the organization and function of potential regulatory elements
TRANSCRIPT

Volume 16 Number 17 1988 Nucleic Acids Research
Conservation of hfcstone H2A/H2B intergene regions: a role for the H2B specific element inV divergent transcription
Richard A.Sturm*, Stephen Dalton+ and Julian R.E.Wells
Department of Biochemistry, University of Adelaide, PO Box 498, Adelaide, South Australia, 5000Australia
Received January 6, 1988; Revised and Accepted May 31, 1988 Accession nos X07756-XO7761 (incl.)
ABSTRACTThe organization and function of potential regulatory elements associated with the promoters
of chicken H2A and H2B genes pairs have been examined. The Intergene regions of sixdispersed and divergently-transcribed H2A/H2B gene pairs contain several extremely wellconserved and spaced blocks of sequence homology. Adjacent coding regions are on average342 base-pairs apart. Respective TATA boxes are separated by 180 base-pairs and within thisconfined region there are four CCAAT boxes and a previously Identified 13 base-pair H2B-speclflc element (H2B-box) which has homology to the octamer motif present In a number ofgene promoter/enhancer elements. Transcription of H2A and H2B genes from wild-type andmutant constructs was measured In transient assays by transfectlon into HeLa cells, and Inpermanently transformed clanal cell lines. In vitro separation of the two genes at a uniquelntergenlc site significantly decreased transcription of each gene. This suggested that theH2A/H2B gene parrs contained overlapping promoters. Deletion or point mutagenesls of theH2B-speclflc element decreased the levels of H2B and the H2A transcripts Indicating that thissequence Is a common regulatory element of both genes In the divergent-pair conflgeratlon.
INTRODUCTION
Genomlc organization studies of hlstone genes have revealed an apparently random gene order
for higher eukaryotes (1,2). However, upon closer examination of the hlstone gene families of
several species, trends do become apparent. A common theme Is the close association of
H2A/H2B and H3/H4 genes. These gene pairs are often divergently transcribed (see gene
organizational Tables in reference 1).
Extensive studies of hlstone gene organization In the chicken genome show a general disorder,
with no long range repeat being evident (3). There are, however, preferred associations, most
notably H1 with H2A and H2B genes. The extent of the H2A/H2B association has been
examined In this report at the DNA sequence level, and conserved sequence elements 5' to
coding regions have been identified. Of particular Interest was a highly conserved promoter
element (5'-CTGATTTGCATAC/G-31 originally described by Harvey and co-workers (4)
positioned six bases 5' to chicken H2B gene TATA boxes, but absent in all other hlstone genes.
The central core region of this motif (ATTTGCAT) was subsequently Identified (In the same or
Inverted orientation] In lmmunoglobulln heavy and light-chain promoters (5,6) and In the
lmmunoglobulin heavy-chain and SV40 enhancer (5.7) In U2 (8,9) and some Ul (10.11) small
nuclear RNA (snRNA) promoters and most recently In the herpes simplex virus thymldlne klnasr.
© IRL Press Limited, Oxford, England. 8 5 7 1
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
promoter (12). In several Instances, this 'octamer' motif has been shown to be essential for
efficient (13) or cell specific promoter activity (14,15).
In conjunction with the examination of the promoter architecture of H2A/H2B gene pairs, an
expression study of one of the gene pairs was initiated. Evidence is presented which suggests
that the highly conserved intergene region is related to the co-ordinated expression of the
H2A/H2B divergent gene pair. The conserved H2B-speciflc element apparently conslstutes a
common regulatory element which Is required for the efficient expression of both genes fri ud».
MATERIALS AND METHODS:
Analysis of Divergent H2A/H2B Gene Pair Genomlc Clones
The relative position of the divergent H2A/H2B gene pairs in the chicken histone gene clusters
can be found In reference 3. The name of each pair presented here derives from the name of the
plasmld or lambda clone from which it had been subcloned; L and R refers to the left and right
side of the clone. Restriction fragments of each divergent gene pair were subcloned Into M13
vectors for sequencing. All sequence analysis was by the M13 dldeoxy chain termination
method (16), and the sequences have been deposited with the EMBL Data Library (accession
numbers J0O864, X07756-X07766). Data analysis utilized the Staden computer programs.
M13 Site Directed Mutagenesls
The methods of Adelman et al., (17) and, Chan and Smith (18) were utilized for deletion and
substitution mutagenesis of intergene promoter elements. Synthetic ollgonucleotldes were as
follows: H2B-box deletion, 5'-dTATTTATAGCCCCAAGCATTCCTTTC-3'; H2B-box base
substitutions, 5'KlGATCCrAG<;CCCrCTAGGCIVVCriXlACWW\GCATTCCrTTCG-3': and CCAAT box
deletion 5'drCGCACTCrTTTCGGAACTCGA-3\ Screening for mutant phage was performed by in
situ plaque hybridization using each of the 3aP-labelled mutagenesls ollgonucleotldes. Plaques
that hybridized strongly relative to negative parental controls, following high temperature
washes, were picked and single-strand M13 preparations sequenced with the H2B primer
extension oligomer (26-mer, (19) see below) to confirm the mutant genotype.
Cell Culture and HeLa Cell Transfection
HeLa cells were grown in Dulbecco modified Eagle medium supplemented with 10% fetal calf
serum. A 10 cm dish of HeLa cells seeded at 8 x 10s - 1 x 106 cells per plate was transfected
with 5 ng of test DNA, together with 20 |ig of carrier DNA, using the calcium phosphate
precipitation technique (20). The calcium precipitate was left on the cells for 18-20 hours after
which the medium was changed and the cells left for a further 48 hours. Cytoplasmlc RNA was
extracted from the cells by lyslng them in 500 ul of 0.5% NP4O, 10 mM NaCl. 10 mM Tris-HCl
pHS.O, 10 mM EDTA followed by centrifugatlon to remove nuclei. The supernatant was
collected and subjected to phenol/chloroform extraction and ethanol precipitation. RNA was
treated with RNase-free DNase I (Promega Biotech) before primer extension analysis. Permanent
clonal cell-lines were generated by methods previously described (21).
8572
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
Primer Extension Analysis
Synthetic DNA primers were generously provided by Dr. D. Skmgle and S. Rogers. The primers
used were as follows:
H2A primer 5'-dACK;GACTXlAACACTCAGAGAGCAAAC-3'
H2B primer S'-dGGCTCGGGCATAGTGGCACAACGCGC-S'
HI primer S'-dGGCGGGAGCGGTCTCGGACATCGCGG-a1
Primers were 5' end-labelled with T4 pohmucleotlde klnase and y-32P-ATP, Ing of primer mixed
with RNA, heated to 80°C for 3 minutes then allowed to anneal at 42°C In 200 mM NaCl, 10 mM
Trls-HCl pH 8.3 for 1.5 hours. Following hybridization, samples were Incubated with reverse
transcriptase (1 hour, 42°C) In 60 mM NaCL 10 mM Tris-HCl pH 8.3, 10 mM dlthlothreitol, 500
mM dNTPs and 8 mM MgCLj (22). Extension products were electrophoresed on 6%
poh/acrylamlde sequencing gels and detected by exposure to X-ray film. Quantltatlon of
radioactivity displayed on autoradlograms was performed on a Zelneh Soft Laser Densltometer
(Model SL-5O4-XL). Densttometry was carried out In duplicate, on two exposures of the
autoradiogram.
Preparation of Chick Embryo RNA
5-day chick embyro RNA was prepared by a guanldlne-HCl method (23). The RNA was size
fractionated on sucrose gradients and fractions containing hlstone mRNA (about 9S) collected.
Transcription In Isolated Nuclei
Nuclei were Isolated from cells and pulse-labeled with a-32P-UTP as described previously (24).
Nascent 32P-labeled RNA was extracted from nuclei and used to probe fllter-immobillzed hlstone
gene Inserts as described (21).
RESULTS:
DNA Sequence Organization of Six Chicken H2A/H2B Divergent Pairs
In all. thirty-six core plus six H1 hlstone genes have been mapped In the chicken genome
(3,25,26.27). Amongst the general disorder of these genes in dispersed groups, six closely
associated H2A/H2B gene pairs have been found; one pair In each of the recomblnants
pCH3.3E/p7AT, pCH3.5E, pCH22.OB and XCH1-10 and. two pairs In the recombinant
pCH 11 .OE (see reference 3 for details). Gross mapping Indicated that these gene pairs,
although dispersed In different clusters, maintained a very similar organization. All six
lntergene regions have now been sequenced and relevant details are presented In Figure 1.
Because we wish to emphasize the nature of conserved promoter elements only the sequence
between the TATA boxes Is presented here. Complete sequence data are available on request
(19) and have been submitted to the EMBL Data Library (accession numbers J00864 and
X07756-X07766). Proteins encoded by the H2B genes of pCH3.3E/p7AT, pCH3.5E and XCH1-
10 correspond to the H2B.1 variant described by Urban et al., (28). The H2B subtype variant
encoded In pCH22.0B has yet to be assigned. Insufficient DNA sequence data are available to
8573
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
£ubctoneJCM12.CB
paa.urani « i
OtetancaB«tw*en
ATQ341U l351353
1S3
H2A H2B
TTT 10 . WOMXC 13
13
2 1
t*
H
14
.cow 2
.CCWTB., 2
Q*^*TC. . J
.ccu». 2CCUTO. 2
1 ..coin 411 ..caoow.. 301 . cam . 421 ..ccun . 431 ..ccun .. 431 T V T .. 31
..CCMTO.
. 0OUH0-.
. .CGU».
CCUKI .
coucr
comic..
XX
XX
XX
22
21
21
. .CTur i iguma. (
c-nuoTTOCiTJC 0
. CT0CTTK33TW.. S
.cnjcmoatTM . 0
CTtjacmxnc. 0ciouiwon. 7
PCH3.3E
* * * * * * * * * *AGAGGGGCTATAAATAAATGOCTA* *PCH22.0BGCanTmTXXCTTCCTGCTGGGGTOGTGOGGCACITrT^TO
ATTGa3GQ3TTAACCMTCGGAAO^QXGTAGAATACa>g>ATTGAACAGCGCGA
CTAATTTCCATACCGCCTCTATAAGTAC• * * * * * * * * • •PCH3.5E
CGCGTTGTGGTG^OXXACAAACAOGGACTrcCAOXMTCCCAGnx^
GAGCTQCTGCTTTGCATAGC0CCGCTATAAAGOCG* * * * * * * * * * * *
pCHll .OE l e f t
GOXETTTGTGGTX>aXCACAAACACGAACTTCCAGCOU^
OGC>GCTCCKrTITC»TAGCGCOGCTATAAAGGCE* * • • • * * • * * * *
pCHll.OE r i g h tC I T T
CTAATCTGCATACAGOGACTATAAAAGAGG* * * * * * * * * * * *
CH1-10
CCCCTTTTATCCCITCa>Q330GGGACTAG0GCOCGGTGATTGGCTQaTACAA^
ATAACAGXCTATAAAAA3GGG
8574
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
classify the pCHl l.OE-encoded proteins. Both of the H2A subtype variants reported In the
literature can be Identified from the sequence of the divergent pair H2A genes. pCH3.3E/p7AT,
pCH22.OB. pCH3.5E and as far as the DNA sequence of pCH 11.OE left and right H2A genes
extend, all correspond to the H2A. 1 subtype variant (28). The XCH1-10 H2A gene encodes a
H2A.2 protein variant
The lntergene sequence of five of the six divergently transcribed H2A/H2B gene pairs differ
considerably (Figure IB) but several highly conserved regions are apparent when the sequences
are compared (Figure LA). The genes are very closely linked; the coding regions are, on average,
342 base-pairs apart; the TATA boxes are separated by only 180 base-pairs and are m the same
relative position for each gene pair. A striking feature observed when the sequence of each
confined lntergene region Is examined Is the presence of four motifs with homology to the
CCAAT box (29). These are all regularly spaced with three of them having the consensus
"CCAATG" (CAT1. CAT2 and CAT3 of Figure IB) positioned at approximately 40, 90 and 110
bases upstream of and directed towards the H2B TATA boxes. The fourth CCAAT box (CAT4 of
Figure 1A) with a consensus sequence "GCCTATC" Is an average of 28 bases upstream from the
H2A TATA boxes.
The position of the pCH3.3E/p7AT TATA box designated In Figure 1 differs from that assigned
by D'Andrea et al. (30). The only A-rich region to be found at a position consistent with the other
divergent pairs Is a run of three A residues. This Is an extremely poor match to the consensus
TATA box whereas the region suggested by D'Andrea Is an extremely good match. However, the
mRNA cap site of this H2A gene has been determined (from primer extension analysis presented
below and SI mapping (19)) and Is consistent with this assignment of AAA as the TATA box.
Two of the lntergene regions have almost identical nucleotlde sequences. DCH3.5E H2A/H2B
and the left H2A/H2B pair of pCHl l.OE have only seven base differences between them. The
sequence of the pCH3.3E H2AmRNA 5' untranslated sequence Is almost the same as that of the
pCH3.5E/pCHl l.OE L H2A genes. All three of these pairs are In gene clusters that are
immediately next to one another on the chicken chromosome (3). Gene conversion or gene
duplication processes may have been involved in generating these homologles.
A fifth region of homology with almost exact conservation for 13 bases (5'-
CrCATITGCATAC/G-31 located 6 bases upstream of the H2B TATA box (Figure LA, H2B box),
has been described previously (4). This element is closely associated with the H2B gene
Figure 1: Architecture of chicken H2A/H2B lntergene regions.(A) Six divergent H2A/H2B intergene regions are compared and a consensus of conservedregions and distances compiled. The average distance between divergent pair ATG Initiationcodons are indicated In addition to the subclone from which they were derived (see reference 3).(B) The sequences of the H2A/H2B lntergene regions between the TATA elements outlined In (A)are listed (accession numbers X07756-X07761). The TATA box is underlined by a continuousline, CAT 1,2,3 and 4 elements are underlined by dashed lines and the H2B-box is underscoredby asterisks.
8575
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
CAT< CAT3 CAT2 CAT! H2B dot
M 1 2
3 4
6 8 -«-H2B H2B - • •
H1
C
«-H2A H2A
M 1
68 — i H2B
H1
H2A
Figure 2: Expression of H2A and H2B genes as separate entitles.(A) A diagrammatic representation of the HI. H2A and H2B genes within the 7 kb EcoRIgenomlc subfragmeni used for expression analysis. The H2A/H2B lntergene region has been
8576
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
expanded to show the relative position and orientation of CCAAT boxes and position of thelntergene Xhol site with respect to these elements. B=BamHI, E=EcoRI. S=SmaI. X=XhoI.(B) RNA was Isolated from HeLa cells 72 hours after being transfected with. Lane 1. pJ7E: Lane2, pJ7E-H2A (H2Agene deleted); Lane 3. pJ7E; Lane 4. pJ7E-H2B (H2B gene deleted) andanalysed by primer extension analysis. A 68 base pair pBR322 Hpall marker Is Indicated (M).(C) Lane 1 represents quantitative primer extension analysis using the three 26 base chickenhlstone specific primers (Materials and Methods) on size fractionated 5 day chick embryo RNA.A pBR322 Hpall marker lane Is shown to Indicate the different extension lengths of the threeprimers; H2A extends to a major band at 49 bases. HI to 58 bases and H2B to 68 bases.
promoters of most organisms (31) and contains the octamer homology lATTTGCAT) that Is a
functional element of a number of gene promoter/enhancer elements (5-12).
Expression of a Chicken H2A/H2B Gene Pair and Mapping of the Divergent Promoter Region.
The gene pair chosen for study was the one for which the complete gene sequences were first
available (4,19,30). A 7kb genomal EcoRI fragment from X.CH-05 (3) was subcloned Into the
vector pAT153 (32) for propagation and the done named p7AT. The 7kb Inserts from p7AT
derivatives were re-subcloned Into pJL4- (an SV40 late replacement vector with the Ncol-Ncol
region excised. 33) for transient expression In HeLa cells. In addition to containing the complete
H2A/H2B divergent pair an H1 gene Is contained within the Insert (34, see Figure 2A). Gene-
speclflc 26 base synthetic oligonucleotlde primers were designed such that the length of In vitro
primer extension products from each gene transcript present In the same mixture could be
readily Identified by size. Typically, major bands of 49, 58 and 68 bases are observed for H2A,
H1 and H2B extension products respectively. When the primers are used In excess the assay
becomes quantitative (22) and the level of transcription from each gene can be measured.
The data In lanes 1 and 3 of Figure 2B show directly comparable quantitative primer extension
results on RNA derived from HeLa cells transfected with pJ7E (the p7AT Insert cloned Into
pJL4-). The relative level of each gene's expression in HeLa cells Is similar to that of endogenous
transcripts in 5-day chick embryo RNA (19, Figure 2C). Using primers of similar specific activity
it can be seen that H2B and HI transcripts are well expressed whereas the H2A gene is poorly
expressed.
The H2A and H2B genes of p7AT are each contained entirely on separate Xhol restriction
fragments (3, Figure 2A). Moreover, the lntergene region of the two genes is split by a Xhol
recognition sequence and expression of the HI gene of p7AT is unaffected by Xhol digestion (19).
By Independent removal of each Xhol fragment of p7AT. it was possible to examine the
expression of each gene in the absence of the other. The two single gene deletions constructs
leave one CCAAT box present In p7AT minus H2A (containing HI and H2B genes) and three in
p7AT minus H2B (containing H1 and H2A genes). In each case the level of H1 gene
transcription serves as an internal control (35). The fragments containing the separated H2A
and H2B genes were transferred to the pJL4- vector and tested in the HeLa cell transient assay.
Significant decreases in the levels of H2B (about 17-fold) and H2A (7-fold) transcription are seen
when these separated genes are expressed In HeLa cells Figure 2B, lanes 2 and 4 respectively:
8577
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
TABLE 1SUMMARY OF TRANSIENT ASSAY DATA
TransfectedConstruct
pJ7E
PJ7E-H2A
DJ7E-H2B
pJ5E
pJCATf
PJH2BSJ;
Relative Steadv-State
H2B
1.0
0.06
-
1.0
0.13
0.11
mRNA Levels
H2A
1.0
-
0.15
1.0
0.47
0.32
Table 1: Summary of data for transient expression of constructs In HeLa cells.HeLa cells In exponential growth phase were transfected with the constructs described andsteady-state RNA levels generated from the exogenous genes detected by primer extensionanalysis. Results are presented relative to the expression of wild-type genes, with the HI geneas an internal reference control, as determined by laser densitometry of autoradlograms. Eachexperiment was performed In duplicate on at least two separate occasions.
Table 1). These experiments demonstrate that the two genes appear to have overlapping
promoter elements. It is perhaps surprising that H2B transcription should decrease so
markedly considering that CATl and the H2B box sequence remain Intact in the construct
tested. However, this result is not confined to the HeLa expression system, but is also seen in
Xenopus oocyte injection experiments (19).
Mutagenesls of the H2A/H2B Intergene Region
Separation of the H2A/H2B genes in the Intergene region affects the level of both transcripts,
but does not determine whether the two genes share any of the conserved DNA sequence
elements of the Intergene region. A site-directed mutagenesls approach was used to
independently remove or modify conserved tntergenlc regions to see if the transcription of either
gene was affected. A 5 kb BamHI fragment containing all of the HI, H2A/H2B gene system in
the 7 kb EcoRI fragment of p7AT (see Figure 2A), was excised and cloned into M 13mp8.
Synthetic DNA deletlon/mutagenesls primers (see Materials and Methods) were used to
independently delete or mutate the CATl and H2B box elements of the intergene region (see
Figures 1A and 2A). Mutagenesls of templates was confirmed by dl-deoxy sequencing using the
H2B 26-base extension primer as a sequencing primer (data not shown).
Functional testing of each 5 kb BamHI mutated fragment was performed by separately
8578
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
1 2 3 M
IH2B
H2A-»
CATf Construct5-TTCTCOAGTTCCQA[CCAATQ|AAAOAGTGCGAAAQO-3- pJ5E5-TTCTCGAGTTCCGA AAAAAGTGCGAAAGG-3' pJCAT-j"
HJB-Box'5'-QAATGCTT|CTCATTTQCATAQ|AQGQQCTA-3' pJ5E
6-GAATGCTTICTCAGTAGCCTAGIAQOGOCTA-31 pJH2B» • • b t
Figure 3: The H2B-speclflc element Is required for efficient expression of H2A/H2B gene pairs invivo.The 5 kb BamHI fragment containing HI, H2A and H2B genes was subcloned Into M13mp8 anddeletion or substitution mutagenesls was performed before being Inserted Into pJL4-. HeLa cellswere transfected with. Lane 1, pJ5E (wild type fragement); Lane 2, pJCATr (CAT1 box deleted);or Lane 3, pJH2BgE (mutated H2B box). A pBR322 Hpall marker lane Is shown (68 basemarker). Sequences resulting from site directed mutagenesls are shown and are compared tocorresponding unmutated (wild type) sequences In the native 5 kb BamHl fragment ('Indicatesbase substitutions in the H2B box).
subclonlng them into pJL4- and assaying following transfectlon Into HeLa cells. RNA was
extracted three days after cells were transfected and subjected to primer extension analysis
using the three gcne-speclfic primers (Figure 3). Expression of the exogenous H2B and H2A
genes was decreased by approximately 9-fold and 3-fold, respectively, as a result of base
substitutions within the H2B-bax (Figure 3. lane 3; Table 1). This was the first Indication that
the H2B-speciflc element Is required for efllclent expression of the divergently transcribed H2A
gene tn vivo. Notably, the decrease In levels of H2A mRNA m HeLa cells was not as severe as
detected when the H2B gene was removed by gross deletion (Figure 2B). This Implies that other
elements wtthln the lntergene region, besides the H2B-spedllc motif, are required for efficient
expression of both genes. Consistent with this Is the observation that deletion of the CAT1 box
8579
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018
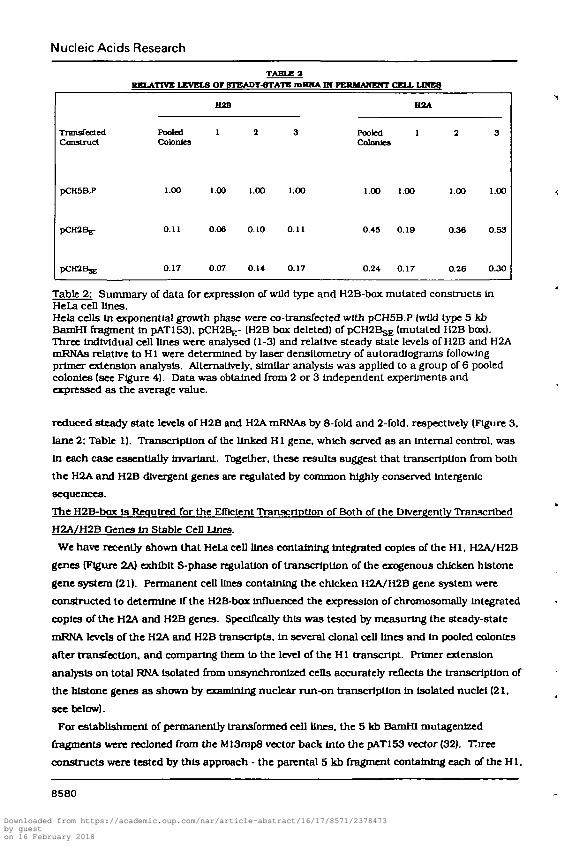
Nucleic Acids Research
TABLE 2HELATTVg LEVEL8 OF BTEADT-8TATB mBWA Pf PKMMAWEWT CELL LIHE8
TransfectedConstruct
pCH5BJ>
P C H 2 B E -
PCH2BSE
PootedColonies
1.00
0.11
0.17
1
1.00
0.06
0.07
2
1.00
0.10
0.14
3
1.00
0.11
0.17
PooledColonies
1.00
0.4S
0.24
H2A
1
1.00
0.19
0.17
2
1.00
0.36
0.26
3
1.00
0.53
0 3 0
Table 2: Summary of data for expression of wild type and H2B-box mutated constructs InHeLa cell lines.Hcla cells In exponential growth phase were co-transfected with pCH5B.P {wild type 5 kbBamHI fragment in pAT153). pCrreBj;- (H2B box deleted) of pCfttBgj. (mutated H2B box).Three Individual cell lines were analysed (1-3) and relative steady state levels of H2B and H2AmRNAs relative to HI were determined by laser densltometry of autoradlograms followingprimer extension analysis. Alternatively, similar analysis was applied to a group of 6 pooledcolonies (see Figure 4). Data was obtained from 2 or 3 independent experiments andexpressed as the average value.
reduced steady state levels of H2B and H2A mRNAs by 8-fold and 2-fold, respectively (Figure 3,
lane 2: Table 1). Transcription of the linked H1 gene, which served as an Internal control, was
In each case essentially Invariant. Together, these results suggest that transcription from both
the H2A and H2B divergent genes are regulated by common highly conserved lntergenlc
sequences.
The H2B-box Is Required for the Efficient Transcription of Both of the Divergently Transcribed
H2A/H2B Genes In Stable Cell Lines.
We have recently shown that HeLa cell lines containing Integrated copies of the H1. H2A/H2B
genes (Figure 2A) exhibit S-phase regulation of transcription of the exogenous chicken his tone
gene system (21). Permanent cell lines containing the chicken H2A/H2B gene system were
constructed to determine If the H2B-box Influenced the expression of chromosomally Integrated
copies of the H2A and H2B genes. Specifically this was tested by measuring the steady-state
mRNA levels of the H2A and H2B transcripts, in several clonal cell lines and in pooled colonies
after transfectton, and comparing them to the level of the HI transcript Primer extension
analysis on total RNA Isolated from unsynchronized cells accurately reflects the transcription of
the hlstone genes as shown by examining nuclear run-on transcription In isolated nuclei (21,
see below).
For establishment of permanently transformed cell lines, the 5 kb BamHI mutagenlzed
fragments were recloned from the M13mp8 vector back into the pAT153 vector (32). TTiree
constructs were tested by this approach - the parental 5 kb fragment containing each of the HI,
8580
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018
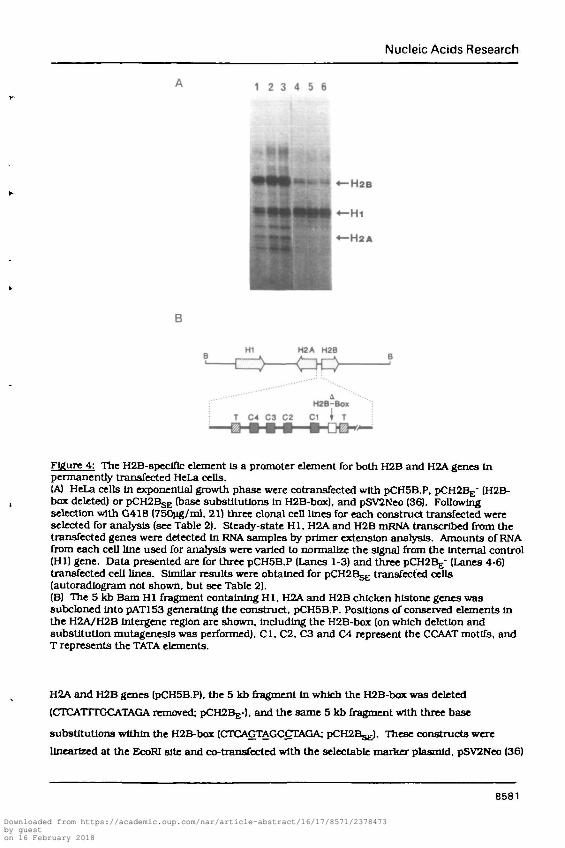
Nucleic Acids Research
1 2 3 4 5 6
B
H2A H2B
H2B-BOX
T C< C3 C2 C1 • T
Figure 4: The H2B-speclflc element Is a promoter dement for both H2B and H2A genes Inpermanently transfected HeLa cells.(A) HeLa cells In exponential growth phase were cotransfected with pCH5B.P, pCH2Bg- (H2B-box deleted) or pCH2BsE (base subsUtutlons In H2B-box), and pSV2Neo (36). Followingselection with G418 (750ng/ml, 21) three clonal cell Unes for each construct transfected wereselected for analysis (see Table 2). Steady-state HI, H2A and H2B mRNA transcribed from thetransfected genes were detected In RNA samples by primer extension analysis. Amounts of RNAfrom each cell line used for analysis were varied to normalize the signal from the internal control(HI) gene. Data presented are for three pCH5B.P (Lanes 1-3) and three pCMBg" (Lanes 4-6)transfected cell lines. Similar results were obtained for pCTTCBsg transfected cells(autoradlogram not shown, but see Table 2).(B) The 5 kb Bam HI fragment containing HI, H2A and H2B chicken hlstone genes wassubcloned Into pAT153 generating the construct, pCH5B.P. Positions of conserved elements Inthe H2A/H2B lntergene region are shown, including the H2B-box (on which deletion andsubstitution mutagenesls was performed). Cl, C2, C3 and C4 represent the CCAAT motifs, andT represents the TATA elements.
H2A and H2B genes (pCH5B.P), the 5 kb fragment In which the H2B-box was deleted
(CTCATTTGCATAGA removed; PCH2BJ;-). and the same 5 kb fragment with three base
substitutions within the H2B-box (CTCAGTAGCCTAGA; pCraBgg). These constructs were
linearized at the EcoRI site and co-transfected with the selectable marker plasmld, pSV2Neo (36)
8581
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
Into HcLa cells by the calcium phosphate precipitation method (20). Following selection of
transformed, G418-reslstant cell lines, levels of transcripts generated from the exogenous
chicken hlstone genes were determined by primer extension analysis (Figure 4) and the results
are summarized In Table 2.
Deletion of the H2B-speclflc motif resulted in a distinct decline in H2B steady-state mRNA
levels for the three pCH2Bj;- transfected cell lines tested (Figure 4, Table 2). A similar effect was
detected with pCFQB^ cell lines as a consequence of point mutations being inserted Into the
H2B-box region. Therefore, a decrease In H2B mRNA levels caused by deletion of the H2B-box
was not a result of general promoter disruption as base substitutions within this region, which
conserved overall promoter architecture and the relative spatial arrangement of promoter
elements, had a similar effect to the gross deletion. Compared to H2B transcript levels In
"parental" cell lines (pCH5B.P-transfected), this represented a decrease ranging from 6 to 17-fold
(see Table 2) as determined by laser densltometry of autoradlograms. In addition to the analysis
of Individual clonal cell-lines, groups of six colonies for each DNA transfectlon were pooled RNA
extracted and primer p-r^mahm analysis carried out. Expression of HI, H2A, and H2B genes
displayed a <tfm»ar pattern when compared to Individual cell-lines. Transcript levels of the
divergently transcribed H2A gene were also reduced, generally by a factor of 2 to 6 fold (see
Table 2). It Is, therefore, apparent that the H2B-box exerts a regulatory effect on both the H2A
and H2B genes in vivo, thus confirming the transient assay results presented In Figure 3 and
summarised in Table 1.
The H2B-Box Modulates Transcription of H2B and H2A Genes in the Divergent Pair.
To directly show that the effect of mutating the H2B-bax on decreasing H2B and H2A steady
state mRNA levels was at the transcrlptlonal level, relative rates of gene transcription were
compared m cell lines which had been permanently transformed with pCH5B.P (wild-type H2B-
box) or pCH2BgE (base substitutions In H2B box). This was done by Isolating nuclei from these
cell lines and pulse labelling them with a-32P-UTP. Nascent labelled transcripts were hybridized
to filter-immobilised complimentary hlstone gene Inserts In order to quantitate the relative rates
of H2A and H2B gene transcription over the pulse labelling Interval using transcription of the
HI gene as an Internal control (21). The contribution of endogenous (human) gene transcription
was determined by probing the same DNA dots with 32P-RNA extracted from nuclei which had
no transfected chicken histone gene copies (untransfected HeLa cells). Under the stringent
hybridisation conditions employed, the signal generated from endogenous genes was generally
less than 5% of that detected In pCH5B.P-transfected cell lines (see Figure 5). In cell lines
transfected with pCH5B.P. transcripts from exogenous HI, H2A and H2B genes were readily
detected over background levels from human genes (Figure 5). Transcription of the chicken H2B
gene In cell lines transfected with the mutant construct pCH2BgE was decreased by a factor of
about 5-fold relative to transcription of the unmutated gene In pCH5B.P-transfected cells.
Importantly, the decrease In H2B gene transcription was accompanied by a 4-fold decrease In
8582
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
1.0 W 10
1.2 # 3.1
0.9 # 0.8IIFigure 5: The H2B-spedflc element Is required Tor efficient transcription of divergent H2B andH2A genes.Nuclei from untransfected (HeLa), pCH5B.P-transfected (wild type fragment In pAT153) orpCH2BgE transfected (base substitutions In H2B-box) cells were pulse labelled with a-^P-UTPand 3x10* cpm of labelled RNA was hybridized to 2.5 ng of filter Immobilized, single strandedchicken historic gene Insert (see Materials and Methods). Relative levels of hybridized ^P-RNAwere quantltated by scintillation spectrometry and each set expressed relative to levels of HI•^P-transcrlpts (Internal control: assigned value of 1.0). Each experiment was performed Induplicate. The top. middle and bottom row of dots corresponds to RNA used to probe HI, H2Band H2A DNA dots respectively.
transcription of the divergent H2A gene. This is consistent with the decrease in levels of H2B
and H2A steady state mRNAs detected by primer extension analysis (see Table 2) and Indicates
that the H2B-bax Is a transcriptlonal element shared by both genes.
DISCUSSION:
The apparently random organization of hlstone genes in the chicken genome belles the
extraordinarily similar architecture of the six divergently transcribed H2A/H2B gene pairs which
occur In different clusters of the hlstone multlgene family. The coupling of these H2A and H2B
genes cannot be mistaken for a remnant of evolution of the gene family In the chicken. DNA
sequence analysis of five of the six pairs shows that some 5' regions have been evolving
independently; the H2B box was first recognized In this context (4).
The selection pressure keeping the majority of these two genes together Is possibly the
stolchlometrtc requirement of the proteins coded for by each gene. The equlmolar appearance of
H2A and H2B In the nucleosome can be achieved by assembly from appropriately sized pools or
by a regulatory control operating during nucleosome assembly. It Is possible that a shared
promoter region for each gene pair enables coordinate control of H2A and H2B mRNA
production. Thus each protein might accumulate at an appropriate level for nucleosome
assembly.
Several lines of evidence support the contention that the H2A/H2B lntergene region
constitutes an overlapping promoter. This was first suggested by the observation that
separation of these genes at an tntergenlc site resulted in a marked decrease In the expression
8583
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
of both genes (Figure 2B). Experiments aimed at delineating sequence elements required for the
efficient transcription of both genes have Identified a dual role for the H2B-bax. Although
transcrtptlon of both genes was significantly decreased when the H2B box was either deleted or
contained point mutations, this effect was less than that when the two genes were separated at
the Xho I site (Table 1) These observations are consistent with there being additional lntergenlc
elements, besides the H2B box, required for the efficient tn vivo expression of the gene pair.
While deletion of the CAAT1 box had an apparent effect on expression of H2B and H2A genes, it
Is recognized that such a deletion may have Interrupted overall promoter architecture and may
not accurately reflect the role of this sequence In the overlapping promoter. Whether disruption
of any other CAAT element In the intergene region has an affect on transcription similar to the
H2B-box is unknown.
A consistent relative decrease in H2B and H2A transcript levels (compared to the internal
control HI gene) were detected in clonal cell lines transfected with H2B-box deleted or base-
substituted constructs. As the H2B-box Influences transcription from H2B and H2A genes
(when in the divergent configuration) It Is likely that this element functions as a transcrlptlonal
amplifier of both genes, perhaps by temporally modulating levels of transcrlptlonal initiation
during S-phase of the cell cycle. Indeed, this sequence has been shown to be involved in the
cell-cycle regulation of an isolated human H2B gene, where point mutations within the H2B-box
had no effect on basal transcription but prevented an increase in utilization of the promoter
during DNA synthesis (37). As yet, no subtype-specific sequence element has been implicated In
the S-phase regulation of H2A genes, but in the case of the chicken H2A/H2B genes studied
here this could be provided by coupling of the H2B-box as a promoter element of the H2A gene
as well as the H2B gene of each pair to mediate the Induction of transcription during DNA
replication.
The H2B-box contains within Its sequence the octamer homology located in the control regions
of several other non cell cycle regulated genes (5-12): the octamer motif has been found tn
sequence elements implicated In controllng basal transcription (8-12), tissue spedfic-
promoter/enhancer function (5-7, 14, 15, 38) and DNA replication (39). It is possible that
various levels of combinatorial control, such as the sequence context in which the octamer motif
Is embedded (40), operate to provide differential gene regulation in these cases. The overall
conservation of Intergene sequences In H2A/H2B gene pairs (Figure 1) and an apparent role of
the H2B box element in influencing both H2A and H2B gene expression may explain the
preference for the paired arrangement of these genes which has been retained throughout
evolution (1).
We thank Richard D'Andrea and Leeanne Coles for the gifts of the chicken hlstone genomal
clones described in the text. RA.S. and S.D. were supported by Australian Commonwealth
8584
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
Postgraduate Research Awards and this work was funded by a grant to the Department of
Biochemistry establishing a Commonwealth Special Research Centre for Gene Technology.
Present addresses: *Cold Spring Harbor Laboratory, PO Box 100, Cold Spring Harbor, New York,NY 11724, USA and +Research Institute for Molecular Pathology, A-1030 Wien, Dr. Bohr-Gasse 7,Austria
REFERENCES1. Maxson, R. Conn, R. Kedes, L. and Mohun. T. (1983). Ann Rev. Genet. 17. 239-277.2. HentscheL C.C. and Blmstlel. MX. (1981). Cell 25. 301-313.3. D'Andrea, RJ. , Coles. L.S., Lesnikowskl, C. Table. L. and Wells. J.RE. (1985). Mol.
CellBiol. 5,3108-3115.4. Harvey, RP.. Robins, A.J. and Wells, J.RE. (1982). NucL Acids Res. 10, 7851-7863.5. Falkner, F.G., and Zachau, H.G. (1984). Nature 310, 71-74.6. Parslow. T.G.. Blair. D.L.. Murphy, WJ. and Granner, D.K. (1984). Proc. Natl. Acad.
Sd. USA 81. 2650-2654.7. Falkner, F.G., Macikat, R and Zachau, H.G. (1986). Nud. Acids Res. 14, 8819-8827.8. Ares, M., Jr.. Mangin, M. and Weiner. A.M. (1985). Mol. CelL Blol. 5. 1560-1570.9. MattaJ. I.W., Lienhard. S., Jirtchy, J. and DeRobertis, E.M. (1985). Nature 316, 163-
167.10. Ciliberto. G.. Buckland. R, Cortese, R and Phillpson, L. (1985). EMBO J. 5. 1537-
1543.11. KroLA..Lund. E. and Dahlberg, J.E. (1985). EMBO J. 4, 1529-1535.12. Parslow, T.G., Jones, S.D., Bond, B. and Yamamoto. K.R (1987). Science 235, 1498-
1501.13. Slve, H.L., Heintz. N.. and Roeder. RG. (1986). Mol. Cell. BloL 6, 3329-3340.14. Dreyfus, M., Doyen, N. and Rougen. F. (1987). EMBO J. 8, 1685-1690.15. Wirth. T., Staudt, L. and Baltimore, D. (1987). Nature 329. 174-178.16. Sanger, F., Nlcklen, S. and Coulson, A.R (1977). Proc. Natl. Acad. ScL USA 74,
5463-5467.17. Adelman, J.P., HayfUck, J.S.. Vasser, M. and Seeburg, P.H. (1983). DNA 2. 183-193.18. Chan, V-L., and Smith, M. (1984). NucL Acids Res. 12, 2407-2419.19. Sturm, RA. (1985). Ph.D. Thesis, University of Adelaide. Adelaide. South Australia.20. Graham. F.L., and van der Eb, AJ. (1973). Virology 52. 456-467.21. Dalton. S. and Wells. J.RE. (1988). EMBO J. 7, 49-56.22. McKnight, S.L., Gavis, E.R. Kingsbury. R and Axel, R (1981). Cell 25, 385-398.23. Brooker. J.D., May, B.K. and Elliott, W.H. (1980). Eur. J. Blochem. 106, 17-24.24. Dalton. S., Coleman, J .R and Wells, J.RE. (1986). Mol. CeU BioL 6, 601-606.25. Wang. S-W.. Robins, AJ.. D'Andrea. R and Wells. J.RE. (1985). Nud. Adds Res. 13.
1369-1387.26. Grandy. D.K and Dodgson, J.B. (1987). Nud. Adds Res. 15, 1063-1080.27. Coles, L.S., Robins, AJ., Madley. L.K. and Wells, J.RE. (1987). J. Blol. Chem. 262,
9656-9663.28. Urban. M.K.. Franklin. S.G. and Zweidler. A. (1979). Biochemistry 18. 3952-3960.29. Breathnach, R and Chambon. P. (1981). Ann. Rev. Biochetn. 50. 349-383.30. DAndrea. R. Harvey. R, and Wells, J.RE. (1981). NucL Acids Res. 9, 3119-3128.31. Wells. D.E. (1986). NucL Acids Res. 14, r l l9-rl49.32. Twigg, AJ . and Sherratt. D. (1980). Nature 283, 216-218.33. Gough. M.N., Metcalf. D.. Gough.J., Grail, D. and Dunn, A.R (1985). EMBO J. 4,
645-653.34. Coles, L.S. and Wells, J.RE. (1985). NucL Acids Res. 13. 585-594.35. Younghusband. H.B., Sturm. R and Wells. J.RE. (1986). Nud. Adds Res. 14, 635-
644.36. Southern, E.M. and Berg, P. (1982). J. Mol. Appl. Genet 1. 327-341.
8585
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018

Nucleic Acids Research
37. LaBella, F., Stve, H.L., Roeder, RG. and Helntz, N. (1988). Genes and Development 232-39.
38. Gerster, T.. Matthias. P.. Thall, M., Jlricny, J. and Schaflher, W. (1987). EMBO J. 6,1323-1330.
39. Pruljn. GJ.M., van Drlel, W. and van der Vllet, P.C. (1986). Nature 322, 656-659.40. Sturm, R, Baumruker, T.. Franza, B.R Jr. and Herr, W. (1987). Genes &
Development 1, 1147-1160.
8586
Downloaded from https://academic.oup.com/nar/article-abstract/16/17/8571/2378473by gueston 16 February 2018