the hydrolysis of a-d-glucosides by amyloglucosidase … · the hydrolysis of a-d-glucosides by...
TRANSCRIPT

THE JOURNAL OFBIOLOQICAL CHEMISTRY Vol. 237, No. 4, April 1962
Printed in U.S.A.
The Hydrolysis of a-D-Glucosides by Amyloglucosidase from
AspergiZZus niger *
JOHN H. PAZUR AND KJELL KLEPPE
From the Department of Biochemistry and Nutrition, University of Nebraska, Lincoln 3, Nebraska
(Received for publication, October 6, 1961)
The purification of an amyloglucosidase from Aspergillus niger by an ion exchange chromatographic procedure was recently described (1). Examination of the purified amyloglucosidase by ultracentrifugal and gel electrophoretic methods has shown that the amyloglucosidase was indeed separated from the other carbo- hydrases of A. niger. Further, the electrophoretic and ultra- centrifugal patterns indicate a high degree of homogeneity in the amyloglucosidase preparation. The molecular weight of the amyloglucosidase was found to be 97,000, which is large in com- parison with values for other fungal carbohydrases (2, 3). In earlier studies (4), it was shown that the amyloglucosidase was capable of hydrolyzing the a~-~-(1 -+ 4) and the a-~-(1 -+ 6) glucosidic linkages in oligosaccharides and presumably in starch and glycogen. There was, however, a marked difference in the rates of hydrolysis of the two types of bonds. Since the CY-D- (1 --f 3) glucosidic linkage may also be present in these polysac- charides (5, 6), it was of interest to determine whether this type of linkage was hydrolyzed by the amyloglucosidase. The latter was achieved by subjecting nigerose, a disaccharide of glucose containing an a-~-(1 -+ 3) bond (7), to the action of amyloglu- cosidase and examining the reaction mixture for the production of D-glucose. In these experiments, it was found that the nig- erase was hydrolyzed to D-glucose and that the rate of hydrolysis of the a-~-(1 ---f 3) bond was intermediate between the rates for the cr-D-(1 -+ 4) and the cr-D-(1 -+ 6) bonds.
Results of earlier studies with oligosaccharides containing different glucosidic linkages (8) and labeled with Cl4 in the re- ducing moiety (1) have demonstrated that the amyloglucosidase acts as a typical a-D-glucosidase, in that a single glucosyl unit is liberated from the nonreducing end of the substrate. Several types of a-D-glucosides have now been tested in an attempt to determine the substrate range of this glucosidase. Included in this list are maltulose (4.c-D-glucopyranosyl-D-fructose) , malto- bionic acid (4-a-D-glucopyranosyl-D-gluconic acid), sucrose (a-D-glucopyranosyl-P-D-fructofuranoside), tUranOSe (&x-D-gh- copyranosyl-D-fructose), trehalose (cr-D-glucopyranosyl-cr-D-glu- copyranoside) , 3-cr-D-glucopyranosyl-D-arabinose, l-&D-&l- copyranosyl-D-glycerol, phenyl cr-D-glucoside, and methyl a+D-glucoside. Paper chromatographic examination of the en- zymic digests of these compounds have shown that all of the com- pounds were susceptible to amyloglucosidase. However, there was a large variation in the relative rates of hydrolysis of the glu- cosides, perhaps indicating differences in the ability of the various
* Published with the approval of the Director as paper No. 1145, Journal Series, Nebraska Agricultural Experiment Station. Supported in part by a grant from the Miles Laboratories, Inc., Elkhardt, Indiana.
glucosides to form an enzyme-substrate complex with the amy loglucosidase.
EXPERIMENTAL PROCEDURE AND RESULTS
Enzyme Source-The amyloglucosidase was isolated from an enzyme preparation of Aspergillus nigerl by chromatography on diethylaminoethyl (DEAE) cellulose as described in detail in an earlier publication (1). The enzyme solution as obtained from the chromatographic column contained 0.12 mg of nitrogen per ml and possessed an activity of 300 units (1) of amyloglucosidase activity per ml. For analysis in the ultracentrifuge, the enzyme solution was concentrated lo-fold in the cold room, and, as a result, the solut.ion used for the analysis contained approximately 0.7 y0 protein. This solution was dialyzed against 0.1 M sodium chloride for 24 hours before use. The concentrated enzyme also was employed in the gel electrophoretic experiments. For the studies on the hydrolysis of the various glucosides, enzyme solu- tions containing 15 units per ml, 150 units per ml, and 300 units per ml were prepared by appropriate dilution of the amyloglu- cosidase preparation with 0.05 M citrate buffer, pH 4.8.
cr-o-Glucosides-Methyl or-D-glucoside was prepared by re- fluxing D-glucose in methyl alcohol in the presence of hydrogen chloride. The methyl a-D-glucoside was crystallized from ethyl alcohol and subjected to chromatography on cellulose columns in order to remove traces of glucose from the preparation (9). The melting point of the crystalline methyl a-D-glucoside was 166-167”.
Phenyl cy-D-glucoside was prepared from 1,2,3,4,6-penta- acetyl+D-glucose and phenol by a published procedure (10). The final product was recrystallized twice from absolute ethyl alcohol and melted at 172-173”.
Pure maltose (4-cr-D-glucopyranosyl-D-glucose) was available in the laboratory and had been isolated from a /?-amylase hy- drolysate of starch (11). The specific rotation of the maltose was +130”.
Isomaltose (6-or-D-glucopyranosyl-D-glucose) was prepared from maltose with the transglucosylase of Aspergillus oryzae (11). The product, when examined by paper chromatography, did not contain D-glucose, maltose, or other reducing compounds. The specific rotation of the isomaltose was +120”.
Nigerose (3.a-D-glucopyranosyl-D-glucose) was also isolated from the enzymolysate of maltose with the transglucosylase of A. oryzae by adsorption on charcoal and elution with ethyl alcohol (7). The compound was further purified by a paper
1 The enzyme preparation is available under the registered trademark of Diazyme from the Miles Chemical Company, Clif- ton, New Jersey.
1002
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

April 1962 J. H. Pazur and K. Kleppe
FIG. 1. Sedimentation velocity photographs obtained in the ultracentrifuge with the purified amyloglucosidase of A. niger
chromatographic procedure (11). The final product yielded one spot on a paper chromatogram, indicating that it was free from reducing contaminants.
Maltobionic acid was prepared by a published procedure by oxidation of maltose (12). To insure that traces of unoxidized maltose were removed from the preparation, the maltobionic acid was further purified by the paper chromatographic method.
Maltulose was prepared by alkali isomerization of maltose (10). Unchanged maltose in the reaction mixture was hydro- lyzed to n-glucose by treatment of the neutralized reaction mix- ture with amyloglucosidase for a short period. The resulting solution, consisting of n-glucose, a small amount of n-fructose, and maltulose, was chromatographed on paper and the maltulose was extracted with water. The sample was dried in a desiccator. An acid hydrolysate of the purified maltulose contained n-fruc- tose and n-glucose in equal molar ratios.
Glycerol glucoside (1-oc-n-glucopyranosyl-n-glycerol) (13) was kindly provided by Dr. E. Hehre, Albert Einstein Medical College, New York. Arabinose glucoside (3-ar-o-glucopyranosyl- n-arabinose) (14) was furnished by Dr. H. Isbell, National Bureau of Standards, Washington, D. C. The turanose and trehalose were products of Nutritional Biochemicals Corporation, Cleveland, Ohio, and sucrose was a product of Mallinckrodt Chemical Works, New York.
Sedimentation and Diffusion Measurements-The sedimenta- tion coefficients at four different concentrations of the enzyme, (3000 units per ml, 1500 units per ml, 750 units per ml, and 375 units per ml) were obtained in a Spinco ultracentrifuge, model E. The enzyme solutions were prepared as described above, and measurements were made at 59,780 r.p.m. Approximately 10 photographs were obtained for each experiment at intervals ranging from 8 minutes to 32 minutes. A photograph of typical patterns is reproduced in Fig. 1. The sedimentation coefficients were evaluated from the data by the method described by Schachman (15). Extrapolation of a plot of the values to in- finite dilution yielded a value for the sedimentation coefficient (s20,J of 5.44 s.
A diffusion coefficient for the amyloglucosidase was also ob- tained/by use of the ultracentrifuge. A synthetic boundary cell was used, and the speed was 4000 r.p.m. Two different experi- ments were performed with the same concentration of enzyme, and the values obtained for the diffusion coefficients were in good agreement. The average value for D~o,~ was 5.05 x lo-’ cm* per second. A value of 5.57 x 10-r cm2 per second was obtained for the diffusion coefficient D20,,,,, in a moving boundary cell in an Aminco-Stern electrophoresis apparatus.
Estimation of molecular weight for a protein from sedimenta- tion velocity and diffusion coefficient values also requires that
the partial specific volume of the protein and the density of the solvent be known. The density of 0.1 M sodium chloride at 20.0” is, within a small error, unity. Data in the literature in- dicate that the partial specific volumes of protein solutions range between 0.70 ml per g to 0.75 ml per g. On the basis of the literature values for proteins similar to amyloglucosidase, a partial specific volume for amyloglucosidase of 0.74 ml per g has been employed in the calculation. These values were used in the conventional formula for calculating molecular weights (15). The molecular weight for amyloglucosidase was found to be 97,000 f 5%.
Electrophoresis on Acrylamide GeP-The apparatus used for electrophoresis was similar to that originally described by Smithies (16). The acrylamide gel was prepared from acryla- midc essentially as outlined in the literature (17). Gel strips of 25.5 x 3.2 X 0.6 cm and a buffer of 0.1 M Tris-EDTA-borate, pH 8.9, were employed in the experiments. The sample of enzyme was introduced into the gel by means of a rectangular strip of cellulose acetate paper which had been dipped into the enzyme solution. The electrophoresis was conducted for 13 hours at 300 volts, at 10 ma, and at 4”. On completion of the experiment, the gel strips were removed from the trays and dipped for 1 minute into a solution of 0.1 y0 ponceau S3 in meth- anol-water-glacial acetic acid (5 : 5 : 1 by volume). The gel strips were then washed for 3 to 4 days in methanol-water-glacial acetic acid (5:5: 1 by volume) and finally in 20 y0 glycerol-water for 1 day. The protein areas appeared as pink bands in a colorless gel. Amyloglucosidase was found as a single sharp band at approximately 6 cm from the origin toward the positive pole. The nonpurified enzyme sample of Diazyme, when ex- amined by this method, yielded several bands. A paper electro- phoretic pattern for a purified amyloglucosidase of A. niger has already been published (2) and further substantiated the con- clusion that the amyloglucosidase preparation consists of a single protein species.
Hydrolysis of a-n-Glucosides by Amyloglucosidase-The mode of action of amyloglucosidase was first studied qualitatively by identifying the hydrolytic products from the various substrates by paper chromatographic methods. In general, the following procedure was used in these experiments. Approximately 10 pmoles of compound were dissolved in 0.1 ml of enzyme solution (300 units per ml) buffered at pH 4.8 with 0.05 M citrate-diso- dium phosphate buffer. This solution was incubated at room temperature, and a small crystal of thymol was added to prevent
2 The collaboration of Dr. R. Dam in this experiment is grate- fully acknowledged.
3 A product of the National Aniline Division, Allied Chemical Corporation, New York 6, New York.
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Hydrolysis of a-o-Glucosides Vol. 237, No. 4
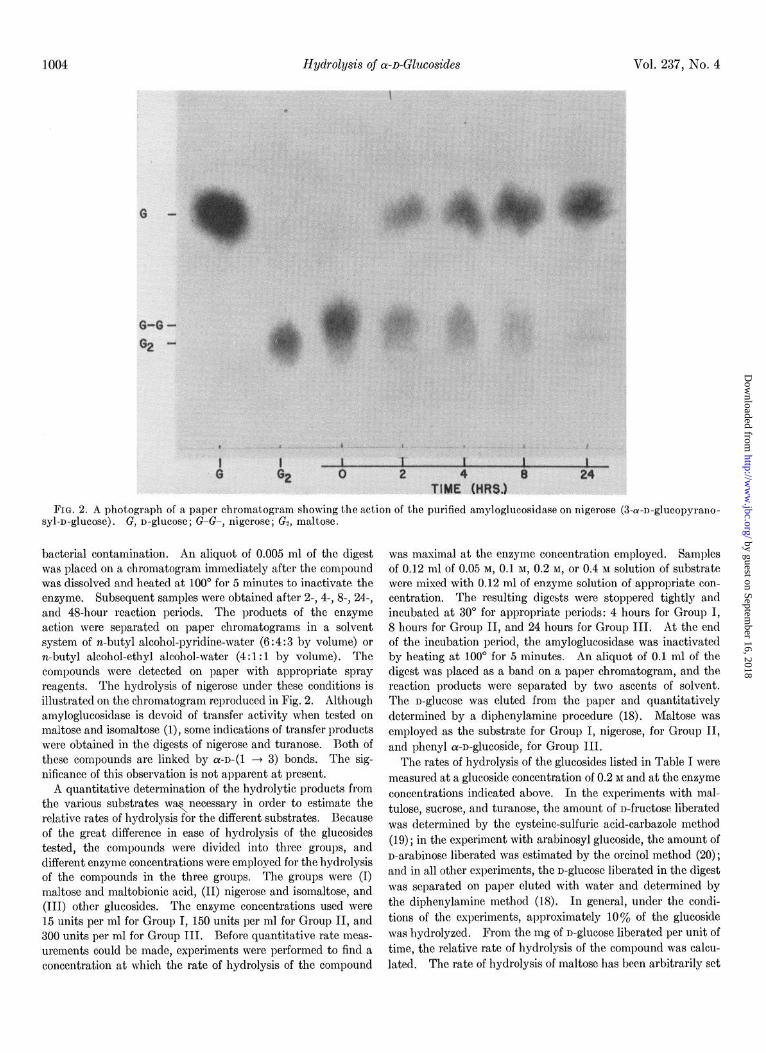
FIG. 2. A photograph of a paper chromatogram showing the action of the purified amyloglucosidase on nigerose (3-a-n-glucopyrano- syl-n-glucose). G, n-glucose; G-G-, nigerose; Gs, maltose.
bacterial contamination. An aliquot of 0.005 ml of the digest was placed on a chromatogram immediately after the compound was dissolved and heated at 100“ for 5 minutes to inactivate the enzyme. Subsequent samples were obtained after 2-, 4-, 8-, 24-, and 4%hour reaction periods. The products of the enzyme action were separated on paper chromatograms in a solvent system of n-butyl alcohol-pyridine-water (6 : 4 : 3 by volume) or n-butyl alcohol-ethyl alcohol-water (4: 1: 1 by volume). The compounds were detected on paper with appropriate spray reagents. The hydrolysis of nigerose under these conditions is illustrated on the chromatogram reproduced in Fig. 2. Although amyloglucosidase is devoid of transfer activity when tested on maltose and isomaltose (l), some indications of transfer products were obtained in the digests of nigerose and turanose. l3oth of these compounds are linked by a-~-(1 3 3) bonds. The sig- nificance of this observation is not apparent at present.
A quantitative determination of the hydrolytic products from the various substrates was,necessary in order to estimate the relative rates of hydrolysis for the different substrates. Because of the great difference in ease of hydrolysis of the glucosides tested, the compounds were divided into three groups, and different enzyme concentrations were employed for the hydrolysis of the compounds in the three groups. The groups were (I) maltose and maltobionic acid, (II) nigerose and isomaltose, and (III) other glucosides. The enzyme concentrations used were 15 units per ml for Group I, 150 units per ml for Group II, and 300 units per ml for Group III. Before quantitative rate meas- urements could be made, experiments were performed to find a concentration at which the rate of hydrolysis of the compound
was maximal at the enzyme concentration employed. Samples of 0.12 ml of 0.05 M, 0.1 M, 0.2 M, or 0.4 M solution of substrate were mixed with 0.12 ml of enzyme solution of appropriate con- centration. The resulting digests were stoppered tightly and incubated at 30” for appropriate periods: 4 hours for Group I, 8 hours for Group II, and 24 hours for Group III. At the end of the incubation period, the amyloglucosidase was inactivated by heating at 100” for 5 minutes. An aliquot of 0.1 ml of the digest was placed as a band on a paper chromatogram, and the reaction products were separated by two ascents of solvent. The n-glucose was eluted from the paper and quantitatively determined by a diphenylamine procedure (18). Maltose was employed as the substrate for Group I, nigerose, for Group II, and phenyl a-n-glucoside, for Group III.
The rates of hydrolysis of the glucosides listed in Table I were measured at a glucoside concentration of 0.2 M and at the enzyme concentrations indicated above. In the experiments with mal- tulose, sucrose, and turanose, the amount of n-fructose liberated was determined by the cysteine-sulfuric acid-carbazole method (19); in the experiment with arabinosyl glucoside, the amount of n-arabinose liberated was estimated by the orcinol method (20) ; and in all other experiments, the n-glucose liberated in the digest was separated on paper eluted with water and determined by the diphenylamine method (18). In general, under the condi- tions of the experiments, approximately 10% of the glucoside was hydrolyzed. From the mg of n-glucose liberated per unit of time, the relative rate of hydrolysis of the compound was calcu- lated. The rate of hydrolysis of maltose has been arbitrarily set
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

April 1962 J. H. Pazur and K. Kleppe 1005
TABLE I
Relative rates of hydrolysis of or-o-glucosides by purified amyloglucosidase
cl+Glucoside
Maltose
Maltobionic acid
Nigerose
Isomaltose
Arabinosyl gluco-
side Maltulose
Sucrose
Phenyl glucoside Glycerol glucoside
Turanose
Trehalose.
Methyl glucoside
structure
4-a-D-Glucopyrano- syl-D-glucose
4-a-D-Glucopyrano- syl-D-gluconic acic
3-a-I)-Glucopyrano- syl-D-glucose
6-a-D-Glucopyrano- syl-D-glucose
3-a-D-Glucopyrano- syl-D-arabinose
4-cx-D-Glucopyrano- syl-D-fructose
a-D-Glucopyrano- syl-8-D-fru&ofu- ranoside
Phenyl cu-D-glucosid 1-c-D-Glucopyrano-
syl-D-glycerol 3-a-D-Glucopyrano-
syl-D-fructose Lu-D-Glucopyrano-
Syl-a-D-&lCOpy-
ranoside Methyl a-D-gluco-
side
n-Glucose liberated
mgjunit/hr
2.3 X 10-l
0.4 x 10-l
1.5 X 10-Z
0.83 X 10-Z
0.19 X 10-Z
0.16 X 10-Z
0.16 X 1O-2
00
34
6.6
3.6
1.66
1.39
1.39
0.10 x 10-2 0.87 0.10 x IO-2 0.87
0.77 x 10-a 0.67
0.17 x 10-a 0.074
0.083 X lo- 3 0.072
lelative rate of lydroly-
sis
at 100, and the relative values for the hydrolysis of the other glucosides have been calculated on this basis (Table I).
DISCUSSION
The cellulose-ion exchange chromatographic method is a simple and rapid procedure for purifying amyloglucosidase from micro- organisms. Fig. 1 shows that the amyloglucosidase obtained from A. n%er sediments across the cell in a single symmetrical boundary, pointing to a high degree of purification of the amylo- glucosidase. When the diffusion coefficients at the various time intervals were calculated and plotted against time, a horizontal line relationship was obtained, indicating homogeneity in the preparation. Gel electrophoretic and paper electrophoretic patterns published earlier (2) also point to a high degree of purification in the amyloglucosidase. The molecular weight of the amyloglucosidase, calculated from the measured sedimenta- tion coefficient and diffusion coefficient and estimated partial specific volume of the enzyme, was found to be 97,000 f 5%. This molecular size for the amyloglucosidase is considerably larger than that for other carbohydrases from microorganisms
(2, 3). The chromatogram reproduced in Fig. 2 shows that the
amyloglucosidase is capable of hydrolyzing the ar-D-(1 + 3) linkage of nigerose. The rate of hydrolysis (see Table I) is approximately twice that for the a-~-(1 ---t 6) linkage of isomal- tose and approximately 7% of the rate for the cr-D-(1 -+ 4)
linkage of maltose. Clearly, the amyloglucosidase is capable of hydrolyzing the three types of bonds in oligosaccharides, and it
is logical to expect that these bonds in branched polysaccharides of the starch type would also be hydrolyzed by the enzyme. Indirect evidence for this suggestion has been published from other laboratories (21,22).
The rate of hydrolysis of the various oc-n-glucosides varied approximately 1400-fold, with maltose being hydrolyzed the fastest and methyl a-n-glucoside, the slowest. Since the amylo- glucosidase acts as an a-n-glucosidase (1, 25, 24), the enzyme- substrate complex most probably involves a combination of enzyme with the glucosyl unit of the substrate. The difference that exists in the glucosyl units of maltose and methyl oc-n-glu- coside is in the conformation of the units. On the basis of in- direct evidence (25, 26), it has been suggested that the glucosyl unit of maltose is in the “boat” conformation whereas that of the methyl a-n-glucoside is in one of the “chair” forms (27). This conformational difference may account for the great differ- ence in the hydrolysis rates of the two compounds. Hydrolysis rates intermediate between those for maltose and methyl a-D-
glucoside may indicate the ease with which the glucosyl units in the compound can transform into the conformation preferred by the enzyme. The conformational aspects of the structure of glycosides may also be in part an explanation for the differences that have been reported in the hydrolysis rates of various galac- tosides by a bacterial fi-n-galactosidase (28).
SUMMARY
Amyloglucosidase of ilspergillus niger purified by chromatog- raphy on diethylaminoethyl cellulose columns appears homoge- neous on ultracentrifugation and on gel electrophoresis. The molecular weight of the enzyme was 97,000 =t 5%. The amylo- glucosidase was capable of hydrolyzing the a-~-(1 + 4), the or-~-(1 --f 6)) and the a-~-(1 --) 3) glucosidic bonds of oligosac- charides. The enzyme also effected the hydrolysis of a variety of other glucosides, but there were large differences in the rates of hydrolysis of these compounds.
REFERENCES
1. PAZUR, J. H., AND ANDO, T., J. Biol. Chem., 234, 1966 (1959). 2. PAZUR, J. H., AND ANDO, T., Arch. Biochem. Biophys., 93, 43
(1961). 3. DIXON, M., AND WEBB, E. C., Enzvrnes, Academic Press, Inc..
New’York, 1958, p. i79. - 4. PAZUR. J. H.. AND ANDO. T.. J. Biol. Chem.. 235. 297 (1960). 5. ABDELIAP&R, M., HA&L&N, J. K., MON&O~&RY, k., AND
SMITH, F., J. Am. Chem. Sot., 74,497O (1952). 6. WOLFROM, M. L., AND THOMPSON, A., J. Am. Chem. SOL, 77,
6403 (1955). 7. PAZUR, J. H., BUDOVICH, T., AND TIPTON, C. L., J. Am. Chem.
Sot., 79, 625 (1957). 8. BARKER, S. A., BOURNE, E. J., AND FLEETWOOD, J. G., J.
Chem. Sot., 4865 (1957). 9. PAZUR, J. H., MARSH, J. M., AND ANDO, T., J. Am. Chem. Sot.,
81, 2170 (1959). 10. BATES, J., Polarimetry, Saccharimetry and the Sugars, National
Bureau of Standards, Washington, D. C., 1942, pp. 467, 494, 517.
11. PAZUR, J. H., AND FRENCH, D., J. Biol. Chem., 196, 265 (1952). 12. FITTING, C., AND PUTMAN, E. W., J. Biol. Chem., 199, 573
(1952). 13. SAWAI, T., AND HEHRE, E., Bacterial. Proc., 161, (1961). 14. ISBELL, H. S., AND SCHAFFER, R., J. Am. Chem. Sot., 76, 1887
(1956). 15. SCHACHMAN, H. K., in S. P. COLOWICK AND N. 0. KAPLAN
(Editors), Methods in Enzymology, Vol. IV, Academic Press, Inc., New York, 1957, pp. 32-103.
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1006 Hydrolysis of a-D-Glucosides Vol. 237, No. 4
16. SMITHIES, 0.. Biochem. J., 71, 585 (1959). 17. RAYMOND, S.; AND WANG,.Y. J., AnaE. Biochem., 1, 391 (1960). 18. PAZUR. J. H.. J. Biol. Chem.. 205.75 (1953). 19. DISC&, Z., ‘in D. GLICK (Edi&),‘Methods of Biochemical
Analysis, Vol. 11, Interscience Publishers, Inc., New York, 1955, p. 331.
20. BROWN, A. H., Arch. Biochem. Biophys., 11, 269 (1946). 21. BURGER, M., AND BERAN, K., Czechoslov. Chem. Communs.,
22, 291 (1957). 22. UEDA, S., Bull. Bgr. Chem. Sot. (Japan), 21, 379 (1957).
23. HELFERICH, B., AND JOHANNIS, J., Z. Physiol. Chem., 320, 75 (1960).
24. WIESMEYER, II., AND COHN, M., Biochim. et Biophys. Acta, 39, 427 (1960).
25. REEVES, R. E., J. Am. Chem. Sot., 76, 4595 (1954). 26. MAYER, F. C., AND LARNER, J., J. Am. Chem. Sot., 81, 188
(1959). 27. REEVES, R. E., Advances in Carbohydrate Chem., 6, 123 (1951). 28. WALLENFELS, K., LEHMANN, J., AND MALHOTRA, 0. P., Bio-
them. Z., 333, 209 (1960).
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

John H. Pazur and Kjell KleppeAspergillus niger-d-Glucosides by Amyloglucosidase from αThe Hydrolysis of
1962, 237:1002-1006.J. Biol. Chem.
http://www.jbc.org/content/237/4/1002.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/237/4/1002.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on September 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from