the rubro-olivo-cerebellar teaching circuit
TRANSCRIPT

Medical Hypotheses 5: 799-807, 1979
THE RLJBRO-OLIVO-CEREBELLAR TEACHING CIRCUIT
P. R. Kennedy, Dept. of Physiology, University of Western Ontario,
London, Ontario, Canada, N6A 5Cl.
ABSTRACT
The hypothesis proposes a neural teaching circuit, and invokes the
parvocellular Red Nucleus as the key nucleus in that circuit acting on the inferior olivary nucleus to maintain the efficiency of motor learning in
primates, while acknowledging the inferior olive to be the key nucleus by
which learning instructions go to Purkinje cells in the cerebellar cortex. Anatomical, physiological and pathological data is reviewed in an attempt to attribute a function to the evasive parvocellular Red Nucleus.
Key Words: Red Nucleus, Olivary Nucleus, Cerebellum, Dentate Nucleus,
Learning, Motor Skills, Movement Disorders, Tremor
HYPOTHESIS
The Rubro-olivary pathway, as part of a neural teaching circuit involving
the cerebellum, acts as a phasic control system on the Inferior Olivary
nucleus to maintain the efficiency of Motor Learning.
INTRODUCTION
Theories on motor learning were put forward in 1969 by David Marr who
proposed that inferior olivary impulses could teach Purkinje cells the ele- ments of motor movements. As he put it (2,23)..."Each olivary cell is
thought to be driven by an instruction from the cerebral cortex for that movement to take place. During learning, the cerebrum organizes the move-
ment, and in so doing, causes the appropriate olivary cells to fire in a
particular sequence. This causes the purkinje cells to learn the contexts within which their corresponding elemental movements are required, so that the next time such a context occurs the mossy fiber activity (i.e., all non- olivary inputs) stimulates the purkinje cell, which evokes the relevant movement." In other words, the inferior olive and its climbing fibres which synapse on purkinje cell dendrites, constitute the "Teaching Line" to the cerebellum.
799

It must be mentioned that the theory was preceded by Brindley's similar work (3,4) and paralleled by Albus (1) who envisaged unrequired purkinje cell responses being suppressed, rather than required purkinje cell responses being facilitated. Hebb in 1949 (15) had proposed learning by synaptic change, giving as an example climbing fibre synapses on purkinje cell den- drites. Marr revised his own theory in 1970 (2) when he suggested that "corrections can eventually be run by the cerebellar purkinje cells, without reference to the cerebrum. If a number of similar mossy fiber inputs have been learned and later an unlearned input is presented which is near enough to those which have been learned, then the purkinje cell may treat the new input as if it had been learned." Gilbert in 1974 (11) suggested a design to improve the speed and versatility of the cerebellum. In 1972 It0 (19) restated his previous proposal in the light of Marr's theory, that cerebellar function is an adaptive feedforward control mechanism for movements, so that active movements programmed in the cerebellum can be performed without ref- erence to the end effect. One essential point common to all these theories is that the inferior olivary nucleus and its climbing fibres constitute the "teaching line" to the purkinje cells of the cerebellar cortex.
Some evidence for this theory has appeared. The evidence of Gilbert and Thach (12) is the most interesting. They showed that in trained Rhesus mon-
keys, the adaptation of arm control to unaccustomed resistances is accom- panied by increased climbing fibre activity in the form of complex spikes recorded from the cerebellar cortex. In other words, during the time that monkeys learn to adapt to changes in resistance on their arms, the inferior olive increases its output to the purkinje cells, thus suggesting that the olive perhaps "teaches" the purkinje cells.
Llinas has produced (22) further evidence for a type of motor learning, namely, learning to compensate for ataxia, which involves the olive. He vestibulectomized rats and allowed them six months to compensate for the ataxia. Then he pharmacologically destroyed their olives and the ataxia returned, suggesting that the retention of the ability to compensate for the ataxia resided in the olive and/or its connections. He further showed that in vestibulectomized rats that had compensated for the ataxia, removal of their cerebellar cortices, without destroying the olives, did not result in a return of the ataxia. This showed that retention of compensation resided, NOT in the cerebellar cortex, but in the inferior olive and/or other struc- tures.
Llinas' results suggest that in a phylogenetically early type of motor learning (that is, adapting to ataxia), the inferior olive is independent of the cerebellar cortex. Higher up the phylogenetic scale, however, Nashner (29) has shown that in humans with cerebellar dysfunction there is a loss of adaptive gain in postural control. It may be that as one ascends the phylo- genetic scale the cerebellum becomes involved with the olive in learning. It would seem, too, that as one ascends the scale, the complexity of theories on learning ought to increase.
The vestibulo-ocular reflex (VOR) is another example of increasing com- plexity as one ascends the phylogenetic scale. Ito (19) showed that adaptive gain of rabbits' VOR was abolished by destruction of the inferior olives. Robinson (14,35) showed a similar loss after destruction of cats' flocculus.
800

Miles, however, concluded (27) that the situation is different in Rhesus
monkeys: The flocculus does not control changes in the gain of the VOR.
One interpretation of these apparently divergent results could be that as one
ascends the phylogenetic scale, complexity increases.
Returning to motor learning, the question is posed: Is motor learning
in primates simply a matter of the inferior olive teaching the purkinje cells, or is it more complex? Is it possible that there is not just a teaching line
but a teaching circuit as well? Such a circuit is proposed, not because an
increase in olivary activity during learning should be thought a deficient explanation, but because it does not explain why the olive is relatively silent in between periods of learning. Does the olive know that the pur- kinje cells have learnt? Does it get "shut off" and remain "quiet"?
The first question is why it should have to know that the purkinje cells
have learnt. Rubia and Kolb (37) have shown in the decerebrate cat that com-
plex spike activity shows phasic and tonic characteristics during passive
movements of the paw. They conclude that complex spike activity can directly signal peripheral events. In the actively performing chronic monkey, Gilbert
and Thach (12) showed phasic activity of complex spikes during motor learn- ing. This phasic activity seemed more than coincidental with the learning,
suggesting that the factors causing the phasic activity in the decerebrate
cats should be different from those in the learning monkeys. Since it is generally believed that motor learning in chronic monkeys takes place active- ly at a supraspinal level, it would seem appropriate that the phasic activity
controlling factors should be integrated functionally and anatomically with the neural learning factors. The teaching circuit described below is an at-
tempt to delineate such a phasic control system.
DISCUSSION
Anatomy
The proposed circuit is illustrated in diagram 1, and is for primates
only. The circuit consists of the principal nucleus of the inferior olivary
complex and its climbing fibres, purkinje cells and their efferents, dentate nucleus, dentato-rubral tract, parvocellular red nucleus and rubro-olivary tract. Note that the teaching circuit includes the teaching line. Massion has stressed (24) important phylogenetic differences by showing that, i.n cats, the parvocellular red nucleus is much smaller than the magnocellular
part, and the dominant output of the red nucleus is the rubrospinal tract
from the magnocellular part, but, in primates, the parvocellular red nucleus and rubro-olivary tract is dominant, while the magnocellular part and the rubrospinal tract virtually disappear. The main output of the red nucleus in primates is to the principal inferior olive (8,24,34,40). The main af- ferent to the red nucleus from the cerebellum in primates is from the dentate
nucleus (10,16), whose afferents come from the neocerebellum (lateral cere- bellar hemispheres) (7). Descending afferents to parvocellular red nucleus come from motor areas 8, 6, 4, and supplementary, from sensory areas 5 and 7,, and from visual cortex areas 17, 18, and 19 (10,16).
The development of motor skills is matched by the development of the principal inferior olive, lateral cerebellar hemispheres, dentate nucleus
801
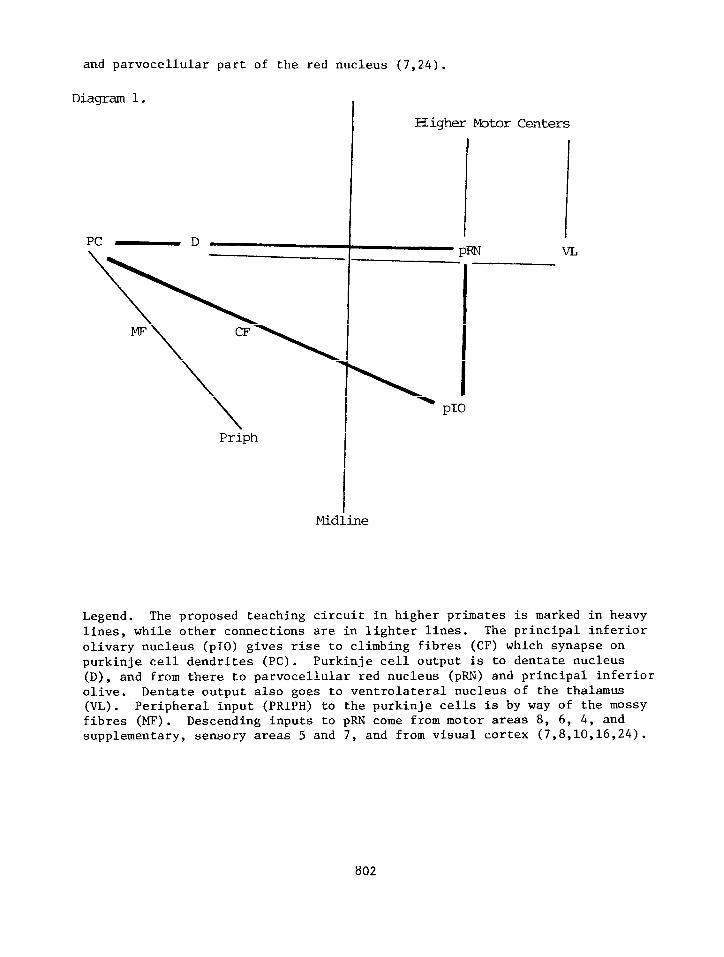
and parvocellular part of the red nucleus (7,24).
Diagram 1.
Higher W&or Centers
PC - D W’J VL
PI0
Priph
Midline
Legend. The proposed teaching circuit in higher primates is marked in heavy
lines, while other connections are in lighter lines. The principal inferior
olivary nucleus (~10) gives rise to climbing fibres (CF) which synapse on
purkinje cell dendrites (PC). Purkinje cell output is to dentate nucleus
(D), and from there to parvocellular red nucleus (pRN) and principal inferior olive. Dentate output also goes to ventrolateral nucleus of the thalamus
W). Peripheral input (PRIPH) to the purkinje cells is by way of the mossy fibres (MF). Descending inputs to pRN come from motor areas 8, 6, 4, and supplementary, sensory areas 5 and 7, and from visual cortex (7,8,10,16,24).
802

Physiology
A physiological study by Oka and Jinnai in cats (30) confirms the rubro-
olivary pathway, the dentato-rubral pathway, and the connection from parietal
association cortex (area 5) to parvocellular red nucleus.
Otero (31) studied red nucleus activity during learned movements in
Rhesus monkeys, and found, among other things, that the peak distribution of
onset times of red nucleus responses occurred 120 ms after precentral gyrus
peak responses. He concluded the red nucleus activity is dependent on the
combined actions of two inputs: (i) A central program from precentral gyrus
and/or cerebellum; and (ii) Sensory feedback from movement relayed via post-
central gyrus and/or cerebellum.
In 1965, Passouant et al. (32) studied the relationship between the red
nucleus and the inferior olive. They found that cutaneous stimulation of the forepaw and electrical stimulation of the red nucleus, in cats, could simul-
taneously be recorded in the inferior olive. As the stimuli coincided, at-
tenuation of the peripheral impulse occurred.
In 1969, Miller, Nezlina, and Oscarsson (26) stimulated in the region of
the red nucleus in cats and recorded in a narrow and lateral strip of the contralateral pars intermedia of the anterior lobe of the cerebellum. They found that such stimulation evoked climbing fibre responses. They showed too
. . . "that the same climbing fiber responses were activated from the midbrain, the contralateral motor cortex, and the dorsolateral funiculus of the spino-
olivo-cerebellar tract. The convergence from the DLF-SOCP incidentally per- mits the conclusion that the mesencephalic paths relay through the inferior
olive."
These interesting conclusions hint at an involvement of the red nucleus in motor control in cats: perhaps too, in primates.
Pathology
Lesions of the dentato-rubro-olivary pathway in 29 human cases have been
discussed by Jellinger (17). Discrete lesions of the medial and dorsal red nucleus in humans (corresponding to PRN in other primates) have been reported by Lapresle and Hamida (18). These lesions result in hypertrophic degenera- tion in the ipsilateral olive, whereas lesions in other parts of the midbrain did not affect the olive. Despite these reports and despite consideration of clinical syndromes, no useful knowledge has been gained regarding underlying function.
Experimental lesions likewise shed no light on the fundamental function of the rubro-olivary connection. Harmaline induced tremor of 6-12 Hz is generated by rhythmic firing of olivary neurones (28). Gogolak et al. (13) have recorded firing of the red nucleus in synchrony with cerebellar dis- charge in rabbits. Ranish and Soechting (33) produced electrolytic lesions in and around the RN in 2 monkeys and found tremor during "fast, ballistical- ly initiated" flexion movements of the forearm.
Pathology, clinical or experimental, tells little of the fundamental
803

function of the rubro-olivo-cerebellar connection.
CONCLUSION
Having considered the neural structures of the proposed teaching circuit, and not having found an exclusive function for at least one of its key com- ponents, namely the red nucleus, one is tempted to speculate on function of the circuit as a phasic control system, focussing particularly on the parvo- cellular red nucleus and principal inferior olive.
The principal inferior olive receives its input, not from the periphery, but from motor cortical areas 4 and 6, sensory cortical areas 1,2,3,5 and SII, and visual cortical areas 17,18, and 19. This descending input relays in the parvocellular red nucleus (5,6), as well as going directly to the olive (6). It is here suggested that this descending input is met at the red nucleus by input from the dentate nucleus, and that the merging of both in- puts determines the output of the parvocellular red nucleus to the principal inferior olive. This parvocellular red nucleus output, it is suggested, will "shut off" the inferior olive when a movement has been learnt.
When performing a controlled, learned movement dentate output precedes the second precentral response (25,39) (or "intended response" of Evarts and Tanji (9)): Indeed, it is held (6,38) that dentate output controls movement from moment to moment. It is here proposed that during a controlled, learned movement a corollary discharge goes from dentate to parvocellular red nucleus This corollary discharge causes parvocellular red nucleus to act on principal inferior olive to effect "shut off" of the principal olive when the movement has been learnt and to keep it "quiet" during controlled learned movements. How could this be possible? Consider: When a new instruction from higher motor centers acts directly on the inferior olive it is suggested that it also goes as corollary discharge to the parvocellular red nucleus. This descending input causes the red nucleus to remove its "quieting" effect from the olive. The olive now becomes active and instructs the purkinje cells, which send impulses to the dentate nucleus under the influence of peripheral feedback via mossy fibre input (which feedback is a direct consequence of the new instruction). This "newly learnt" dentate output merges at parvocellular red nucleus with the original instruction from the higher motor centers, and recognizing it as a "matching" instruction, shuts it off. The higher motor center effect on the red nucleus is thus removed and the red nucleus, once again under the influence of the dentate, now "shuts off" the principal ol- ive, and keeps it "quiet" during controlled learned movements.
Failure of the red nucleus to "shut off" and "quieten" the principal inferior olive should result in a continuation of the learning, so that the olive would continue to instruct the purkinje cells. Thus more purkinje cells than necessary would learn the elements of the movement (or one pur- kinje cell would learn more than is necessary), and the efficient execution of the movement would be affected. To illustrate: If a certain number of purkinje cells are required to execute a certain range of movement, and too many purkinje cells are instructed due to a red nucleus lesion, then the range that has been learnt by the purkinje cells will be executed, resulting in overshoot. Subsequent attempts at correction would result in tremor, which is the classical sign of a red nucleus lesion in clinical neurology.
804

If the function of the parvocellular red nucleus is to inhibit "flooding"
of the purkinje cells with useless new instructions, in other words to inhi-
bit "overlearning", then one could consider the teaching circuit as a phasic
control system which inhibits useless instructions and thus maintains the
efficiency of motor learning. A practical conclusion is that the Red Nucleus with its connections is ripe for anatomical, electrophysiological, and rever- sible lesion investigations all of which now seem possible (21,36).
The author wishes to thank his mentor, Dr. V.B.Brooks, and his colleagues, Drs. A-Miller and FFG.Ross, for their constructive critisms.
REFERENCES
1. Albus JS. A theory of cerebellar function. Math. Biosci. 10: 25, 1971.
2. Blomfield S, Marr D. How the cerebellum may be used. Nature 227: 1224,
1970.
3. Brindley GS. The use made by the cerebellum of the information that it receives from sense organs. IBRO Bull. 3 (3): 80, 1964 (Abstract).
4. Brindley GS. Nerve net models of plausible size that perform many simple learning tasks. Proc. R. Sot. Lond. [Biol.] 174: 173, 1969.
5. Brooks VB. Motor programs revisited. in press in Posture and Movement: Perspective for Integrating Sensory and Motor Research on the Mammalian Nervous System. (RE Talbott, DR Humphrey, eds) Raven Press, New York,
1979.
6. Brooks VB. Control of intended limb movements by the lateral and inter- mediate cerebellum. in press in Integration in the Nervous System. (H Asanuma, VJ Wilson, eds) Igako-Shoin, Tokyo, 1979.
7. Ghan-Palay V. Cerebellar Dentate Nucleus. Springer-Verlag, Berlin, 1977.
8. Courville J, Otabe S. The Rubro olivary projection in the macaque. An experimental study with silver impregnation methods. J. Comp. Neurol. 158: 479, 1974.
9. Evarts EV, Tanji J. Reflex and intended responses in motor cortex pyrami- dal tract neurons of monkey. J. Neurophysiol. 39: 1069, 1976.
10. Flumerfelt BA, Otabe S, Courville J. Distinct projections to the red nucleus from the dentate and interposed nuclei in the monkey. Brain Res. 50: 408, 1973.
11. Gilbert PFC. A theory of memory that explains the function and structure of the cerebellum. Brain Res. 70: 1, 1974.
12. Gilbert PFC, Thach WT. Purkinje cell activity during motor learning. Brain Res. 128: 309, 1977.
805

13. Gogolak G, Jindra R, Stumpf Ch. Effect of harmaline on the cerebello- rubral system. Experientia 33 (10): 1352, 1977.
14. Haddad GM, Friendlich AR, Robinson DA. Compensation of nystagmus after 8th nerve lesions in vestibulo-cerebellectomized cats. Brain Res. 135:
192, 1977.
15. Hebb DO. The Organization of Behaviour: A Neuropsychological Theory. Wiley, New York, 1949.
16. Humphrey DR, Gold R, Reed DJ. Origins of cortical and cerebellar projec- tions to the red nucleus in the monkey. Abstracts, Proc. Sot. Neurosci.,
November, 1978.
17. Jellinger K. Hypertrophy of the inferior olives. Report on 29 cases.
Zand. Neurol. 205: 153, 1973.
18. Lapresle J, Hamida MB. The dentato-olivary pathway. Somatotopic relation- ship between the dentate nucleus and the contralateral inferior olive.
Arch. Neurol. 22: 135, 1970.
19. Ito M. Neural design of the cerebellar motor control system. Brain Res.
40: 81, 1972.
20. Ito M, Miyashita Y. The effects of chronic destruction of the inferior olive upon visual modification of the horizontal vestibulo-ocular reflex
of rabbits. Proc. Japan Acad. Sci. 51: 716, 1975.
21. Kennedy PR, Ross H-G. Cooling in the brainstem of chronic monkeys. To be published. Physiol. Behav.
22. Llinas R, Walton K, Hillman DE, Sotelo C. Inferior olive: its role in learning. Science 190: 1230, 1975.
23. Marr D. A theory of cerebellar cortex. J. Physiol. (London) 202: 437, 1969.
24. Massion J. The mammalian red nucleus. Physiol. Rev. 47 (3): 383, 1967.
25. Meyer-Lohmann J, Conrad B, Matsunami K, Brooks VB. Effects of dentate cooling on precentral unit activity following torque pulse injections into elbow movements. Brain Res. 94: 237, 1975.
26. Miller S, Nezlina N, Oscarsson 0. Climbing fiber projection to the cere- bellar anterior lobe activated from structures in midbrain and from spinal cord. Brain Res. 14: 234, 1969.
27. Miles FA, Braitman DJ. Effects of prolonged optical reversal of vision on the vestibulo-ocular reflex: some neurophysiological observations. Abstracts, Proc. Sot. Neurosci., November, 1978.
28. deMontigny C, Lamarre Y. Activity in the olivo-cerebella-bulbar system of the cat during ibogaline- and oxotremorine tremor. Brain Res. 82: 369, 1974.
29. Nashner LM. Adapting reflexes controlling the human posture. Exp. Brain Res. 26: 59, 1976.
30. Oka H, Jinnai K. Electrophysiological study of parvocellular red nucleus neurons. Brain Res. 149: 239, 1978.
31. Otero JB. Comparison between red nucleus and precentral neurons during learned movements in the monkey. Brain Res. 101: 37, 1976.
806

32. Passouant P, Cadilhac J, Baldy-Moulinier M, et Cabanac P. Don&es experimentales sur l'olive bulbaire. Revue Neurologique 113 (5): 489, 1965.
33. Ranish NA, Soechting JF. Studies on the control of some simple motor
tasks. Effects of thalamic and red nucleus lesions. Brain Res. 102:
339, 1976.
34. Robertson LT, Stotler WA. The structure and connections of the developing inferior olivary nucleus of the rhesus monkey. J. Comp. Neurol. 158 (2):
167, 1974.
35. Robinson DA. Adaptive gain control of vestibulo-ocular reflex by the cerebellum. J. Neurophysiol. 39: 954, 1976.
36. Ross H-G, Kennedy PR, Brooks VB. Frequency changes of complex spikes of monkey's cerebellar Purkinje cells during cooling of the inferior olive.
Proc. Can. Fed. Biol. Sot., Vancouver, 1979 (Abstract).
37. Rubia FJ, Kolb FP. Responses of cerebellar units to a passive movement in
the decerebrate cat. Exp. Brain Res. 31: 387, 1978.
38. Strick PL. Cerebellar involvement in "volitional" muscle responses to
load changes. p 85 in Cerebral Motor Control in Man: Long Loop Mecha-
nisms. vol 4 (JE Desmedt, ed) Karger, Basel, 1978.
39. Vilis T, Hore J, Meyer-Lohmann J, Brooks VB. Dual nature of the pre-
central responses to limb perturbations revealed by cerebellar cooling.
Brain Res. 117: 336, 1976.
40. Walberg F. Descending connections to the inferior olive. J. Comp. Neurol.
104: 77, 1956.
807