therapy of an animal model of human gastric cancer … amplification and/or ... (genex,...
TRANSCRIPT

[CANCER RESEARCH 52, 2771-2776, May 15, 1992]
Therapy of an Animal Model of Human Gastric Cancer Using a Combination ofAnti-erbB-2 Monoclonal Antibodies1
Philip G. Kasprzyk, Sun Uk Song, Pier Paolo Di Fiore, and C. Richter King2
Molecular Oncology, Inc., Gaithersburg, Maryland 20878 [P. G. K., S. V. S., C. R. K.J. and Laboratory of Cellular and Molecular Biology, National Cancer Institute,N1H, Bethesda, Maryland 20892 [P. P. DJ
ABSTRACT
Amplification and/or overexpression of the erhti-2 gene have beendemonstrated in 20-30% of adenocarcinomas of the breast, ovary, lung,and stomach and are associated with aggressive clinical course and poorprognosis. Interference with erbB-2 function by the use of monoclonalantibodies is a promising approach to the treatment of these diseases. Inthis study we demonstrate that a combination of two anti-erôB-2-specificantibodies inhibited the growth of human gastric tumor cells in vitro.This combination antibody therapy also inhibited the growth of humantumor cell lines growing as xenografts in nude mice and was able todramatically reduce established tumors. This is the first reported observation of tumor regression induced by anti-erAB-2 monoclonal antibodies.Treatment was not curative in that tumors regrew after 6 weeks. Treatment with either single antibody alone did not inhibit cell growth ortumor formation. Pulse chase and t>rosine kinase activity experimentswere used to investigate the activity of the erhH-2 gene productim>liS5"''" '). The formation of complexes by two antibodies was found to
interfere with receptor function and mimic some properties of a typicalreceptor ligand. Selective interference of the erbB-2 receptor by combination antibody therapy may be advantageous for the treatment of humancancers.
INTRODUCTION
Amplification and/or overexpression of the erbB-2 gene havebeen shown in 20-30% of adenocarcinomas of the breast (1-5), ovary (3), lung (6), and stomach (7). Two lines of evidenceimplicate erbB-2 overexpression in the pathogenesis of humanneoplasia. First, overexpression has been linked with poorprognosis in breast (2-3, 8-11), ovarian (3, 12), stomach (13),and lung cancer (14), indicating that overexpression profoundlyeffects the cancer cell. Second, artificial overexpression of erbB-2 induces a transformed phenotype in NIH/3T3 fibroblasts (15,16) as well as in mammary epithelial cells (17), suggesting thatoverexpression can contribute directly to the development ofthe malignant phenotype.
Because of extensive homology between gpl85"*B"2 and the
epidermal growth factor receptor, it is widely assumed that theactivation of growth signal transduction might proceed throughsimilar mechanisms. One proposed mechanism involves receptor dimerization or oligomerization, which is thought to be animportant step in the activation of the epidermal growth factorreceptor intrinsic tyrosine kinase function (18, 19). In this studyinterference with receptor-receptor interactions was evaluatedas a potential therapeutic approach to the treatment of cancerswith erbB-2 overexpression. Previous studies have evaluatedthe use of single monoclonal antibodies directed against erbB-2 (20) and the related epidermal growth factor receptor protein(21) as potential therapeutic agents for the treatment of cancer.
Received 8/29/91; accepted 3/11/92.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by Grant 2 R44 CA50077-02 from the Department of Health andHuman Services.
2To whom requests for reprints should be addressed, at Molecular Oncology,
Inc., 19 Firstfield Road, Gaithersburg, MD 20878.
Our studies focused on the synergistic effect of a combinationof two anti-erAB-2 antibodies. We chose to evaluate combinations of monoclonal antibodies because binding to gpl85"*B~2
should induce complex lattices of highly constrained proteinon the cell surface. We reasoned that these lattices might alterreceptor function in a manner different from that of the bindingof single monoclonal antibodies that can be expected to inducereceptor dimerization. Our results show profound growth inhibition with the combination of two antibodies when comparedto single-antibody treatment.
MATERIALS AND METHODS
Monoclonal Antibodies. Mice were immunized using a membranepreparation of N/erbB-2 cells (NIH/3T3 cells engineered to overexpressthe human erbB-2 protein). Following tests of polyclonal antibodyresponse using immunoprecipitation, three fusions were conductedusing the myeloma cell line Ag8.653 and standard techniques. Thesefusions produced approximately 1500 hybridoma clones which wereeach screened using enzyme-linked immunosorbent assay. Membranesisolated from N/erbB-2 cells were bound to polystyrene plates, andculture medium was added to allow antibody-antigen interaction. 1mmunoglobulin binding was detected using a biotinylated goat anti-mouse antibody, streptavidin horseradish peroxidase, and o-phenyl-enediamine hydrochloride. Positive reacting hybridomas were pickedand counter-screened using membranes from wild-type NIH/3T3 membranes. The molecular specificity was confirmed by immunoprecipitation analysis. Five hybridomas were picked with anti-eréB-2-specifÃcreactivity and cloned by limiting dilution; two of these were designatedas e21 and e23 and were used in this study. Ascites was prepared byadministering injections of IO1'hybridoma cells to pristane-primed
mice. Antibodies were isolated in large amounts from ascites fluid andpurified by high-performance liquid chromatography with a Gamma-bind Ultra column (Genex, Gaithersburg, MD). SDS-PAGE3 was run
under nonreducing conditions using Coomassie blue staining with asingle band at M, 180,000 observed, indicating a >98% purified preparation. From 1 ml of ascites approximately 8-15 mg of antibody wereroutinely purified.
Cell Lines and Tissue Culture. The human gastric tumor cell lineused in these studies, N87, has been previously described (22) and wasroutinely subcultura! in RPMI 1640 supplemented with 10% fetalbovine serum. The cell lines SK-Br-3, MDA-MB-468, and MDA-MB-231 (breast) and SK-OV-3 (ovarian) were routinely subcultured inimproved minimal essential medium (IMEM) supplemented with 5%fetal bovine serum. Cultures were maintained in humidified incubatorsat 37°Cin an atmosphere of 5% CO2 and 95% air. Cells were tested
for Mycoplasma using a ribosomal RNA hybridization method (Gen-Probe, San Diego, CA).
Growth Inhibition Assays. A single cell suspension of 10,000 cells/well was plated in a serum-free defined media of RPMI 1640 containingbovine serum insulin (5 Mg/mO, human transferrin (10 ¿tg/ml),17-/3-estradiol (10 nM), sodium selenite (5 nM), and 10 mM 4-(2-hydroxy-ethyl)-l-piperazineethanesulfonic acid or in RPMI 1640 containing 2%fetal bovine serum. PBS, e21, e23, or a combination of e21 and e23(0.1, 1, 10 Mg/ml final concentration) was then added. The plates were
5The abbreviations used are: SDS-PAGE, sodium dodecyl sulfate-polyacryl-amide gel electrophoresis; PBS, phosphate-buffered saline; MTT, 3-(4-5-dime-thylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; cDNA, complementaryDNA.
2771
on April 20, 2017. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

erbB-2 THERAPY FOR GASTRIC CANCER
incubated for 5-7 days in a CO2 incubator with humidity at 37°C.Cell RESULTS
viability was monitored by one of two different methods. The first, theMTT assay (23), is based on the ability of live cells to reduce atetrazolium-based compound, MTT, to a purplish colored formazanproduct that can be measured spectrophotometrically. After 7 days, 50/il of MTT reagent (0.1 mg) were added and allowed to incubate for 4h at 37°C.Ninety % of the media was then removed, and the crystals
were solubilized in 0.175 ml dimethyl sulfoxide with absorbance measured at 540 nm in a Molecular Devices max kinetic microplate reader.The second method involves the cell number measurement in mono-layer cultures by crystal violet staining (24). Cells were plated as aboveand after 7 days cells were fixed by the addition of 20 ^1 of a 11%glutaraldehyde solution. After being shaken on a BélicoOrbital Shakerfor 15 min the plates were washed three times with deionized water.Plates were then air-dried and stained by the addition of 100 /<!of a0.1% solution of crystal violet dissolved in 200 mM borate, pH 6.0.After being shaken for 20 min at room temperature, excess dye wasremoved by extensive washing with deionized water, and the plateswere air-dried prior to solubilization in 100 /il of 10% acetic acid.Absorbance was measured at 590 nm in the microplate reader.
Antibody Specificity. Subconfluent SK-Br-3 monolayers were meta-bolically labeled with ["SJCys (specific activity, 1000 Ci/mmol). Total
cell proteins were immunoprecipitated with 10 n\ of the indicatedantibodies. The immune complexes were recovered by protein G-Aga-rose (Genex) and analyzed by SDS-PAGE on an 8-16% Tris-glycinegel. The gel was exposed to film at —¿�70°Covernight with an intensifying
screen.Western Blots. Cells or tumors were lysed in sample buffer which
contained 0.125 M Tris-HCl, 4% SDS, 0.002% bromophenol blue, and15% glycerol. Five % /3-mercaptoethanol was added after the proteinconcentration was determined. Samples (10 Mg total protein) wereboiled for 3 min, fractionated by SDS-PAGE on 8-16% Tris-Glycinegel (Novex, Encinitas, CA), and transferred to nitrocellulose. Detectionof gpl85"*B"2 was performed with a monoclonal antibody (E2-4001;
Molecular Oncology, Inc.) to the COOH-terminal portion of the
protein.Southern Blots. DNA was extracted from cell lines and human
placenta tissue using guanidine thiocyanate and cesium gradient cen-trifugation. DNA (15 ^g) was cleaved with restriction enzyme Hindlll,separated by electrophoresis on a 1% agarose gel, transferred to nitrocellulose, and probed with a radioactive erbB-2 cDNA probe as previously described (25). The cDNA probe corresponds to the entire erbB-2 protein coding region.
In Vivo Antitumor Assay. Tumor cells (5 x 106/mouse) were injected
s.c. into the flanks of BNX (beige, nude, xid) mice. The day after cellinoculation treatment was begun which consisted of four trial groups(3 mice/group), each given 0.2-ml i.p. treatment injections twice aweek. Tumor growth was monitored at least once a week and reportedas an average relative tumor volume. The effect of treatment after theformation of small tumors was also carried out. Cells were injectedusing the same treatment protocol as above except that the treatmentwas begun 4 days after cell injection instead of 1 day after. Animal carewas in accordance with institutional guidelines. Statistical analysis wascarried out using a SAS Computer Package (SAS Institute, Cary, NC).
gpl85"4i" Stability Assay. Subconfluent N87 cell monolayers werepulse-labeled l h with 20 /<Ci[35S]cysteine and then chased with 5 mM
Cys in the presence of antibody for 24 h. Total cellular protein wasimmunoprecipitated as described above using a monoclonal antibodydirected against the COOH terminus of gpl85"*B2 coupled to Sepha-rose and analyzed by SDS-PAGE. The gel was exposed to film at-70°Covernight with an intensifying screen.
Tyrosine Phosphorylation. Cells were plated as in the protein stabilityassay. After 1 h, cells were processed, and proteins were extracted insample buffer for electrophoresis as in the antibody specificity experiment. Following electrophoresis the proteins were electroblotted ontonitrocellulose paper and incubated with anti-phosphotyrosine IgG(monoclonal; Upstate Biotechnology, Inc.) and immunodetected using12!I-protein A. The gel was exposed to film at —¿�70"Covernight with an
intensifying screen.2772
Monoclonal antibodies directed against the extracellular domain of gpl85'r*B2 were prepared by immunizing mice using a
membrane preparation from NIH/3T3 cells engineered to ov-erexpress the human erbB-2 protein (N/erbB-2 cells). Two ofthese, designated e21 and e23, were used in this study. Bothantibodies specifically immunoprecipitated a single 35S-labeledprotein of molecular weight 185,000 from SK-Br-3 cells (abreast cancer cell line which overexpresses gpl85'r*B'2 protein)
(26) as shown in Fig. 1. No immunoprecipitation was detectedin cells which do not overexpress the gpl85"*B2 protein (e.g.,MDA-MB-468; data not shown).
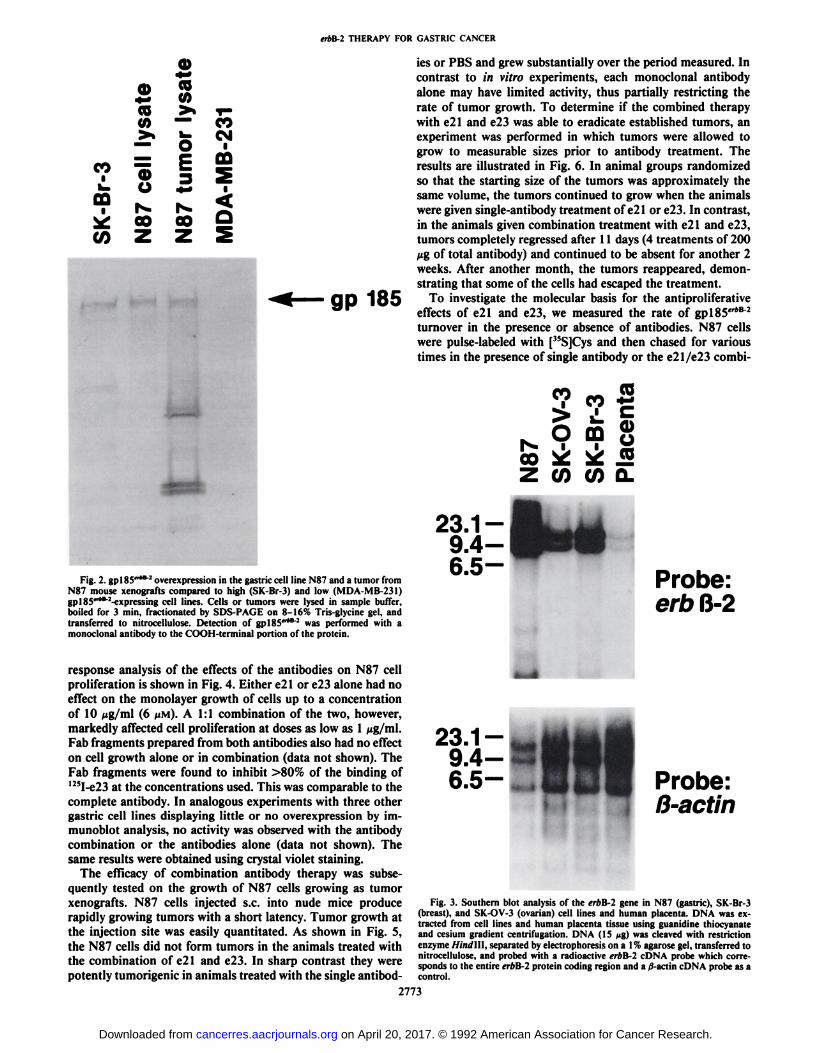
Since the erbB-2 oncogene is overexpressed frequently in atleast 20% of stomach cancers and these tumors have a poorclinical course, we investigated the gastric cell line N87, whichoverexpresses gpl85"*B 2 at high levels. An immunoblot of the
N87 cell line and a tumor formed by implantation of N87 cellsinto a nude mouse is shown in Fig. 2 compared to the breastcell lines SK-Br-3 (high level of gpl85"*B2 overexpression) andMDA-MB-231 (low level of gpl85"*B2 overexpression). Thelevels of erbB-2 gene amplification in N87, as shown in Fig. 3,surpassed those found in the well-characterized SK-Br-3 andSK-OV-3 cell lines (26).
The effect of these antibodies on growth was first studied invitro using a semiautomated colorimetrie MTT assay. A dose-
OC
CM0)
COCM0)
200- —¿�
97.4- —¿�
-gp185
il I69-
46-
30-
14.3-
Fig. 1. Specificity of monoclonal antibodies e21 and e23. Subconfluent SK-Br-3 monolayers were metabolically labeled with ["SJCys and immunoprecipitated with the indicated antibodies.
on April 20, 2017. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
crAB-2 THERAPY FOR GASTRIC CANCER
QQ 185
Fig. 2. gpl85"*B 2overexpression in the gastric cell line N87 and a tumor fromN87 mouse xenografts compared to high (SK-Br-3) and low (MDA-MB-231)gpl85"*t:-expressing cell lines. Cells or tumors were lysed in sample buffer,boiled for 3 min, fractionated by SDS-PAGE on 8-16% Tris-glycine gel, andtransferred to nitrocellulose. Detection of gplSS""" was performed with amonoclonal antibody to the COOH-terminal portion of the protein.
ies or PBS and grew substantially over the period measured. Incontrast to in vitro experiments, each monoclonal antibodyalone may have limited activity, thus partially restricting therate of tumor growth. To determine if the combined therapywith e21 and e23 was able to eradicate established tumors, anexperiment was performed in which tumors were allowed togrow to measurable sizes prior to antibody treatment. Theresults are illustrated in Fig. 6. In animal groups randomizedso that the starting size of the tumors was approximately thesame volume, the tumors continued to grow when the animalswere given single-antibody treatment of e21 or e23. In contrast,in the animals given combination treatment with e21 and e23,tumors completely regressed after 11 days (4 treatments of 200fig of total antibody) and continued to be absent for another 2weeks. After another month, the tumors reappeared, demonstrating that some of the cells had escaped the treatment.
To investigate the molecular basis for the antiproliferativeeffects of e21 and e23, we measured the rate of gplSS"*82
turnover in the presence or absence of antibodies. N87 cellswere pulse-labeled with p'SjCys and then chased for varioustimes in the presence of single antibody or the e21/e23 combi-
O DÛ
coco
23.19.46.5 Probe:
erb ß-2
response analysis of the effects of the antibodies on N87 cellproliferation is shown in Fig. 4. Either e21 or e23 alone had noeffect on the monolayer growth of cells up to a concentrationof 10 Mg/ml (6 fiM). A 1:1 combination of the two, however,markedly affected cell proliferation at doses as low as 1 ng/m\.Fab fragments prepared from both antibodies also had no effecton cell growth alone or in combination (data not shown). TheFab fragments were found to inhibit >80% of the binding of125I-e23at the concentrations used. This was comparable to the
complete antibody. In analogous experiments with three othergastric cell lines displaying little or no overexpression by immunoblot analysis, no activity was observed with the antibodycombination or the antibodies alone (data not shown). Thesame results were obtained using crystal violet staining.
The efficacy of combination antibody therapy was subsequently tested on the growth of N87 cells growing as tumorxenografts. N87 cells injected s.c. into nude mice producerapidly growing tumors with a short latency. Tumor growth atthe injection site was easily quantitated. As shown in Fig. 5,the N87 cells did not form tumors in the animals treated withthe combination of e21 and e23. In sharp contrast they werepotently tumorigenic in animals treated with the single antibod-
2773
Probe:ß-actin
Fig. 3. Southern blot analysis of the erbB-2 gene in N87 (gastric), SK-Br-3(breast), and SK-OV-3 (ovarian) cell lines and human placenta. DNA was extracted from cell lines and human placenta tissue using guanidine thiocyanateand cesium gradient centrifugation. DNA (15 *ig) was cleaved with restrictionenzyme Hind\\\, separated by electrophoresis on a 1% agarose gel, transferred tonitrocellulose, and probed with a radioactive erbB-2 cDNA probe which corresponds to the entire erbB-2 protein coding region and a 0-actin cDNA probe as acontrol.
on April 20, 2017. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

eròB-2 THERAPY FOR GASTRIC CANCER
50000 r
CD
O
30000 •¿�
-J3L^r-.p1PBS
0.1 1 10 0.1 1 10 0.1 1 10
e21 & e23 e21 e23
CONCENTRATION Utg/ml]
Fig. 4. EfTects of e21 and e23 on the growth of human N87 gastric tumor cellsin a monolayer MTT growth assay. A single cell suspension of 10,000 cells/wellwas plated in a defined medium consisting of RPMI 1640 supplemented withinsulin, human transferrin, 17-/3-estradiol, sodium selenite, and 4-(2-hydroxy-ethyl)-l-piperazineethanesulfonic acid buffer. Antibodies and combinations werethen added. After 7 days, MTT reagent was added and allowed to incubate for 4h at 37'C. Ninety % of the media was then removed, and the crystals were
solubilized in dimethyl sulfoxide. Absorbance was measured at 540 nm withresults representing the average of eight wells, with SD noted. Under the conditions used, the cell number is directly proportional to MTT reduction.
1.0 2.0 3.0 4.0 5.0 6.0 7.0
Weeks following introduction of tumor cells
Fig. S. Effects of treatment with e21 (O), e23 (A), a combination of e21 ande23 (•),or PBS (D) on the growth of N87 tumor xenografts in BNX mice. Tumorcells (5 x 10" mouse) were injected s.c. into the flanks of BNX (beige, nude, xid)
mice. Treatment begun on day 1 consisted of four trial groups (3 mice/group),each given 0.2-ml i.p. injections twice a week of either PBS (D), 200 ;<upurifiede21 (O), 200 I/Kpurified e23 (A), or a mixture of 100 ng purified e21 and 100 UKof purified e23 (•)for 3 weeks. Tumor growth is reported as an average relativetumor volume ±SE. At day 51 measurement of the tumor volume betweendifferent treatment groups was determined to be statistically significant (P <0.025) using analysis of variance. Two repeats of the experiment gave the sameresults.
nation. The results of a 24-h chase are shown in Fig. 7. Theantibody combination induced rapid degradation ofgp!85fr*82, while the individual antibody treatment had little
or no effect.A possible explanation for the decreased half-life of
gplSS"*82 is that the combined treatment with e21 and e23acts by constraining gpl85"*B"2 into an activated conformation,
thus mimicking an agonist ligand. This kind of mechanism hasbeen postulated for other agonistic antibodies (20). If the e21/
e23 combination mimics the ligand, then treatment shouldresult in increased gpl85cr*B2 autophosphorylation upon e21/
e23 combined treatment. We used antiphosphotyrosine immu-noblots to test this hypothesis. As shown in Fig. 8, increases inthe tyrosine phosphors hi ion of gpl85"*B 2from N87 cells were
observed 15 min and l h after the addition of the singleantibodies or the antibodies in combination. (The same resultswere observed at 30 min and 2 h; data not shown.) This suggeststhat activation of tyrosine kinase activity may be necessary butis probably not sufficient for growth inhibition.
120
0 4 7 11 14 19
Days after therapy started
Fig. 6. Effect of treatment after the formation of small tumors. Cells wereinjected using the same treatment protocol as in Fig. 5 except that the treatmentwas begun 4 days after cell injection instead of 1 day after.
CM
+CO i-
0) 0) 0) 0) 0)S •¿�* •¿�« •¿�J2 n ra (0 to (03 .C .C .C £i C.u. O O O O O
0)V)(O
"t
:t—¿�gp 185
200-
97.4-
69-46-
30-
14.3-
Fig. 7. Effect of antibody binding on erbB-2 protein turnover. SubconfluentN87 cell monolayers were pulse-labeled l h with 20 »Ci["SJcysteine and thenchased with 5 m\i Cys in the presence of e21 alone, e23 alone, or a 1:1combination of e21 and e23 (10 fig/ml) for 24 h. Total cellular protein wasimmunoprecipitated as described in Fig. 1 using a monoclonal antibody directedagainst the COOH terminus of gplSS"*""2 coupled to Sepharose and analyzed bySDS-PAGE. The gel was exposed to film at —¿�70'Covernight with an intensifying
screen.
2774
on April 20, 2017. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

erbñ-2THERAPY FOR GASTRIC CANCER
COCM
CMCM CMO 0) 0 0
n« -gpi85
15 min
~W -gp185
1 hr
Fig. 8. Measurement of tyrosine phosphorylalion of gplSS"*8"' after incubation
with antibody combination. Cells were plated as in Fig. 7. After l h cells wereprocessed as in Fig. 1. The proteins were electroblotted onto nitrocellulose paperand incubated with anti-phosphotyrosine IgG (monoclonal; Upstate Biotechnology, Inc.) and immunodetected using 1!*I-protein A. The gel was exposed to filmat —¿�70"C overnight with an intensifying screen.
DISCUSSION
Our results are the first reported observation of tumor regression induced by anti-erftB-2 monoclonal antibodies. While complete regression was observed, the treatment was not curativein that tumors grew back in 6 weeks. Other investigators havefocused their studies on the use of single anti-erbB-2 (or HER-2) antibodies against the growth of human breast cancer cellsin vitro and against the formation of tumors by fibroblaststransformed by the overexpression of human erbB-2 (or HER-2). Previous studies have shown that two anti-neu antibodiescan inhibit the growth of tumors by murine cells transformedby the mutationally activated neu oncogene (27). This effect isalso seen with the inhibition of leukemic tumor cell growthusing anti-transferrin monoclonal antibodies (28). In a recentpublication by Hancock et al. (29), the authors studied thesynergistic effect of an antibody against c-erbB-2 plus cis-diaminedichloroplatinum.
Our results suggest that each antibody singly can stimulategpl85"*B"2 autophosphorylation; the combined antibody treatment also stimulates gpl85'r*B2 autophosphorylation but actsadditionally by altering the turnover rate of gpl85"*B2. Since
combination antibody treatment probably acts primarily tospatially constrain the erbB-2 molecule, our results raise theintriguing possibility that two different receptor-receptor interactions might take place during the response of gpl85"*B2 tonatural Hgand. One receptor-receptor conformation is requiredfor tyrosine kinase activation, which might be mimicked bycertain monoclonal antibodies that activate erbB-2 autophosphorylation (20, 30) or e21 and e23. Such a monoclonal anti-
erbB-2 antibody has been proposed as a therapeutic. Followingkinase activation, another sterically distinct receptor-receptorinteraction may then facilitate internalization and down-regulation. This alignment may be induced when the erbB-2 proteinmolecule is constrained by binding a combination of anti-ereB-2 e21 and e23. This model is supported by the finding that theFab fragments do not show antigrowth activity in combination,nor do other combinations of nnti-erbB-2 monoclonal antibodies that we have tested. Alternatively, the combined e21/e23treatment may induce a nonphysiological pattern of receptoroligomerization that shunts gplSS"*8"2 to the down-regulation
pathway.Regardless of the mechanism, our results demonstrate that a
combination of anti-receptor antibodies leads to different andmore potent antitumor activities than single antibodies, andtherefore, combination antibody therapy may be a useful approach to the treatment of human malignancies overexpressinggplSS"*8 2.This approach may be particularly important in the
treatment of gastric cancer, a disease which responds poorly tocurrent systemic chemotherapies.
ACKNOWLEDGMENTS
We are grateful to Adi Gazdar for providing the N87 human gastriccell line. We thank Teresa Clayton, John Henry, and Laura Porter forexcellent technical assistance and Michael Herman and Nancy Turnerfor critical reading of the manuscript.
REFERENCES
1. King, C. R., Kraus, M. H., and Aaronson, S. A. Amplification of a novel v-iT/ill related gene in a human mammary carcinoma. Science (WashingtonDC), 229: 974-976, 1985.
2. Slamon, D. J., Clark, G. M., Wong, S. G., Levin, W. J., Ullrich, A., andMcGuire, W. L. Human breast cancer: correlation of relapse and survivalwith amplification of the HER-2/neu oncogene. Science (Washington DC),235: 177-182, 1987.
3. Slamon, D. J., Godolophin, W., Jones, L. A., Holt, J. A., Wong, S. G.,Keith, D. E., Levin, W. J., Stuart, S. G., Udove, J., Ullrich, A., and Press,M. F. Studies of the Her-2/neu proto-oncogene in human breast and ovariancancer. Science (Washington DC), 244: 707-712, 1989.
4. Yokota, J., Yamamoto, T., Toyoshima, K., Sugimura, T., Yamamoto, T.,Terada, M., Battifora, H., and Cline, M. J. Amplification of c-erbB-2 oncogene in human adenocarcinomas in vivo. Lancet, /: 765-767, 1986.
5. King, C. R., Swain, S. M., Porter, L., Steinberg, S. M., Lippman, M. E., andGelman, E. P. Heterogenous expression of erbB-2 messenger RNA in humanbreast cancer. Cancer Res., 49:4185-4191, 1989.
6. Schneider, P. M., Hung, M-C, Chiocca, S. M., Manning, J., Zhao, X., Fang,H., and Roth, J. A. Differential expression of the erbB-2 gene in humansmall cell and non-small cell lung cancer. Cancer Res., 49:4968-4971, 1989.
7. Park, J., Rhim, J. S., Park, S., Kimm, S., and Kraus, M. Amplification,overexpression and rearrangement of the erbB-2 protooncogene in primaryhuman stomach carcinomas. Cancer Res., 49: 6605-6609, 1989.
8. Wright, C, Angus, B., Nicholson, S., Sainsbury, J. R., Cairns, J., Gullick,W. J., Kelly, P., Harris, A. L., and Home, C. H. Expression of c-erbB-2oncoprotein: a prognostic indicator in human breast cancer. Cancer Res., 49:2087-2090, 1989.
9. Tandon, A. K., Clark, G. M., Chamness, G. C., Ullrich, A., and McGuire,W. L. Her-2/neu oncogene protein and prognosis in breast cancer. J. Clin.Oncol., 7: 1120-1128, 1989.
10. Paik, S., Hazan, R., Fisher, E. R., Sass, R. E., Fisher, B., Redmond, C.,Schlessinger, J., Lippman, M. E., and King, C. R. Pathological findings fromthe National Surgical Adjuvant Breast and Bowel Project: prognostic significance of erbB-2 protein overexpression in primary breast cancer. J. Clin.Oncol.,«: 103-112, 1990.
11. Borg, A., Tandon, A. K., Sigurdsson, H.. Clark, G. M., Perno, M., Fuqua,S. A. W., Killander, D., and McGuire, W. L. Her-2/ncu amplification predictspoor survival in node-positive breast cancer. Cancer Res., 50: 4332-4337,1990.
12. Berviinek, A., Kamel, A., Whitaker, R., Kerns, B., Oil, G., Kinney. R., Soper,J. T., Dodge, R., Clarke-Pearson, D. L., Marks, P., McKenzie, S., Yin, S.,and Bast, R. C. Overexpression of HER-2/neu is associated with poor survivalin advanced epithelial ovarian cancer. Cancer Res., 50: 4087-4091, 1990.
13. Yonemura, Y., Ninoomiya, I., Yamaguchi, A., Fushida, S., Kimura, H.,Ohoyama, S., Miyazaki, I., Endou, Y., Tanaka, M., and Sasaki, T. Evaluationof immunoreactivity for erbB-2 protein as a marker of poor short term
2775
on April 20, 2017. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ÃrAB-2THERAPY FOR GASTRIC CANCER
prognosis in gastric cancer. Cancer Res., SI: 1034-1038, 1991.14. Kern, J. A., Schwartz, D. A., Nordberg, J. E., Weiner, D. B., Greene, M. I.,
Torney, L., and Robinson. R. A. p 185""1expression in human lung adeno-carcinomas predicts shortened survival. Cancer Res., 50: 5184-5191, 1990.
15. Di Fiore, P. P., Pierce, J. H., Kraus, M. H., Segallo, O., King, C. R., andAaronson, S. A. erbB-2 is a potenl oncogene when overexpressed in NIH/3T3 cells. Science (Washington DC), 237: 178-182, 1986.
16. llml/i.ik. R. M , Schlessinger, J., and Ullrich, A. Increased expression of theputative growth factor pl85 (HER2) causes transformation and tumorigen-esis of NIH3T3 cells. Pro. Nati. Acad. Sci. USA, 84: 7159-7163, 1987.
17. Pierce, J. H., Arnstein, P., DiMarco, E., Artrip, J., Kraus, M. H., Lonardo.F., DiFiore, P. P., and Aaronson, S. A. Oncogenic potential of erbB-2 inhuman mammary epithelial cells. Oncogene, 6: 1189-1194, 1991.
18. Yarden, Y.. and Ullrich, A. Molecular analysis of signal transduction bygrowth factors. Biochemistry, 27: 3114-3118, 1988.
19. Schlessinger, J. The epidermal growth factor receptor as a multifunctionalallosteric protein. Biochemistry, 27: 3119-3123, 1988.
20. Hudziak, R. M., Lewis, G. D., Winget. M., Fendly, B. M.. Shepard. H. M.,and Ullrich, A. pl85HER2 Monoclonal antibody has antiproliferative effects
in vitro and sensitizes human breast tumor cells to tumor necrosis factor.Mol. Cell. Biol., 9: 1165-1172, 1989.
21. Divgi, C. R., Welt, S., Kris, M., Real, F. X., Yeh, S. D. J.,, Gralla. R.,Merchant, H Schweighart, S., Unger, M., Larson, S. M., and Mendelsohn,J. Phase I and imaging trial of indium Ill-labeled anti-epidermal growthfactor receptor monoclonal antibody 225 in patients with squamous cell lungcarcinoma. J. Nail. Cancer Inst., S3: 97-104, 1991.
22. Park, J-G.. Frucht. H., LaRocca, R. V., Bliss, D. P., Kurita, Y., Chen, T. R.,Henslee, J. G., Trepel, J. B., Jensen, R. T., Johnson, B. E., Bang, Y-J., Kim,.11'.. and Gazdar, A. Characteristics of cell lines established from human
gastric carcinoma. Cancer Res., 50: 2773-2780, 1990.23. Mossman, T. Rapid colorimetrie assay for cellular growth and survival:
application to proliferation and cytotoxic assays. J. Immunol. Methods, 65:55-63, 1983.
24. Sibler, W. K., and Eppenberger. U. Quantification of cells cultured on 96-well plates. Anal. Biochem.. ¡82:16-19. 1989.
25. King, C. R., Swain, S. M., Porter. L.. Steinberg, S. M., Lippman, M. E., andGelman, E. P. Heterogeneous expression of erbB-2 messenger RNA in humanbreast cancer. Cancer Res., 49: 4185-4191, 1989.
26. Kraus, M. H., Popescu, N. C., Amsbaugh, S. C., and King, C. R. Overexpres-sion of the EGF receptor-related proto-oncogene iréB-2in human mammarytumor cell lines by different molecular mechanisms. EMBO J., 6: 605-610,1987.
27. Drebin, J. A., Link, V. C., and Greene, M. I. Monoclonal antibodies reactivewith distinct domains of the neu oncogene-encoded pl85 molecule exertsynergistic anti-tumor effects in vivo. Oncogene. 2: 273-277, 1988.
28. White, S., Taetle, R., Seligman. P. A.. Rutherford. M., and Trowbridge, I.S. Combinations of anti-transferrin receptor monoclonal antibodies inhibithuman tumor cell growth in vitro and in vivo: evidence for synergisticantiproliferalive effects. Cancer Res., 50: 6295-6301, 1990.
29. Hancock, M. C, Langton, B. C, Chan, T., Toy, P., Monahan, J. J., Mischak,R. P., and Shawver, L. K. A monoclonal antibody against the c-erbB-2protein enhances the cytotoxicity of m-diaminedichloroplatinum againsthuman breast and ovarian tumor cell lines. Cancer Res., 51: 4575-4580,1991.
30. Kumar, R., Shepard, H. M., and Mendelsohn, J. Regulation of phosphoryl-ation of the c-erbB-2/HEK2 gene product by a monoclonal antibody andserum growth factor(s) in human mammary carcinoma cells. Mol. Cell. Biol.,//: 979-986, 1991.
2776
on April 20, 2017. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1992;52:2771-2776. Cancer Res Philip G. Kasprzyk, Sun Uk Song, Pier Paolo Di Fiore, et al.
B-2 Monoclonal AntibodieserbCombination of Anti-Therapy of an Animal Model of Human Gastric Cancer Using a
Updated version
http://cancerres.aacrjournals.org/content/52/10/2771
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on April 20, 2017. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from