Kissenzentrifugation war ein Pellet am Boden des
Ultrazentrifugenröhrchens auszumachen, welches in PBSM gelöst wurde.
Probeweise wurden bereits mit dem gelösten Pellet ELISA-Platten beschichtet und
mit diversen Mäuseseren, die in der IFA postiv bzw. negativ für MNV reagiert hatten,
erprobt. Hierbei zeigte sich eine Substratumsetzung bei IFA positiven Seren,
während die meisten IFA negativen Seren nicht reagierten. Die Ausnahmen ließen
eine Verunreinigung vermuten. Daher wurde mit dem gelösten Pellet eine
Aufreinigung über einen Cäsiumchloridgradienten durchgeführt. Am Ende der
Zentrifugation war eine deutliche Bande im Ultrazentrifugenröhrchen zu erkennen,
die abgenommen wurde (Abbildung 11).
Abbildung 11: Kissenzentrifugation; Bande nach der Gradientenzentrifugation.
Eine Dialyse zur Entfernung des Cäsiumchlorids erfolgte nicht, da beim Coating der
ELISA-Platten hoch verdünnt wurde. Doch auch hier reagierten einige negative
Seren. Dies sprach für eine immer noch vorhandene Verunreinigung der Virusbande.
Da es sich bei der RAW-Zelllinie um Mäusezellen handelt, kam die Vermutung auf,
dass die falsch positiven Seren mit RAW-Zellresten reagiert hatten, die trotz

Ergebnisse
61
Aufreinigung durch Zentrifugation und Ultrazentrifugation noch vorhanden waren. Um
dies zu überprüfen wurden RAW-Zellen angezüchtet und nicht infiziert. Die weitere
Behandlung und Aufreinigung entsprach jedoch der Behandlung infizierter RAW-
Zellen. Nach der Kissenzentrifugation war ebenfalls ein Pellet am Boden
auszumachen. Dieses wurde ebenfalls in PBSM gelöst und an ELISA-Platten
gebunden. Hier reagierten die IFA-negativen Seren teilweise ebenfalls mit einer
Substratumsetzung, dies bestätigte die Vermutung, dass einige Mäuse Antikörper
gegen RAW-Zellbestandteile besitzen. Um die Mäuseseren zu erkennen, die mit
RAW-Zellbestandteilen reagieren, wurde entschieden, eine parallele Zellkontrolle
beim ELISA durchzuführen.
Damit die OD-Werte von MNV-Reaktionsvertiefungen, in denen vermutlich auch
noch RAW-Bestandteile gebunden waren, vergleichbar zu Zellkontroll-
Reaktionsvertiefungen waren, wurden gleichzeitig zur MNV-Anzucht RAW-Zellen der
gleichen Passage ohne Infektion angezüchtet. Sobald die zur Infektion vorgesehene
Hälfte der Zellkulturflaschen infiziert wurde, wurde die andere Hälfte (Kontrollcharge)
eingefroren. Beim Coating der ELISA-Platten wurde für MNV-Antigen und RAW-
Antigen die gleiche Verdünnung mit dem Coatingbuffer gewählt.
Zur Einschätzung, ob ein Serum Antikörper gegen MNV enthält, wurde der korrigierte
OD-Wert eingeführt. Dieser ergibt sich aus der Differenz des OD-Werts der MNV-
Antigen-beschichteten Kavität zur RAW-Antigen-beschichteten Kavität. Zur
Bestimmung der Grenzwerte für die Serumbeurteilung und zur Überprüfung der
Robustheit des ELISA wurde eine Validierung durchgeführt. Die einzelnen
Messdaten sind im Anhang zu finden.

Ergebnisse
62
4.5.2.1 ELISA-Validierung
Zuerst wurde ein Positivkontroll- und ein Negativkontroll-Pool aus eindeutig in der
IFA beprobten Seren zusammengestellt. Jeder Pool wurde gründlich durchmischt,
sterilfiltriert und in kleinen Chargen bei -20° C bis zur Verwendung gelagert. Erst eine
große Menge Serum erlaubte eine Validierung in entsprechender Proben-Anzahl.
Außerdem kann anhand dieser Pools eine Orientierung bei der Herstellung von
neuen Chargen des ELISA stattfinden, um vergleichbare OD-Werte zu erhalten.
Beim alltäglichen Gebrauch des ELISA können die Pools genutzt werden, um
Abweichungen beim Anwendungsprotokoll sowie Qualitätsmängel des antigen-
beschichteten Materials aufzuzeigen.
Durch die Verwendung einer Differenz für den korrigierten OD-Wert wurden
korrigierte OD-Werte kleiner null für die Berechnung von Mittelwert und
Standardabweichung gleich null gesetzt und auf die Bildung eines
Variationskoeffizienten verzichtet.
Für die Validierung des ELISA wurden folgenden Messungen durchgeführt:
4.5.2.2 Intraserielle Präzision des Positivkontroll- und des Negativkontroll-Pools
Der Positivkontroll-Pool wurde in einem Reaktionsansatz zehnmal untersucht, der
Negativkontroll-Pool im gleichen Ansatz sechsmal. Hierbei ergab sich aus den
korrigierten OD-Werten des Positivkontroll-Pools ein Mittelwert von 2,76 bei einer
Standardabweichung von 0,07. Der Variationskoeffizient betrug 2,59%. Für den
Negativ-Kontroll-Pool betrug der Mittelwert 0,11 bei einer Standardabweichung von
0,04.
4.5.2.3 Interserielle Präzision des Positivkontroll- und des Negativkontroll-Pools
Der Positivkontroll- als auch der Negativkontroll-Pool wurden an zehn verschiedenen
Tagen als Einzelprobe getestet. Hierbei ergab sich aus den korrigierten OD-Werten
des Positivkontroll-Pools ein Mittelwert von 2,82 bei einer Standardabweichung von
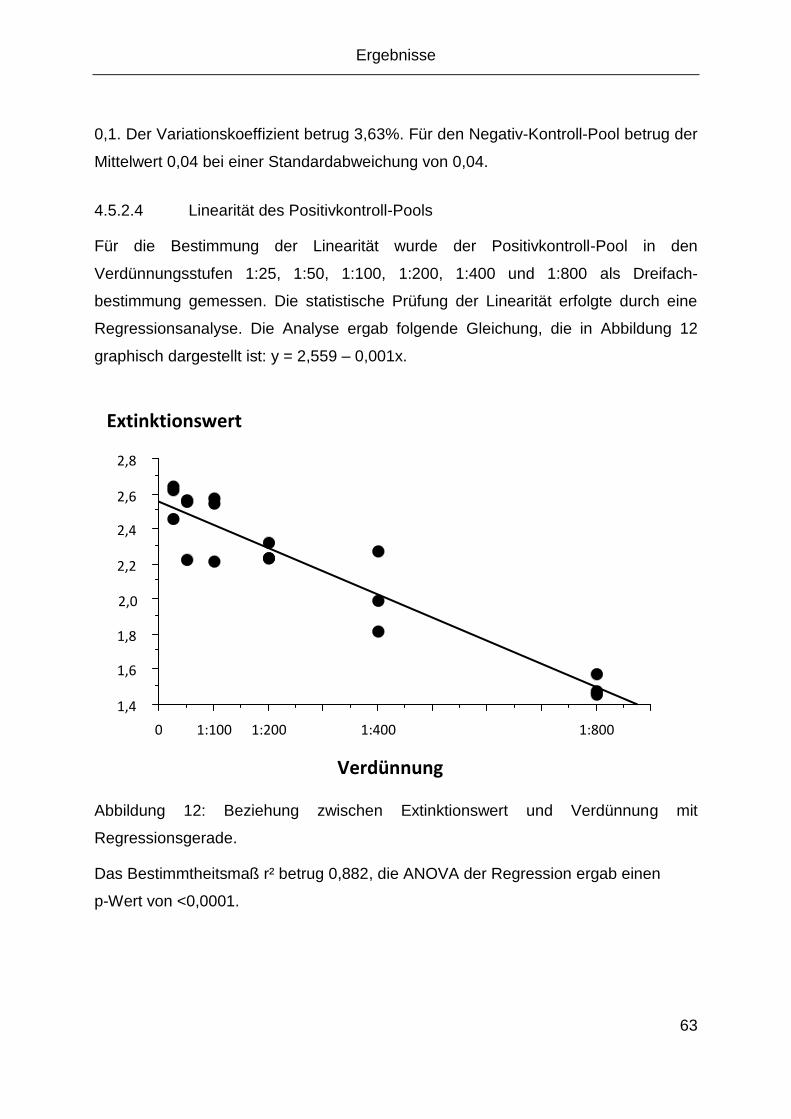
Ergebnisse
63
0,1. Der Variationskoeffizient betrug 3,63%. Für den Negativ-Kontroll-Pool betrug der
Mittelwert 0,04 bei einer Standardabweichung von 0,04.
4.5.2.4 Linearität des Positivkontroll-Pools
Für die Bestimmung der Linearität wurde der Positivkontroll-Pool in den
Verdünnungsstufen 1:25, 1:50, 1:100, 1:200, 1:400 und 1:800 als Dreifach-
bestimmung gemessen. Die statistische Prüfung der Linearität erfolgte durch eine
Regressionsanalyse. Die Analyse ergab folgende Gleichung, die in Abbildung 12
graphisch dargestellt ist: y = 2,559 – 0,001x.
Abbildung 12: Beziehung zwischen Extinktionswert und Verdünnung mit
Regressionsgerade.
Das Bestimmtheitsmaß r² betrug 0,882, die ANOVA der Regression ergab einen
p-Wert von <0,0001.
1,4
1,6
1,8
2,2
2,4
2,6
2,8
0 1:100 1:200 1:400 1:800
2,0
Verdünnung
Extinktionswert

Ergebnisse
64
4.5.2.5 Einzelmessungen an IFA positiven und negativen Seren
Um den ELISA als Testverfahren mit der IFA zu vergleichen, wurden 68 negative und
76 positive Seren in einem Versuchsansatz mit dem ELISA getestet. Hierbei ergab
sich eine Verteilung der Messwerte, die in Abbildung 13 zu sehen ist.
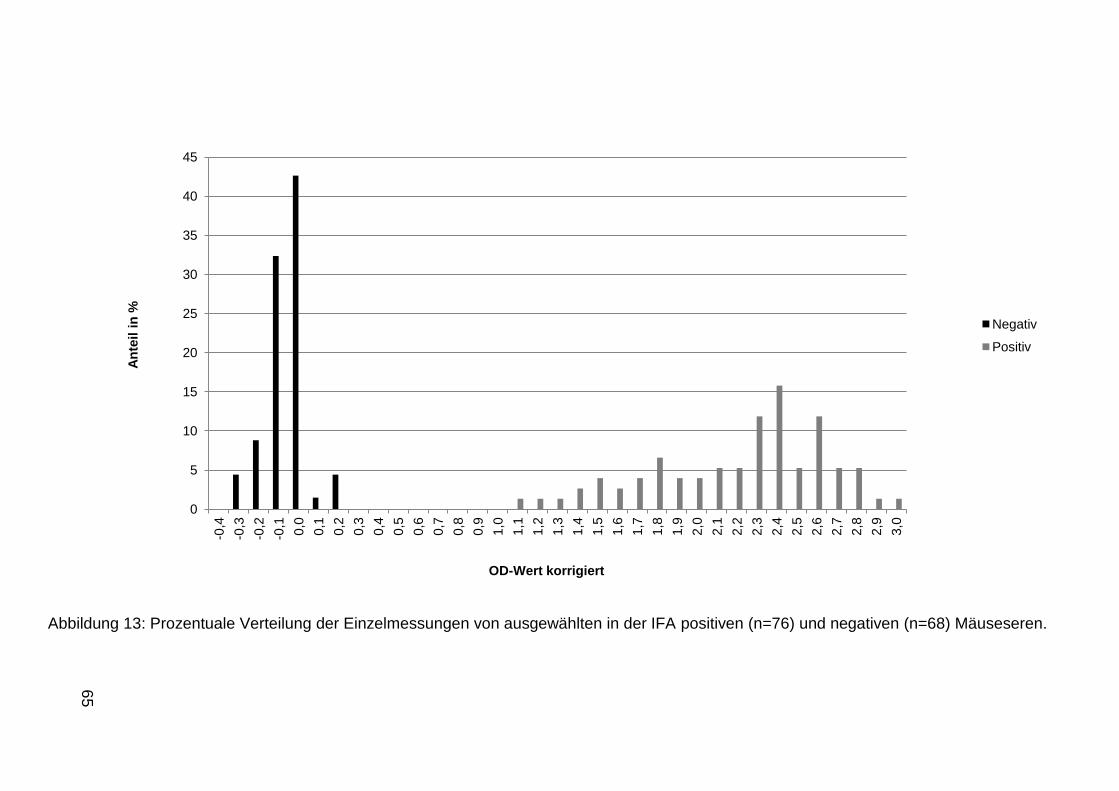
65
Abbildung 13: Prozentuale Verteilung der Einzelmessungen von ausgewählten in der IFA positiven (n=76) und negativen (n=68) Mäuseseren.
0
5
10
15
20
25
30
35
40
45
-0,4
-0,3
-0,2
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0
2,1
2,2
2,3
2,4
2,5
2,6
2,7
2,8
2,9
3,0
An
teil i
n %
OD-Wert korrigiert
Negativ
Positiv

Ergebnisse
66
Zusätzlich wurden anschließend weitere 205 Seren, die in der IFA negativ für MNV-
Antikörper beurteilt worden waren, untersucht. Hierbei ergab sich ein Spektrum für
den korrigierten OD-Wert eines MNV-negativen Serums von -0,385 bis 1,4. Die
Verteilung ist in Abbildung 14 dargestellt.
Abbildung 14: Prozentuale Verteilung der Einzel-OD-Wertbestimmung von 205 in der
IFA negativen Mäuseseren mit Hilfe des ELISA.
Anhand der Verteilung wurden folgende Grenzwerte für die hergestellte Charge
festgelegt, um eine Bewertung von Testseren zu ermöglichen:
OD korrigiert < 0 bis 1 negativ
OD korrigiert > 1 bis 1,5 fraglich
OD korrigiert > 1,5 positiv
4.6 Elektronenmikroskopische Aufnahmen
Die Aufreinigung über den CsCl-Gradienten führte nicht zu auswertbaren Grids
(Objektträger für elektronenmikroskopische Aufnahmen). Daher erfolgte wie
beschrieben eine Aufreinigung durch wiederholte Pelletierung. Verschiedene
Aufnahmen sind in Abbildung 15 zu sehen. Das Virus ist ca. 35 nm groß. In
Abbildung 15a ist die Kelchstruktur der Oberfläche auszumachen, Pfeile weisen auf
Virionen hin.
0
5
10
15
20
25
-0,4 -0,3 -0,2 -0,1 0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,9 1 1,1 1,4
An
teil i
n %
OD-Wert korrigiert
Negativ

Ergebnisse
67
Abbildung 15: Elektronenmikroskopische Aufnahmen von MNV-Virionen
(durch schwarze Pfeile gekennzeichnet).
a b
d c

Ergebnisse
68
4.7 Serologische Untersuchung von Anzeigertieren des zentralen
Tierlaboratoriums auf MNV
Um einen Überblick über die Verbreitung von MNV im ZTL zu erhalten, wurden
Seren, die im Rahmen der Jahresuntersuchung 2008 von Anzeigertieren gewonnen
worden waren, zusätzlich mittels IFA auf gegen MNV gerichtete Antikörper
untersucht. Die Ergebnisse sind in Tabelle 12 zusammengefasst.
Tabelle 12: Überblick über die Verbreitung von MNV im ZTL im Jahr 2008
Haltungssystem Anzahl Tiere
positiv/getestet
Prävalenzrate (%)
Isolator 0/13 0
SPF-Zuchtbarriere 0/6 0
Experimentelle Haltungsbarriere* 121/205 59
Vergleich von zwei Tierräumen der experimentellen Haltungsbarriere,
in denen alle Mäuse getestet wurden
Raum mit offenen Käfigen 174/200 87
Raum mit Filterdeckelkäfigen 104/155 67
*enthält eine Einheit mit einzeln belüfteten Käfigsystemen - individually ventilated
cages (IVC) - mit 22/40 (55%) positiven Anzeiger-Mäusen
4.8 Infekionsversuche
1. Tierversuch
Im ersten Infektionsversuch wurden C3-Il10-/-- und B6-Il-10-/--Mäuse beider
Geschlechter infiziert. Um einen Überblick über den bisher unbekannten
Infektionsverlauf zu bekommen, wurden Sektionen nach zwei, vier und acht Wochen
durchgeführt.

Ergebnisse
69
2. Tierversuch
Im zweiten Infektionsversuch wurden C3-Il10-/-- und B6-Il-10-/--Mäuse sowie der
jeweilige Wildtyp infiziert. Folgernd aus den Daten der INFγ-Messungen des ersten
Versuchs wurden zwei Sektionstermine (zwei und vier Wochen nach Infektion)
festgelegt.
3. Tierversuch
Im dritten Infektionsversuch wurden Mäuse infiziert, um Daten des zweiten Versuchs
zu ergänzen. Außerdem wurden männliche B6-Il-10-/- - Mäuse sowie männliche und
weibliche Mäuse des zugehörigen Wildtyps infiziert und nach einer Woche seziert,
um Erkenntnisse über den Infektionsstatus (vor allem die Stimulation der
Lymphozyten) der Tiere eine Woche nach der Infektion zu bekommen. Eine
zusätzliche Tiergruppe B6-wt Weibchen wurde infiziert und nach 3, 6, 9 und 12
Tagen seziert. Durch diese Gruppe sollte Aufschluss darüber geschaffen werden, wo
und ab wann das Virus per PCR nachzuweisen ist. Daher wurde der
Mesenteriallymphknoten der Tiere für den PCR-Nachweis verwandt.
4. Tierversuch
Im vierten Infektionsversuch wurden keimfreie männliche und weibliche C3-Il10-/-
Mäuse, die aus einem Isolator des ZTL stammten, mit MNV infiziert und vier Wochen
bis zur Sektion in einem Mikroisolator gehalten. Dieser Versuch sollte zeigen, welche
Einflüsse MNV auf Tiere ohne jegliche natürliche mikrobiologische Flora hat.
5. Tierversuch
In fünften Versuch wurden C3-Il10-/-- und B6-Il-10-/--Mäuse zuerst mit MNV infiziert.
Zwei Wochen später erfolgte eine zusätzliche Infektion mit H. hepaticus. Die
Kontrolltiere wurden nur mit H. hepaticus infiziert. Mit diesem Versuch sollte ein
möglicher Einfluss der MNV-Infektion auf die spätere H. hepaticus-Infektion
untersucht werden. Die Sektion erfolgte dann sechs Wochen nach Infektion mit H.
hepaticus.

Ergebnisse
70
4.8.1 Klinik
In Tierversuch 1 bis 4 zeigten die mit MNV infizierten Mäuse bis zum Versuchsende
ein ungestörtes Allgemeinbefinden. Zu den Kontrolltieren war kein Unterschied
festzustellen. In Tierversuch 5 zeigten die Tiere bis 40 Tage nach Infektion mit H.
hepaticus keine Auffälligkeiten. Erst am Sektionstermin war sowohl bei der
Doppelinfektionsgruppe als auch bei der Kontrollgruppe ein leicht aufgekrümmter
Rücken festzustellen. Die Tiere fraßen und tranken noch.
4.8.2 Sektion
Die Sektion der Tiere aus Tierversuch 1 bis 4 ergab keinerlei makroskopische
Hinweise auf ein Infektionsgeschehen. Es bestand kein feststellbarer Unterschied zu
den Kontrolltieren. In Tierversuch 5 waren sowohl bei der Doppelinfektions- als auch
bei der Kontrollgruppe makroskopische Veränderungen feststellbar. Es handelte sich
hierbei um ein geschwollenes Kolon.
4.8.3 Serologie
Die Untersuchung der gewonnenen Seren zeigte in der Immunfluoreszenz bereits
am Tag 7 nach Infektion (Tierversuch 3) eine positive Reaktion. Ab Tag 9 p.i. lag die
Serokonversion aller mit MNV infizierten Tiere (Tierversuch 1 bis 5) bei 100 %. Die
Kontrolltiere hatten keine nachweisbaren Antikörper gegen MNV. Dies galt auch für
die Kontrolltiere, die in einem separaten Filterdeckelkäfig im Käfigregalschrank mit
den infizierten Tieren gehalten wurden.

Ergebnisse
71
4.8.4 Lymphozytenstimulation
4.8.4.1 Versuch 1
Um sicher zu gehen, dass genügend Lymphozyten für eine Stimulation aus den
Mesenteriallymphknoten der Tiere zu Verfügung standen, wurden im ersten Versuch
die Mesenteriallymphknoten nach Stamm und Geschlecht gepoolt. Die Lymphozyten
wurden sechsfach pro Pool aufgetragen. Die IFNγ-Messwerte des ersten
Infektionsversuchs sind in Abbildung 16 graphisch dargestellt. Hierbei zeigte sich,
dass B6-Il10-/--Mäuse im Allgemeinen geringere Werte in der Lymphozyten-
stimulation aufwiesen als C3-Il10-/--Mäuse; ein geschlechtlicher Unterschied
innerhalb der Stämme konnte nicht beobachtet werden. Acht Wochen nach Infektion
war kein weiterer Anstieg von IFNγ festzustellen. Die Lymphozytenstimulation der
weiblichen Kontrolltiere (C3-Il10-/-) war nicht effektiv.
Die Präparation eines einzelnen MLK lieferte genügend Lymphozyten für den
Versuchsaufbau. Um mehr verwertbare Daten bezüglich der Lymphozyten-
Stimulation zu erhalten, wurden ab dem 2. Tierversuch Einzelpräparationen der
Mesenteriallymphknoten pro Tier angefertigt, die dreifach aufgetragen wurden.
Tabelle 13: Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 1 mit Hilfe einer einfaktoriellen ANOVA für die 4 Gruppen:
Kontrolle, 2 Wochen p.i., 4 Wochen p.i., 8 Wochen p.i.
Maus-Stamm B6-Il10-/- w B6-Il10-/- m C3-Il10-/- w C3-Il10-/- m
S; p<0,0001 S; p<0,0001 NS; p=07358 S; p<0,0001
S: Signifikant, p≤0,05; NS: Nicht signifikant, p>0,05
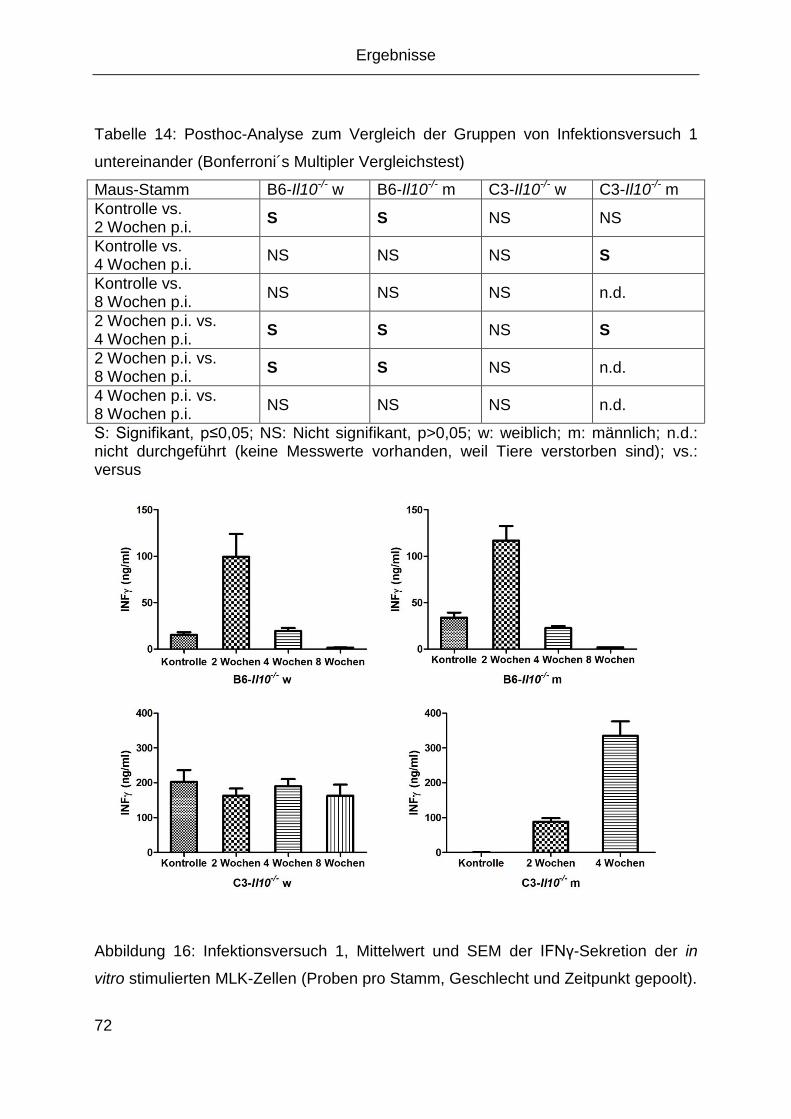
Ergebnisse
72
Tabelle 14: Posthoc-Analyse zum Vergleich der Gruppen von Infektionsversuch 1
untereinander (Bonferroni´s Multipler Vergleichstest)
Maus-Stamm B6-Il10-/- w B6-Il10-/- m C3-Il10-/- w C3-Il10-/- m
Kontrolle vs. 2 Wochen p.i.
S S NS NS
Kontrolle vs. 4 Wochen p.i.
NS NS NS S
Kontrolle vs. 8 Wochen p.i.
NS NS NS n.d.
2 Wochen p.i. vs. 4 Wochen p.i.
S S NS S
2 Wochen p.i. vs. 8 Wochen p.i.
S S NS n.d.
4 Wochen p.i. vs. 8 Wochen p.i.
NS NS NS n.d.
S: Signifikant, p≤0,05; NS: Nicht signifikant, p>0,05; w: weiblich; m: männlich; n.d.: nicht durchgeführt (keine Messwerte vorhanden, weil Tiere verstorben sind); vs.: versus
Abbildung 16: Infektionsversuch 1, Mittelwert und SEM der IFNγ-Sekretion der in
vitro stimulierten MLK-Zellen (Proben pro Stamm, Geschlecht und Zeitpunkt gepoolt).

Ergebnisse
73
4.8.4.2 Versuch 2
Die IFNγ-Sekretion der Darmlymphozyten aller im 2. Tierversuch mit MNV infizierten
Mäuse war zwei Wochen p.i. erhöht im Vergleich zur Kontrollgruppe (signifikant bei
B6-Il10-/- Mäusen und dem zugehörigen Wildtyp). Vier Wochen p.i. war die INFγ-
Sekretion im Vergleich zur Kontrollgruppe ebenfalls erhöht, signifikant jedoch nur bei
C3-Il10-/--Mäusen.
Tabelle 15: Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 2 mit Hilfe einer einfaktoriellen ANOVA für die 3 Gruppen:
Kontrolle, 2 Wochen p.i., 4 Wochen p.i.
Maus-Stamm B6-Il10-/- B6-wt C3-Il10-/- C3-wt
S; p=0,0094 S; p=0,0012 S; p=0,0221 NS; p=0,2042
S: Signifikant, p≤0,05; NS: Nicht signifikant, p>0,05
Tabelle 16: Posthoc-Analyse zum Vergleich der Gruppen von Infektionsversuch 2
untereinander (Bonferroni´s Multipler Vergleichstest)
Maus-Stamm B6-Il10-/- B6-wt C3-Il10-/- C3-wt
Kontrolle vs. 2 Wochen p.i. S S NS NS
Kontrolle vs. 4 Wochen p.i. NS NS S NS
2 Wochen vs. 4 Wochen p.i. NS S NS NS
S: Signifikant, p≤0,05; NS: Nicht signifikant, p>0,05; vs.: versus

Ergebnisse
74
Abbildung 17 Infektionsversuch 2, Mittelwert und SEM der INFγ-Sekretion der in vitro
stimulierten MLK-Zellen (Einzeltierproben, keine Unterscheidung nach Geschlecht)
Kontrolle 2 Wochen 4 Wochen0
1
2
3
4
5
B6-Il10-/-
INF (
ng
/ml)
Kontrolle 2 Wochen 4 Wochen0
1
2
3
4
5
B6-wt
INF (
ng
/ml)
Kontrolle 2 Wochen 4 Wochen0
5
10
15
20
25
C3-Il10-/-
INF (
ng
/ml)
Kontrolle 2 Wochen 4 Wochen0
5
10
15
20
25
C3-wt
INF (
ng
/ml)

Ergebnisse
75
4.8.4.3 Versuch 3
Die Werte der Lymphozytenstimulation sieben Tage nach der Infektion mit MNV
ergaben eine signifikante Erhöhung der INFγ-Sekretion für männliche B6-Il10-/-
Mäuse im Vergleich zur Kontrollgruppe. Auch beim Wildtyp sind die gemessenen
Werte erhöht, jedoch nicht signifikant.
Tabelle 17: Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 3 mit einem ungepaarten t-Test
Maus-Stamm B6-Il10-/- B6-wt
Kontrolle vs. 1 Woche p.i. S; p=0,0154 NS; p=0,1346
S: Signifikant, p≤0,05; NS: Nicht signifikant, p>0,05; vs.: versus
Abbildung 18: Infektionsversuch 3, Mittelwert und SEM der INFγ-Sekretion der in
vitro stimulierten MLK-Zellen (Einzeltierproben, keine Unterscheidung nach
Geschlecht).
Kontrolle 1 Woche Kontrolle 1 Woche0
1
2
3
4
5
B6-Il10-/-
B6-wt
INF (
ng
/ml)

Ergebnisse
76
4.8.4.4 Versuch 4
Die Auswertung der Lymphozytenstimulation aus dem 4. Tierversuch zeigte keine
signifikanten Unterschiede zwischen den keimfreien mit MNV infizierten C3-Il10-/-
Mäusen vier Wochen nach der Infektion und der zugehörigen Kontrollgruppe.
Tabelle 18: Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 4 mit einem ungepaarten t-Test
Maus-Stamm C3-Il10-/- (keimfrei)
Kontrolle vs. 4 Woche p.i. NS, p=0,8988
S: Signifikant, p≤0,05; NS: Nicht signifikant, p>0,05; vs.: versus
Abbildung 19: Infektionsversuch 4, Mittelwert und SEM der INFγ-Sekretion der in
vitro stimulierten MLK-Zellen (Einzeltierproben, keine Unterscheidung nach
Geschlecht).
Kontrolle 4 Wochen0
1
2
3
4
5
Keimfreie C3-Il10-/-
INF (
ng
/ml)

Ergebnisse
77
4.8.4.5 Versuch 5
Im 5. Tierversuch wurden keine Unterschiede in der INFγ-Sekretion der mit H.
hepaticus infizierten Kontrollgruppe zur Doppelinfektionsgruppe festgestellt.
Tabelle 19 Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 5 mit einem ungepaarten t-Test
Mausstamm C3-Il10-/- B6-Il10-/-
Kontrolle (Helico) vs. Doppelinfektion (MNV+Helico)
NS, p=0,5697 NS, p=0,5155
S: Signifikant, p≤0,05; NS: Nicht signifikant, p>0,05; vs.: versus; Helico: H. hepaticus
Abbildung 20: Infektionsversuch 5, Mittelwert und SEM der INFγ-Sekretion der in
vitro stimulierten MLK-Zellen (Einzeltierproben, keine Unterscheidung nach
Geschlecht).
Helico MNV+Helico Helico MNV+Helico0
10
20
30
40
50
60
C3-Il10-/-
B6-Il10-/-
INF (
ng
/ml)

Ergebnisse
78
4.8.5 PCR-Nachweis von MNV
Lunge, Leber, Nieren bis Tag 12 nach Infektion sowie Milzen bis Tag 28 nach
Infektion wurden per PCR negativ auf MNV getestet. Weitere Untersuchungen zu
späteren Zeitpunkten erfolgten nicht. Wie aus Tabelle 20 hervorgeht, wurde MNV im
Mesenteriallymphknoten, Darm sowie Kot bis zu acht Wochen nach Infektion
nachgewiesen. Die zugehörigen nicht infizierten Kontrolltiere wurden negativ auf
MNV getestet.
Tabelle 20: MNV-Nachweis via PCR (positiv getestet / insgesamt getestet)
Organe/Proben Tage p.i. Milz MLK Ileum Kolon Kot
3 0/4 4/4 Nicht getestet Nicht getestet 3/3
6 0/4 4/4 3/3 3/3 2/2
9 0/3 3/3 Nicht getestet Nicht getestet 3/3
12 0/3 2/2 Nicht getestet Nicht getestet 3/3
14 0/17 * Nicht getestet Nicht getestet 12/12
28 0/17 * 4/12 12/12 12/12
56 Nicht getestet * 4/8 4/8 4/8
*benutzt für Zytokin-Bestimmung

Ergebnisse
79
4.8.6 PCR-Nachweis von Helicobacter hepaticus
Zum Nachweis einer Infektion mit H. hepaticus wurden Kotproben der Tiere mittels
PCR untersucht. Die Ergebnisse sind Tabelle 21 zu entnehmen.
Tabelle 21: H. hepaticus-Nachweis via PCR (positiv getestet / insgesamt getestet)
positiv
Infektion mit MNV und H. hepaticus
12/12
Infektion mit H. hepaticus (Kontrollen)
9/11
Alle mit H. hepaticus infizierten Mäuse, auch die PCR-negativ getesteten Tiere,
zeigten in der Sektion typische pathologische Veränderungen, die für eine
Erkrankung der Tiere sprachen (siehe 4.8.2).
4.8.7 Histologie
Die infizierten Tiere aus Versuch 1 bis 4 zeigten kaum histologisch auswertbare
Unterschiede im Vergleich zur jeweiligen Kontrollgruppe. Nur bei mit MNV infizierten
C3-Il10-/--Mäusen waren besonders vier Wochen nach der Infektion (Tierversuch 2)
geringgradige fokale entzündliche Läsionen feststellbar (siehe Abbildung 22). In
Tierversuch 4 wurde bei den keimfreien mit MNV infizierten C3-Il10-/--Mäusen
histologisch nur die Anwesenheit einer höheren Anzahl mononukleärer Zellen im
Vergleich zur Kontrollgruppe festgestellt (siehe Abbildung 23). Die Auswertung von
Tierversuch 5 ergab sowohl bei der Doppelinfektions- als auch bei der Kontrollgruppe
eine hochgradige multifokale Dickdarmentzündung (siehe Abbildung 21) ohne
signifikante Unterschiede in der Score-Bewertung (siehe Tabelle 22). Es wurde
jedoch bei einigen Tieren der Doppelinfektionsgruppe ein etwas geringerer
histologischer Score-Wert ermittelt.

Ergebnisse
80
Tabelle 22: Statistische Auswertung der histologischen Scorewerte von
Infektionsversuch 5 mit einem ungepaarten t-Test
Mausstamm C3-Il10-/- B6-Il10-/-
Zäkum, Helico vs. MNV+Helico
NS; p=0,4105 NS; p=0,0977
Kolon, Helico vs. MNV+Helico
NS; p=0,1102 NS; p=0,3201
S: Signifikant, p≤0,05; NS: Nicht signifikant, p>0,05; vs.: versus; Helico: H. hepaticus
Abbildung 21: Infektionsversuch 5, Mittelwert und SEM des histologischen Score
(Jackson) von Zäkum und Kolon bei der Kontrollgruppe (Helico mit H. hepaticus
infiziert) und der Doppelinfektionsgruppe (MNV+Helico mit MNV und H. hepaticus
infiziert).
Helico MNV+Helico Helico MNV+Helico0
5
10
15
20
Zäkum
C3-Il10-/-
B6-Il10-/-
His
tolo
gis
ch
er
Sco
re
Helico MNV+Helico Helico MNV+Helico0
5
10
15
20
Kolon
C3-Il10-/-
B6-Il10-/-
His
tolo
gis
ch
er
Sco
re

81
Abbildung 22: Kolonabschnitt einer C3-Il10-/--Maus 4 Wochen p.i. mit MNV (A-C) und die zugehörige Kontrolle (D-F); HE-Färbung, der schwarze
Balken entspricht 100µm. Im Vergleich zur Kontrolle ist eine milde Hyperplasie des Epithels, ein gemischtzelliges Infiltrat in Mukosa und
Submukosa sowie ein geringgradiges Ödem festzustellen.
E
B
C
F
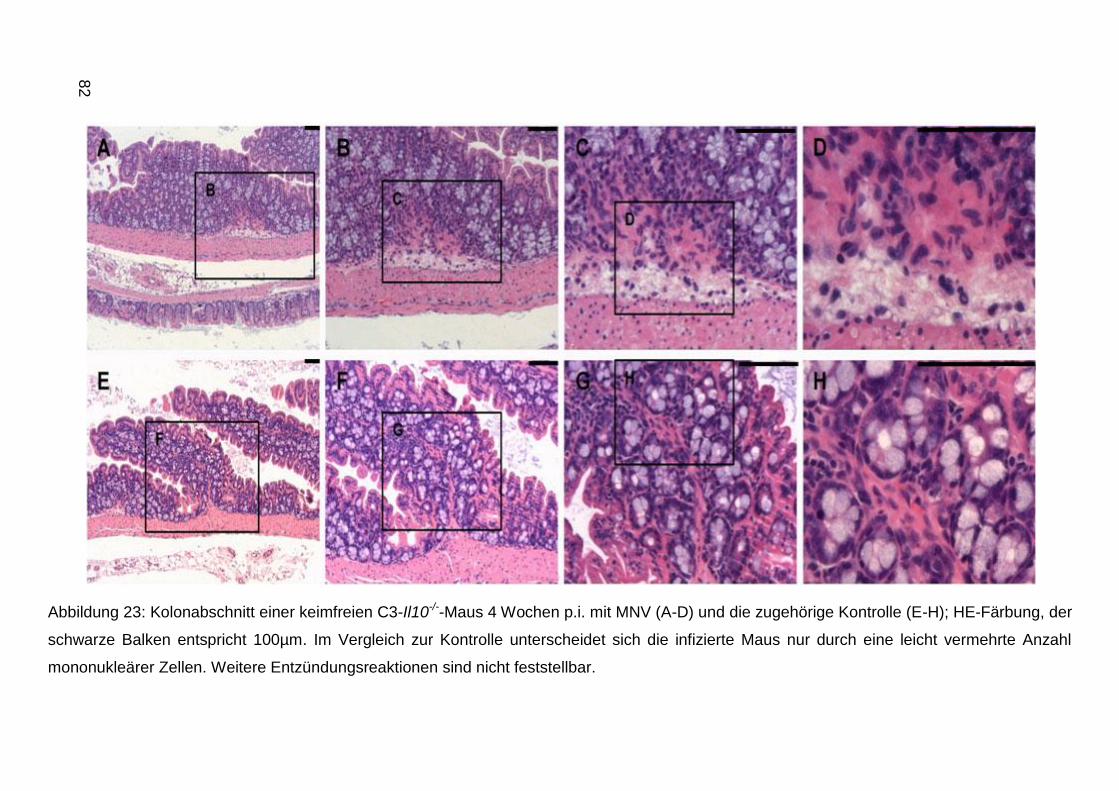
82
Abbildung 23: Kolonabschnitt einer keimfreien C3-Il10-/--Maus 4 Wochen p.i. mit MNV (A-D) und die zugehörige Kontrolle (E-H); HE-Färbung, der
schwarze Balken entspricht 100µm. Im Vergleich zur Kontrolle unterscheidet sich die infizierte Maus nur durch eine leicht vermehrte Anzahl
mononukleärer Zellen. Weitere Entzündungsreaktionen sind nicht feststellbar.

83
Abbildung 24: Kolonabschnitt einer B6-Il10-/--Maus infiziert mit MNV und H. hepaticus (A-D) und die zugehörige Kontrolle (E-H), nur infiziert mit
H. hepaticus; HE-Färbung, der schwarze Balken entspricht 100µm. Doppelinfektion und Kontrolle weisen eine vergleichbare mittel- bis
hochgradige transmurale, gemischtzellige Enteritis mit prominenter Hyperplasie der Enterozyten auf.

Diskussion
84
5 Diskussion
5.1 Zellkultur und Quantifizierung
Der erste Nachweis eines Norovirus bei der Maus wurde 2003 beschrieben (KARST
et al. 2003). Es konnte gezeigt werden, dass MNV-1 in Makrophagen und
dendritischen Zellen repliziert (WOBUS et al. 2004).
An der MHH wurde MNV aus Anzeiger-Mäusen isoliert, die Kontakt zur Einstreu von
Mäusen der Quarantäneabteilung des ZTL hatten. Da die Mäuse eine schwere
kombinierte Immundefizienz hatten, wurde eine mögliche aktive bzw. persistierende
Infektion vermutet. Spätere Studien von Goto et al. untermauern eine langfristige
MNV-Ausscheidung bei Prkdcscid-Mäusen nach einer Infektion mit MNV (GOTO et al.
2009). Klinische Symptome gab es nicht. Ein Antikörpernachweis war zu diesem
Zeitpunkt nicht etabliert. Außerdem war ein serologischer Nachweis aufgrund der
Immundefizienz der Anzeiger-Mäuse ungeeignet.
Mit Hilfe von RAW 264.7-Zellen gelang eine Isolierung und Vermehrung des MNV-
Stammes. MNV-typisch trat ein starker CPE auf. Um Infektionsversuche durchführen
zu können, musste das angezüchtete Virus quantifiziert werden. Ein zuerst
durchgeführter Plaque-Test zur Virusquantifizierung ließ sich nicht auswerten, da das
Virus keine auszählbaren Löcher im Zellrasen verursachte. Daraufhin wurde ein
TCID50-Test durchgeführt, der ausgewertet werden konnte. Ungefähr zeitgleich
wurden ebenfalls Isolate beschrieben, bei denen ein Plaque-Test nicht auswertbar
war (MÜLLER et al. 2007). Nur der TCID50-Test ist geeignet für eine
Virusquantifizierung des vorliegenden Isolats. Für einen Vergleich verschiedener
Isolate sollte daher der TCID50-Test angewandt werden.
5.2 Sequenzierung
Zur Sequenzierung des MNV-Isolats wurde unter anderem der Primer 15T-aTag
eingesetzt (MÜLLER et al. 2007). Aufgrund seines Aufbaus eignet sich dieser Primer

Diskussion
85
sehr gut, um unbekannte Noroviren zu sequenzieren. Er verfügt über einen zum
Poly-Adenin-Schwanz des Virus-Genoms komplementären Anteil von 15 Thymin
kombiniert mit einer variablen Base (V=A+C+G), sowie einer zusätzliche Sequenz,
die als spätere Primerbindungsstelle („Tag“) verwendet wurde.
Das komplette Genom wurde in der Nukleotiddatenbank des NCBI unter dem Namen
Murine norovirus/Hannover1/2007/DEU mit der accession number EU854589
veröffentlicht. Ein Vergleich mittels BLAST (ALTSCHUL et al. 1990) zeigte eine hohe
Übereinstimmung mit MNV-4 (accession number FJ446719). Die Kenntnis der
genetischen Sequenz des MNV-Isolats ermöglichte dann einen Abgleich der bereits
für die Detektion entwickelten Primer, um diese an die vorliegende Sequenz
anzupassen.
5.3 Nachweisverfahren
MNV-PCR
Um MNV isolieren zu können, musste eine direkte Detektionsmethode in Form der
PCR entwickelt werden. Da unterschiedliche MNV-Isolate voneinander stark
abweichende Sequenzen haben können, wurden zuerst anhand bereits
veröffentlichter MNV-Sequenzen Primer zur Detektion entwickelt, sowie Primer aus
anderen Veröffentlichungen erprobt. Das durch die eigenen Untersuchungen
bevorzugte Primerpaar 5004f/5219r ist ohne Abweichung in der Sequenz des Isolats
vorhanden. Im Vergleich zu anderen ausgetesteten Primerpaaren war hier zwar nur
eine mittelstarke Bande zu sehen, jedoch kam es bei aufgearbeiteten Organ- und
Kotproben nicht zu einer Mehrfachbandenbildung, die für einen PCR-Nachweis
unerwünscht ist (siehe Abbildung 7 S. 55).
Da das Isolat aus der Milz einer NOD-Prkdcscid-Maus gewonnen worden war, wurden
im Tierversuch auch die Milzen der infizierten Mäuse entnommen und per PCR auf
das Vorhandensein von MNV-RNA getestet. Hierbei stellte sich heraus, dass alle
Milzen negativ getestet wurden. Es konnte jedoch Erreger-RNA im
Gastrointestinaltrakt der Mäuse nachgewiesen werden. Dies bestätigt die

Diskussion
86
Untersuchungsergebnisse von Goto et al. (GOTO et al. 2009). Auch acht Wochen
nach der Infektion war im Kot der Mäuse noch MNV-RNA nachzuweisen, bei Paik et
al. wurde zehn Wochen p.i. MNV-RNA im Kot nachgewiesen (PAIK et al. 2010). Der
RNA-Nachweis lässt jedoch nicht unbedingt eine Aussage über die Infektiösität zu
(MÜLLER 2009).
Unsere Ergebnisse zeigen, dass sich Mäusekot gut für einen Erregernachweis
eignet. Die Mäuse müssen nicht getötet werden. Häufig setzen sie bei einer
Fixierung Kot ab. Dies bedeutet relativ wenig Stress im Vergleich zu einer
Blutentnahme für einen serologischen Nachweis. Bisher wurde aufgrund der
genetischen Variabilität bei murinen Noroviren allerdings kein Universalprimerpaar
zum Nachweis verschiedener bekannter muriner Norovirusstämme gefunden. Es
müssen also mehrere Primerpaare verwandt werden. Ein PCR-Nachweis ist daher
eher für die Überwachung eines Infektionsversuchs mit murinem Norovirus
bekannter Sequenz geeignet (HENDERSON 2008). Im Rahmen dieser Doktorarbeit
kam der PCR-Nachweis zum Einsatz, um die Virusverteilung im Tier und die Dauer
der Ausscheidung zu bestimmen. Das Zäkum wurde im Rahmen der Versuche
immer komplett für die Histologie entnommen. Es sollte bei zukünftigen Versuchen
beprobt werden, da Goto et al. Kot und Zäkum für den PCR-Nachweis von MNV
empfehlen (GOTO et al. 2009).
Serologische Nachweisverfahren
Für den Nachweis einer Infektion mit MNV wurde mit Hilfe des isolierten MNV-
Stammes im Rahmen der Dissertation ein serologisches Testverfahren in Form eines
ELISA entwickelt. Aufgrund der serologischen Kreuzreaktivität verschiedener MNV-
Isolate (HSU et al. 2006; THACKRAY et al. 2007) eignet sich der serologische Test
sehr gut für die Routine-Untersuchung auf eine Infektion mit MNV. Zu beachten ist
beim serologischen Nachweis, ob die Maus in der Lage ist, Antikörper zu bilden, und
ob die Infektion lange genug zurückliegt, dass bereits Antikörper gebildet wurden.
Humane Noroviren haben im Gegensatz zu MNV verschiedene Serotypen. Hinzu

Diskussion
87
kommt eine hohe Durchseuchung beim Menschen, was den serologischen Nachweis
in der Humandiagnostik weniger sinnvoll erscheinen lässt.
MNV-IFA (entwickelt durch die Firma Biodoc)
Die Herstellung der Objektträger, die mit MNV infizierten Zellen beschichtet sind,
erfolgte technisch angelehnt an Kraft und Meyer. (KRAFT u. MEYER 1990).
Dadurch, dass die Auswertung durch einen Untersucher am Mikroskop durchgeführt
wird, können hier Seren mit hohem unspezifischen Hintergrund erkannt werden.
Auch Seren mit niedrigem Titer können durch den Untersucher anhand der typischen
Markierung infizierter Zellen erkannt werden. Oft werden in Veröffentlichungen
positive Befunde aus ELISA und MFI mit der IFA bestätigt bzw. validiert (HSU et al.
2005).
MNV-ELISA
Der ELISA ermöglicht die automatisierte und quantitative Auswertung, da hier mit
einem Photometer der OD-Wert bestimmt wird. Anfangs wichen die Ergebnisse des
im Rahmen dieser Dissertation entwickelten ELISA teilweise von den Ergebnissen
der Immunfluoreszenz ab. Einige Mäuse, die eindeutig negativ in der
Immunfluoreszenz auf MNV getestet wurden, reagierten im MNV-ELISA positiv. Ein
kommerzieller rekombinanter ELISA, hergestellt zu wissenschaftlichen Zwecken
durch Charles River Laboratories, reagierte ebenfalls negativ. Dies sprach für eine
unspezifische Reaktion aufgrund von ungenügender Aufreinigung des Virusantigens.
Eine zusätzliche Aufreinigung über einen CsCl-Gradienten, wie sie in diversen
Veröffentlichungen beschrieben war (HSU et al. 2005), führte nicht zu besseren
Ergebnissen. Daraufhin wurde untersucht, ob die besagten Seren auch mit
Bestandteilen der für die Virusanzucht verwendeten RAW-Zellen reagierten, da es
sich bei der Zelllinie um Maus-Makrophagen handelt. Die Zellen wurden in der
Zellkultur angezüchtet und nicht infiziert. Es erfolgte der gleiche
Aufreinigungsprozess, der zur MNV-Antigenaufreinigung angewandt wurde.
Anschließend wurden Untersuchungskavitäten beschichtet. Wurden nun die IFA-
negativen Seren getestet, so reagierten einige Seren mit einer deutlichen Erhöhung

Diskussion
88
des OD-Werts. Die „falsch“ positiven Seren reagierten also auf einen Bestandteil der
Zellkultur. Andere Veröffentlichungen haben diese unerwünschte Reaktion bisher
nicht beschrieben.
Um falsch positive Seren zu erkennen, wurde daraufhin eine Zellkontrolle dauerhaft
eingeführt. Damit die Kontrollzellkultur vergleichbar zu den infizierten Zellkulturen
war, wurden beide Zellkulturen (nicht infiziert und infiziert mit MNV) parallel möglichst
gleich behandelt inklusive der Aufreinigung. Bewertet für die diagnostische Aussage
wurde die Differenz aus MNV-OD-Wert und RAW-OD-Wert. Durch Einführung des
korrigierten OD-Werts wurde eine hohe Übereinstimmung zu den Testergebnissen
der IFA erreicht. Um falsch positive Ergebnisse zu reduzieren, wurde auch ein
fraglicher Bereich definiert. Im Idealfall ergab der Differenzwert bei einem MNV-
negativen Serum den Wert 0, bei einem MNV-positiven Serum einen Wert >0, der
abhängig von der Verdünnung des Antigens bei der Beschichtung der ELISA-Platte
und dem Antikörpertiter des Serums war.
Um bei der Beschichtung für eine größtmögliche Konstanz zu sorgen, wurde ein
positiver Kontrollserumpool angelegt, aliquotiert und eingefroren. Bei der Erstellung
einer neuen Charge ELISA-Platten sollten ähnliche OD-Werte erreicht werden, wenn
der positive Kontrollserumpool aufgetragen wurde. Der positive Kontrollserumpool
wurde außerdem für die Validierung eingesetzt und als Positivkontrolle bei ELISA-
Tests mitgeführt. Bei korrigierten OD-Werten, die im fraglichen Bereich zwischen 1
bis 1,5 lagen, sollte der OD-Wert der Zellkontrolle getrennt beurteilt werden. Fand
hier eine starke Reaktion (hoher OD-Wert) statt, so konnte von einer unspezifischen
Reaktion ausgegangen werden, die durch nicht gegen MNV gerichtete Antikörper
hervorgerufen wurde. War der korrigierte OD-Wert im fraglichen Bereich, der OD-
Wert der Zellkontrolle jedoch niedrig, so konnte es sich um ein Serum mit
schwachem Antikörpertiter gegen MNV handeln. Es empfahl sich die erneute
Untersuchung des Serums mit einem anderen Testverfahren (z. B. IFA) oder eine
erneute Blutabnahme zur Serumgewinnung, um dieses dann mit dem ELISA zu
testen.

Diskussion
89
5.4 Prävalenz
Mit Hilfe der serologischen Testverfahren wurde das Vorkommen von MNV in
verschiedenen Hygieneeinheiten im ZTL untersucht. Hierbei zeigte sich in der
experimentellen Barriere eine hohe Prävalenz für MNV, die weltweit in vielen
Versuchstierhaltungen festgestellt wurde (MÄHLER u. KÖHL 2009; PRITCHETT-
CORNING et al. 2009). Dagegen waren Zuchttiere aus Isolatoren und SPF-Haltung
MNV-negativ. Die Art der Haltung der Mäuse hatte also einen großen Einfluss auf
das Vorhandensein von MNV-Antikörpern. Dieses Ergebnis lässt sich einerseits
durch eine bereits erfolgte Sanierung von anderen Erregern durch Embryotransfer
und Hysterektomie in der Vergangenheit, sowie andererseits durch die strengen
Hygiene-Auflagen erklären (HENDERSON 2008). In der experimentellen Haltung
sind die Hygiene-Auflagen für Pflegepersonal und Experimentatoren weniger streng
gefasst und überwacht. Hier werden nach Quarantänisierung und Freigabe durch
das mikrobiologische Labor des ZTL auch importierte Tiere gehalten. Die
Prävalenzdaten wurden erfasst, bevor MNV als Problem erkannt wurde.
Werden die Tiere ohne Filterdeckel gehalten, kann sich MNV über kontaminierte
Einstreu noch besser in einem Haltungsraum verbreiten. Dies lässt sich aus dem
Vergleich der Prävalenz der Haltung im offenen Käfig (87%) zur Haltung im
Filterdeckelkäfig (67%) schließen. Eine nochmals niedrige Prävalenz (55%) weist die
Haltung im IVC-Regal auf. Durch die getrennte Belüftung jedes einzelnen Käfigs
kann eine Übertragung über die Zuluft von Käfig zu Käfig verhindert werden. Gerät
der Erreger jedoch beim Experiment oder Umsetzen in den Käfig, so kann von einer
hohen Infektionsrate ausgegangen werden. Nach der Infektion im Tierversuch war
eine Serokonversion bei allen infizierten Tieren festzustellen.
Ein separater Filterdeckelkäfig mit nicht infizierten Kontrolltieren stand zusammen mit
den Filterdeckelkäfigen der infizierten Tiere im gleichen belüfteten Käfigschrank. Die
Haltung in einem Filterdeckelkäfig kombiniert mit der richtigen Reihenfolge der
Tierbetreuung (nicht infizierte Käfige zuerst versorgen) sowie dem Käfigwechsel
unter einer Sicherheitswerkbank haben bei oben genanntem Käfig mit Kontrolltieren

Diskussion
90
erfolgreich eine Kontamination mit MNV verhindert. Sowohl PCR- als auch Serologie-
Diagnostik für MNV waren bei diesen Mäusen negativ.
5.5 MNV im Tierversuch
Bei den immunkompetenten Mäusen, die im Rahmen dieser Dissertation untersucht
wurden, war die Infektion mit dem isolierten MNV-Stamm zu den untersuchten
Zeitpunkten auf den Magen-Darm-Trakt beschränkt. Die Infektion mit dem isolierten
MNV-Stamm führte zu einer verstärkten IFNγ-Sekretion der T-Zellen der infizierten
Mäuse (SPF-Haltung), wenn diese in vitro stimuliert wurden, auch im subakuten
Infektionsstadium 4 Wochen p.i. Die histologische Auswertung zeigte eine
geringfügige Erhöhung des Histologie-Scorewerts auf. So wurde bei einigen C3-
Il10-/--Mäusen 4 Wochen nach der Infektion eine milde Hyperplasie des Epithels, ein
gemischtzelliges Entzündungszell-Infiltrat in Mukosa und Submukosa sowie ein
geringgradiges Ödem festgestellt. Der isolierte MNV-Stamm kann daher einen
kolitogenen Stimulus auf die IL10-defiziente Maus haben. Diese Beobachtungen
(veränderte Histologie und gesteigerte IFNγ-Sekretion) wurden vor allem bei Mäusen
mit dem genetischen Hintergrundstamm C3H/HeJBir gemacht.
Im Infektionsversuch mit keimfreien C3-Il10-/--Mäusen gab es keinen signifikanten
Unterschied bei der Messung der IFNγ-Sekretion zwischen infizierten und nicht
infizierten Mäusen. Histologisch war lediglich eine leicht vermehrte Anzahl
mononukleärer Zellen festzustellen, die für eine Aktivierung der intestinalen
Immunantwort steht. Hieraus lässt sich schlussfolgern, dass murine Noroviren nur in
Kombination mit einer aktiven Darmflora eine verstärkte IFNγ-Sekretion und
histologische Veränderungen hervorrufen.
Die Tötung der keimfreien Mäuse zur Organentnahme für die Histologie sowie zur
Gewinnung von Lymphozyten zur Stimulation wurde 4 Wochen p.i. durchgeführt. In
vorhergehenden Versuchen mit SPF-Mäusen war eine Erhöhung der IFNγ-Sekretion
nach 2 und 4 Wochen gemessen worden. Daher wurde der Zeitpunkt für die Sektion

Diskussion
91
4 Wochen nach der Infektion gewählt. Veränderungen zu einem früheren Zeitpunkt
p.i. können nicht ausgeschlossen werden.
Der Doppelinfektionsversuch zeigte, dass eine vorhergehende Infektion mit MNV
eine 14 Tage später folgende Infektion mit H. hepaticus nicht verstärkte. Anhand der
Befunde der Lymphozytenstimulation aus dem zweiten Infektionsversuch wurde der
Infektionstermin mit H. hepaticus 2 Wochen nach der Infektion mit MNV festgesetzt.
Die histologische Auswertung zeigte keine signifikanten Unterschiede zwischen
Doppelinfektion und Kontrollgruppe, jedoch war der Histologie-Score bei einigen
Mäusen aus der Doppelinfektionsgruppe etwas geringer. Dies könnte für einen leicht
protektiven Effekt der MNV-Infektion sprechen. Dieser ließe sich erklären durch eine
Abschwächung der Immunantwort auf H. hepaticus durch MNV. Da das Virus
antigenpräsentierende Makrophagen befällt und zerstört, könnte die überschießende
Immunreaktion gegen H. hepaticus herabgesetzt sein. Diese Ergebnisse stehen im
Gegensatz zu einer Veröffentlichung von Lencioni et al. (LENCIONI et al. 2008). Hier
wurden jedoch ein anderes Mausmodell (MDR1α-defiziente Maus), ein anderer
Infektionszeitpunkt, ein anderes Norovirus (MNV-4) und eine andere Helicobacter-Art
gewählt. Die Infektion mit H. bilis erfolgte 7 Tage nach der Infektion mit MNV. Die
Tötung der Tiere mit anschließender Sektion erfolgte bei Lencioni et al. variabel nach
Schweregrad der klinischen Symptome.
5.6 Ausblick
Die problemlose Anzucht von MNV ermöglicht neue Erkenntnisse zur Tenazität des
Erregers, was gerade im Bereich Hygieneforschung genutzt werden kann, um gegen
humane Noroviren wirksame Desinfektionsmittel und Hygienestrategien zu
entwickeln (CANNON et al. 2006).
Im Zellkulturmodell lassen sich Pathomechanismen ergründen, wie Noroviren Zellen
schädigen und sich in ihnen vermehren. Dies ermöglicht unter Umständen die
Entwicklung einer Impfung oder Akut-Therapie gegen humane Noroviren. Erste

Diskussion
92
Antikörper wurden durch Müller et al. auf Kreuzreaktivität und Effizienz erprobt
(MÜLLER 2009).
MNV in der Labortierhaltung
Die serologischen Testverfahren ermöglichen eine Routine-Überwachung der
Hygiene in Labortierhaltungen aufgrund der vorhandenen serologischen
Kreuzreaktivität bei verschiedenen murinen Noroviren (HSU et al. 2006). Der
Nachweis per PCR ist möglich, jedoch gibt es bisher kein Primerpaar, mit dem alle
murinen Noroviren detektiert werden können. Dies sollte bei Hygieneuntersuchungen
bedacht werden. Ein negatives PCR-Ergebnis muss nicht zwangsläufig für einen
negativen Bestand stehen (HENDERSON 2008).
MNV hat in der vorliegenden Arbeit keine klinischen Auswirkungen auf das
Tiermodell der IL10-defizienten Maus gehabt. Jedoch kann zum Beispiel eine akute
MNV-Infektion bei IL10-defizienten Mäusen im Tierversuch zu falschen
Vergleichswerten bei einer in vitro Lymphozytenstimulation in der INFγ-Bestimmung
führen. Auch leichte histologische Veränderungen sind möglich. Diese Effekte
können Ergebnisse in Versuchen mit der IL10-defizienten Maus als Tiermodell für
CED verändern. Ein Einfluss von MNV auf andere Mausmodelle kann nicht
ausgeschlossen werden. Weitere Untersuchungen hierzu sind empfehlenswert und
notwendig. Es wurden zudem MNV-Isolate mit unterschiedlichen Eigenschaften
beschrieben. So kann MNV-1 bei schwer immundefizienten Mäusen (RAG2- und
STAT1-defizient) zu tödlichen systemischen Erkrankungen führen (KARST et al.
2003).
Da eine Diagnostik im Rahmen der Routine-Überwachung möglich ist, sollte MNV
daher im Tierbestand langfristig ausgeschlossen werden. Hygienemaßnahmen und
Standardmethoden der Bestandssanierung haben Erfolg gezeigt (ARTWOHL et al.
2008; COMPTON 2008).

Zusammenfassung
93
6 Zusammenfassung
Marcel Achard
„Charakterisierung eines murinen Norovirus – Auswirkungen des Virus auf die
experimentelle chronisch entzündliche Darmerkrankung bei der IL10-defizienten
Maus“
Im Rahmen dieser Dissertation wurde ein murines Norovirus (MNV), das aus
Anzeiger-Mäusen des Zentralen Tierlaboratoriums (ZTL) der Medizinischen
Hochschule Hannover (MHH) isoliert wurde, charakterisiert. Die genetische Sequenz
wurde bestimmt und wies eine 95% Übereinstimmung zu MNV-4 auf. Sie wurde
unter der Zugangsnummer EU854589 veröffentlicht. Zusätzlich wurden
elektronenmikroskopische Aufnahmen der Virusstruktur erstellt.
Die erfolgreiche Anzucht und Vermehrung von MNV in RAW 264.7-Zellen
ermöglichte die Etablierung diagnostischer Tests in Form von PCR und Serologie
(ELISA). Mit Hilfe der serologischen Diagnostik wurde die Prävalenz für MNV im
Zentralen Tierlaboratorium der MHH für den Überwachungszeitraum 2007/2008
erfasst. MNV-positive Anzeiger-Mäuse wurden nur in der experimentellen Barriere
gefunden. Die Prävalenzrate bewegte sich je nach Tierraum zwischen 8% und 61%.
Die Auswirkung von MNV auf das Tiermodell der IL10-defizienten Maus (Il10-/-)
wurde anhand von zwei genetischen Hintergrundstämmen, C3H/HeJBirZtm (C3)
sowie C57BL/6JZtm (B6), im Infektionsversuch untersucht. Alle infizierten Mäuse
wiesen eine Serokonversion auf. Es kam zu keiner klinischen Beeinträchtigung. MNV
war bei den infizierten Mäusen nur in Organen des Gastrointestinaltraktes per PCR
nachweisbar. Kot, Kolon und Mesenteriallymphknoten waren als Probenmaterial
besonders geeignet für einen PCR-Erregernachweis.

Zusammenfassung
94
Es konnten keine schwerwiegenden histologischen Veränderungen im Vergleich zu
nicht mit MNV infizierten Kontrollmäusen ausgemacht werden. Bei einigen mit MNV
infizierten C3-Il10-/--Mäusen wurden jedoch sehr milde fokale entzündliche
Veränderungen im Kolon festgestellt. Die Interferon (INF)γ-Sekretion in vitro
stimulierter Darmlymphozyten von mit MNV infizierten Mäusen war 2 Wochen nach
der Infektion erhöht im Vergleich zur nicht infizierten Kontrollgruppe. Bei infizierten
C3-Il10-/--Mäusen war die IFNγ-Sekretion 4 Wochen nach der Infektion weiterhin
erhöht.
Keimfrei gehaltene C3-Il10-/--Mäuse, die mit MNV infiziert wurden, wiesen 4 Wochen
nach der Infektion keine entzündlichen histologischen Veränderungen oder eine
gesteigerte IFNγ-Sekretion auf. Demnach kann MNV nur in Verbindung mit einer
endogenen bakteriellen Darmflora einen kolitogenen Stimulus erzeugen.
Ein Doppelinfektionsversuch mit MNV und 14 Tage später mit Helicobacter (H.)
hepaticus sollte zeigen, ob eine vorhergehende Infektion mit MNV eine entzündliche
Darmerkrankung, ausgelöst durch H. hepaticus, verstärken kann. Die Kontrollgruppe
wurde nur mit H. hepaticus infiziert. Alle Mäuse wurden 6 Wochen nach der Infektion
mit H. hepaticus seziert. Die Befunde der Klinik, Histologie und IFNγ-Sekretion
wiesen keine deutlichen Unterschiede auf. Hieraus lässt sich folgern, dass MNV die
Entwicklung einer durch H. hepaticus hervorgerufenen Entzündung bei der IL10-
defizienten Maus nicht verstärkt. Dennoch zeigen die vorhergehenden Versuche,
dass die Interaktion von MNV mit einer endogenen apathogenen Darmflora bei der
IL10-defizienten Maus zu einer milden Entzündungsreaktion im Darm führen kann.
Dies macht MNV zu einem nicht zu vernachlässigenden Faktor im Tierversuch.

Summary
95
7 Summary
Marcel Achard
“Characterization of a murine norovirus – the impact of the virus on experimental
inflammatory bowel disease in IL10-deficient mice”
Aim of this study was to characterize a murine norovirus (MNV) that was isolated
from sentinel-mice of the Central Animal Facility (Ztm) at Hannover Medical School.
Genetic sequencing was performed and showed 95% identity with MNV-4. The
sequence was published under the accession number EU85489. Additionally, the
virus structure of MNV was depicted by electron microscopy.
The successful replication of MNV in RAW 264.7-cells allowed us to establish several
diagnostic tests as PCR and serology (ELISA). Therefore, it was possible to
determine the prevalence of MNV at the Ztm for the years of 2007 and 2008. MNV-
positive mice were only found in the experimental barrier. Prevalence among animal
rooms differed from 8% to 61%.
In order to analyse the influence of MNV on IL10-defficient mice (Il10-/-), Il10-/- mice
on genetic background strains C3H/HeJBirZtm (C3) and C57BL/6JZtm (B6) were
experimentally infected. Although seroconversion could be detected in all infected
animals, no clinical effects were visible. The detection of MNV in infected mice was
solely possible in organs of the gastro-intestinal tract. Faeces, colon and mesenteric
lymph nodes were suitable material for the detection of MNV via PCR.
Histological examinations of MNV-positive mice did not show grave differences in
comparison to control mice. Nevertheless, few individuals belonging to the C3-Il10-/-
group showed mild focal inflammations of the colon. In vitro stimulated intestinal
lymphocytes of MNV-positive mice secreted significantly more interferon (IFN)γ two

Summary
96
weeks post infection (p.i.) than those of the control group. Infected animals belonging
to the C3-Il10-/- group showed an increased IFNγ-secretion up to four weeks p.i.
Sterile housed C3-Il10-/--mice that were infected with MNV neither showed
inflammatory changes of the colon nor an increased secretion of IFN-γ four weeks
p.i. Therefore, MNV is only able to induce a colitogenic stimulus in combination with
an endogenous bacterial intestinal flora.
A dual infection experiment with MNV and Helicobacter (H.) hepaticus (14 days
subsequent to the MNV infection) was intended to clarify the question, whether a
preceding infection with MNV aggravates inflammatory diseases of the bowel. A
control group was only infected with H. hepaticus. All mice were sacrificed six weeks
p.i. with H. hepaticus. No obvious differences in clinical signs, intestinal pathology or
INFγ secretion were observed. Therefore, it is suggested that MNV does not
exacerbate inflammatory diseases resulting from a H. hepaticus infection in IL10-
deficient mice. Nevertheless, the preceding experiments demonstrate the interaction
of MNV with the endogenous, non-pathogenic intestinal flora in IL10-deficient mice.
Thus, an infection with MNV should be considered as a confounding variable in
animal experimentation using mouse models.

Anhang
97
8 Anhang
Protokoll zur Isolation von RNA aus Zellkulturüberstand bzw. Organen (gemäß
Hersteller Macherey-Nagel)
1) Bis zu 30 mg Gewebe werden zerkleinert bzw. homogenisiert und in ein
steriles Eppendorfgefäß gegeben; bei Zellkulturüberstand ist dieser Schritt
nicht nötig.
2) 350 µl RA1-Puffer und 3,5 µl ß-Mercaptoethanol werden hinzugegeben;
anschließend wird die Probe für 1 min gevortext.
3) Ein NucleoSpin®- Filter wird in ein Sammelröhrchen gesteckt;
anschließend wird die Probe auf den Filter aufgetragen und für 1 min bei
11000 x g zentrifugiert.
4) Der NucleoSpin®- Filter wird verworfen, zum Filtrat werden 350 µl 70%
Ethanol gegeben.
5) Nach mehrmaligem Durchmischen mit der Pipette wird das Lysat auf eine
NucleoSpin® RNA II-Säule gegeben und für 30 s bei 11000 x g
zentrifugiert.
6) Das Sammelröhrchen wird verworfen, die Säule wird in ein neues
Sammelröhrchen gestellt. 350 µl Membrane Desalting Buffer werden auf
die Säule gegeben, dann wird für 1 min bei 11000 x g zentrifugiert.
7) 95 µl vorbereitete DNase-Reaktionsmixtur werden auf die Säule gegeben,
danach erfolgt eine 15 minütige Inkubation bei Raumtemperatur.
8) Ein erster Waschschritt erfolgt mit 200 µl RA2-Puffer, der auf die Säule
gegeben wird. Dann wird für 30 s bei 11000 x g zentrifugiert. Die Säule wird
in ein neues Sammelröhrchen gesetzt. Ein zweiter Waschschritt erfolgt mit
600 µl RA3-Puffer für 30 s bei 11000 x g. Die Flüssigkeit im
Sammelröhrchen wird verworfen, die Säule wird in das Sammelröhrchen
zurückgestellt. Ein dritter Waschschritt erfolgt mit 250 µl RA3-Puffer für 2
min bei 11000 x g. Anschließend wird die Säule in ein nukleasefreies
Sammelröhrchen gesetzt.

Anhang
98
9) Zum Abschluss wird die RNA mit 40 – 60 µl RNase-freiem Wasser eluiert,
das auf die Säule gegeben wird. Es erfolgt ein letzter Zentrifugationsschritt
bei 11000 x g für 1 min.

Anhang
99
Protokoll zur Isolation von RNA aus Kot (gemäß Hersteller Macherey-Nagel,
modifiziert durch Kunden)
1) Zur Kotprobe werden 350 µl des vorbereiteten ß-Mercaptoethanol-RA1
gegeben.
2) Anschließend wird die das Gemisch für 2 min gevortext.
3) Ein NucleoSpin®- Filter wird in ein Sammelröhrchen gesteckt;
anschließend wird das Gemisch auf den Filter aufgetragen und für 1 min
bei 11000 x g zentrifugiert.
4) Der Überstand wird in ein neues Gefäß überführt, das sichtbare Pellet
verworfen.
5) Zum Überstand werden 350 µl 70% Ethanol gegeben.
6) Nach einer Inkubation von 5 min wird das Gemisch nach mehrmaligem
Durchmischen mit der Pipette auf eine NucleoSpin® RNA II-Säule
aufgetragen und für 30 s bei 11000 x g zentrifugiert.
7) Das Sammelröhrchen wird verworfen, die Säule wird in ein neues
Sammelröhrchen gestellt. 350 µl Membrane Desalting Buffer werden auf
die Säule gegeben, dann wird für 1 min bei 11000 x g zentrifugiert.
8) 95 µl vorbereitete DNase-Reaktionsmixtur werden auf die Säule gegeben,
danach erfolgt eine 15 minütige Inkubation bei Raumtemperatur.
9) Ein erster Waschschritt erfolgt mit 200 µl RA2-Puffer, der auf die Säule
gegeben wird. Dann wird für 30 s bei 11000 x g zentrifugiert. Die Säule wird
in ein neues Sammelröhrchen gesetzt.Ein zweiter Waschschritt erfolgt mit
600 µl RA3-Puffer für 30 s bei 11000 x g. Die Flüssigkeit im
Sammelröhrchen wird verworfen, die Säule wird in das Sammelröhrchen
zurückgestellt.Ein dritter Waschschritt erfolgt mit 250 µl RA3-Puffer für 2
min bei 11000 x g. Anschließend wird die Säule in ein nukleasefreies
Sammelröhrchen gesetzt.
10) Zum Abschluss wird die RNA mit 40 – 60 µl RNase-freiem Wasser eluiert,
das auf die Säule gegeben wird. Es erfolgt ein letzter Zentrifugationsschritt
bei 11000 x g für 1 min.

Anhang
100
Rohdaten ELISA-Messung
Intraseriell
Serumpool OD MNV OD RAW OD korrigiert MW SD
+ 3,474 0,773 2,701 + 3,348 0,717 2,631 + 3,505 0,69 2,815 + 3,475 0,681 2,794 + 3,524 0,738 2,786 + 3,535 0,806 2,729 + 3,524 0,713 2,811 + 3,492 0,739 2,753 + 3,44 0,726 2,714 + 3,593 0,709 2,884 2,7618 0,071530568
- 0,673 0,591 0,082 - 0,651 0,523 0,128 - 0,757 0,607 0,15 - 0,752 0,592 0,16 - 0,677 0,612 0,065 - 0,7 0,609 0,091 0,11266667 0,03886215
+: MNV-positiv; -: MNV-negativ
Interseriell
Positivserumpool
Tag OD MNV OD RAW OD korrigiert MW SD
1 3,601 0,714 2,887 2 3,671 0,843 2,828 3 3,52 0,706 2,814 4 3,72 0,776 2,944 5 3,737 0,905 2,832 6 3,627 0,8 2,827 7 3,801 0,949 2,852 8 3,594 0,67 2,924 9 3,836 1,133 2,703 10 3,559 0,96 2,599 2,821 0,10247927

Anhang
101
Negativserumpool
Tag OD MNV OD RAW OD korrigiert MW SD
1 0,691 0,612 0,079
2 0,613 0,658 -0,045
3 0,705 0,653 0,052
4 0,809 0,732 0,077
5 0,638 0,656 -0,018
6 0,784 0,773 0,011
7 0,725 0,617 0,108
8 0,503 0,521 -0,018
9 0,726 0,759 -0,033
10 0,903 0,812 0,091 0,0304 0,05721344
Linearität
Serumverdünnung OD MNV OD RAW OD korrigiert MW SD
1:25 3,495 0,874 2,621
3,527 0,885 2,642
3,4 0,938 2,462 2,575 0,09842256
1:50 2,946 0,721 2,225
3,318 0,76 2,558
3,438 0,876 2,562 2,44833333 0,19342268
1:100 2,741 0,527 2,214
3,077 0,534 2,543
3,129 0,549 2,58 2,44566667 0,20148035
1:200 2,74 0,415 2,325
2,624 0,389 2,235
2,647 0,409 2,238 2,266 0,05111751
1:400 2,149 0,332 1,817
2,592 0,321 2,271
2,327 0,335 1,992 2,02666667 0,22897671
1:800 1,826 0,248 1,578
1,725 0,244 1,481
1,693 0,237 1,456 1,505 0,06444377

Anhang
102
Einzelmessungen
Negativ-Seren N=68
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert
Prozentualer Anteil (von 68 negativen Seren)
11146 - 3,032 3,375 -0,34 4,41
8428 - 1,56 1,883 -0,32 4050 - 2,122 2,434 -0,31 8422 - 0,167 0,402 -0,24 8,82
4053 - 2,146 2,369 -0,22 4052 - 0,414 0,592 -0,18 8423 - 0,207 0,359 -0,15 11144 - 1,171 1,32 -0,15 8418 - 0,235 0,383 -0,15 11145 - 0,478 0,606 -0,13 32,35
4054 - 0,386 0,512 -0,13 8426 - 0,172 0,297 -0,13 11143 - 0,322 0,425 -0,10 4057 - 0,278 0,375 -0,10 8431 - 2,744 2,841 -0,10 4051 - 0,379 0,468 -0,09 4058 - 0,491 0,577 -0,09 10096 - 0,296 0,38 -0,08 4049 - 0,54 0,623 -0,08 11153 - 0,257 0,336 -0,08 4059 - 0,386 0,463 -0,08 11161 - 1,432 1,503 -0,07 10095 - 0,218 0,287 -0,07 8427 - 0,2 0,265 -0,07 11158 - 0,274 0,336 -0,06 8420 - 0,164 0,222 -0,06 11156 - 0,236 0,288 -0,05 10098 - 0,298 0,35 -0,05 10089 - 0,331 0,382 -0,05 10093 - 0,287 0,334 -0,05 8430 - 0,178 0,223 -0,05 8421 - 0,23 0,273 -0,04 42,64
10097 - 0,303 0,345 -0,04 8425 - 0,154 0,187 -0,03 4055 - 0,381 0,414 -0,03 8419 - 0,272 0,305 -0,03 8429 - 0,175 0,203 -0,03 11149 - 0,235 0,26 -0,03 4769 - 0,456 0,481 -0,03 10088 - 0,372 0,395 -0,02 11150 - 0,226 0,244 -0,02 4048 - 0,411 0,423 -0,01 4056 - 0,393 0,403 -0,01 8424 - 0,185 0,194 -0,01 11147 - 0,212 0,221 -0,01

Anhang
103
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert Prozentualer Anteil
(von 68 negativen Seren)
10083 - 0,577 0,586 -0,01 10099 - 0,265 0,273 -0,01 11152 - 0,26 0,267 -0,01 10084 - 0,258 0,26 0,00 10092 - 0,31 0,305 0,01 11155 - 0,146 0,126 0,02 10090 - 0,31 0,286 0,02 11151 - 0,324 0,298 0,03 10087 - 0,212 0,181 0,03 10086 - 0,611 0,578 0,03 11159 - 0,214 0,18 0,03 11154 - 0,296 0,26 0,04 10081 - 0,267 0,23 0,04 11160 - 0,217 0,177 0,04 10082 - 0,228 0,186 0,04 10085 - 0,303 0,253 0,05 11148 - 0,312 0,255 0,06 10100 - 0,319 0,231 0,09 4771 - 0,525 0,433 0,09 11157 - 1,54 1,441 0,10 1,47
10094 - 1,306 1,155 0,15 4,41
10091 - 0,822 0,653 0,17 4770 - 0,535 0,324 0,21

Anhang
104
Positiv-Seren N=76
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert
Prozentualer Anteil (von 76 positiven Seren)
8338 +/- EL + 1,509 0,447 1,06 1,32
4131 + 1,824 0,608 1,22 1,32
10180 NSR EL+ 3,347 2,087 1,26 1,32
4454 +++ 2,26 0,828 1,43 2,63
10176 NSR EL+ 2,861 1,428 1,43 10181 NSR EL+ 3,313 1,83 1,48 3,95
4130 ++ 2,126 0,611 1,52 8725 ++ 3,348 1,812 1,54 8727 + 2,356 0,767 1,59 2,63
8344 + 2,188 0,578 1,61 4125 ++ 2,257 0,611 1,65 3,95
4453 + 2,41 0,683 1,73 8339 + 2,336 0,6 1,74 10173 +++ 2,66 0,879 1,78 6,58
8666 + 2,16 0,369 1,79 4451 ++ 2,884 1,061 1,82 8658 + 3,502 1,675 1,83 4126 ++ 2,641 0,781 1,86 4127 + 2,373 0,476 1,90 3,95
8718 ++ 3,019 1,099 1,92 4132 ++ 2,563 0,625 1,94 8723 + 2,487 0,525 1,96 3,95
8343 +++ 2,993 1,017 1,98 8676 ++ 3,073 1,057 2,02 4450 +++ 3,473 1,407 2,07 5,26
10191 + 3,339 1,259 2,08 4124 ++ 2,812 0,725 2,09 10174 ++ 2,723 0,596 2,13 8711 ++ 2,807 0,638 2,17 5,26
8677 ++ 2,882 0,71 2,17 10172 NSR EL+ 3,547 1,344 2,20 8648 + 3,119 0,913 2,21 8341 + 2,597 0,348 2,25 11,84
10178 + 2,851 0,593 2,26 8674 ++ 3,38 1,115 2,27 8675 + 3,283 1,007 2,28 4129 +++ 3,092 0,808 2,28 MAG03 +++ 2,675 0,386 2,29 8724 + 2,549 0,23 2,32 8669 +++ 3,342 1,021 2,32 MAG02 ++ 2,912 0,569 2,34 10175 +++ 3,122 0,776 2,35 15,79
8679 + 3,202 0,84 2,36 8719 +++ 3,155 0,792 2,36 4452 ++ 3,236 0,858 2,38 10171 NSR EL+ 3,512 1,123 2,39 8671 + 3,215 0,822 2,39

Anhang
105
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert Prozentualer Anteil
(von 76 positiven Seren)
8656 + 3,5 1,101 2,40 8650 ++ 3,223 0,82 2,40 8667 + 3,341 0,934 2,41 MAG04 ++ 2,849 0,438 2,41 MAG05 ++ 2,869 0,458 2,41 8668 + 3,104 0,686 2,42 8678 + 3,09 0,613 2,48 5,26
8716 ++ 2,955 0,478 2,48 8665 + 3,322 0,803 2,52 MAG01 ++ 3,029 0,487 2,54 8663 ++ 3,485 0,923 2,56 11,84
8680 +++ 3,263 0,685 2,58 8722 +++ 3,396 0,815 2,58 10177 ++ 3,079 0,497 2,58 10190 +++ 3,332 0,742 2,59 10189 +/- EL + 3,464 0,864 2,60 10004 +++ 3,386 0,773 2,61 10001 +++ 3,152 0,537 2,62 8662 + 3,233 0,596 2,64 10185 +++ 3,083 0,427 2,66 5,26
8661 + 3,307 0,638 2,67 8660 + 3,442 0,736 2,71 10002 +++ 3,293 0,585 2,71 8681 ++ 3,163 0,417 2,75 5,26
8657 ++ 3,403 0,61 2,79 8720 ++ 3,557 0,758 2,80 8670 ++ 3,392 0,587 2,81 4503 +++ 3,116 0,276 2,84 1,32
10003 +++ 3,431 0,436 3,00 1,32
+: schwach positive Reaktion; ++: Mittelstarke positive Reaktion; +++: Stark positive Reaktion NSR: Nicht spezifische Reaktion; +/-: Grenzwertige Reaktion; EL+: Charles-River-ELISA positiv

Anhang
106
Negativ-Seren N=205
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert Prozentualer Anteil
(von 205 Seren)
3786 - 0,782 1,167 -0,385 0,98
3830 - 0,93 1,284 -0,354
2276 - 1,711 2,048 -0,337 2,44
3122 - 0,162 0,48 -0,318
3118 - 1,232 1,517 -0,285
3559 - 0,352 0,622 -0,27
3567 - 0,507 0,776 -0,269
3563 - 0,66 0,892 -0,232 4,88
3551 - 0,602 0,828 -0,226
3555 - 0,804 1,021 -0,217
3787 - 1,378 1,579 -0,201
3782 - 0,957 1,15 -0,193
4835 - 1,416 1,608 -0,192
3547 - 0,45 0,614 -0,164
3778 - 0,898 1,058 -0,16
3837 - 0,567 0,719 -0,152
3557 - 0,426 0,578 -0,152
3036 - 0,505 0,652 -0,147 8,78
2275 - 0,839 0,984 -0,145
3794 - 0,548 0,688 -0,14
3790 - 0,689 0,823 -0,134
2343 - 0,599 0,732 -0,133
2994 - 0,673 0,8 -0,127
2530 - 0,353 0,464 -0,111
2154 - 0,658 0,755 -0,097
2212 - 0,657 0,737 -0,08
3017 - 0,829 0,898 -0,069
3014 - 1,006 1,075 -0,069
3793 - 0,505 0,573 -0,068
2995 - 0,823 0,888 -0,065
2494 - 0,512 0,575 -0,063
3561 - 0,306 0,365 -0,059
2204 - 0,476 0,534 -0,058
2496 - 0,376 0,432 -0,056
3834 - 1,717 1,772 -0,055
3554 - 0,747 0,793 -0,046 22,44
2274 - 1,63 1,675 -0,045
3549 - 0,321 0,365 -0,044
2550 - 0,964 1,006 -0,042
3562 - 0,75 0,792 -0,042
3813 - 0,687 0,727 -0,04
3833 - 1,02 1,056 -0,036
2528 - 0,344 0,378 -0,034
2344 - 0,784 0,815 -0,031
3801 - 0,551 0,582 -0,031
3553 - 0,71 0,739 -0,029
2384 - 1,323 1,348 -0,025

Anhang
107
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert Prozentualer Anteil
(von 205 Seren)
2551 - 0,433 0,458 -0,025
3016 - 0,56 0,584 -0,024
3548 - 0,39 0,412 -0,022
4833 - 0,776 0,796 -0,02
3029 - 0,725 0,742 -0,017 3119 - 1,342 1,359 -0,017
2532 - 0,43 0,445 -0,015
3032 - 0,544 0,558 -0,014
2524 - 1,253 1,26 -0,007
3564 - 1,457 1,464 -0,007
2200 - 0,846 0,851 -0,005
3781 - 0,868 0,873 -0,005
3797 - 0,599 0,602 -0,003
3010 - 0,617 0,615 0,002
2529 - 0,417 0,407 0,01
2999 - 0,942 0,93 0,012
2805 - 0,142 0,128 0,014
3550 - 0,56 0,544 0,016
3809 - 0,518 0,502 0,016
neg.Kontrolpool - 0,879 0,862 0,017
3802 - 1,029 1,01 0,019
2192 - 0,492 0,47 0,022
4836 - 0,292 0,267 0,025
3005 - 0,937 0,911 0,026
2385 - 0,82 0,791 0,029
3006 - 0,635 0,604 0,031
2194 - 0,523 0,49 0,033
3818 - 1,004 0,968 0,036
3817 - 0,732 0,692 0,04
2672 - 0,832 0,791 0,041
3565 - 0,331 0,288 0,043
2195 - 0,891 0,845 0,046
3024 - 0,727 0,678 0,049
3810 - 0,837 0,788 0,049
3560 - 0,612 0,562 0,05 22,93
3821 - 0,772 0,719 0,053
3015 - 0,941 0,887 0,054
2197 - 0,576 0,521 0,055
2998 - 0,66 0,604 0,056
3030 - 0,443 0,385 0,058
4834 - 0,341 0,283 0,058
3558 - 0,497 0,437 0,06
3025 - 0,912 0,852 0,06
3803 - 0,648 0,587 0,061
3000 - 0,515 0,446 0,069
2341 - 1,088 1,019 0,069
2996 - 0,614 0,542 0,072
2342 - 0,685 0,613 0,072
2340 - 1,28 1,206 0,074

Anhang
108
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert Prozentualer Anteil
(von 205 Seren)
2160 - 0,679 0,604 0,075
3038 - 0,545 0,47 0,075
2673 - 0,716 0,638 0,078
2552 - 0,871 0,792 0,079
3556 - 0,778 0,699 0,079 2193 - 0,498 0,418 0,08
2671 - 0,499 0,414 0,085
3783 - 0,939 0,852 0,087
3779 - 1,259 1,168 0,091
2196 - 0,812 0,714 0,098
2152 - 0,693 0,59 0,103
3039 - 0,838 0,735 0,103
3785 - 0,892 0,789 0,103
3808 - 0,689 0,585 0,104
3566 - 0,499 0,394 0,105
3026 - 1,089 0,983 0,106
3028 - 0,405 0,295 0,11
2958 - 0,783 0,671 0,112
2993 - 0,739 0,626 0,113
2526 - 1,004 0,883 0,121
2670 - 0,948 0,826 0,122
2153 - 0,708 0,578 0,13
3796 - 0,851 0,718 0,133
3806 - 0,859 0,724 0,135
3805 - 0,593 0,457 0,136
2203 - 0,642 0,504 0,138
3791 - 0,946 0,804 0,142
3018 - 1,309 1,166 0,143
3013 - 0,933 0,789 0,144
3033 - 0,887 0,741 0,146
2213 - 0,863 0,716 0,147
3001 - 0,689 0,541 0,148
3829 - 0,989 0,836 0,153 12,68
3835 - 0,951 0,797 0,154
3121 - 0,302 0,147 0,155
2527 - 1,757 1,595 0,162
3003 - 0,73 0,568 0,162
2531 - 0,47 0,306 0,164
3799 - 0,961 0,794 0,167
2188 - 0,594 0,417 0,177
3811 - 0,596 0,411 0,185
3815 - 0,758 0,573 0,185
3812 - 0,936 0,748 0,188
3820 - 0,739 0,55 0,189
3011 - 0,971 0,78 0,191
3552 - 0,733 0,539 0,194
3819 - 0,966 0,77 0,196
4943 - 0,639 0,442 0,197
3031 - 0,722 0,523 0,199

Anhang
109
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert Prozentualer Anteil
(von 205 Seren)
2273 - 2,05 1,847 0,203
3807 - 0,792 0,585 0,207
3795 - 0,843 0,634 0,209
2523 - 1,456 1,244 0,212
3784 - 1,721 1,495 0,226 2522 - 1,52 1,293 0,227
2201 - 0,902 0,665 0,237
3035 - 1,161 0,924 0,237
2495 - 0,605 0,366 0,239
3814 - 1,11 0,86 0,25 10,73
3034 - 0,803 0,537 0,266
3792 - 0,85 0,577 0,273
3827 - 0,984 0,71 0,274
3831 - 1,056 0,778 0,278
2497 - 0,688 0,409 0,279
3823 - 0,694 0,415 0,279
3826 - 1,061 0,776 0,285
3544 - 0,675 0,39 0,285
3800 - 0,686 0,4 0,286
2997 - 2,061 1,774 0,287
3648 - 3,094 2,805 0,289
2954 - 0,785 0,495 0,29
2525 - 1,165 0,873 0,292
4946 - 0,751 0,456 0,295
3023 - 0,828 0,531 0,297
3789 - 1,497 1,19 0,307
3004 - 0,969 0,661 0,308
3825 - 1,224 0,904 0,32
3009 - 1,199 0,877 0,322
3027 - 0,991 0,666 0,325
3804 - 0,829 0,488 0,341
3816 - 0,993 0,643 0,35 3,9
3836 - 1,048 0,696 0,352
3022 - 1,483 1,123 0,36
3020 - 1,297 0,935 0,362
3822 - 1,713 1,333 0,38
3002 - 1,281 0,893 0,388
3019 - 1,331 0,933 0,398
2151 - 1,608 1,18 0,428
4830 - 1,412 0,951 0,461 2,44
3012 - 1,381 0,907 0,474
3788 - 1,516 1,006 0,51
2211 - 1,143 0,614 0,529
3546 - 1,684 1,153 0,531
3037 - 1,494 0,913 0,581 3,41
3824 - 1,361 0,774 0,587
3021 - 2,079 1,459 0,62
3832 - 1,861 1,228 0,633
4832 - 2,26 1,623 0,637

Anhang
110
Biodoc-Nr. IFA OD MNV OD RAW OD korrigiert Prozentualer Anteil
(von 205 Seren)
3007 - 1,482 0,845 0,637
3545 - 1,78 1,131 0,649
3120b - 1,515 0,854 0,661 2,44
3780 - 1,877 1,194 0,683
2966 - 1,802 1,117 0,685 3649 - 2,998 2,276 0,722
3008 - 3,578 2,832 0,746
3650 - 1,365 0,482 0,883 0,49
3647 - 4,636 3,653 0,983 0,49
3828 - 2,185 1,078 1,107 0,49
3798 - 2,428 1,006 1,422 0,49

Anhang
111
Werte der Interferon γ-Messung (ng/ml) nach in vitro Stimulation
Infektionsversuch 1
B6-Il10-/- m
B6-Il10-/- w
Kontrolle 2 Wochen 4 Wochen 8 Wochen Kontrolle 2 Wochen 4 Wochen 8 Wochen
21,002 162,35 27,691 1,804 11,481 179 10,3 2,093
40,778 164,02 25,914 1,6 10,358 172 22,539 1,974
24,267 119 20,508 1,6 13,52 78,676 18,599 1,6
43,129 81,62 15,981 1,6 6,696 52,232 17,027 1,6
22,279 84,952 19,176 1,6 9,89 66,46 16,575 1,6
52,36 89,109 27,472 3,06 12,541 48,36 32,708 1,6
27,089
38,835
15,756
9,782
10,59
18,992
C3-Il10-/- m
C3-Il10-/- w
Kontrolle 2 Wochen 4 Wochen Kontrolle 2 Wochen 4 Wochen 8 Wochen
0,3 79,938 395 216 103 188 117
0,3 61,699 292 321,004 121 184 315
0,472 99,934 443 278,172 144 276 134
0,3 112 382 199 205 202 128
0,3 60,732 160 104 237 122 176
0,3 117 342 356,347 167 171 109
64,899
201
128
402
67,924
99,694
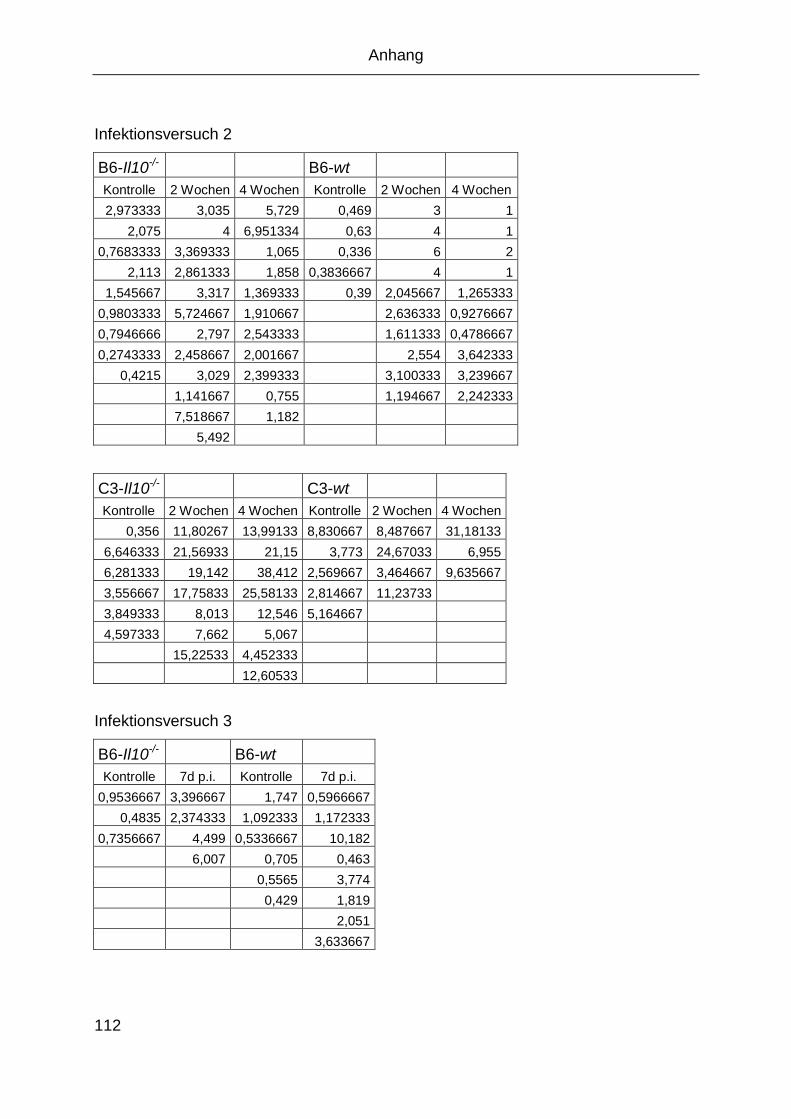
Anhang
112
Infektionsversuch 2
B6-Il10-/-
B6-wt
Kontrolle 2 Wochen 4 Wochen Kontrolle 2 Wochen 4 Wochen
2,973333 3,035 5,729 0,469 3 1
2,075 4 6,951334 0,63 4 1
0,7683333 3,369333 1,065 0,336 6 2
2,113 2,861333 1,858 0,3836667 4 1
1,545667 3,317 1,369333 0,39 2,045667 1,265333
0,9803333 5,724667 1,910667
2,636333 0,9276667
0,7946666 2,797 2,543333
1,611333 0,4786667
0,2743333 2,458667 2,001667
2,554 3,642333
0,4215 3,029 2,399333
3,100333 3,239667
1,141667 0,755
1,194667 2,242333
7,518667 1,182
5,492
C3-Il10-/-
C3-wt
Kontrolle 2 Wochen 4 Wochen Kontrolle 2 Wochen 4 Wochen
0,356 11,80267 13,99133 8,830667 8,487667 31,18133
6,646333 21,56933 21,15 3,773 24,67033 6,955
6,281333 19,142 38,412 2,569667 3,464667 9,635667
3,556667 17,75833 25,58133 2,814667 11,23733 3,849333 8,013 12,546 5,164667
4,597333 7,662 5,067
15,22533 4,452333
12,60533
Infektionsversuch 3
B6-Il10-/-
B6-wt
Kontrolle 7d p.i. Kontrolle 7d p.i.
0,9536667 3,396667 1,747 0,5966667
0,4835 2,374333 1,092333 1,172333
0,7356667 4,499 0,5336667 10,182
6,007 0,705 0,463
0,5565 3,774
0,429 1,819
2,051
3,633667

Anhang
113
Infektionsversuch 4
C3-Il10-/- ( keimfrei)
Kontrolle 4 Wochen
0,9279273 0,756581
0,969763 1,001276
1,777933 1,445699
1,157028 0,830935
1,817827
Infektionsversuch 5
C3-Il10-/-
B6-Il10-/-
H.hepaticus MNV + H.hepaticus H.hepaticus MNV + H.hepaticus
33,9378 26,05871 38,18218 46,39897
48,97928 50,26017 42,69127 9,08253
38,03638 45,06061 57,02307 24,43101
53,10282 54,62005 16,87314 12,40349
36,76929 55,69619 40,08938 36,34947
51,71358 50,76093
61,21308

Anhang
114
TJL-Histologie-Werte für Infektionsversuch 5
Zäkum C3-Il10-/-
B6-Il10-/-
Helico Helico+MNV Helico Helico+MNV
5 4 8 6
6 4 6 5
5 5 7 5
6 6 5 5
5 5 7 7
7 7
5
Kolon Helico Helico+MNV Helico Helico+MNV
11 10 15 18
14 7 14 12
11 10 14 6
12 12 11 12
10 7 17 13
10 11
12
Helico: Infektion mit H. hepaticus; Helico+MNV: Doppelinfektion mit MNV und H.
hepaticus

Literaturverzeichnis
115
9 Literaturverzeichnis
ALTSCHUL, S. F., W. GISH, W. MILLER, E. W. MYERS u. D. J. LIPMAN (1990): Basic local alignment search tool. J Mol Biol 215, 403-410 ARTWOHL, J. E., J. E. PURCELL u. J. D. FORTMAN (2008): The use of cross-foster rederivation to eliminate murine norovirus, Helicobacter spp., and murine hepatitis virus from a mouse colony. J Am Assoc Lab Anim Sci 47, 19-24 BAERT, L., C. E. WOBUS, E. VAN COILLIE, L. B. THACKRAY, J. DEBEVERE u. M. UYTTENDAELE (2008): Detection of murine norovirus 1 by using plaque assay, transfection assay, and real-time reverse transcription-PCR before and after heat exposure. Appl Environ Microbiol 74, 543-546 BAILEY, D., L. B. THACKRAY u. I. G. GOODFELLOW (2008): A single amino acid substitution in the murine norovirus capsid protein is sufficient for attenuation in vivo. J Virol 82, 7725-7728 BALISH, E. u. T. WARNER (2002): Enterococcus faecalis induces inflammatory bowel disease in interleukin-10 knockout mice. Am J Pathol 160, 2253-2257 BAUMGART, D. C. u. S. R. CARDING (2007): Inflammatory bowel disease: cause and immunobiology. Lancet 369, 1627-1640 BAUMGART, D. C. u. W. J. SANDBORN (2007): Inflammatory bowel disease: clinical aspects and established and evolving therapies. Lancet 369, 1641-1657 BERG, D. J., N. DAVIDSON, R. KÜHN, W. MÜLLER, S. MENON, G. HOLLAND, L. THOMPSON-SNIPES, M. W. LEACH u. D. RENNICK (1996): Enterocolitis and colon cancer in interleukin-10-deficient mice are associated with aberrant cytokine production and CD4(+) TH1-like responses. J Clin Invest 98, 1010-1020 BLAKENEY, S. J., A. CAHILL u. P. A. REILLY (2003): Processing of Norwalk virus nonstructural proteins by a 3C-like cysteine proteinase. Virology 308, 216-224

Literaturverzeichnis
116
BLEICH, A. (2003) Chronisch entzündliche Darmerkrankungen im Modell der Interleukin-10 defizienten Maus: QTL- und Kandidatengenanalysen. Hannover, Tierärztliche Hochschule Hannover, Institut für Versuchstierkunde der Medizinischen Hochschule Hannover, Diss. BLEICH, A., M. MAHLER, C. MOST, E. H. LEITER, E. LIEBLER-TENORIO, C. O. ELSON, H. J. HEDRICH, B. SCHLEGELBERGER u. J. P. SUNDBERG (2004): Refined histopathologic scoring system improves power to detect colitis QTL in mice. Mamm Genome 15, 865-871 BOIRIVANT, M., I. J. FUSS, A. CHU u. W. STROBER (1998): Oxazolone colitis: A murine model of T helper cell type 2 colitis treatable with antibodies to interleukin 4. J Exp Med 188, 1929-1939 BOK, K., V. G. PRIKHODKO, K. Y. GREEN u. S. V. SOSNOVTSEV (2009): Apoptosis in murine norovirus-infected RAW264.7 cells is associated with downregulation of survivin. J Virol 83, 3647-3656 BRISTOL, I. J., M. A. FARMER, Y. CONG, X. X. ZHENG, T. B. STROM, C. O. ELSON, J. P. SUNDBERG u. E. H. LEITER (2000): Heritable susceptibility for colitis in mice induced by IL-10 deficiency. Inflamm Bowel Dis 6, 290-302 BÜCHLER, G. (2010) Charakterisierung des Hauptsuszeptibilitätslocus Cdcs1 für experimentelle, chronisch entzündliche Darmerkrankungen. Hannover, Tierärztliche Hochschule Hannover, Institut für Versuchstierkunde der Medizinischen Hochschule Hannover, Diss. BURICH, A., R. HERSHBERG, K. WAGGIE, W. ZENG, T. BRABB, G. WESTRICH, J. L. VINEY u. L. MAGGIO-PRICE (2001): Helicobacter-induced inflammatory bowel disease in IL-10- and T cell-deficient mice. Am J Physiol Gastrointest Liver Physiol 281, G764-778 CAHILL, R. J., C. J. FOLTZ, J. G. FOX, C. A. DANGLER, F. POWRIE u. D. B. SCHAUER (1997): Inflammatory bowel disease: an immunity-mediated condition triggered by bacterial infection with Helicobacter hepaticus. Infect Immun 65, 3126-3131 CANNON, J. L., E. PAPAFRAGKOU, G. W. PARK, J. OSBORNE, L. A. JAYKUS u. J. VINJE (2006): Surrogates for the study of norovirus stability and inactivation in the environment: A comparison of murine norovirus and feline calicivirus.

Literaturverzeichnis
117
J Food Prot 69, 2761-2765 CHACHU, K. A., D. W. STRONG, A. D. LOBUE, C. E. WOBUS, R. S. BARIC u. H. W. T. VIRGIN (2008): Antibody is critical for the clearance of murine norovirus infection. J Virol 82, 6610-6617 COMPTON, S. R. (2008): Prevention of murine norovirus infection in neonatal mice by fostering. J Am Assoc Lab Anim Sci 47, 25-30 COMPTON, S. R., F. X. PATURZO u. J. D. MACY (2010): Effect of murine norovirus infection on mouse parvovirus infection. J Am Assoc Lab Anim Sci 49, 11-21 DOLIN, R., N. R. BLACKLOW, H. DUPONT, R. F. BUSCHO, R. G. WYATT, J. A. KASEL, R. HORNICK u. R. M. CHANOCK (1972): Biological properties of Norwalk agent of acute infectious nonbacterial gastroenteritis. Proc Soc Exp Biol Med 140, 578-583 DOOM, C. M., H. M. TURULA u. A. B. HILL (2009): Investigation of the impact of the common animal facility contaminant murine norovirus on experimental murine cytomegalovirus infection. Virology 392, 153-161 ELSON, C. O. (1999): Experimental models of intestinal inflammation. New insights into mechanisms of mucosal homeostasis. In: Handbook of Mucosal Immunology Academic Press, San Diego, 1007-1024 ELSON, C. O., K. W. BEAGLEY, A. T. SHARMANOV, K. FUJIHASHI, H. KIYONO, G. S. TENNYSON, Y. CONG, C. A. BLACK, B. W. RIDWAN u. J. R. MCGHEE (1996): Hapten-induced model of murine inflammatory bowel disease: mucosa immune responses and protection by tolerance. J Immunol 157, 2174-2185 FARROKHYAR, F., E. T. SWARBRICK u. E. J. IRVINE (2001): A critical review of epidemiological studies in inflammatory bowel disease. Scand J Gastroenterol 36, 2-15 FIELDS, B. N., K. Y. GREEN, D. M. KNIPE u. P. M. HOWLEY (2007): Caliciviridae: The Noroviruses. In: Fields Virology, 5th Edition Wolters Kluwer Health/Lippincott Williams & Wilkins, Philadelphia, 949-979

Literaturverzeichnis
118
GERONDOPOULOS, A., T. JACKSON, P. MONAGHAN, N. DOYLE u. L. O. ROBERTS (2010): Murine norovirus-1 cell entry is mediated through a non-clathrin-, non-caveolae-, dynamin- and cholesterol-dependent pathway. J Gen Virol 91, 1428-1438 GOTO, K., N. HAYASHIMOTO, M. YASUDA, T. ISHIDA, S. KAMEDA, A. TAKAKURA u. T. ITOH (2009): Molecular detection of murine norovirus from experimentally and spontaneously infected mice. Exp Anim 58, 135-140 HAN, K. R., Y. CHOI, B. S. MIN, H. JEONG, D. CHEON, J. KIM, Y. JEE, S. SHIN u. J. M. YANG (2010): Murine norovirus-1 3Dpol exhibits RNA-dependent RNA polymerase activity and nucleotidylylates on Tyr of the VPg. J Gen Virol 91, 1713-1722 HENDERSON, K. S. (2008): Murine norovirus, a recently discovered and highly prevalent viral agent of mice. Lab Anim (NY) 37, 314-320 HENSLEY, S. E., A. K. PINTO, H. D. HICKMAN, R. J. KASTENMAYER, J. R. BENNINK, H. W. VIRGIN u. J. W. YEWDELL (2009): Murine norovirus infection has no significant effect on adaptive immunity to vaccinia virus or influenza A virus. J Virol 83, 7357-7360 HOLLANDER, G. A., S. J. SIMPSON, E. MIZOGUCHI, A. NICHOGIANNOPOULOU, J. SHE, J. C. GUTIERREZ-RAMOS, A. K. BHAN, S. J. BURAKOFF, B. WANG u. C. TERHORST (1995): Severe colitis in mice with aberrant thymic selection. Immunity 3, 27-38 HSU, C. C., L. K. RILEY u. R. S. LIVINGSTON (2007): Molecular characterization of three novel murine noroviruses. Virus Genes 34, 147-155 HSU, C. C., L. K. RILEY, H. M. WILLS u. R. S. LIVINGSTON (2006): Persistent infection with and serologic cross-reactivity of three novel murine noroviruses. Comp Med 56, 247-251 HSU, C. C., C. E. WOBUS, E. K. STEFFEN, L. K. RILEY u. R. S. LIVINGSTON (2005):

Literaturverzeichnis
119
Development of a microsphere-based serologic multiplexed fluorescent immunoassay and a reverse transcriptase PCR assay to detect murine norovirus 1 infection in mice. Clin Diagn Lab Immunol 12, 1145-1151 HYDE, J. L. u. J. M. MACKENZIE (2010): Subcellular localization of the MNV-1 ORF1 proteins and their potential roles in the formation of the MNV-1 replication complex. Virology 406, 138-148 HYDE, J. L., S. V. SOSNOVTSEV, K. Y. GREEN, C. WOBUS, H. W. VIRGIN u. J. M. MACKENZIE (2009): Mouse norovirus replication is associated with virus-induced vesicle clusters originating from membranes derived from the secretory pathway. J Virol 83, 9709-9719 KARST, S. M., C. E. WOBUS, M. LAY, J. DAVIDSON u. H. W. T. VIRGIN (2003): STAT1-dependent innate immunity to a Norwalk-like virus. Science 299, 1575-1578 KASTENMAYER, R. J., K. A. PERDUE u. W. R. ELKINS (2008): Eradication of murine norovirus from a mouse barrier facility. J Am Assoc Lab Anim Sci 47, 26-30 KATPALLY, U., N. R. VOSS, T. CAVAZZA, S. TAUBE, J. R. RUBIN, V. L. YOUNG, J. STUCKEY, V. K. WARD, H. W. T. VIRGIN, C. E. WOBUS u. T. J. SMITH (2010): High-resolution cryo-electron microscopy structures of murine norovirus 1 and rabbit hemorrhagic disease virus reveal marked flexibility in the receptor binding domains. J Virol 84, 5836-5841 KELMENSON, J. A., D. P. POMERLEAU, S. GRIFFEY, W. ZHANG, M. J. KAROLAK u. J. R. FAHEY (2009): Kinetics of transmission, infectivity, and genome stability of two novel mouse norovirus isolates in breeding mice. Comp Med 59, 27-36 KIM, M., H. LEE, K. O. CHANG u. G. KO (2010): Molecular characterization of murine norovirus isolates from South Korea. Virus Res 147, 1-6 KRAFT, V. u. B. MEYER (1990): Seromonitoring in small laboratory animal colonies. A five year survey: 1984-1988. Z Versuchstierkd 33, 29-35 KÜHN, R., J. LÖHLER, D. RENNICK, K. RAJEWSKY u. W. MÜLLER (1993): Interleukin-10-deficient mice develop chronic enterocolitis. Cell 75, 263-274

Literaturverzeichnis
120
KULLBERG, M. C., J. M. WARD, P. L. GORELICK, P. CASPAR, S. HIENY, A. CHEEVER, D. JANKOVIC u. A. SHER (1998): Helicobacter hepaticus triggers colitis in specific-pathogen-free interleukin-10 (IL-10)-deficient mice through an IL-12- and gamma interferon-dependent mechanism. Infect Immun 66, 5157-5166 LENCIONI, K. C., A. SEAMONS, P. M. TREUTING, L. MAGGIO-PRICE u. T. BRABB (2008): Murine norovirus: an intercurrent variable in a mouse model of bacteria-induced inflammatory bowel disease. Comp Med 58, 522-533 LÖHLER, J. K., D. RENNICK, K. RAJEWSKY u. W. MÜLLER (1995): Interleukin-10-deficient mice: a model of chronic mucosal inflammation. Tygat G BJ, van Deventer S, Med. Inflammatory bowel disease. Dordrecht: Klüwer Academic publishers 401-407 MADSEN, K. L., J. S. DOYLE, L. D. JEWELL, M. M. TAVERNINI u. R. N. FEDORAK (1999): Lactobacillus species prevents colitis in interleukin 10 gene-deficient mice. Gastroenterology 116, 1107-1114 MADSEN, K. L., J. S. DOYLE, M. M. TAVERNINI, L. D. JEWELL, R. P. RENNIE u. R. N. FEDORAK (2000): Antibiotic therapy attenuates colitis in interleukin 10 gene-deficient mice. Gastroenterology 118, 1094-1105 MÄHLER, M. u. W. KÖHL (2009): A serological survey to evaluate contemporary prevalence of viral agents and Mycoplasma pulmonis in laboratory mice and rats in western Europe. Lab Anim (NY) 38, 161-165 MÄHLER, M. u. E. H. LEITER (2002): Genetic and environmental context determines the course of colitis developing in IL-10-deficient mice. Inflamm Bowel Dis 8, 347-355 MANUEL, C. A., C. C. HSU, L. K. RILEY u. R. S. LIVINGSTON (2008): Soiled-bedding sentinel detection of murine norovirus 4. J Am Assoc Lab Anim Sci 47, 31-36 MATSUMOTO, S., Y. OKABE, H. SETOYAMA, K. TAKAYAMA, J. OHTSUKA, H. FUNAHASHI, A. IMAOKA, Y. OKADA u. Y. UMESAKI (1998): Inflammatory bowel disease-like enteritis and caecitis in a senescence accelerated mouse P1/Yit strain. Gut 43, 71-78

Literaturverzeichnis
121
MCCARTNEY, S. A., L. B. THACKRAY, L. GITLIN, S. GILFILLAN, H. W. VIRGIN u. M. COLONNA (2008): MDA-5 recognition of a murine norovirus. PLoS Pathog 4, e1000108 MOOLENBEEK, C. u. E. J. RUITENBERG (1981): The "Swiss roll": a simple technique for histological studies of the rodent intestine. Lab Anim 15, 57-59 MOORE, K. W., A. O'GARRA, R. DE WAAL MALEFYT, P. VIEIRA u. T. R. MOSMANN (1993): Interleukin-10. Annu Rev Immunol 11, 165-190 MOST, C. (2001) Untersuchungen zur genetischen Disposition für chronische Darmentzündungen bei der IL10-defizienten Maus. Hannover, Tierärztliche Hochschule Hannover, Institut für Versuchstierkunde der Medizinischen Hochschule Hannover, Diss. MÜLLER, B. (2009) Molekulare Charakterisierung muriner Noroviren – phylogenetische und antigene Eigenschaften. Berlin, Mathematisch-Naturwissenschaftliche Fakultät I der Humboldt-Universität zu Berlin, Diss. MÜLLER, B., U. KLEMM, A. MAS MARQUES u. E. SCHREIER (2007): Genetic diversity and recombination of murine noroviruses in immunocompromised mice. Arch Virol 152, 1709-1719 MUMPHREY, S. M., H. CHANGOTRA, T. N. MOORE, E. R. HEIMANN-NICHOLS, C. E. WOBUS, M. J. REILLY, M. MOGHADAMFALAHI, D. SHUKLA u. S. M. KARST (2007): Murine norovirus 1 infection is associated with histopathological changes in immunocompetent hosts, but clinical disease is prevented by STAT1-dependent interferon responses. J Virol 81, 3251-3263 MURRAY, P. J. (2006): Understanding and exploiting the endogenous interleukin-10/STAT3-mediated anti-inflammatory response. Curr Opin Pharmacol 6, 379-386 NEUMANN, D., T. TSCHERNIG, D. POPA, A. SCHMIEDL, G. PEREZ DE LEMA, K. RESCH u. M. U. MARTIN (2006):

Literaturverzeichnis
122
Injection of IL-12- and IL-18-encoding plasmids ameliorates the autoimmune pathology of MRL/Mp-Tnfrsf6lpr mice: synergistic effect on autoimmune symptoms. Int Immunol 18, 1779-1787 NICKLAS, W., P. BANEUX, R. BOOT, T. DECELLE, A. A. DEENY, M. FUMANELLI u. B. ILLGEN-WILCKE (2002): Recommendations for the health monitoring of rodent and rabbit colonies in breeding and experimental units. Lab Anim 36, 20-42 OKAYASU, I., S. HATAKEYAMA, M. YAMADA, T. OHKUSA, Y. INAGAKI u. R. NAKAYA (1990): A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 98, 694-702 PAIK, J., Y. FIERCE, R. DRIVDAHL, P. M. TREUTING, A. SEAMONS, T. BRABB u. L. MAGGIO-PRICE (2010): Effects of murine norovirus infection on a mouse model of diet-induced obesity and insulin resistance. Comp Med 60, 189-195 PANWALA, C. M., J. C. JONES u. J. L. VINEY (1998): A novel model of inflammatory bowel disease: mice deficient for the multiple drug resistance gene, Mdr1a, spontaneously develop colitis. J Immunol 161, 5733-5744 PARK, G. W., L. BARCLAY, D. MACINGA, D. CHARBONNEAU, C. A. PETTIGREW u. J. VINJE (2010): Comparative Efficacy of Seven Hand Sanitizers against Murine Norovirus, Feline Calicivirus, and GII.4 Norovirus. J Food Prot 73, 2232-2238 PERDUE, K. A., K. Y. GREEN, M. COPELAND, E. BARRON, M. MANDEL, L. J. FAUCETTE, E. M. WILLIAMS, S. V. SOSNOVTSEV, W. R. ELKINS u. J. M. WARD (2007): Naturally occurring murine norovirus infection in a large research institution. J Am Assoc Lab Anim Sci 46, 39-45 PERRY, J. W., S. TAUBE u. C. E. WOBUS (2009): Murine norovirus-1 entry into permissive macrophages and dendritic cells is pH-independent. Virus Res 143, 125-129 PERRY, J. W. u. C. E. WOBUS (2010): Endocytosis of murine norovirus 1 into murine macrophages is dependent on dynamin II and cholesterol.

Literaturverzeichnis
123
J Virol 84, 6163-6176 POWRIE, F. (1995): T cells in inflammatory bowel disease: protective and pathogenic roles. Immunity 3, 171-174 POWRIE, F., M. W. LEACH, S. MAUZE, L. B. CADDLE u. R. L. COFFMAN (1993): Phenotypically distinct subsets of CD4+ T cells induce or protect from chronic intestinal inflammation in C. B-17 scid mice. Int Immunol 5, 1461-1471 PRITCHETT-CORNING, K. R., J. COSENTINO u. C. B. CLIFFORD (2009): Contemporary prevalence of infectious agents in laboratory mice and rats. Lab Anim 43, 165-173 RASCHKE, W. C., S. BAIRD, P. RALPH u. I. NAKOINZ (1978): Functional macrophage cell lines transformed by Abelson leukemia virus. Cell 15, 261-267 RILEY, L. K., C. L. FRANKLIN, R. R. HOOK, JR. u. C. BESCH-WILLIFORD (1996): Identification of murine helicobacters by PCR and restriction enzyme analyses. J Clin Microbiol 34, 942-946 ROERS, A., L. SIEWE, E. STRITTMATTER, M. DECKERT, D. SCHLÜTER, W. STENZEL, A. D. GRUBER, T. KRIEG, K. RAJEWSKY u. W. MÜLLER (2004): T cell-specific inactivation of the interleukin 10 gene in mice results in enhanced T cell responses but normal innate responses to lipopolysaccharide or skin irritation. J Exp Med 200, 1289-1297 SADLACK, B., H. MERZ, H. SCHORLE, A. SCHIMPL, A. C. FELLER u. I. HORAK (1993): Ulcerative colitis-like disease in mice with a disrupted interleukin-2 gene. Cell 75, 253-261 SCHULTZ, M. u. R. B. SARTOR (2000): Probiotics and inflammatory bowel diseases. Am J Gastroenterol 95, S19-21 SELLON, R. K., S. TONKONOGY, M. SCHULTZ, L. A. DIELEMAN, W. GRENTHER, E. BALISH, D. M. RENNICK u. R. B. SARTOR (1998): Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10-deficient mice. Infect Immun 66, 5224-5231 SHAMES, B., J. G. FOX, F. DEWHIRST, L. YAN, Z. SHEN u. N. S. TAYLOR (1995): Identification of widespread Helicobacter hepaticus infection in feces in commercial mouse colonies by culture and PCR assay.

Literaturverzeichnis
124
J Clin Microbiol 33, 2968-2972 STUART, A. D. u. T. D. BROWN (2006): Entry of feline calicivirus is dependent on clathrin-mediated endocytosis and acidification in endosomes. J Virol 80, 7500-7509 TAKEDA, K., B. E. CLAUSEN, T. KAISHO, T. TSUJIMURA, N. TERADA, I. FORSTER u. S. AKIRA (1999): Enhanced Th1 activity and development of chronic enterocolitis in mice devoid of Stat3 in macrophages and neutrophils. Immunity 10, 39-49 THACKRAY, L. B., C. E. WOBUS, K. A. CHACHU, B. LIU, E. R. ALEGRE, K. S. HENDERSON, S. T. KELLEY u. H. W. T. VIRGIN (2007): Murine noroviruses comprising a single genogroup exhibit biological diversity despite limited sequence divergence. J Virol 81, 10460-10473 TORRES, M. I. u. A. RIOS (2008): Current view of the immunopathogenesis in inflammatory bowel disease and its implications for therapy. World journal of gastroenterology : WJG 14, 1972-1980 WARD, J. M., C. E. WOBUS, L. B. THACKRAY, C. R. EREXSON, L. J. FAUCETTE, G. BELLIOT, E. L. BARRON, S. V. SOSNOVTSEV u. K. Y. GREEN (2006): Pathology of immunodeficient mice with naturally occurring murine norovirus infection. Toxicol Pathol 34, 708-715 WEI, J., Y. JIN, T. SIMS u. K. E. KNIEL (2010): Manure- and biosolids-resident murine norovirus 1 attachment to and internalization by Romaine lettuce. Appl Environ Microbiol 76, 578-583 WEI, J., Y. JIN, T. SIMS u. K. E. KNIEL (2011): Internalization of Murine Norovirus 1 by Lactuca sativa during Irrigation. Appl Environ Microbiol 77, 2508-2512 WOBUS, C. E., S. M. KARST, L. B. THACKRAY, K. O. CHANG, S. V. SOSNOVTSEV, G. BELLIOT, A. KRUG, J. M. MACKENZIE, K. Y. GREEN u. H. W. VIRGIN (2004): Replication of Norovirus in cell culture reveals a tropism for dendritic cells and macrophages. PLoS Biol 2, e432

Literaturverzeichnis
125
YANG, H., J. D. OHMEN, Y. MA, L. G. BENTLEY, S. R. TARGAN, N. FISCHEL-GHODSIAN u. J. I. ROTTER (1999): Additional evidence of linkage between Crohn's disease and a putative locus on chromosome 12. Genet Med 1, 194-198 YEN, D., J. CHEUNG, H. SCHEERENS, F. POULET, T. MCCLANAHAN, B. MCKENZIE, M. A. KLEINSCHEK, A. OWYANG, J. MATTSON, W. BLUMENSCHEIN, E. MURPHY, M. SATHE, D. J. CUA, R. A. KASTELEIN u. D. RENNICK (2006): IL-23 is essential for T cell-mediated colitis and promotes inflammation via IL-17 and IL-6. J Clin Invest 116, 1310-1316 YEOM, S. C., S. A. YU, E. Y. CHOI, B. C. LEE u. W. J. LEE (2009): Prevalence of Helicobacter hepaticus, murine norovirus, and Pneumocystis carinii and eradication efficacy of cross-fostering in genetically engineered mice. Exp Anim 58, 497-504 ZSCHEMISCH, N.-H. (2004) Untersuchungen zur Rolle von Cyclin E1 in der Zellzykluskontrolle muriner Hepatozyten. Hannover, Fachbereich Chemie der Universität Hannover, Diss.

Abbildungsverzeichnis
126
Abbildungsverzeichnis
Abbildung 1: Schema des MNV-Genoms; unterhalb der Leseraster sind die
verschiedenen Proteinprodukte mit ihrem Gewicht in Kilodalton aufgeführt; N, N
Terminus; 3A, NTPase 3A-like; P, Vpg Protease; RdRp, RNA dependent RNA
Polymerase; VP1, VP2 Hauptkapsidproteine (HENDERSON 2008). .................. 3
Abbildung 2: Elektronenmikroskopische Aufnahme von MNV-1 (markiert durch
schwarze Pfeile) (HSU et al. 2005). .................................................................... 4
Abbildung 3: Dreidimensionale Rekonstruktion anhand cryoelektronen-
mikroskopischer Aufnahmen von humanem Norovirus (NV, virus-like particles),
murinem Norovirus (MNV) und dem Virus der hämorrhagischen Erkrankung
beim Kaninchen (RHDV, virus-like particles) im Vergleich (KATPALLY et al.
2010). .................................................................................................................. 5
Abbildung 4: Pathogenese der CED bei IL10-defizienten Mäusen, modifiziert nach
Powrie (POWRIE 1995) und Elson (ELSON 1999). .......................................... 16
Abbildung 5: Vergleich verschiedener Primerpaare im Gradienten. ......................... 54
Abbildung 6: Vergleich der Primerpaare 5580f/5671r und 5205f/5469r mit 5
verschiedenen Proben, die aus Organen und Kot isoliert wurden. .................... 54
Abbildung 7: Vergleich der Primerpaare 5205f/5469r und 5004f/5219r. ................... 55
Abbildung 8: Verwandschaft vom isolierten MNV-Stamm Hannover1/2007/DEU
erstellt durch BLAST (ALTSCHUL et al. 1990). ................................................. 57
Abbildung 9: TCID50-Test; der Indikator des Mediums zeigt lebende Zellen (gelb) an;
zweites Bild nach der Anfärbung mit Kristallviolett (lebende Zellen stark lila). .. 58
Abbildung 10: MNV-positives Mausserum (+++) in der Immunfluoreszenz. MNV-
infizierte Zellen sind grüngelblich markiert. ........................................................ 59
Abbildung 11: Kissenzentrifugation; Bande nach der Gradientenzentrifugation. ...... 60
Abbildung 12: Beziehung zwischen Extinktionswert und Verdünnung mit
Regressionsgerade. .......................................................................................... 63
Abbildung 13: Prozentuale Verteilung der Einzelmessungen von ausgewählten in der
IFA positiven (n=76) und negativen (n=68) Mäuseseren. .................................. 65

Abbildungsverzeichnis
127
Abbildung 14: Prozentuale Verteilung der Einzel-OD-Wertbestimmung von 205 in der
IFA negativen Mäuseseren mit Hilfe des ELISA. ............................................... 66
Abbildung 15: Elektronenmikroskopische Aufnahmen von MNV-Virionen (durch
schwarze Pfeile gekennzeichnet). ..................................................................... 67
Abbildung 16: Infektionsversuch 1, Mittelwert und SEM der IFNγ-Sekretion der in
vitro stimulierten MLK-Zellen (Proben pro Stamm, Geschlecht und Zeitpunkt
gepoolt). ............................................................................................................ 72
Abbildung 17 Infektionsversuch 2, Mittelwert und SEM der INFγ-Sekretion der in vitro
stimulierten MLK-Zellen (Einzeltierproben, keine Unterscheidung nach
Geschlecht) ....................................................................................................... 74
Abbildung 18: Infektionsversuch 3, Mittelwert und SEM der INFγ-Sekretion der in
vitro stimulierten MLK-Zellen (Einzeltierproben, keine Unterscheidung nach
Geschlecht). ...................................................................................................... 75
Abbildung 19: Infektionsversuch 4, Mittelwert und SEM der INFγ-Sekretion der in
vitro stimulierten MLK-Zellen (Einzeltierproben, keine Unterscheidung nach
Geschlecht). ...................................................................................................... 76
Abbildung 20: Infektionsversuch 5, Mittelwert und SEM der INFγ-Sekretion der in
vitro stimulierten MLK-Zellen (Einzeltierproben, keine Unterscheidung nach
Geschlecht). ...................................................................................................... 77
Abbildung 21: Infektionsversuch 5, Mittelwert und SEM des histologischen Score
(Jackson) von Zäkum und Kolon bei der Kontrollgruppe (Helico mit H. hepaticus
infiziert) und der Doppelinfektionsgruppe (MNV+Helico mit MNV und H.
hepaticus infiziert). ............................................................................................ 80
Abbildung 22: Kolonabschnitt einer C3-Il10-/--Maus 4 Wochen p.i. mit MNV (A-C) und
die zugehörige Kontrolle (D-F); HE-Färbung, der schwarze Balken entspricht
100µm. Im Vergleich zur Kontrolle ist eine milde Hyperplasie des Epithels, ein
gemischtzelliges Infiltrat in Mukosa und Submukosa sowie ein geringgradiges
Ödem festzustellen. ........................................................................................... 81
Abbildung 23: Kolonabschnitt einer keimfreien C3-Il10-/--Maus 4 Wochen p.i. mit MNV
(A-D) und die zugehörige Kontrolle (E-H); HE-Färbung, der schwarze Balken
entspricht 100µm. Im Vergleich zur Kontrolle unterscheidet sich die infizierte

Abbildungsverzeichnis
128
Maus nur durch eine leicht vermehrte Anzahl mononukleärer Zellen. Weitere
Entzündungsreaktionen sind nicht feststellbar. ................................................. 82
Abbildung 24: Kolonabschnitt einer B6-Il10-/--Maus infiziert mit MNV und H. hepaticus
(A-D) und die zugehörige Kontrolle (E-H), nur infiziert mit H. hepaticus; HE-
Färbung, der schwarze Balken entspricht 100µm. Doppelinfektion und Kontrolle
weisen eine vergleichbare mittel- bis hochgradige transmurale, gemischtzellige
Enteritis mit prominenter Hyperplasie der Enterozyten auf. ............................... 83

Tabellenverzeichnis
129
Tabellenverzeichnis
Tabelle 1: Interaktion verschiedener Umweltfaktoren auf das CED-Modell der IL10-
defizienten Maus ............................................................................................... 14
Tabelle 2: Ausprägung der Kolitis bei IL10-defizienten Inzuchtstämmen ................. 14
Tabelle 3: Verwendete Mausstämme und ihre Abkürzung ....................................... 18
Tabelle 4: Zusammensetzung von Altromin 1324 TPF ............................................. 19
Tabelle 5: Primer zur Detektion von MNV ................................................................ 22
Tabelle 6: Primer zur Klonierung von MNV .............................................................. 23
Tabelle 7: Primer zur Sequenzierung von MNV ........................................................ 23
Tabelle 8: Primer zur Kontrolle der RNA-Umschreibung zu cDNA ........................... 23
Tabelle 9: Primer zur Detektion von Helicobacter (H.) hepaticus ............................. 24
Tabelle 10: Primmerpaare zum Nachweis von MNV mit Hilfe des Quiagen-Taq PCR
Core-Kit (Primersequenzen siehe S. 22f.) ......................................................... 34
Tabelle 11: Übersichtsdiagramm der Infektionsversuche ......................................... 46
Tabelle 12: Überblick über die Verbreitung von MNV im ZTL im Jahr 2008 ............. 68
Tabelle 13: Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 1 mit Hilfe einer einfaktoriellen ANOVA für die 4 Gruppen:
Kontrolle, 2 Wochen p.i., 4 Wochen p.i., 8 Wochen p.i. ..................................... 71
Tabelle 14: Posthoc-Analyse zum Vergleich der Gruppen von Infektionsversuch 1
untereinander (Bonferroni´s Multipler Vergleichstest) ....................................... 72
Tabelle 15: Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 2 mit Hilfe einer einfaktoriellen ANOVA für die 3 Gruppen:
Kontrolle, 2 Wochen p.i., 4 Wochen p.i.............................................................. 73
Tabelle 16: Posthoc-Analyse zum Vergleich der Gruppen von Infektionsversuch 2
untereinander (Bonferroni´s Multipler Vergleichstest) ....................................... 73
Tabelle 17: Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 3 mit einem ungepaarten t-Test ............................................ 75
Tabelle 18: Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 4 mit einem ungepaarten t-Test ............................................ 76

Tabellenverzeichnis
130
Tabelle 19 Statistische Auswertung der Lymphozytenstimulation von
Infektionsversuch 5 mit einem ungepaarten t-Test ............................................ 77
Tabelle 20: MNV-Nachweis via PCR (positiv getestet / insgesamt getestet) ............ 78
Tabelle 21: H. hepaticus-Nachweis via PCR (positiv getestet / insgesamt getestet) 79
Tabelle 22: Statistische Auswertung der histologischen Scorewerte von
Infektionsversuch 5 mit einem ungepaarten t-Test ............................................ 80

Erklärung
131
Erklärung
Hiermit erkläre ich, dass ich die Disseration
„Charakterisierung eines murinen Norovirus – Auswirkungen des Virus auf die
experimentelle chronisch entzündliche Darmerkrankung bei der IL10-defizienten
Maus“
selbständig verfasst habe.
Bei der Anfertigung wurden folgende Hilfen Dritter in Anspruch genommen:
Die Sequenzierung von MNV wurde von der Firma MWG Biotech AG durchgeführt.
Die elektronenmikroskopischen Aufnahmen erfolgten im Zentrallabor
Elektronenmikroskopie der MHH mit freundlicher Unterstützung von Prof. Dr.
Ungewickell, Gerhard Preiß und Jutta Milzer (Institut für Virologie, MHH). Die
Messung der IFNγ-Sekretion erfolgte durch die Arbeitsgruppe PD Dr. Neumann im
Institut für Pharmakologie der MHH.
Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten
(Promotionsberater oder anderer Personen) in Anspruch genommen. Niemand hat
von mir unmittelbar oder mittelbar entgeltliche Leistungen für Arbeiten erhalten, die
im Zusammenhang mit dem Inhalt der vorgelegten Dissertation stehen.
Ich habe die Dissertation am Institut für Versuchstierkunde der Medizinischen
Hochschule Hannover angefertigt. Die Dissertation wurde bisher nicht für eine
Prüfung oder Promotion oder für einen ähnlichen Zweck zur Beurteilung eingereicht.
Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig
und der Wahrheit entsprechend gemacht habe.
Hannover, den
_____________________________
Marcel Achard

Danksagung
132
Danksagung
Mein Dank gilt
Herrn Prof. Dr. Hedrich MHH Herrn Prof. Bleich, Ph.D. MHH Herrn Prof. Dr. Mähler MHH, Firma Biodoc
am ZTL der MHH
Herrn Dr. Zschemisch Frau Smoczek Frau Wehling Frau Wiebe Frau Janus, Ph. D. Frau Büchler, Ph. D.
in der Firma Biodoc
Frau Dr. Köhl & dem gesamten Labor-Team
im Institut für Virologie der MHH
Frau Milzer
im Zentrallabor Elektronenmikroskopie der MHH
Herrn Prof. Ungewickell Herrn Preiß
im Institut für Pharmakologie der MHH
PD Dr. Neumann & seiner Arbeitsgruppe
im Institut für Virologie der TiHo Hannover
Prof. Dr. Haas Frau Winter, Ph.D.
und ganz besonders
meinen Eltern und Freunden, die mich unterstützt haben.