the plastid ribosomal proteins (1): identification of all ... · summary identification of ......
TRANSCRIPT

1
30S PRP ms-su-revised.doc
JBC (protein synthesis and post-translational modifications)
The Plastid Ribosomal Proteins (1):
Identification of all the Proteins in the 30S Subunit of an Organelle
Ribosome (Chloroplast)
Running title: Plastid Ribosomal 30S Subunit Proteins
Kenichi Yamaguchi, Klaus von Knoblauch, and Alap R. Subramanian*
From the Department of Biochemistry, The University of Arizona, Tucson, USA
and the Max-Planck-Institut für moleculare Genetik, Berlin-Dahlem, Germany
*Corresponding Author:
Dr. A. R. Subramanian, Department of Biochemistry, BioSciences West, The University
of Arizona, Tucson, AZ 85721-0088
Fax: +1 520 621 9288
E-mail: [email protected]
The protein and nucleic acid sequences reported in this paper have been deposited in
SWISS-PROT and GenBank/EMBL data bases, respectively. For (over 30) new
accession numbers, see Table I and text.
Copyright 2000 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on June 28, 2000 as Manuscript M004350200 by guest on July 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
ABBREVIATIONS
Abbreviations used are: PRP, plastid ribosomal protein; PSRP, plastid-specific ribosomal
protein; RP, ribosomal protein; TP30, total protein of 30S subunit; 2D-PAGE, 2-
dimensional polyacrylamide gel electrophoresis; HPLC, high-performance liquid
chromatography; LC/MS, reversed-phase HPLC coupled with electrospray ionization
mass spectrometry; MALDI-TOF, matrix-assisted laser desorption/ionization time-of-
flight; ESI MS electrospray ionization mass spctrometer; EST, expressed sequence tag;
AA, amino acid(s); NTE/CTE, N-terminal/C-terminal extension.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

3
SUMMARY
Identification of all the protein components of a plastid (chloroplast) ribosomal 30S
subunit has been achieved, using 2-dimensional gel electropholesis, HPLC purification,
N-terminal sequencing, PCR-based screening of cDNA library, nucleotide sequencing
and mass spectrometry (ESI, MALDI-TOF, and LC/MS). Twenty-five proteins were
identified, of which 21 are orthologues of all E. coli 30S ribosomal proteins (S1 to S21),
and 4 are plastid-specific ribosomal proteins (PSRP's) that have no homologues in the
mitochondrial, archaebacterial, or cytosolic ribosomal protein sequences in databases.
Twelve of the 25 plastid 30S ribosomal proteins (PRP's) are encoded in the plastid
genome whereas the remaining 13 are encoded by the nuclear genome. Post-translational
transit peptide cleavage sites for the maturation of the 13 cytosolically synthesized PRP's,
and post-translational N-terminal processing in the maturation of the 12 plastid
synthesized PRP's are described. Post-translational modifications in several PRP's were
observed: alpha-N-acetylation of S9; N-terminal processings leading to five mature forms
of S6 and two mature forms of S10, C-terminal and/or internal modifications in S1, S14,
S18, and S19, leading to two distinct forms differing in mass and/or charge (the
corresponding modifications are not observed in E. coli). The four PSRP's in spinach
plastid 30S ribosomal subunit (PSRP-1, 26.8 kDa, pI 6.2; PSRP-2, 21.7 kDa, pI 5.0;
PSRP-3, 13.8 kDa, pI 4.9; PSRP-4, 5.2 kDa, pI 11.8) comprise 16% of the total protein
mass (67.6 kDa) of the 30S subunit (429.3 kDa). PSRP-1 and PSRP-3 show sequence
similarities with hypothetical photosynthetic bacterial proteins, indicating their possible
origins in photosynthetic bacteria. We propose the hypothesis that PSRP's form a "plastid
translational regulatory module" on the 30S ribosomal subunit structure, for the possible
mediation of nuclear factors on plastid translation.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

4
INTRODUCTION
The plastid (chloroplast) ribosome is a plant-specific, organelle ribosome that is
responsible for the synthesis of fewer than a hundred polypeptides (all encoded in the
plastid genome) but these include components of some of the most abundant plant
proteins. The plastid translation system is thus a pivotal component of the plant
biosystem. The largest functional entity of a translational system is the ribosome,
composed of both RNA and 50-100 distinct proteins. The rRNA’s in all categories of
ribosomes have been fully identified, but complete characterization of ribosomal proteins
has been achieved for only a few types of ribosomes (1-3). Protein characterization for
the organelle ribosomes of plastid/chloroplast has been in progress (reviewed in refs. 4-6)
and the results have indicated a significantly larger number of proteins than in the E. coli
ribosome.
The translation and its regulation in plastids differ significantly from that in eubacteria.
For example, only 30% of chloroplast mRNA's contain ribosome binding sites (RBS)
even though a canonical Shine-Dalgarno sequence is present in all sequenced plastid 16S
rRNA's (7). Plastid mRNA is edited in most plants, and the mature mRNA levels remain
relatively unchanged through dark/light transitions, whereas protein synthesis rates
increase dramatically upon illumination (8,9). Nuclear-encoded factors mediate light-
regulated translation (10) as well as transcription and post-transcriptional processes in
plastids (11,12), and many isolated nuclear mutants interfere with polysome assembly
(13). Thus, unlike in E. coli or cyanobacteria, the plastid translation system functions
with additional regulatory elements under the control of the nuclear genome. It is possible
that some of these controls is exerted through modifications in the plastid ribosome.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

5
Thus, a key to the understanding of plastid/chloroplast translation system could be a
precise elucidation of its component parts, and in particular those components that are
unique.
Two chloroplast-specific ribosomal proteins (PSRP's) in the 30S ribosomal subunit,
with no obvious homology to any E. coli ribosomal proteins have been previously
identified (14-16). Since then, the complete list of ribosomal proteins in yeast and
mammalian ribosomes (2-3) and many mitochondrial ribosomal proteins (17) have been
reported, but no homologues of the PSRP's exist in these ribosomes. The 30S ribosomal
subunit is responsible for the initiation of translation and therefore would be an
appropriate site for regulatory intervention. Expecting to find additional PSRP's, we
started a proteomic analysis of the plastid 30S subunit, using 2D-PAGE,
electroblotting/protein sequencing, HPLC separation, PCR-based screening/DNA
sequencing, and mass spectrometry (LC/MS, ESI MS and MALDI-TOF MS). A
summary of the results from this study are presented below, including identification of all
the proteins in the plastid 30S subunit, post-translational modifications, and identification
of two additional novel PSRP's. The protein/DNA sequence data reported here have been
deposited in SWISS-PROT and GenBank/EMBL databases. A corresponding study of the
plastid 50S subunit is in preparation for publication.
EXPERIMENTAL PROCEDURES
Chloroplast Ribosome and 30S Subunits - Preparation of spinach (Spinacia oleracea,
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

6
cv. Alwaro) chloroplast ribosomes, and zonal purifications of 70S and 30S subunits
(5000 A260 units of crude ribosome/zonal rotor for 70S purification and 3000 A260 units
of purified 70S/zonal rotor for 30S isolation) were done as described (15,18). The
approximate conversion factor, 1 A260 unit of 30S = 60 ug protein was used for protein
estimation. TP30 (total protein of 30S subunit) was extracted basically according to
Hardy et al. (19). For 2D-PAGE analysis, TP30 extract was dialyzed (Spectra/Por
membrane, 3,500 MWCO) for 16 hr against 8M urea, 0.1% beta-mercaptoethanol and
concentrated on Sephadex G-50. For HPLC, the extract (1 mg TP30) was dialyzed using
the same membrane for 4 hr successively against 30%, 10%, and 5% acetic acid
(containing, 0.1% beta-mercaptoethanol), dried in a Speed-Vac, and dissolved in 500 ul
of 5% acetic acid, 0.1% beta-mercaptoethanol. For LC/MS analysis, the extract was
dialyzed using a Slide-A-Lyser unit (3,500 MWCO, Pierce) for 16 hr against 5% acetic
acid, and concentrated using Microcon-3 (Amicon).
Reversed-phase HPLC - The HPLC system was assembled with a WellChrom Maxi-
Star K-1000 HPLC pump (KNAUER, Germany), injection valve, No.7125 (Rheodyne,
USA), and UV detector, UV-VIS S-3702 (Soma, Japan). Separation of TP30 was
performed basically according to Kamp et al. (20) using Vydac C18 column (4.6 mm x
250 mm) by a gradient of 0.1% trifluoroacetic acid (TFA) and 0.08% TFA in isopropanol
at flow rate of 0.5 ml/min.
Protein Electrophoresis and Electroblotting - SDS-PAGE was done by the method of
Laemmli et al. (21). 2D-PAGE (basic and acidic) were done as described previously
(22,23). Electroblotting was according to Walsh et al. (24) using a tank-blotting chamber.
PVDF-membrane (Sequi-Blot, BIO-RAD) was used as transfer membrane. After
electroblotting, the blot was rinsed in water and stained with 0.1% Amido Black 10B
(Sigma) in 50% methanol for 5 min, and destained with 50% methanol until the
background color disappeared. It was rinsed 3 times in water, dried on Whatman filter
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

7
paper and stored at -20 oC until used.
Protein sequencing - Protein sequencing was done at the Laboratory for Protein
Sequencing and Analyses (University of Arizona) on an Applied Biosystem 477A
Protein/Peptide sequencer interfaced with a 120A HPLC analyzer to determine
phenylthiohydantoin (PTH) amino acids. At the conversion step, 50 ul volume (out of
135 ul) is injected into Vydac C18 column for detection of PTH-AA; the remaining
sample going to fraction collector for further analysis.
PCR Screening and Nucleotide Sequencing - A lambda gt11 spinach cDNA library
(25) was screened by thermal gradient PCR using a Mastercycler gradient PCR apparatus
(Eppendorf Scientific, Inc., USA). Thermal gradient PCR was performed for: 3 min at 94
oC, 35 cycles of 1 min at 94
oC, 1 min at 43-60
oC, 1 min 30 sec at 72
oC with 0.25U of
Taq DNA polymerase (Gibco-BRL) in a 20 ul reaction volume containing 1 ul spinach
cDNA library (~108 pfu), 5 uM gene specific primer (or 10 uM degenerate primer), 5 uM
arm primer (PF or PR), 20 uM each dNTP, 1.5 mM MgCl2, and 50 mM KCl in 20 mM
Tris-HCl (pH 8.4). The oligonucleotide primers used were:
PF: 5'-CGGGATCCGGTGGCGACGACTCCTGGAGCCCG-3'
PR: 5'-CGGGATCCCAACTGGTAATGGTAGCGACCGGC-3'
PS6F1: 5'-CAAGAGGAAGAACAAAGCAG-3'
PS6R1: 5'-TADATICCRTCIARRTARTTRTT-3'
PS6R2: 5'-CTGCTTTGTTCTTCCTCTTG-3'
PS9F1: 5'-CARGAYAAYCCIYTITGGYTICARTA-3'
PS9F2: 5'-GGTCAAGCACAAGCAATATCTC-3'
PS9R1: 5'-ACTGACCTTCAACAATGCCC-3'
PS1F1: 5'-GGTGACATGATTCGTAATCCGAAG-3'
PF (forward primer) and PR (reverse primer) are complementary to the cloning site of
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

8
lambda gt11. Degenerate primers, PS6R1 and PS9F1, were designed from PRPS6 peptide
1 (sequence region: NNYLDGIY) and PRPS9 peptide 1 (sequence region:
QGNPLWLQY), respectively (see also Table I). PRPS6 gene specific primers, PS6F1
and PS6R2, were designed from the nucleotide sequence of PCR product (PF/PS6R1)
which was amplified using primer sets PF and PS6R1. PRPS9 gene specific primers,
PS9F2 and PS9R1, were designed from the nucleotide sequence of PCR product (PS9F1/
PR). PS1F1 was designed from the nucleotide sequence in exon 5 of PRP S1 (26).
PCR products were analyzed by agarose gel electrophoresis using 1% agarose gel and
visualized by ethidium bromide staining. Nucleotide sequences were analysed at the
DNA Sequencing Facility (Arizona Research Laboratories, University of Arizona) using
Applied Biosytems model 377 DNA sequencer.
Mass Spectrometry - Mass spectrometry was done at the Mass Spectrometry Facility
(Department of Chemistry, University of Arizona.) Electrospray ionization mass
spectrometry (ESI-MS) was carried out using a Finnigan LCQ system. Liquid
chromatography - mass spectrometry (LC/MS) was done by the same LCQ system
interfaced with a Michrom HPLC system (Magic 2002) using a Microbore C18 column
(150 mm x 1mm). The solvent system used was 0.1% TFA in 98% acetonitrile (solvent
A) and 0.1% TFA in 90% acetonitrile (solvent B). 50 pmol PRP in 10 ul 4% acetic acid
was subjected to ESI-MS. For LC/MS, 20 l TP30 (60 pmol) in solvent A was injected to
the Microbore C18 column. Matrix-assisted laser desorption/ionization time-of-flight
mass spectrometry (MALDI-TOF MS) was carried out on a Bruker Reflex-III. 100 fmol
PRP (HPLC pool) was deposited using sinapinic acid (SA) as a matrix. On MALDI-MS
analysis for endoproteinase Lys-C digest of PRP S9, alpha-cyano-4-hydroxycinnamic
acid (CCA) was used as a matrix. Samples were prepared by mixing one part of
protein/peptide sample with nine parts of matrix (saturated SA or CCA in 0.1% TFA,
30% acetonitrile), and 1 ul of this solution was applied onto the sample probe. External
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

9
calibration was carried out using three ion peaks from cytochrome c: [M+2H]2+
, 6,181;
[M+H]+ , 12.361; [2M+H]
+ , 24,723.
Enzymatic and chemical cleavage – "In-gel" digestion of PRPS6 and PRPS9 were
performed basically according to Hellman et al. (27). Each of the 5 spots were excised
from Coomassie blue stained TP30 gels (200pmol each PRP's /gel) and equilibrated with
mixing for 40 min at 30 oC in 2 ml of 50 mM Tris-HCl, pH 8.5 in 50% acetonitrile. The
gel pieces were completely dried in a Speed-Vac, then rehyderated with 150 ul of 50 mM
Tris-HCl, pH 8.5, 0.02% polyoxyethylenesorbitan monolaurate (Tween 20), 10%
acetonitrile, containing 0.4 g (ca. 1/40 enzyme/substrate ratio by weight) endoproteinase
Lys-C (Sigma), and incubated at 30 oC for 16 hr. The enzyme reaction was stopped by
the addition of 1/10 reaction volume of 10% TFA. Gel pieces were transferred in 500 ul
of 0.1% TFA in 60% acetonitrile and peptides were extracted by shaking at 30 oC for 80
min. The extract was dried in a Speed-Vac and dissolved in 50 ul of 5% acetic acid.
Peptides were purified by reversed-phase HPLC using Vydac C4 (150 mm x 4.6 mm) or
C8 (50 mm x 4.6 mm) columns in TFA-acetonitrile system. The purified peptides were
then dried and dissolved in 25 ul of 30% acetic acid, and stored at -20 oC until used.
Deblocking of PRPS9 was performed according to the method of Wellner et al. (28). A
PRPS9 spot (200 pmol/spot) was excised from the TP30 blot and incubated in 30 ul of
anhydrous trifluoroacetic acid for 3 min at 45 oC. The spot was dried under a gentle
stream of nitrogen for 1 min and placed in a 1.5 ml Eppendorf tube. The tube was
incubated at 60 oC for 12 hr, and the blot was then sequenced.
Computer analyses - The program BLAST (National Center for Biotechnology
Information, NCBI) was used for sequence searches. Protein sequence searches were
performed using the "blastp" program versus nr (non-redundant Database of GenBank
CDS translations, PDB, SwissProt, PIR, and PRF). EST searches were done using the
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

10
"tblastn" program versus dbEST (non-redundant Database of GenBank, EMBL, DDBJ
EST Divisions). ORF's (open reading frame) from cDNA sequences were analyzed using
the "map" program from GCG software (29). Isoelectric points and sequence masses
were calculated using "peptidesort" (29). Sequence alignments and comparisons were
performed using "pileup" and "gap" programs (29). LC/MS data was analyzed using
ThermoQuest Finnigan Xcalibur data system. Masses were deconvoluted from resulting
m/z using "BIOMASS Deconvolution" program and charge states were convoluted using
"BIOMASS Calculation".
RESULTS
Proteins of Chloroplast Ribosomal 30S Subunit - In order to identify all the PRP’s in
pure chloroplast 30S ribosomal subunit, spinach chloroplast 70S ribosomes were first
purified on a zonal gradient and then run on a second dissociating zonal gradient to
obtain 30S and 50S subunits. Efficient dissociation of chloroplast ribosomes required the
development of a phosphate-containing dissociating buffer (see 15 for details and typical
gradient profiles). TP30 was extracted from 30S subunits and proteins were resolved by
2D-PAGE (Fig. 1A). In further experiment, the resolved proteins were transferred onto
PVDF membrane, stained with Amido Black, spots excised and subjected to N-terminal
sequencing. As seen in Fig. 1A, many of the protein spots were composite-looking,
indicating partial, overlapping separations. The experiment was therefore repeated, with
each of the spots excised into 2 or 3 segments for N-terminal analysis. The sequence data
from all the experiments are summarized in Table 1.
In most instances the N-terminal sequences were sufficient to allow protein identification
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

11
from similarity using the BLAST search. In a few cases (N-terminally blocked proteins or
those showing insufficient similarity for identification), internal peptide sequences were
obtained as described in Experimental Section. The chloroplast proteins (PRP's) are
designated in Table I as S1, S2, ... S21, as per their sequence similarity to corresponding
E. coli ribosomal proteins. Fig. 1B shows the positions of all the identified proteins in the
2D-PAGE of Fig. 1A. The designations alpha, beta, PSRP-1 etc. are discussed in a later
section.
In order to confirm that all spinach chloroplast 30S subunit PRP's appeared in the 2D-
PAGE pattern, two additional experiments were performed. The 2-D system used in Fig.
1A resolves proteins of pI 4.5 or greater (22); acidic proteins of lower pI do not migrate
into the gel. By running TP30 in another 2D-PAGE system, suitable for acidic proteins
(pI 5.5 or lower, 23), such acidic proteins are resolved. A few TP30 spots appeared in the
2D-PAGE (acidic) gel, but N-terminal sequencing did not reveal any new sequences.
However if a N-blocked PRP and an unblocked protein were to migrate to a single spot, it
would appear under Edman degradation that the spot contained only one protein.
Therefore, we separated 30S subunit PRP's (TP30) on a reversed phase HPLC column,
where hydrophobic interactions rather than net charge or peptide chain length are the key
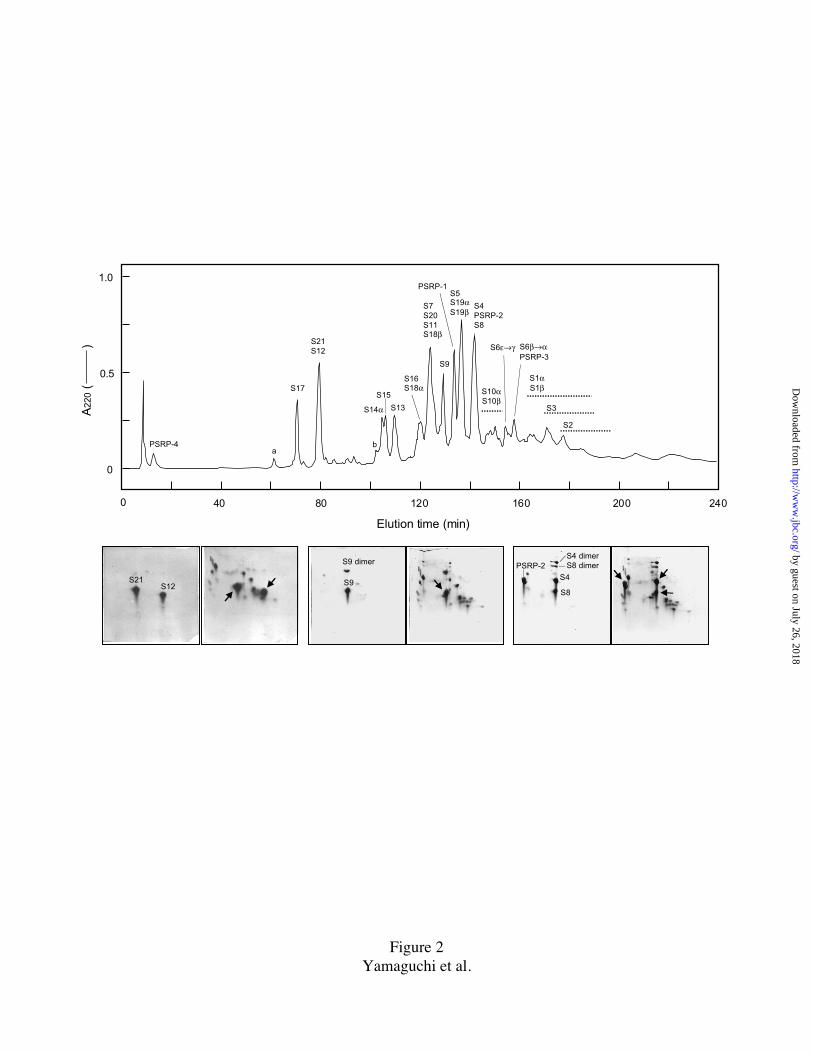
to resolution. HPLC may often separate proteins that are unresolved by 2D-PAGE. Fig.2
shows the separation obtained by this procedure. We determined the N-terminal
sequences of the material in the individual peaks, but this experiment only confirmed the
previous data set and provided no new significant sequences. Identification of the PRP's
in the individual peaks is also shown in Fig.2. Interestingly, PRP's S2, and S3 show poor
tailing separations in both HPLC and 2D-PAGE (Figs. 1 and 2). The minor unmarked
peaks/spots in the HPLC and 2D-PAGE separations were all subjected to N-terminal
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

12
analysis but did not yield any new significant data, other than the indication that some
chloroplast PRP's are dimerized under the experimental conditions.
The yields of PTH-AA recovered in the first three cycles of Edman degradation is
summarized in Fig. 3. In those cases where the PRP exists in multiple forms, the yields
were summated. Most of the proteins showed yields of approximately the same amount
indicating that they exist in stoichiometric amounts. There were a few exceptions, i.e.
PRP's S2, S3, S8, S11, and PSRP-3; and S9 which showed zero yield (discussed later).
The 2D-PAGE spots of S2 and S3 were always unusually elongated (tailing in 1st
dimension): for N-terminal analysis the concentrated region (i.e. only a portion), was
excised. S2 and S3 are most likely stoichiometric, unmodified proteins. For proteins S8
and S11, we do not have any further data to choose between relatively poor yields at
Edman degradation or partial N-terminal modification. For PSRP-3 (irregular protein
spot, Fig1), analysis of different parts of the spot indicated that possibly two-thirds of the
protein was N-terminally blocked.
The N-terminal sequences in Table I allowed the identification of 18 E. coli
orthologues and many post-translationally modified forms in spinach chloroplast 30S
subunit. The orthologues of the three other E. coli 30S subunit ribosomal proteins (E. coli
30S subunit contains 21 RP's: S1 to S21), namely, S6, S9, and S10, were identified from
additional derived protein sequences obtained by cloning and nucleotide sequencing.
The N-terminal and internal peptide sequences of PRPS6 and PRPS9 did not show
significant similarity to any proteins in databases. We therefore screened a spinach
lambda gt11 cDNA library (25) using inosine-containing degenerate oligonucleotide
primers designed from the sequences of PRPS6 peptide 1 and PRPS9 peptide 1. Thermal
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

13
gradient PCR allowed us to find the optimal amplification conditions using the
degenerate (PS6R1 and PS9F1) and lambda arm primer (PF or PR) sets. Amplified PCR
products (PF/PS6R1 and PS9F1/ PR) were sequenced, and further PCR amplifications
were done using sets of gene-specific primers based on the DNAsequence (forward,
PS6F1 and PS9F2; reverse, PS6R2 and PS9R1). The nucleotide sequences so obtained
lacked the transit peptide N-terminal coding regions of both PRPS6 and PRPS9, but the
data provided the complete mature protein sequences (and partial transit peptide
sequences). The cDNA nucleotide sequences of PRPS6 and PRPS9 are available under
GenBank accession numbers, AF250383 and AF242547. Comparison of spinach PRPS6
and PRPS9 protein sequences with homologues in a higher plant (Arabidopsis thaliana),
an alga (Porphyra purpurea), a photosynthetic bacterium (Synechocystis PCC6803) and
E. coli are shown in Fig. 4, confirming the identity assignments.
As compared to E. coli S6, spinach PRPS6 contains a 39 amino acid long, negatively
charged N-terminal extension (NTE), a 9 amino acid insert in the interior, and truncation
of the negatively charged, 39 amino acid long C-terminal region. This and the fact that
whereas PRPS6 shows amino acid identity of 83.4% to Arabidopsis F1N19.8 derived
sequence, but only 31.2%, 36.0%, and 27.7%, respectively to the S6 protein sequences
from P. purpurea, Synechocystis PCC6803, and E. coli, indicates a high level of
divergence in higher plant PRPS6 evolution.
As compared to E. coli S9, a 28 amino acid long NTE is present in spinach PRPS9, but
the C-terminal region in both proteins is well conserved. The chloroplast protein shows
amino acid identities of 82.8%, 52.6%, 54.1%, and 45.4%, respectively, to S9 protein of
A. thaliana, P. purpurea, Synechocystis PCC6803, and E. coli. The S9 proteins are well
conserved in their C-terminal half, particularly in the last 30 amino acids.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

14
PRP S10 was identified by a different procedure. The 22 residue long N-terminal
sequence shown in Table I did not allow identification by BLASTP search, while
TBLASTN search identified similarity to three EST clones of ice plant. We obtained one
of the ice plant EST clones, L0-67, and completely sequenced it. The sequence (available
under the GenBank accession number AF245665) codes for a 197 amino acid long PRP
precursor, comprising a putative chloroplast transit peptide and a mature protein with
homology to algal and eubacterial S10 ( Fig. 4). The ice plant PRPS10 thus acted as a
bridge between the N-terminal sequence of spinach chloroplast S10 and eubacterial and
algal chloroplast S10 sequences in databases.
With these 3 identifications (that of S10 being the weaker since we did not have its
cDNA sequence), the orthologues of all E. coli 30S proteins, S1 to S21, were identified in
spinach chloroplast ribosomal 30S subunit. Therefore the 4 additional proteins (PSRP-1
to PSRP-4), which show no significant similarity to any proteins in E. coli, yeast or
mammalian ribosomes, are named PSRP's (Plastid Specific Ribosomal Proteins). Two of
them, PSRP-1 and PSRP-4 (earlier named S31, here renamed PSRP-4), have been
previously reported (15,16) by us. We have studied the two newly discovered proteins,
PSRP-2 and PSRP-3, in greater detail and the results will be published elsewhere
(Yamaguchi and Subramanian, manuscript in preparation): Their cDNA sequences are
accessible under GenBank Nrs. AF240462 and AF239218.
Mass Spectrometry - Spinach chloroplast 30S ribosomal subunit proteins (TP30) were
analyzed by LC/MS and several of the peaks from the HPLC run, shown in Fig. 2, were
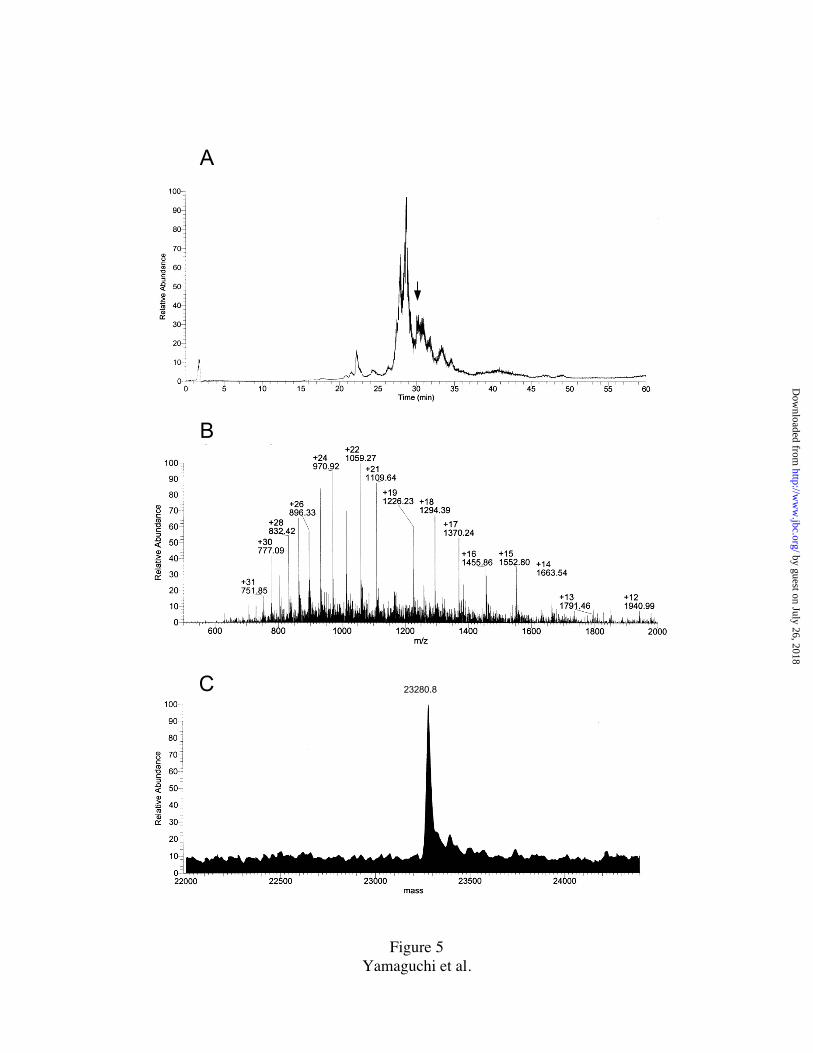
analyzed by MALDI-TOF. Fig. 5A shows the summed mass/charge ratio (m/z)
abundance of spinach 30S subunit PRP’s in the m/z range of 400 to 2,000 versus LC
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

15
elution time. An example of a mass spectrum (summed scans for 30 sec) at the elution
interval of 30.0-30.5 min is shown in Fig. 5B. Protein mass was calculated by
deconvolution of the m/z series. For example, deconvolution of the m/z series in Fig. 5B,
yielded a mass of 23,280.8 Da (Fig. 5C). As the observed mass 23,280.8 Da is consistent
with the sequence mass of PRPS4 (23,279.02 Da), it was identified as the mass of PRP
S4. When sequence mass was not available, ESI MS and/or MALDI-TOF MS of
individual HPLC pools (shown in Fig. 2), containing protein(s) identified by 2D-PAGE,
allowed identification. The results so obtained are summarized in Table II. A total of 22
proteins of spinach chloroplast 30S subunit were thus identified, and their exact
molecular masses determined. The data also included the exact masses for the individual
forms of several PRP's that exist in two or more forms (Table II).
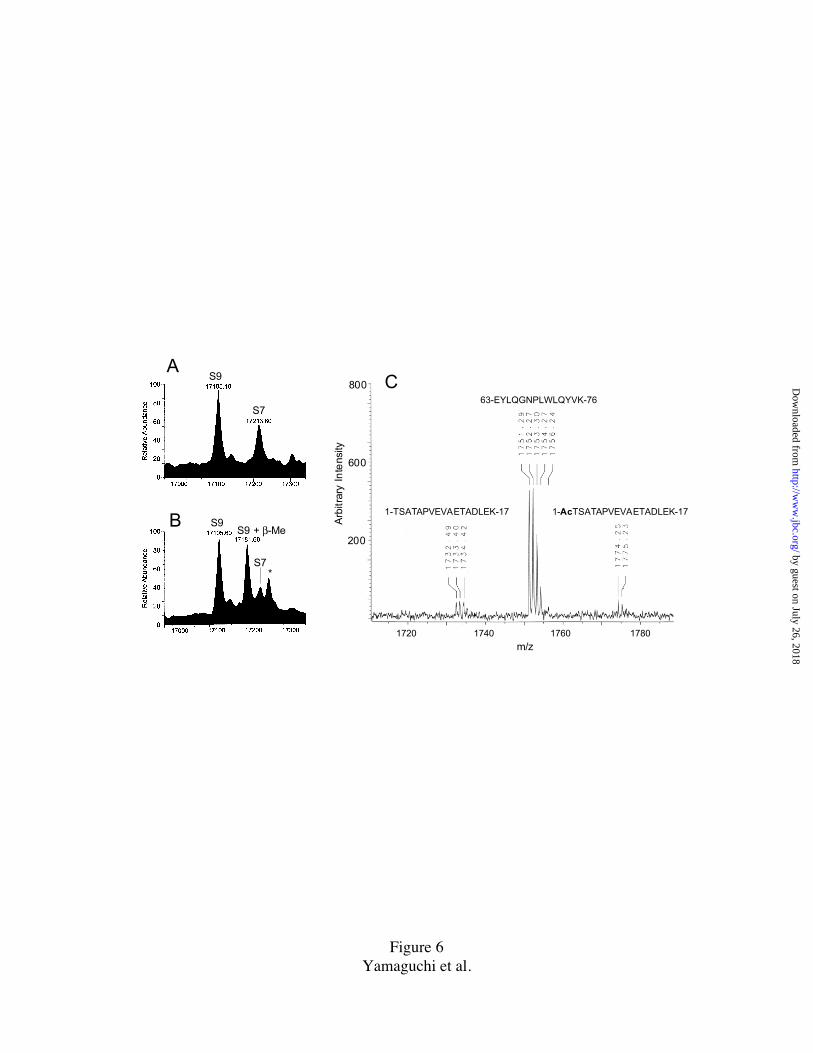
The case of PRPS9 – The spot identified as PRPS9 (Fig. 1) yielded no results in the
initial N-terminal analysis runs indicating N-terminal blockage. However, the N-terminal
sequence of TSATAPVEVA see Table 1) was obtained when the excised spot was
treated with trifluoroacetic acid (TFA) prior to sequence run. As the N-terminal residue
of PRPS9 is identified as threonine, the deblocking reaction could occur by an acid-
catalyzed N to O shift of alpha-N-acetyl group followed by beta elimination (28). From
its cDNA sequence (this paper), the calculated sequence molecular mass of spinach
PRPS9 is 17,063.6. The observed mass values were: 17105.1 (LC/MS of TP30) and
17105.6 (ESI MS of purified PRPS9), instead of 17063.6 (Fig. 6A). The data thus
suggested a post-translational modification of PRPS9 with mass increment of 42 Da,
corresponding to acetylation.
To confirm whether the 42 Da increment was indeed due to N-terminal acetylation,
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

16
purified PRPS9 was digested "in gel" with the endoproteinase Lys-C and the resultant
peptides were analyzed on MALDI-TOF MS (Fig. 6C). In the m/z range of 1700 to 1800,
three peptide ion peaks, resolving the isotope ion, were observed. The m/z peak of
1774.25 corresponds to an expected (acetylated) monoisotopic peptide mass (MH+,
1773.88: Ac-TSATAPVEVAETADLEK, residues 1-17), m/z peak 1751.29 corresponds
to an internal peptide (MH+, 1750.92: EYLQGNPLWLQYVK, residues 63-76), and m/z
1732.49 corresponds to a nonacetylated N-terminal peptide (MH+,1731.87
TSATAPVEVAETADLEK, residues 1-17). The data supports N-acetylation, since the
unmodified N-terminal peptide (m/z 1732.49) could be generated by acid-catalyzed
deblocking (28) when the enzymic reaction was stopped with 1% TFA. From the
sequencing and mass spectrometry results we conclude that the N-terminal threonine
residue of PRPS9 is modified post-translationally, most probably by acetylation.
DISCUSSION
All the proteins in the cytosolic ribosomes of yeast (2) and mammals (3) as well as the
prokaryotic ribosome of E. coli (1) are presently identified from a combination of protein
and DNA sequencing experiments. Similar endeavors have been in progress for the
organelle ribosomes of yeast mitochondria (17) and chloroplasts (reviewed in 4-6). In this
paper we present sufficient protein and nucleic acid data to establish the identification of
all the proteins in the 30S ribosomal subunit of a higher plant chloroplast/plastid
ribosome.
Nomenclature - We propose here a nomenclature for plastid ribosomal proteins and
their genes that accords with the CPGN (Commission on Plant Gene Nomenclature)
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

17
rules. Chloroplast ribosomal proteins are designated PRP (Plastid Ribosomal Protein), in
line with the designation of MRP for mitochondrial ribosomal protein (Abbreviation CRP
is avoided because "C" would suggest "Cytosolic": also, plastid is the general name for
the organelle). Orthlogues of E. coli ribosomal proteins occurring in chloroplast will
retain the E. coli designations, with the prefix PR added. Thus chloroplast/plastid
orthlogues of E. coli S1 to S21 and L1 to L36 are to be designated, PRPS1 to PRPS21
and PRPL1 to PRPL36. Chloroplast ribosomes contain proteins that do not have
homologues in E. coli (eubacteria), or the cytosolic ribosomes of yeast and mammals.
They (six in all, 4 in the 30S subunit, and 2 in the 50S subunit, ms in preparation) are
designated PSRP (Plastid Specific Ribosomal Protein), PSRP-1 to PSRP-4 in the 30S
subunit; the numbers ordered based on pI and molecular mass. The PSRP nomenclature is
recommend over the previous designations, e.g. S30/S22 for PSRP-1 and S31 for PSRP-4
(14-16), because S22/S30/S31 could be imagined to be homologues of cytosolic (or
mitochondrial) ribosomal proteins with those names. The genes for plastid/chloroplast
ribosomal proteins are distributed between the plastid's own genome and the nuclear
genome, and therefore it is informative if the gene name reflects gene location. In
accordance with the approved CPGN designation for plant genes (30), gene names are
written in italics, with nuclear genes having capital first letters, and organelle genes
having lowercase first letters. Thus, the gene for nuclear encoded PRPS1 is PrpS1,
whereas the gene for plastid encoded PRPS2 is prpS2, and gene for nuclear encoded
PSRP-1 is Psrp-1 (note: all six PSRP's are encoded in the nucleus).
Sufficient identification - This work had its main focus on proteins, and therefore
amino acid sequencing was carried out to the point of identification rather than
completion. We kept in view the impending completion of several plant genome
sequencing projects that are in progress, which would provide the complete (derived) RP
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

18
sequences. The genome sequencing data cannot however: a) identify proteins that are
specific to the chloroplast as ribosomal proteins, b) locate maturation cleavage sites in the
precursors of imported PSRP's and PRP's that are encoded in the nucleus, c) identify
post-translational N-terminal processing of PRP's that are encoded in the organelle
genome, d) provide information on occurrence of post-tanslational modifications in
individual RP's, and e) give precise delineation of the exact number of RP's comprising a
ribosome. In this work, even with incomplete protein sequence data, we were able to
accomplish almost all the tasks listed above.
Post-Translational Processing: Chloroplast RP's encoded in the organelle's genome -
It can be concluded from our data that orthologues of all the 21 E. coli 30S subunit RP's
are maintained in the plastid/chloroplast ribosome. Twelve of these proteins are encoded
by the plastid genome in spinach and other land plants, and are synthesized on the plastid
ribosome using formyl-Met-tRNA for initiation. Table III provides information on post-
translational N-terminal processing. Nine of the proteins have the initiating N-terminal
formyl-Met residue excised, but in 3 (PRP's: S15, S16, and S18) only the formyl group is
removed, showing that post-translational N-terminal processing occurs via multiple
pathways in the chloroplast. In E. coli, the excision of N-terminal Met by methionyl
amino peptidase depends on the size of the second amino acid's side chain: if it is bulky,
the N-terminal Met was not removed (31). The concerned side chains of PRP's S15, S16
and S18 are relatively bulky (Lys; Val; Asp, respectively); it is therefore likely that
plastid methionyl amino peptidase is related to the E. coli enzyme. The protein-
chemically determined N-terminal sequences of all the 12 proteins matched 100% with
the derived sequences from spinach plastid DNA, recently available in GenBank database
(accession AJ400848, April 2000), except for a total of 3 amino acid differences in S15
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

19
and S18, which could arise from cultivar difference. We can therefore extend the earlier
conclusion (32.) that the 30S RP genes in the plastid DNA are all functional genes.
Post-Translational Processing: Chloroplast 30S RP's encoded in the nuclear genome -
A total of 13 chloroplast 30S subunit RP's, including 9 E. coli orthologues and 4 plastid-
specific proteins, are encoded by the nuclear genome, and are synthesized on cytosolic
ribosomes as precursors that are considerably larger than the mature PRP's. The extra,
routing, peptide sequence at the N-terminus (transit peptide), directs the precursor
molecule to the chloroplast envelope, where it is cleaved off upon entry into the
organelle. This post-translational processing is performed by a stromal endopeptidase
(33), whose sequence specificity has not been fully delineated. Table III lists the pre- and
post-cleavage peptide sequences for 8, and post-cleavage sequences for 5 PRP's. Since
the N-terminal sequences of all nuclear encoded 30S PRP's have been determined, their
transit-peptide cleavage sites are now directly established.
Post-translational Modifications in 30S PRP's - A significant difference between
plastid RP's and E. coli RP's exists not only in protein composition but also in post-
translational modification. Plastid specific post-translational modification are discussed
below.
PRPS1 exists in two forms (S1alpha and S1 beta, Fig. 1) differing in charge and size.
Their relative amounts were estimated from the yields of PTH- amino acids from Edman
degradation during sequence analysis (S1 alpha, 71%; S1 beta, 29%). SDS-PAGE and
MALDI-TOF MS analysis showed S1 alpha and S1 beta to be 41 kDa and 37 kDa
proteins, respectively. As the N-terminal sequences of the two forms were the same, and
as S1alpha mass corresponded to the mass derived from cDNA sequence (34.), we
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

20
investigated the possibility of S1 beta being the product of a shorter mRNA generated by
alternative splicing (PrpS1 gene comprises 7 exons, ref. 26). If so, two cDNA species
encoding the two forms of S1 should exist, and they could be amplified by PCR. A
spinach cDNA library in lambda gt11 (25) was subjected to PCR amplification using an
appropriate S1-specific forward primer (PS1F) and a reverse primer designed from the
lambda right arm. The experiment yielded only a single amplified species, whose DNA
sequence corresponded to S1 alpha (data not shown). Thus, S1 beta must be derived by
post-translational modification from S1 alpha, either post-translational C-terminal
trimming or protein splicing of an intein.
The mature form of PRPS5 has a significantly long N-terminal extension of 86
residues compared to E. coli S5 (Fig. 7). Edman degradation of PRPS5 yielded an
unidentified N-terminal PTH-amino acid. Derived protein sequence of PRPS5 precursor
from database (accession # CAA63650) combined with the N-terminal sequence of the
mature form (Table I) showed N-terminal cysteine. Pyridylethylation of PRPS5 with 4-
vinyl pyridine under reducing conditions did not yield PTH-pyridylethyl Cys, and
therefore a modified cysteine is indicated. From MS data, the N-terminal Cys is likely to
be modified with 32.7 Da mass increment (Table II ). Evidently, as Edman degradation
was successfully performed to 32 cycles (Table I), the modification in PRPS5 does not
block the N-terminal amino group. Therefore, it is a different modification from that in E.
coli S5, where the protein is N-terminally acetylated, by RimJ gene product (35).
PRPS6 exists in five forms, and as briefly discussed in the Results Section (Table I and
Fig. 4), they differ only in their N-terminal portion. The relative amounts of the alpha to
epsilon forms were, 8%, 19%, 10%, 10% and 53%, respectively, as estimated from the
yield of PTH-amino acids. In E. coli ribosome, S6 occurs in five forms but the forms
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

21
arise by a post-translational, C-terminal glutamylation reaction (36). C-terminal poly-
glutamylation in spinach chloroplast S6 could be ruled out from our LC/MS analysis of
TP30 because we detected proteins with masses corresponding exactly to the PRPS6
forms of alpha, beta and epsilon (as calculated from the derived protein sequences, Fig.
4) and none with multiple increments of mass +129 Da (Glu residue). The forms gamma
and delta were not detected in the LC/MS experiment - probably due to the smaller
amounts and/or less ionization. It has been suggested that the S6 glutamylating enzyme
(gene, rimK) in E. coli recognizes the S6 C-terminal -Glu-Glu-COOH, since the
modification is abolished in an rpsF mutant with -Lys-Glu-COOH at the C-terminus (37).
Spinach chloroplast S6 has -Lys-Tyr-COOH at its C-terminus, in a highly positively
charged C-terminal context (5 of 10 residues basic ), whereas E. coli S6 has -Glu-Glu-
COOH in a highly negatively charged context. BLAST search of the database did not
reveal a plant gene matching the E. coli rimK gene. It is therefore likely that the C-
terminal of PRPS6 is not polyglutamilated. Spinach PRPS6 also shows significant
structural differences as compared to E. coli S6, e.g. a 39 residues long NTE and a C-
terminal truncation of 39 residues (Fig. 4). We suggest that it is likely the multiple forms
of chloroplast S6 arise from five different proteolytic cleavage sites in the PRPS6
precursor. However it is remarkable that, although using different post-translational
pathways at the N- and C-terminal regions, both E. coli S6 and chloroplast S6 exist in
five forms that carry similar overall net charge. Theoretical pI values, calculated from the
sequences for the alpha to epsilon forms of chloroplast S6 are, 4.59, 4.68, 4.78, 5.4 and
5.4, respectively, while the corresponding values E. coli S6 are, 4.75, 4.83, 4.91, 5.02,
and 5.17. Therefore, it appears that multiple forms of an acidic protein, of pI 4.5 - 5.5,
may be a requirement for a ribosomal biological function that is conserved in
photosynthetic organelle evolution. In this context, the X-ray structure of Thermus
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

22
thermophylus S6 shows the N- and C-terminal portions of the S6 protein are located side
by side forming a beta sheet structure (38).
PRPS9 (spinach) is N-terminally modified, probably by alpha-N-acetylation, while E.
coli S9 is a post-translationally unmodified protein. The alignment of bacterial and plant
S9 proteins showed less similarity at the N-terminal portion; even among the higher plant
S9's; but the C-terminal 30 amino acids are highly conserved, also between bacteria
higher plants. Indeed, chloroplast RPS9 was identified by an early experiment in our lab
(39) from its immunological cross-reaction to E. coli S9 antiserum.
PRPS10 exists in two electrophoretically resolvable forms, differing in their N-
terminal portion (Fig. 2 and Table I). Both forms were present in each of the two spots
(Fig. 1, S10 alpha and S10 beta) with the same relative amounts (estimated from PTH
amino acid yields), 60% alpha and 40% beta in either spot. As noted in Results Section,
we obtained the complete mature PRPS10 sequence of ice plant (Mesembryanthemum
crystallinum) chloroplast S10. The N-terminal portions of spinach and ice plant PRPS10
are well conserved, and therefore the post-translatinal modification sites in the two
proteins could be readily identified (Fig. 4). The land plant S10 proteins are significantly
longer than their homologues in E. coli, Synechocystis sp, and P. purpurea (marine alga)
chloroplast, and carry a negatively charged, 35-residue long NTE.
PRPS17 showed (Fig. 2, peak "a") a minor form having the same N-terminal sequence
of the major form, but a smaller mass of 10,614 Da by ESI MS analysis. It could be a
post-translationally modified (C-terminally truncated by 15 AA) form. The amount of the
minor form was insufficient to show a distinct spot on the 2-D gel (Fig. 1).
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

23
PRPS18 exists in two forms that are electrophoretically well separated (Fig. 1). E. coli
S18 exists in a single, N-terminally acetylated (RimI gene, 35) form, whereas both the
alpha (88%) and beta forms (12%) of PRPS18 are not N- terminally blocked. We have
not identified the post-translational modification in chloroplast S18 (mass spectrometry
did not provide data on S18 beta, Table II). The maize and rye (monocot plants)
PRPS18's show a 7-fold repeat of a positively charged, hydrophobic heptapeptide motif
(SKQPFRK) in their NTE, whereas only two such repeats are found in the PRPS18's of
the dicot plants pea and tobacco (40,41). The spinach N-terminal sequence shown here
includes two repeats. When rye PRPS18 was expressed in E. coli cells, 30-40% of the
expressed protein assembled in E. coli ribosomes and polysomes (42). The recently
published, 2.6A resolution, crystal structure of a ribonucleoprotein complex from the
central domain of Thermus thermophilus 30S subunit (containing both S6 and S18)
shows the three dimensional arrangements of S6 and S18 proteins on a junction of 16S
rRNA helices 22 and 23 (43). The N-terminal region of S18 and both the N- and C-
terminal regions of S6 are situated next to each other. These three terminal regions are at
the edge of the 30S subunit platform, where it is likely they play a role in translational
regulation via their net positive (S18) and negative (S6) charges.
Plastid Specific Ribosomal Proteins. The most important finding from the study of
chloroplast ribosomal proteins is likely to be the discovery and identification of plastid
specific ribosomal proteins (PSRP's) in this organelle ribosome. There are 4 such proteins
on the chloroplast 30S subunit. Since the orthologues of all E. coli 30S subunit proteins
exist in plastid 30S subunit (Table I), the origins and functions of the 4 additional
proteins are intriguing. PSRP-1 has 58.3% sequence similarity with the hypothetical
translation product of a light-repressed transcript (lrtA) gene of Synechococcus PCC 7002
(44). Whether lrtA gene product is a component of Synechococcus PCC 7002 ribosome is
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

24
not known. PSRP-1 has been expressed in E. coli (45). The expressed protein was readily
assembled in E. coli 30S subunits, 70S ribosomes, and in polysomes. Incorporation of
PSRP-1 did not interfere with protein synthesis (45). After protein chemical identification
of PSRP-2 and PSRP-3, we cloned and sequenced their cDNA's. Protein sequence of
PSRP-2 showed homology to stromal RNA-binding proteins which are associated with
mRNA processing (46) and mRNA stabilization (47). PSRP-3 sequence showed
homology to the translation product of ycf65, a hypothetical reading frame present in
plastid DNA and in the complete genome sequence of Synechococcus PCC 7002. Thus,
PSRP's 1, 2 and 3 are likely to be proteins that originated in photosynthetic bacteria.
These three PSRP's are acidic proteins with isoelectric points, PSRP-1 (pI=6.21), PSRP-2
(4.99), and PSRP-3 (4.93). Basic charges on most ribosomal proteins contribute to their
binding to the rRNA, whereas the acidic charge on the PSRP's may contribute to
additional protein-protein interactions in the plastid 30S subunit. In contrast, PSRP-4
(pI=11.79) is the most basic protein on spinach chloroplast 30S subunit. We have
previously reported (15) a sequence similarity in the N-terminal regions of PSRP-4 and a
30S subunit protein from Thermus thermophilus. We have now cloned and sequenced
PSRP-4 cDNA from spinach cDNA library. The complete sequence confirms the
homology to the T. thermophilus protein, but to no other bacterial proteins (detailed
results from PSRP's 2-4 will be published separately, ms in preparation). The summed
mass of the 4 PSRP's (67.6 kDa) corresponds to 16% of the total protein mass (429.3
kDa) of the chloroplast 30S subunit.
Plastid 30S subunit contains even more additional protein sequences as compared to E.
coli 30S subunit. Most chloroplast RP's are longer than their bacterial homologues (see
Fig. 7). The summed mass of plastid-specific NTE's, CTE's and PSRP's equals to 19%,
i.e. approximately one-fifth, of the total protein mass on the 30S subunit (429.3 kDa). In
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

25
contrast to this major change in protein composition, only minor changes occur in
chloroplast 30S subunit rRNA structure. For example tobacco chloroplast 16S rRNA (7)
(1,486 nt) is 56 nt shorter than E. coli 16S rRNA (1,542 nt). This truncation is due to the
absence or shortening of helices 6.4, 10, and 17.4 of E. coli 16S rRNA (Fig. 8 A, B).
Interestingly, all three of these helices, while apart in the secondary structure models, are
located together (Fig. 8C) in the 3-D folding model of E. coli 16S rRNA (48). Protein-
RNA cross-linking studies (49) and the crystal structure (5.5 A) of Thermus thermophirus
30S subunit (50) indicate the bottom part of 30S subunit structure as protein-poor.
Whether PSRP's occupy this part of the 30S subunit is speculative, but it is apparent that
a significant remodeling of the bacterial-type 30S subunit has occurred during the
evolution of chloroplast ribosome (localization of PSRP's in the 30S subunit structure is
currently under investigation, in cooperation with us, by cryoelectron microcopy, R.
Agrawal in J. Frank's laboratory).
The mRNA levels in chloroplasts remain relatively unchanged through light/dark
transition, whereas protein synthesis rates increase dramatically upon illumination (8,9).
Nuclear-encoded factors mediate light-regulated translation of plastid mRNA (10) and
nuclear mutantions affect polysome assembly and RNA metabolism (13). We propose
that PSRP's form a plastid-specific, "translational regulatory module" on the chloroplast
30S subunit that mediate some of these effects. Finally, based on the observation that
only 30% of chloroplast mRNA's contain RBS whereas a canonical Shine-Dalgarno
sequence is present in all chloroplast 16S rRNA's (7), and the fact that protein S1 that is
essential for the initial step of translation in bacteria (51, 52) is truncated in chloroplast
30S, a direct role for PSRP's in the initiation of plastid protein synthesis cannot be ruled
out (results on the identification of the 50S subunit PRP's will be publishes elsewhere, ms
in preparation).
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

26
Acknowledgment
We would like to thank the students, Marius Bartsch, Gerd Bolken, Anke Klippel, Klaus
Giese, Stephen Bantz, Wolfgang Wegloehner, and Juergen Schmidt (from Alap
Subramanian's Max-Planck Berlin lab) who made early contributions to this work. Ice
plant EST clone L0-67 was a gift from Dr. J. Cushman.
REFERENCES
1. Wittmann, H. G. (1982) Ann. Rev. Biochem. 51, 155-183
2. Planta, R. J. and Mager, W. H. (1998) Yeast 14, 471-477
3. Wool, I. G., Chan, Y-L., and Glück, A. (1995) Biochem. Cell Biol. 73, 933-947
4. Subramanian, A. R., Stahl, D., and Prombona, A. (1990) in The Molecular Biology of
Plastids (Bogorad, L., and Vasil, I. K., eds) pp. 191-215, Academic Press, New York
5. Harris, E. H., Boynton, J. E., and Gillham, N. W. (1994) Microbiol. Rev. 58, 700-754
6. Subramanian, A, R. (1993) Trends Biochem. Sci. 18, 177-180
7. Sugiura, M., Hirose, T., and Sugita, M. (1998) Annu. Rev. Genet. 32, 437-459
8. Fromm, H. , Devic, M., Fluhr, R, and Edelman, M. (1985) EMBO. J. 4, 291-295
9. Malonë, P., Mayfield, S. P., and Rochaix, J. D. (1988) J. Cell Biol. 106, 609-616
10. Yohn, C. B., Choen, A., Danon, A., and Mayfield, S. P. (1998) Proc. Natl. Acad. Sci.
U. S. A. 95, 2238-2243
11. Rochaix, J. D. (1996) Plant Mol. Biol. 32, 327–341
12. Stern, D. B., Higgs, D. C., and Yang, J. (1997) Trends Plant Sci. 2, 308–315
13. Barkan, A. (1993) Plant Cell 5, 389-402
14. Zhou, D. X. and Mache, R. (1989) Mol. Gen. Genet. 219, 204-208
15. Johnson, C. H., Kruft V., and Subramanian, A. R. (1990) J. Biol. Chem. 265, 12790-
12795
16. Schmidt, J., Srinivasa, B., Weglöhner, W., and Subramanian, A. R. (1993) Biochem.
Mol. Biol. Int. 29, 25-31
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

27
17. Graack, H. R. and Wittmann-Liebold, B. (1998) Biochem. J. 329, 433-448
18. Bartsch, M., Kimura, M., and Subramanian, A. R. (1982) Proc. Natl. Acad. Sci. U. S.
A. 79, 6871-6875
19. Hardy, S. J. S., Kurland, C. G., Voynow, P. and Mora, G. (1969) Biochemistry 8,
2897-2905
20. Kamp, R. M., Yao, Z. -J., Bosserhoff, A. and Wittmann-Liebold, B. (1983) Hoppe-
Seyler's Z. Physiol. Chem. 364, 1777-1793
21. Laemmli, U. K. (1970) Nature 227, 680-685
22. Subramanian, A, R. (1974) Eur. J. Biochem. 45, 541-546
23. Li, K. and Subramanian, A. R. (1975) Anal. Biochem. 64, 121-129
24. Walsh, M. J., McDougall J., and Wittmann-Liebold, B. (1988) Biochemistry 27,
6867-6876
25. Giese, K. and Subramanian, A. R. (1989) Biochemistry 28, 3525-3529
26. Franzetti, B., Zhou, D. X., and Mache, R. (1992) Nucleic Acids Res. 25, 4153-4157
27. Hellman, U., Wernstedt, C., Góñez, J., and Heldin, C-H. (1995) Anal. Biochem. 224,
451-455
28. Wellner, D., Panneerselvam,C., and Horecker, B. L. (1990) Proc. Natl. Acad. Sci. U.
S. A. 87, 1947-1949
29. GCG (1998) Wisconsin Package, Version 10.0, Genetics Computer Group, Madison,
Wisconsin.
30. Price, C. A., Reardon, E. M., and Lonsdale, D. M. (1996) Plant Mol. Biol. 30, 225-
227
31. Hirel, P. H., Schmitter, M.J., Dessen, P., Fayat, G., and Blanquet, S. (1989) Proc.
Natl. Acad. Sci. U. S. A. 86, 8247-8251
32. Schmidt, J., Herfurth, E., and Subramanian, A. R. (1992) Plant Mol. Biol. 20, 459-
465
33. Richter, S. and Lamppa, G. K. (1998) Proc. Natl. Acad. Sci. U. S. A. 95, 7463-7468
34. Franzetti, B., Carol, P., and Mache, R. (1992) J. Biol. Chem. 267, 19075-19081
35. Yoshikawa, A., Isono, S., Sheback, A., and Isono, K. (1987) Mol. Gen. Genet. 209,
481-488
36. Hitz, H., Schäfer, D., and Wittmann-Liebold, B. (1977) Eur. J. Biochem. 75, 497-512
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

28
37. Cumberlidge, A. G. and Isono, K. (1979) J. Mol. Biol. 25,169-189
38. Lindahl, M., Svensson, L. A., Liljas, A., Sedelnikova, S.E., Eliseikina, I. A.,
Fomenkova, N. P., Nevskaya, N., Nikonov, S. V., Garber, M. B., and Muranova, T.
A. (1994) EMBO J. 13,1249-1254
39. Bartsch, M. (1985) J. Biol. Chem. 260, 237-241
40. Weglöhner, W. and Subramanian, A. R. (1991) FEBS Lett. 279,193-197
41. Weglöhner, W., Kauschmann, A., and Subramanian, A. R. (1995) Biochem Mol Biol
Int. 36, 265-273
42. Weglöhner, W., Junemann, R., von Knoblauch, K., and Subramanian, A. R. (1997)
Eur. J. Biochem. 249, 383-392
43. Agalarov, S. C., Prasad, G. S., Funke, P. M., Stout, C. D., and Williamson, J. R.
(2000) Science 288,107-113
44. Tan, X., Varughese, M., and Widger, W. R. (1994) J. Biol. Chem. 269, 20905-20912
45. Bubunenko, M. G. and Subramanian, A. R. (1994) J. Biol. Chem. 269,18223-18231
46. Schuster, G. and Gruissem, W. (1991) EMBO. J. 10, 1493-1502
47. Nakamura, T., Ohta, M., Sugiura, M., and Sugita, M. (1999) FEBS Lett. 460, 437-441
48. Mueller, F. and Brimacombe, R. (1997) J. Mol. Biol. 271, 545-565
49. Brimacombe, R. (1995) Eur. J. Biochem. 230, 365-383
50. Clemons, W. M. Jr,, May, J. L., Wimberly, B. T., McCutcheon, J. P., Capel, M. S.,
and Ramakrishnan, V. (1999) Nature 400, 833-840
51. Subramanian, A. R. (1983) in Progress in Nucleic Acid Research and Molecular
Biology, Vol 28, Cohn, W. E. (ed.), pp. 101-142, Academic Press, New York
52. Sorensen, M. A., Fricke, J., and Pedersen, S. (1998) J. Mol. Biol. 280, 561-569
53. Arnold, R. J. and Reilly, J. P. (1999) Anal. Biochem. 269, 105-112
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

29
FIGURE LEGENDS
FIG. 1. Plastid ribosomal proteins (PRP’s) of spinach chloroplast 30S ribosomal
subunit separated by 2D-PAGE. A. Electropherogram, stained with Coomassie Blue.
TP30 (100 ug) was electrophoresed as described in ref 22. First dimension, at pH 5.0 in
8M urea; second dimension, at pH 6.7 in 0.2% SDS. Inset: a better resolution of the
dotted region.
B, Schematic diagram of the spots in A, with protein identification. Proteins were
identified as describes in Results. The minor spots, mostly in the upper portion of Fig.
1A, did not yield any significant sequence data when subjected to N-terminal
sequencing/identification (they are probably minor adhering impurities or multimeric
aggregates of PRP's). Because S6 spot showed multiple sequences (mixture of five
different forms, α-ε ), the spot was cut in two and sequenced: the upper half showed
mixed sequences of S6α,β, while the lower half yielded mixed sequences of S6γ-ε. Clean
sequences of the five forms of PRPS6 were obtained from HPLC experiment (Fig. 2).
Sequences of both S10α and S10β were present in the two spots of PSRPS10. (Prefix
PRP is omitted in Fig. 1B for convenience).
FIG. 2. Separation of spinach 30S PRP's by reversed-phase HPLC and examples of
peak identification by 2D-PAGE.
One mg of TP30 was resolved by a Vydac C18 column (250 mm x 4.6 mm) using a step
linear gradient of solvent #1: 0.1% TFA and solvent #2: 0.08% TFA in isopropanol (90%
#1/10% #2 from 0 to 10 min, 75% #1/25% #2 at 80 min, 55% #1/45% #2 at 250 min,
followed by a washing step, 20%#1/80% #2 at 260 min) at constant flow rate of 0.5
ml/min. The tailing PRP's are indicated by dotted lines underneath the identified names.
Fractions were analyzed by 2D-PAGE. Stained 2-D gel patterns for 3 pools (S21/S12, S9,
S4/PSRP-2/S8) and co-electrophoresis with TP30 (left, pool; right, pool + TP30) are
shown beneath HPLC-profile. The S6 forms (α-ε) were collected from the run at 2-
minute intervals and analyzed on protein sequencer, yielding clean sequences of the five
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

30
forms (Table I). Small peaks, marked a, S17 fragment (see Discussion), and b, probably
S14β.
FIG. 3. Histogram of N-terminal amino acid yields from sequencer run.
200 pmol of TP30 was subjected to 2D-PAGE/electroblotting and subjected to N-
terminal sequencing. Yield is calculated from the average PTH-AA recovery for the first
three sequencing cycles. Multiple forms of PRP's S1, S6, S10, S14, S18, and S19 are
indicated (see inset ). The average yield for all 30S PRP’s in the first three cycles was
27.9 pmols. Sine the volume for transfer at the PTH conversion step was 135 µl, of which
50 µl was injected into the C18 column for PTH-AA detection (the remaining sample
goes to a fraction collector), the actual recovery is about 37%. The low yields from spots
corresponding to S2, S3, S8, S11, PSRP-3, and zero yield from S9 are discussed in
Results Section.
FIG. 4. Protein sequence alignments for chloroplast PRP's S6 (alpha to epsilon), S9,
and S10 (alpha and beta). The spinach chloroplast sequence is at top. Black arrows
indicate experimentally derived transit peptide cleavage sites in spinach chloroplast PRP
precursors. Gray arrows indicate either alternative transit peptide cleavage sites or
additional processing points after import into plastid. Cleavage/processing points of the
multiple forms of S6 and S10 forms are shown. Dot indicates gap. Underlined sequences
are experimentally determined protein sequences (Table I). The sequence lengths of
mature proteins and precursors, and percents of sequence similarity (S) and sequence
identity (I) of the spinach sequence to other sequences are shown. The first 30 N-terminal
amino acids of Arabidopsis thaliana S6 precursor and the first 50 amino acids of ice plant
(Mesembryanthemum crystallinum) S10 precursor are omitted for representative
convenience. Four of the five forms of E. coli S6 (forms 2-5) are post-translationally
lengthened by C-terminal glutamylation (residues shown in italic). Sol, Spinacia
oleracea; Ath, Arabidopsis thaliana; Mcr, M. crystallinum (ice plant), Ppu, Porphyra
purpurea (brown alga); Syn, Synechocystis PCC6803 (photosynthetic bacterium); Eco,
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

31
Escherichia coli. Accession numberes of new sequences we reported are: Sol S6
(AF250383);; Sol S9 (AF242547); Sol S10 (P06506); and Mcr S10 (AF245665)
FIG. 5. LC/MS analysis of spinach chloroplast 30S ribosomal subunit proteins .
A. Relative abundance of charged molecules with mass to charge ratio (m/z) in the range
from m/z 400 to 2000 versus HPLC elution time. TP30 (60 pmol) was loaded on a
Microbore Vydac C18 column and resolved by a step linear gradient of solvent #1: 0.1%
TFA in 2% acetonitrile and solvent #2: 0.1% TFA in 90% acetonitrile (95% #1/5% #2 at
0 min, 90% #1/10% #2 at 9 min, 70% #1/30% #2 at 12 min, 65% #1/35% #2 at 15 min,
61% #1/39% #2 at 17 min, 53% #1/47% #2 from 30 - 32 min, 46% #1/54% #2 at 50 min,
45% #1/55% #2 at 51 min, 40% #1/60% #2 at 56 min, followed by a washing step, 5%
#1/95% #2 at 57 min).
B. A summary of the mass spectrum, m/z versus abundance, at the elution interval 30.0 –
30.5 min (indicated by arrow in A). Peaks represent individual charged ions; convoluted
charge and m/z values are given above the peaks.
C. Deconvoluted mass spectrum of shown in B. A protein mass of 23,280.8 Da is shown,
corresponding to PRPS4.
FIG. 6. Mass spectrometric evidence for -N-acetylation of PRPS9.
A. Deconvoluted mass spectrum from the LC/MS analysis of TP30 shown in Fig. 5A
indicating PRPS9 (17,105.1 Da) and PRPS7 (17,213.6 Da). The elution interval was
28.3-28.7 min.
B, Another deconvoluted mass spectrum of PRPS9 (17,105.6 Da), obtained by ESI MS
analysis of purified PRPS9. The HPLC pool containing PRPS9 (Fig.2) was analyzed on a
electrospray ionization mass spectrometer. The peak of 17,181.6 Da corresponds to an
adduct of PRPS9 with β-mercaproethanol (+ 76 Da ). A small peak of PRPS7 (17,214.0),
and an adduct of it (indicated with asterisk) with an unknown molecule (+ 45 Da), can
also be observed.. These adducts might arise by the different preparation methods used
for LC/MS and HPLC-purifications.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

32
C. MALDI-TOF MS analysis of "in gel" Lys-C digest of PRPS9. Individual isotopic ions
of 3 peptides are shown in the m/z region, 1,700-1,800. The corresponding peptide
sequences are shown above the ion peaks.
FIG. 7. Histogram comparing the mass changes between E. coli and chloroplast 30S
ribosomal subunit proteins.
Chloroplast protein masses are taken from Table II. For S10 mass the data for ice plant (α
form, 15,298.47 Da, Fig. 4) was applied: for spinach the corresponding data is presently
not available, see Results. Masses of E. coli ribosomal proteins are taken from a recent
MALDI-TOF MS determination (53).
FIG. 8. Secondary structure of tobacco chloroplast 16S rRNA and secondary
structure and 3-D model of E. coli 16S rRNA (locations of helices 6.4, 10 and 17.4).
A, Predicted secondary structure of the 5’ domain of tobacco16S rRNA. B, The secondary
structure of the corresponding domain of E. coli16S rRNA. Three helical regions in E.
coli 16S rRNA (shown in gray boxes) are shortened in the tobacco chloroplast 16S rRNA
structure (note: primary/secondary structure of chloroplast 16S RNA's is highly
conserved). The secondary structures of the two rRNA molecules are identical except in
the 5' domain. C, The locations of the missing chloroplast 16S rRNA helices (17.4, 6.4
and 10) are shown (arrows) in the 3D model for E. coli 16S rRNA (48); h, head; p,
platform; s, shoulder; b, body, in 30S subunit structure. (The 3-D model reproduced with
the kind permission of Dr. Brimacombe (48) and JMB)
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from
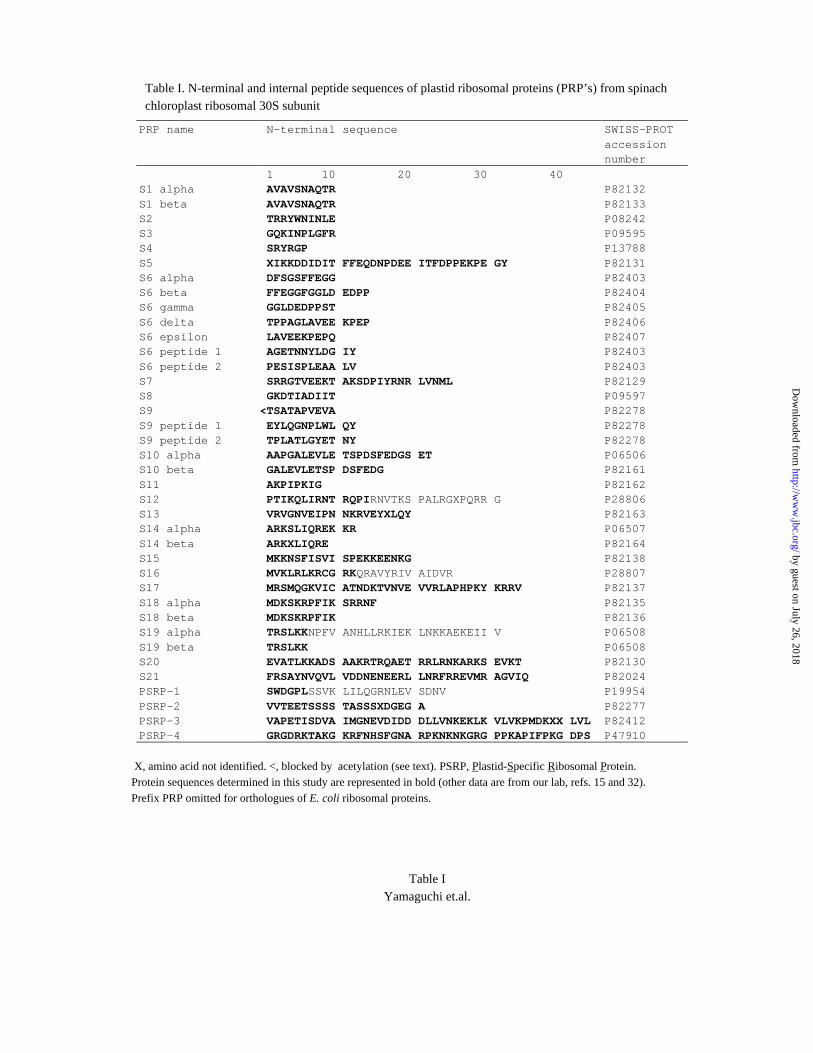
Table I. N-terminal and internal peptide sequences of plastid ribosomal proteins (PRP’s) from spinachchloroplast ribosomal 30S subunit
PRP name N-terminal sequence SWISS-PROTaccessionnumber
1 10 20 30 40S1 alpha AVAVSNAQTR P82132S1 beta AVAVSNAQTR P82133S2 TRRYWNINLE P08242S3 GQKINPLGFR P09595S4 SRYRGP P13788S5 XIKKDDIDIT FFEQDNPDEE ITFDPPEKPE GY P82131S6 alpha DFSGSFFEGG P82403S6 beta FFEGGFGGLD EDPP P82404S6 gamma GGLDEDPPST P82405S6 delta TPPAGLAVEE KPEP P82406S6 epsilon LAVEEKPEPQ P82407S6 peptide 1 AGETNNYLDG IY P82403S6 peptide 2 PESISPLEAA LV P82403S7 SRRGTVEEKT AKSDPIYRNR LVNML P82129S8 GKDTIADIIT P09597S9 <TSATAPVEVA P82278S9 peptide 1 EYLQGNPLWL QY P82278S9 peptide 2 TPLATLGYET NY P82278S10 alpha AAPGALEVLE TSPDSFEDGS ET P06506S10 beta GALEVLETSP DSFEDG P82161S11 AKPIPKIG P82162S12 PTIKQLIRNT RQPIRNVTKS PALRGXPQRR G P28806S13 VRVGNVEIPN NKRVEYXLQY P82163S14 alpha ARKSLIQREK KR P06507S14 beta ARKXLIQRE P82164S15 MKKNSFISVI SPEKKEENKG P82138S16 MVKLRLKRCG RKQRAVYRIV AIDVR P28807S17 MRSMQGKVIC ATNDKTVNVE VVRLAPHPKY KRRV P82137S18 alpha MDKSKRPFIK SRRNF P82135S18 beta MDKSKRPFIK P82136S19 alpha TRSLKKNPFV ANHLLRKIEK LNKKAEKEII V P06508S19 beta TRSLKK P06508S20 EVATLKKADS AAKRTRQAET RRLRNKARKS EVKT P82130S21 FRSAYNVQVL VDDNENEERL LNRFRREVMR AGVIQ P82024PSRP-1 SWDGPLSSVK LILQGRNLEV SDNV P19954PSRP-2 VVTEETSSSS TASSSXDGEG A P82277PSRP-3 VAPETISDVA IMGNEVDIDD DLLVNKEKLK VLVKPMDKXX LVL P82412PSRP-4 GRGDRKTAKG KRFNHSFGNA RPKNKNKGRG PPKAPIFPKG DPS P47910
X, amino acid not identified. <, blocked by acetylation (see text). PSRP, Plastid-Specific Ribosomal Protein.Protein sequences determined in this study are represented in bold (other data are from our lab, refs. 15 and 32).Prefix PRP omitted for orthologues of E. coli ribosomal proteins.
Table IYamaguchi et.al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

Table II. Mass spectrometry of spinach chloroplast 30S ribosomal proteins
LC/MS of TP30
PRP Sequence Elution Observed Mass HPLC pool HPLC pool SDS-PAGEg
name massa timeb massc differenced ESIe MALDIf
(Da) (min) (Da) (Da) (Da) (Da) (kDa)
S1α 40,427.67h ND ND 40,900 43.0S1β NA ND ND 36,890 36.8S2 26,564.55 h ND ND 27,053 29.2S3 24,796.74 38.0 24,950.2 h 153.5 ND 25,154 29.2S4 23,279.02 30.0 23,280.8 h 1.8 23,406 23,481 27.0S5 27,657.39 29.0 27,690.1 h 32.7 27,730 27,784 32.0S6α 16,324.30 35.0 16,326.7 h 2.4 ND 16,423 16.5S6β 15,830.84 35.0 15,830.1 -0.7 15,953 15,951 16.5S6γ 15,146.09 ND ND ND 16.5S6δ 14,278.23 ND ND ND 16.5S6ε 13,854.77 34.5 13,853.6 -1.2 ND 13,865 16.5S7 NA 27.5 17,212.7 h 17,214 17,243 18.5S8 15,367.85 31.5 15,443.8 h 75.9 ND 15,560 15.5S9 17,063.64 28.0 17,105.1 h 41.5 17,106 ND 18.2S10α NA ND ND ND 18.2S10β NA ND ND ND 18.2S11 14,795.21 h ND ND ND 15.5S12 13632.93 22.5 13,784.7 h 151.8 13,786 13,779 15.5S13 NA ND 14,628 h 14,900 15.5S14α/β 11,649.42 24.0 11,745.9 h 96.5 11,762 11,947 14.0S15 10753.60 26.0 10,830.1 h 76.5 10,847 10,847 12.5S16 10434.13 26.5 10,240.9 h -193.2 10,525 10,534 12.5S17 NA 21.5 12,247.8 h 12,354 12,354 16.5S18α NA ND 12,086 h 12,093 14.0S18β NA ND ND ND 14.0S19α/β 10,477.16 28.0 10,477.0 h -0.2 10,477 10,495 12.0S20 NA 27.0 12,917.8 h 12,922 12,920 15.0S21 NA 22.0 13,632.2 h 13,631 13,631 17.8PSRP-1 26,805.03 29.0 26,805.0 h 0.0 ND 26,943 31.0PSRP-2 21,665.02 30.5 21,664.6 h -0.4 21,666 21,658 25.0PSRP-3 13,794.02 h 34.5 ND ND ND 14.0PSRP-4 5,173.82 17.5 5,173.4 h -0.4 5,174 5,175 7.5 i
a Sequence mass calculated from translated DNA sequence using ‘peptidesort’ program (22). b Elution timefrom Fig. 5A. c Mass to the first decimal was obtained using advanced deconvolution d The observed mass
minus sequence mass. e Mass obtained by ESI MS of HPLC pools each containing identified PRP(s), seeFig. 2. f Mass obtained by MALDI-TOF MS of HPLC pools each containing identified PRP(s), see Fig. 2.g Taken from ref. 2. h These mass values were used for Fig. 7. i Measured by Tricine SDS-PAGE.NA, not available. ND, not determined. α/β, which form (α or β) not known.
Table IIYamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from
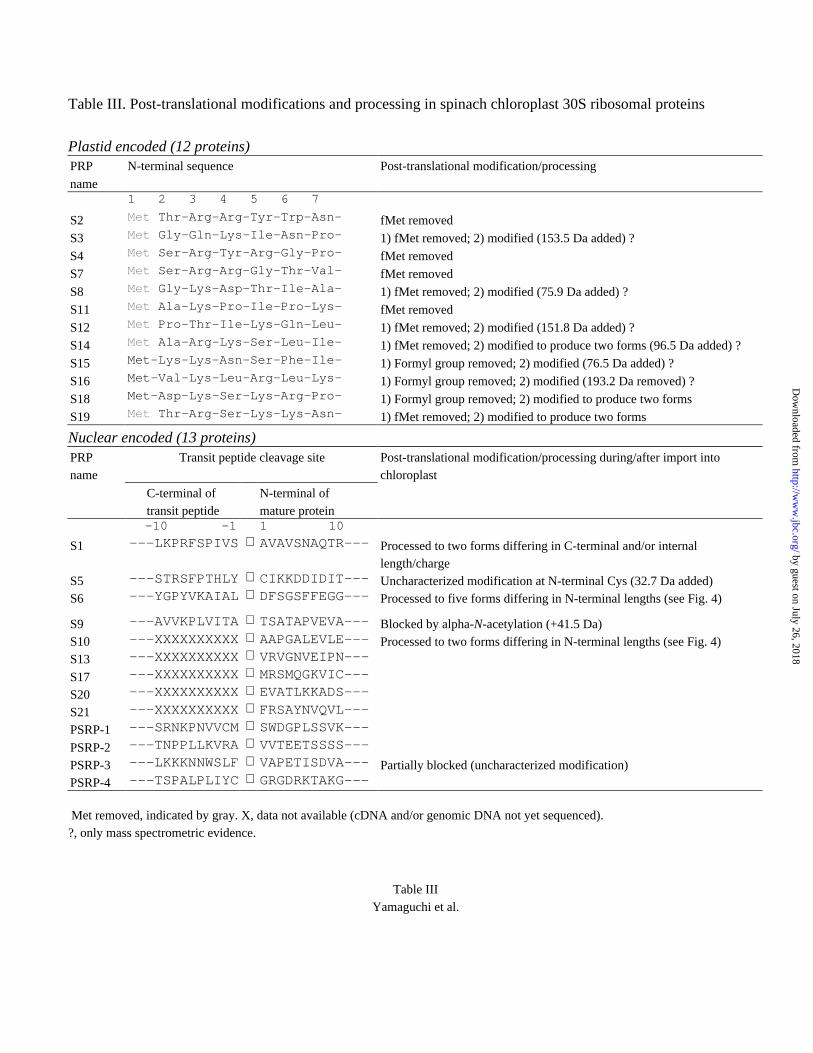
Table III. Post-translational modifications and processing in spinach chloroplast 30S ribosomal proteins
Plastid encoded (12 proteins)PRP N-terminal sequence Post-translational modification/processing
name1 2 3 4 5 6 7
S2 Met Thr-Arg-Arg-Tyr-Trp-Asn- fMet removed
S3 Met Gly-Gln-Lys-Ile-Asn-Pro- 1) fMet removed; 2) modified (153.5 Da added) ?
S4 Met Ser-Arg-Tyr-Arg-Gly-Pro- fMet removed
S7 Met Ser-Arg-Arg-Gly-Thr-Val- fMet removed
S8 Met Gly-Lys-Asp-Thr-Ile-Ala- 1) fMet removed; 2) modified (75.9 Da added) ?
S11 Met Ala-Lys-Pro-Ile-Pro-Lys- fMet removed
S12 Met Pro-Thr-Ile-Lys-Gln-Leu- 1) fMet removed; 2) modified (151.8 Da added) ?
S14 Met Ala-Arg-Lys-Ser-Leu-Ile- 1) fMet removed; 2) modified to produce two forms (96.5 Da added) ?
S15 Met-Lys-Lys-Asn-Ser-Phe-Ile- 1) Formyl group removed; 2) modified (76.5 Da added) ?
S16 Met-Val-Lys-Leu-Arg-Leu-Lys- 1) Formyl group removed; 2) modified (193.2 Da removed) ?
S18 Met-Asp-Lys-Ser-Lys-Arg-Pro- 1) Formyl group removed; 2) modified to produce two forms
S19 Met Thr-Arg-Ser-Lys-Lys-Asn- 1) fMet removed; 2) modified to produce two forms
Nuclear encoded (13 proteins)PRPname
Transit peptide cleavage site Post-translational modification/processing during/after import intochloroplast
C-terminal of N-terminal of transit peptide mature protein -10 -1 1 10
S1 ---LKPRFSPIVS ⇓ AVAVSNAQTR--- Processed to two forms differing in C-terminal and/or internallength/charge
S5 ---STRSFPTHLY ⇓ CIKKDDIDIT--- Uncharacterized modification at N-terminal Cys (32.7 Da added)S6 ---YGPYVKAIAL ⇓ DFSGSFFEGG--- Processed to five forms differing in N-terminal lengths (see Fig. 4)
S9 ---AVVKPLVITA ⇓ TSATAPVEVA--- Blocked by alpha-N-acetylation (+41.5 Da)S10 ---XXXXXXXXXX ⇓ AAPGALEVLE--- Processed to two forms differing in N-terminal lengths (see Fig. 4)S13 ---XXXXXXXXXX ⇓ VRVGNVEIPN---
S17 ---XXXXXXXXXX ⇓ MRSMQGKVIC---
S20 ---XXXXXXXXXX ⇓ EVATLKKADS---
S21 ---XXXXXXXXXX ⇓ FRSAYNVQVL---
PSRP-1 ---SRNKPNVVCM ⇓ SWDGPLSSVK---
PSRP-2 ---TNPPLLKVRA ⇓ VVTEETSSSS---
PSRP-3 ---LKKKNNWSLF ⇓ VAPETISDVA--- Partially blocked (uncharacterized modification)PSRP-4 ---TSPALPLIYC ⇓ GRGDRKTAKG---
Met removed, indicated by gray. X, data not available (cDNA and/or genomic DNA not yet sequenced).?, only mass spectrometric evidence.
Table IIIYamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

A
S1αS1β
S5 PSRP-1
PSRP-2
S10α,β
S3S2
S6
PSRP-3
S4S7
S8S9
S21
S17
S16
S10α,β
S20 S11
S13
S18βS15 S19α
S12
S18α
S14α
S14β
PSRP-4
BS19β
αε
→
Figure 1Yamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from
PSRP-4
S17
S21S12
S14α
S7S20S11S18β
S16S18α
S13
S15
S4PSRP-2S8
S5S19αS19β
PSRP-1
S9
S3
S1αS1βS10α
S10β
S2
0 40 80 120 160 200 240
Elution time (min)
S6ε→γ S6β→αPSRP-3
A22
0 (
)
0.5
1.0
0
ab
S9 dimer
S21 S9S12S8
S4
PSRP-2S4 dimerS8 dimer
Figure 2Yamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from
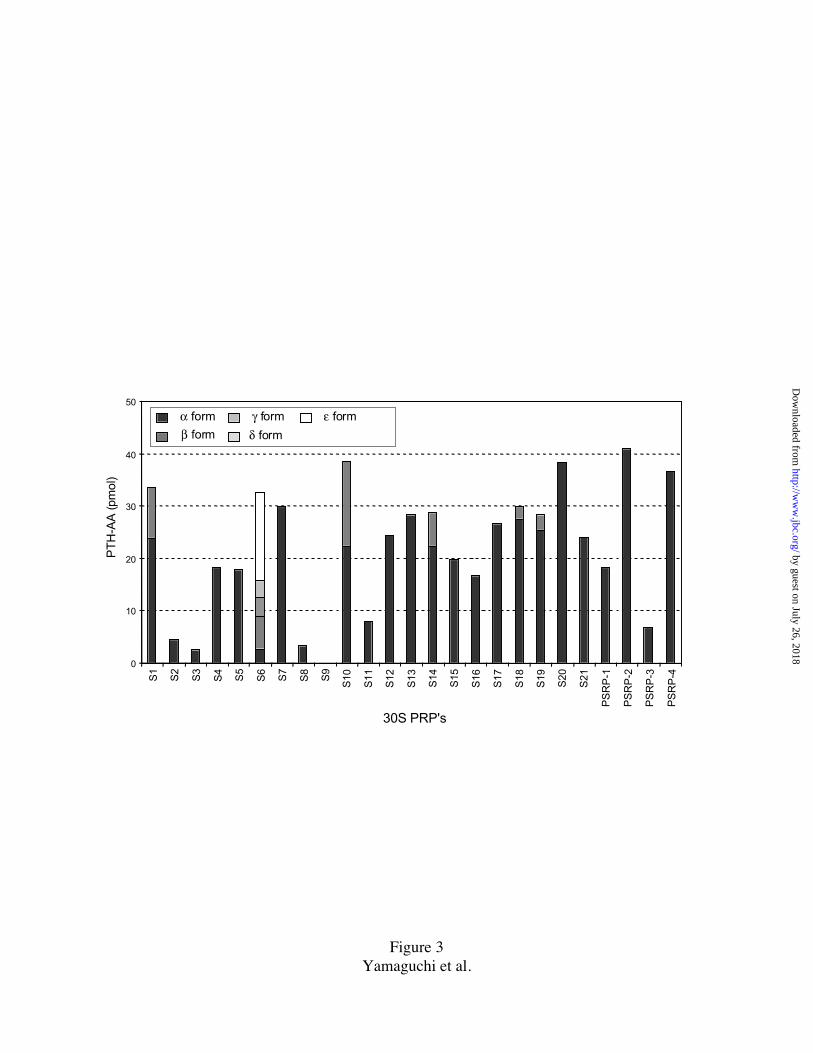
Figure 3Yamaguchi et al.
0
10
20
30
40
50
β form
α form
δ form
γ form ε form
PT
H-A
A (p
mol
)
30S PRP's
S1
S2
S3
S4
S5
S6
S7
S8
S9
S10
S11
S12
S13
S14
S15
S16
S17
S18
S19
S20
S21
PS
RP
-1
PS
RP
-2
PS
RP
-3
PS
RP
-4
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

S6Sol ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~N LKPSRHHYLS NYGP..YVKA IALDFSGSFF EGGFGGLDED PPSTPPAGL. .AVEEKPEPQ CPPGLRQYETAth VSNSTICPLP NVSSQPLLSF SHSLRPFISK SKPMCASIQK RDGSQFVVKS QALDFSGTFF EGGFGS.DDD PTSPSGSGVS TALEDKPEPQ CPPGLRQYETPpu ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~MLQTKNS QEIKLNWYETSyn ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~MLVNSYELEco ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~MRHYEI
ol MAVLRPDMTE DERLTLTQKY EELLVAGGAM YVEVFNRGVI PLAYSIKRKN KAGETNNYLD GIYLLFTYFT KPESISPLEA ALVTDDDVIR SSSFKIRKRKAth MAVLRPDMSE DERLGLTQKY EELLVAGGGM YVEVFNRGVI PLAYSIRKKN KAGETNTYLD GIYLLFTYFT KPESIVPLET VLTADDDIIR SSSFKIKKRKPpu VYLIKSDLNE DRTLCIINEY KSMLTNGGAK NIILQHRGRR HLSYSIKD.. .......YRD GIYVQVNYEG NGKLVQSFEK SLRFDENIIR YLT.....NKSyn MVILRPDLNE ERVSQEVTKY QEFLTNNAAE EVSVKVWGKR RLAYQIRRFN .........D GIYVLFNFNG EGQQIALIER DMRLNDNVMR FLSIKLTPEKEco VFMVHPDQSE .QVPGMIERY TAAITGAEGK IHRLEDWGRR QLAYPINKLH KAH....... ..YVLMNVEA PQEVIDELET TFRFNDAVIR SMVMRTKHAV
Sol Y~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~ α:146 β:141 γ:135 δ:126 ε:121 (>168)Ath YN~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~ (231) S:87.6% I:83.4%Ppu QNKDNKIEG~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~ (112) S:41.3% I:31.2%Syn PEKEKKAKAV AVEA~~~~~~ ~~~~~~~~~~ ~~~~~~~~~ (113) S:46.0% I:36.0%Eco TEASPMVKAK DERRERRDDF ANETADDAEA GDSEEEEEE 1:135 2:134 3:133 4:132 5:131 (131) S:38.3% I:27.7%
S9Sol ~~~~~~~~~~ FASLSFTSNL TPKPQTLPMA RTKP.FSL.S NPAVVKPLVI TATSATAPVE VAETADLEKF VKSRLPGGFA AQTVI..GTG RRKCAIARVVAth MASITNLASS LSSLSFSSQV SQRPNTISFP RANSVFALPA KSARRASLSI TAT.VSAPPE EEEIVELKKY VKSRLPGGFA AQKII..GTG RRKCAIARVVPpu ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~MSTELI KTRAIYSGTG RRKCSVAQVRSyn ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~MQANDS SNKVVYWGTG RRKAAIARVREco ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ MAENQYYGTG RRKSSAARVF
Sol LQEGTGKFII NYRDAKEYLQ GNPLWLQYVK TPLATLGYET NYDVFVKAHG GGLSGQAQAI SLGVARALLK VSASHRAPLK QEGLLTRDSR IVERKKPGLKAth LQEGTGKVII NYRDAKEYLQ GNPLWLQYVK VPLVTLGYEN SYDIFVKAHG GGLSGQAQAI TLGVARALLK VSADHRSPLK KEGLLTRDAR VVERKKAGLKPpu LVPGSGNLVI NGIPGESYLQ FSPNYLRVSY APLQVLGLLN QYDIHVNARG GGLTGQADAI RLGVARALCS INPENRSALK SEGYLTRDPK VKERKKYGLKSyn LVPGQGEVIV NGKPGEIYFN RIANYIQSLK APLETLGLEG EYNILVNAHG GGLTGQADAV KLGVARALCQ LSPENRQPLK AEGYLTRDPR AKERKKYGLHEco IKPGNGKIVI NQRSLEQYF. GRETARMVVR QPLELVDMVE KLDLYITVKG GGISGQAGAI RHGITRALME YDESLRSELR KAGFVTRDAR QVERKKVGLR
Sol KARKAPQFSK R 157 (>197)Ath KARKAPQFSK R (208) S:86.0% I:82.8%Ppu KARKAPQFSK R (137) S:57.7% I:52.6%Syn KARKAPQYSK R (137) S:60.7% I:54.1%Eco KARRRPQFSK R 129 (130) S:54.6% I:45.4%
S10Sol AAPGALEVLE TSPDSF.EDG SETMcr SSSSSNVRVF AAPEVLESQE TGPESYVEEG SETSALGIGA DSDQMAPKQK IRIKLRSYWV PLIEDSCKQI MDAARTTNAK TMGPVPLPTK KRIFCVLKSPPpu ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~MTVTQQPK IRIKLKAYNS SLLNTSCKKI VDTAERTNAI AVGPIPLPTK RRIYCVLRSPSyn ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~MATLQQQK IRIRLKAFDR RLLDTSCDKI VDTANRTNAA AVGPIPLPTK RKIYCVLRSPEco ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~~~~~~ ~~~~~MQNQR IRIRLKAFDH RLIDQATAEI VETAKRTGAQ VRGPIPLPTR KERFTVLISP
Mcr HVHKDARFHF EIRTHQRLID ILYPTAQTID SLMQLDLPAG VDVEVKL~ α:137 β:134 (197)Ppu HVDKDSREHF EIRSHRRIID IHQPSSQTID ALMKLNLPSG VDIEVKL~ (105) S:67.6% I:60.0%Syn HVDKDSREHF ETRTHRRIID IYQPSSKTID ALMKLDLPAG VDIEVKL~ (105) S:72.8% I:61.2%Eco HVNKDARDQY EIRTHLRLVD IVEPTEKTVD ALMRLDLAAG VDVQISLG 103 (103) S:66.7% I:52.0%
α β
γ δ εα β
Figure 4Yamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from
23280.8
A
B
C
Figure 5Yamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from
1720 1740 1760 1780
m/z
Arb
itrar
y In
tens
ity
200
600
800
B
AC
S9S9 + β-Me
S9
S7
S7*
1-AcTSATAPVEVAETADLEK-171-TSATAPVEVAETADLEK-17
63-EYLQGNPLWLQYVK-76
Figure 6Yamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

0
10000
20000
30000
40000
50000
60000
70000
S1
S2
S3
S4
S5
S6
S7
S8
S9
S10
S11
S12
S13
S14
S15
S16
S17
S18
S19
S20
S21
PS
RP
-1
PS
RP
-2
PS
RP
-3
PS
RP
-4
Ribosomal protein
Mas
s (
Da)
Chloroplast
E. coli
Figure 7Yamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

CBA
helix 17.4
helix 6.4
helix 10
helix 17
helix 6
helix 10
h
sp
b
Chloroplast E. coli
Figure 8Yamaguchi et al.
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from

Kenichi Yamaguchi, Klaus von Knoblauch and Alap R. Subramanianof an organelle ribosome (chloroplast)
The plastid ribosomal proteins (1): identification of all the proteins in the 30S subunit
published online June 28, 2000J. Biol. Chem.
10.1074/jbc.M004350200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on July 26, 2018http://w
ww
.jbc.org/D
ownloaded from