the culture of cells derived from mouse blastocysts
TRANSCRIPT

Cell, Vol. 5, 343-349, August 1975, Copyright Q 1975 by MIT
The Culture of Cells Derived from Mouse Blastocysts
Michael I. Sherman Roche Institute of Molecular Biology Nutley, New Jersey 07110
On the fourth day of pregnancy, one day prior to implantation, the mouse embryo has reached the blastocyst stage, and contains an average of 64 cells. These cells can be placed into two classes: an outer shell, one cell layer thick, is called the troph- ectoderm, and an enclosed clump of cells is the inner cell mass (ICM). It is generally accepted that cells derived from proliferation of the trophecto- derm layer form only the trophoblast, or fetal part of the placenta (but see Gardner and Papaiannou, 1975) while the ICM contains the stem cells for all parts of the embryo proper, as well as the extraem- bryonic membranes (yolk sac and amnion). The mammalian blastocyst develops in the relatively Iux- urious environment of the uterus, unlike embryos of lower species. It seemed conceivable that the uterine milieu might not only play a protective and nutritive role, but might also provide signals which trigger various events involved in differentiation of the embryo. Early efforts to investigate this question involved removing embryos from the genital tract prior to implantation and reimplanting them in a va- riety of foreign sites as diverse as the anterior chamber of the eye and the testis in a recipient host. Some of the earliest of these ectopic implants, car- ried out in rats by Nicholas (1942), were quite suc- cessful. Following implantation of 2-4-cell embryos under the kidney capsules of recipient rats, the development of a large variety of recognizable em- bryonic tissues was observed at a frequency of about 20%. In fact, Nicholas (1942) maintained that both proliferation and differentiation were actually accelerated under the kidney capsule compared to normal development in utero. Subsequent investi- gators were not always so successful in demon- strating advanced degrees of embryonic differentia- tion when cleavage stage mouse embryos were placed in ectopic sites (for example, see Fawcett, Wislocki, and Waldo, 1947; Kirby, 1962; Billington, 1965). Usually, only extraembryonic derivatives- that is, yolk sac and trophoblast-were observed. However, both Runner (1947) and Stevens (1968) were able to generate embryonic tissues from ectopic implants of 2-cell mouse eggs, and even those laboratories which were not successful with cleavage stage embryos could demonstrate almost normal embryonic development for a few days when blastocysts were implanted (Kirby, 1963; Billington, 1965; Billington, Graham, and McLaren, 1968). These experiments taken together were at least suggestive that differentiation of a number of em- bryonic cell types could be initiated outside the
Review
uterus. With the establishment of conditions appro- priate for blastocyst culture, these and other ques- tions relating to patterns of embryonic development and cell differentiation could be more directly at- tacked.
Short Term Blastocyst Cultures The first description of conditions amenable to postblastocyst development of mouse embryos in vitro was by Mintz in 1964. Although she used a high serum concentration (50%), subsequent stu- dies by Cole and Paul (1965) and Gwatkin (1966) indicated that blastocysts could be maintained in vitro at serum levels more commonly used for cell culture. The results in these three laboratories were similar: while the trophectoderm layer appeared to develop into giant cells resembling the in vivo tro- phoblast, the ICM failed to proliferate or even sur- vive beyond a few days in all but some isolated cases. Cole and Paul (1965) for example, reported limited proliferation of ICM cells in only 5-20% of the blastocysts they cultured.
Since trophoblast cells did appear to develop well in blastocyst cultures, biochemical studies were carried out to determine how close a relationship existed between trophoblast cells developing in vivo and in vitro. A diverse array of biochemical proper- ties were studied: polyploidization (Barlow and Sherman, 1972), alkaline phosphatase (Sherman, 1972a), esterase (Sherman, 1972b), and, most recently, A5,3/&hydroxysteroid dehydrogenase (Chew and Sherman, 1975). In every case, these biochemical markers were detected in trophoblast cells cultured from blastocysts, or even from 2-cell embryos, and usually the time of appearance in vitro closely paralleled that in vivo. More recently, as culture conditions have been used which sup- port development of ICM derived cells as well as trophoblast, yolk sac specific biochemical markers have also been observed after appropriate periods of culture (N-acetyl-P,D-hexosaminidase, Bell and Sherman, 1973; esterases, Sherman, 1975a). In most of the above cases, the relationship between the production of biochemical markers in vivo and in vitro has been only qualitative; enzyme levels in vitro do not usually reach in vivo levels. Neverthe- less, these results clearly indicate that the initiation of differentiation of trophoblast and yolk sac cells can take place in vitro, independently of the mater- nal environment.
The best demonstration to date that development of the embryo proper can occur outside the uterus involves the series of experiments by Hsu and his colleagues (Hsu, 1971, 1972, 1973; Hsu et al., 1974). They have utilized factors such as a collagen substratum and human cord serum and multiple medium changes in their blastocyst cultures. The

Cell 344
result has been the formation, albeit at low fre- quency (5-10% of blastocysts cultured), of embryos which, by morphological criteria, have reached the ninth day of development. By this stage the three germ layers have formed, and structures such as somites, beating heart, and blood islands are all easily detectable.
Since the maternal milieu does not appear to be necessary for the decision-making processes dur- ing early embryonic development, the information must be intraembryonic. A number of studies, re- viewed recently by Graham (1971) and Mintz(1974) have stressed the importance of cell position in the preimplantation embryo in the eventual roles of these cells in development. The details of these ex- periments are outside the scope of this review, but it is now generally agreed that cells on the outside of the embryo have become determined by the blas- tocyst stage to form trophoblast, while only interior cells (ICM) form the embryo proper.
To study whether cell-cell interactions play a role in the early processes of differentiation, experi- ments have been designed to separate the trophec- toderm from the ICM and to monitor the develop- ment of each in the absence of the other. The initial studies were carried out by Gardner (1971, 1972), who separated the two cell populations of the blas- tocyst microsurgically. He demonstrated that the isolated trophectoderm cell fractions reformed blas- tocyst-like structures lacking an ICM, which he called “trophoblastic vesicles.” When placed in uteri or in ectopic sites, these trophoblastic vesicles gave rise to cells morphologically indistinguishable from normal trophoblast cells of the same age; fur- thermore, they showed a functional property of tro- phoblast cells in that they were capable of implant- ing into the transplantation site. Similar results were found by Snow (1973a,b), who showed that cleav- ing embryos treated with appropriate concentra- tions of 3H-thymidine developed into trophoblastic vesicles, presumably due to radiation-induced death of the ICM: these trophoblastic vesicles formed cells which resembled differentiated tro- phoblast cells morphologically and functionally when transplanted into recipient hosts. Further- more, Ansell and Snow (1975) demonstrated that ,H-thymidine-induced trophoblastic vesicles could be maintained in culture, and grew as a monolayer of giant trophoblast cells. Subsequently, it has been shown that appropriate dosages of X irradiation (Goldstein, Spindle, and Pedersen, 1975), actino- mycin D, cycloheximide, cordycepin (Rowinski, Solter, and Koprowski, 1975), bromodeoxyuridine, cytosine arabinoside, and Colcemid (Sherman and Atienza, 1975) will all selectively kill ICM cells, but will not interfere with the differentiation of tropho- blast. Finally, by the use of a procedure originally
described by Tarkowski and Wroblewska (1967), it has been possible to generate trophoblastic vesi- cles which never contained any ICM cells. Not only do these structures form cells morphologically and functionally similar to mature trophoblast cells in vitro, but they also express A5,3,&hydroxysteroid dehydrogenase activity in the temporal manner ex- pected of differentiated trophoblast cells (Sherman, 1975b).
To date, efforts to demonstrate differentiation of ICM cells in the absence of trophoblast have been less successful. Although Gardner (1971) has shown that disaggregated single ICM cells when in- jected into a second blastocyst can differentiate along with the host ICM cells, efforts to clone isolat- ed blastocyst cells in culture have as yet been un- successful (K. E. Bell and M. I. Sherman, unpub- lished observations). These and other observations (Sherman, 1975b) suggest that cell-cell interaction may be necessary for ICM cell survival, but until ICMs can be maintained in a viable state in the ab- sence of trophectoderm, little can be said about the role of cellular interactions in ICM differentiation.
Long Term Blastocyst Cultures When late gestation mouse embryos are disaggre- gated and the resultant cells maintained in culture, the end result is inevitably a reasonably pure popu- lation of fibroblast cells similar to those described by Todaro and Green (1963). Cole and Paul (1965) attempted to culture mammalian blastocysts for long periods of time in order to determine whether these much earlier embryonic cultures would meet the same fate. They succeeded in culturing cells from disaggregated rabbit blastocysts for an ex- tended period of time (more than 200 generations). The cell types that they obtained were both fibro- blastic and epithelioid (Cole and Paul, 1965). The same investigators failed in their efforts at long term culture of mouse blastocysts. The first success at long term culture of mouse blastocysts was report- ed recently (Sherman, 1975~). It is not clear why these efforts were successful while earlier attempts failed, but the strain of mice used might be one fac- tor (see below). Another is the use of a very rich medium, NCTC-109 (Evans et al., 1964), that has not routinely been used in the past in blastocyst culture studies [although it was used by Cole and Paul (1965) in their studies]. Finally, the fetal calf serum was carefully selected, and heat inactivated (56°C for 30 min) prior to use (at a level of 10%). Under these conditions (see Sherman, 1975c, for further details), more than 90% of the blastocysts hatch from their zona pellucidae and attach to the culture dish. Trophoblast cells grow out as a mono- layer, while the ICM initially remains as a solid clump of cells (for example, see Figure 1 a). Although
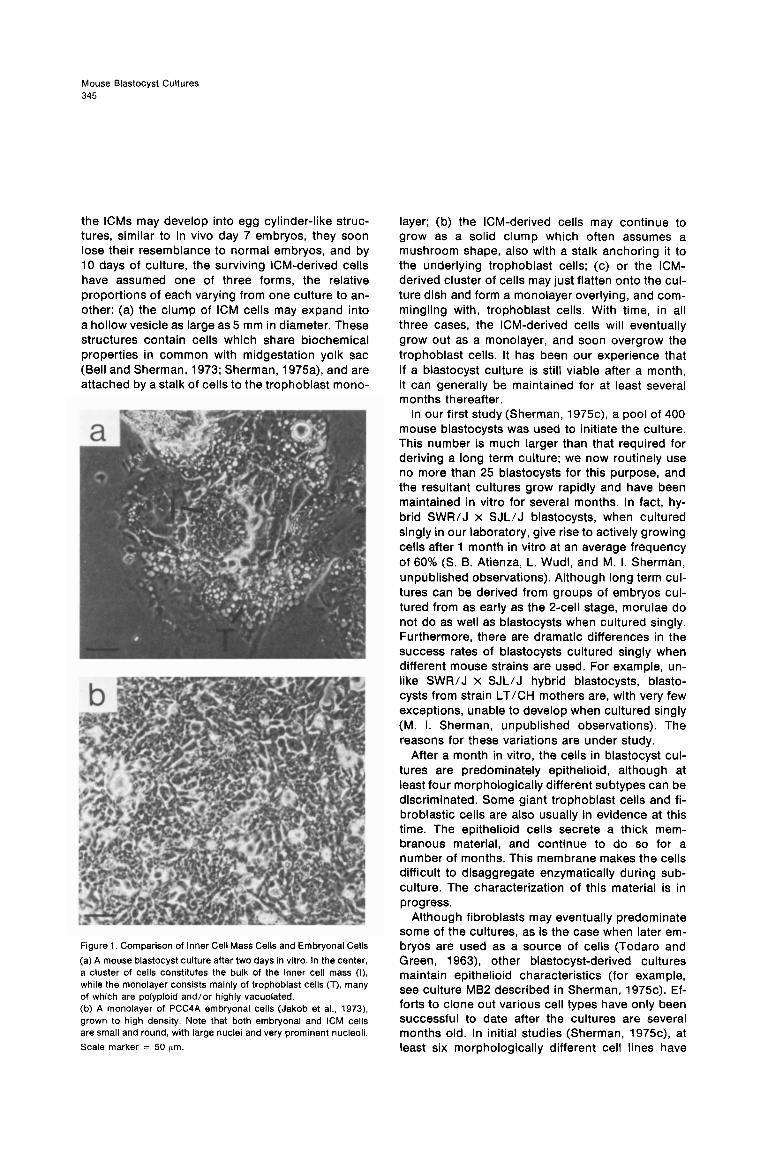
Mouse Blastocyst Cultures 345
the ICMs may develop into egg cylinder-like struc- tures, similar to in vivo day 7 embryos, they soon lose their resemblance to normal embryos, and by 10 days of culture, the surviving KM-derived cells have assumed one of three forms, the relative proportions of each varying from one culture to an- other: (a) the clump of ICM cells may expand into a hollow vesicle as large as 5 mm in diameter. These structures contain cells which share biochemical properties in common with midgestation yolk sac (Bell and Sherman, 1973; Sherman, 1975a), and are attached by a stalk of cells to the trophoblast mono-
Figure 1. Comparison of Inner Cell Mass Cells and Embryonal Cells
(a) A mouse blastocyst culture after two days in vitro. In the center, a cluster of cells constitutes the bulk of the inner cell mass (I). while the monolayer consists mainly of trophoblast cells (T), many of which are polyploid and/or highly vacuolated. (b) A monolayer of PCC4A embryonal cells (Jakob et al., 1973) grown to high density. Note that both embryonal and ICM cells are small and round, with large nuclei and very prominent nucleoli.
Scale marker = 50 pm
layer; (b) the ICM-derived cells may continue to grow as a solid clump which often assumes a mushroom shape, also with a stalk anchoring it to the underlying trophoblast cells; (c) or the ICM- derived cluster of cells may just flatten onto the cul- ture dish and form a monolayer overlying, and com- mingling with, trophoblast cells. With time, in all three cases, the ICM-derived cells will eventually grow out as a monolayer, and soon overgrow the trophoblast cells. It has been our experience that if a blastocyst culture is still viable after a month, it can generally be maintained for at least several months thereafter.
In our first study (Sherman, 1975c), a pool of 400 mouse blastocysts was used to initiate the culture. This number is much larger than that required for deriving a long term culture; we now routinely use no more than 25 blastocysts for this purpose, and the resultant cultures grow rapidly and have been maintained in vitro for several months. In fact, hy- brid SWR/J x SJL/J blastocysts, when cultured singly in our laboratory, give rise to actively growing cells after 1 month in vitro at an average frequency of 60% (S. B. Atienza, L. Wudl, and M. I. Sherman, unpublished observations). Although long term cul- tures can be derived from groups of embryos cul- tured from as early as the 2-cell stage, morulae do not do as well as blastocysts when cultured singly. Furthermore, there are dramatic differences in the success rates of blastocysts cultured singly when different mouse strains are used. For example, un- like SWR/J x SJL/J hybrid blastocysts, blasto- cysts from strain LT/CH mothers are, with very few exceptions, unable to develop when cultured singly (M. I. Sherman, unpublished observations). The reasons for these variations are under study.
After a month in vitro, the cells in blastocyst cul- tures are predominately epithelioid, although at least four morphologically different subtypes can be discriminated. Some giant trophoblast cells and fi- broblastic cells are also usually in evidence at this time. The epithelioid cells secrete a thick mem- branous material, and continue to do so for a number of months. This membrane makes the cells difficult to disaggregate enzymatically during sub- culture. The characterization of this material is in progress.
Although fibroblasts may eventually predominate some of the cultures, as is the case when later em- bryos are used as a source of cells (Todaro and Green, 1963) other blastocyst-derived cultures maintain epithelioid characteristics (for example, see culture MB2 described in Sherman, 1975c). Ef- forts to clone out various cell types have only been successful to date after the cultures are several months old. In initial studies (Sherman, 1975c), at least six morphologically different cell lines have

Cell 346
been isolated by cloning from cultures 6-8 months of age.
Characterization of Blastocyst Derived Cell Llnes Biochemical, immunological, and tumor studies have been carried out on several blastocyst-derived cell lines in an effort to better characterize them. The most widely analyzed cell line is MB4 (Sherman, 197%). MB4 has now been maintained in culture for almost 2 years with little change in morphology. The cells are fibroblastic in nature, although cells with two distinct morphologies have been isolated by cloning. The saturation density of the cell line is the lowest of those examined to date. Like all other blastocyst-derived cell lines tested, it is poorly tumorigenic, although a single tumor has been ob- tained. After passage, the tumor has developed into a rapidly growing, malignant one containing poorly differentiated cells (M. I. Sherman, Ft. A. Miller, and C. B. Richter, manuscript in preparation). Cells in tumor derived cultures resemble the MB4 cell line both morphologically and immunologically (see below).
Along with the other blastocyst cell lines tested, MB4 does not possess any trophoblast-specific bio- chemical markers. This general observation sup- ports the conclusion by morphological criteria that trophoblast cells fail to survive in our cultures for longer than 40 days (Sherman, 197%). MB4 cells do, however, possess some biochemical properties characteristic of midgestation yolk sac but not found in trophoblast or embryo proper, namely the presence of yolk sac-associated esterase (Sher- man, 1972b), an overall preference for butyryl versus acetyl ester substrates, and a very high ratio of N-acetyl-/3,D-hexosaminidase to P-glucuroni- dase activities (Sherman, 197%). On the other hand, neither MB4, nor its derived tumor cells, have the epithelioid morphology characteristic of yolk sac cell lines (Lehman et al., 1974), and MB4 is unreactive with an antiserum generated against yolk sac cells (K. Artzt, personal communication), which cross-reacts with all tested cell lines of endo- dermal origin and a restricted number of tumor lines of mesodermal origin (K. Artzt, H. Jakob, and F. Jacob, manuscript in preparation). However, the yolk sac is a bilayered membrane, and one layer is of mesodermal origin (Snell and Stevens, 1966). Indeed, Jenkinson and Billington (1974) have shown that cells cultured from disaggregated mid- pregnancy yolk sacs have two discrete morpholo- gies. It is therefore possible that MB4 is a mesoder- mally derived yolk sac cell line.
MB2 is a line in which epithelioid cells are pre- dominant. However, cloning studies have revealed that three or more morphologically distinct cell types are present, and at least one of them, with
a stellate morphology, is not epithelioid. The bio- chemical properties so far resemble neither those of trophoblast nor yolk sac cells. One of the cell types produces an orange-yellow pigment which has an absorption spectrum characteristic of a heme protein (Sherman, 1975.c) but which appears not to be hemoglobin (R. A. Miller, personal com- munication). Tumors can be obtained by continu- ous intraperitoneal injections of large numbers of MB2 cells into syngeneic mice (M. I. Sherman, R. A. Miller, and C. B. Richter, manuscript in preparation). The cells grow both in ascitic and solid form. The ascitic form contains structures which are similar to “embryoid bodies” derived from intraperitoneal injections of teratoma cells (see below). The solid tumors consist mainly of epithelioid cells, but can contain pigmented cells and giant cells which bear a striking resemblance to polyploid trophoblast cells. Giant cells have been detected in the parental MB2 cell cultures, but at a very low frequency (Sher- man, 1975~). The parental MB2 cell line cross reacts with the antiserum derived against yolk sac cells (K. Artzt, personal communication), but this does not necessarily indicate a yolk sac derivation for MB2 since, as mentioned above, other endo- derm-derived and some mesoderm-derived cell lines are also positive (Artzt, Jakob, and Jacob, manuscript in preparation).
Two other closely related blastocyst-derived cell lines, MB21 and MB31, have been studied in some detail (Sherman, 1975c). These lines are fibroblastic in nature, although, once again, by cloning studies, two distinct morphologies can be distinguished. Tumors have not yet been found in mice injected with MB21 cells, but one tumor derived from MB31 cells has been tentatively classified as a fibroma (Sherman, Miller, and Richter, manuscript in preparation). The properties of these cells after 8 months in culture were in some respects unlike those of fibroblasts derived from late embryos; for example, MB21, and especially MB31, had very high levels of alkaline phosphatase, and the majority of MB21 and MB31 metaphases contained the diploid number of chromosomes (Sherman, 1975~). On the other hand, after a further 4 months of culture, these distinguishing properties had disappeared: alkaline phosphatase levels were very low, and both cultures contained mainly hypotetraploid cells. Fur- ther studies will have to be carried out to determine whether MB21 and MB31 now possess any signifi- cant differences from other murine fibroblast lines.
A number of other blastocyst-derived long term cultures have now been obtained. In some cases, morphologies have been observed which are not present in the cell lines described above. These new cultures have not as yet been further characterized.

Mouse Blastocyst Cultures 347
Ftelatlonship Between Blastocyst-Derived and Teratoma-Derived Cell Lines In 1958, Stevens noted that males of strain 129 mice had a relatively high incidence of spontaneous tes- ticular teratomas; this observation led to a ready source of these multidifferentiating tumors, which are otherwise rare. Stevens (1958) named the undif- ferentiated tumor stem cells “embryonal cells” be- cause he felt that they strongly resembled very early embryonic cells morphologically, and because they were pluripotential, giving rise to the large variety of differentiated cell types in the tumor. (It should be noted that the term embryonal used in this sense defines a particular cell type, and is different from embryonic, which encompasses a variety of dif- ferent cell species.) Stevens (1968, 1970) demon- strated the close relationship between embryonal cells and early embryonic cells by transplanting preimplantation embryos under the testis capsule. Growths were produced which contained embry- onal cells, among others, and which were indistin- guishable from spontaneously occurring teratomas. Furthermore, Artzt et al. (1973) have observed that a cell surface antigen on embryonal cells also ap- pears on the cells of the morula. These and other relationships between embryonal and early embry- onic cells have been reviewed in detail elsewhere (Damjanov and Solter, 1974; Martin, 1975).
When injected intraperitoneally into syngeneic hosts, embryonal cells give rise to multicellular vesi- cles called “embryoid bodies” (Pierce, Dixon, and Verney, 1960; Stevens, 1980). Aggregates which appear to be akin to embryoid bodies can also be generated in a number of ways from embryonal cells in vitro (Rosenthal, Wishnow, and Sato, 1970; Kahan and Ephrussi, 1970; Nicolas et al., 1975; Martin and Evans, 1975; Sherman, 1975d). When maintained in culture, embryoid bodies attach to the culture dish and give rise to a monolayer of cells, some of which can be recognized as embryonal stem cells. Others assume a variety of morphologies -epithelioid, fibroblastic, neuroblastic, myoblastic, etc. (Rosenthal et al., 1970; Kahan and Ephrussi, 1970; Lehman et al., 1974; Gearhart and Mintz, 1974; Teresky et al., 1974).
Since blastocyst transplants can give rise to tera- tomas (Stevens, 1970), it would not be surprising if the cells developing in blastocyst cultures resem- ble those observed when embryoid bodies are placed in vitro. In fact, shortly after attachment of the blastocyst to the culture dish, the cells of the ICM bear a striking resemblance to embryonal cells (Figure 1). It is, perhaps, unexpected that even though a variety of different cell types can be gen- erated in long term blastocyst cultures, the em- bryonal-like cells do not seem to persist under our culture conditions. Along with the fact that none
of the established cell lines possess cells morpholo- gically similar to embryonal cells, two of the cell lines, MB2 and MB21, have been tested with an anti- serum specific for embryonal cells (Artzt et al., 1973) and fail to cross-react (C. Babinet, personal communication). Conversely, an antiserum derived against MB4 cells fails to react with embryonal cell lines (Miller, Ruddle, and Sherman, 1975). In addi- tion, unlike embryonal cells (Rosenthal et al., 1970; Kahan and Ephrussi, 1970; Jakob et al., 1973; Leh- man et al., 1974; Martin and Evans, 1974), blasto- cyst-derived cell lines do not possess stable diploid, or near-diploid, chromosome numbers (although one of the lines, MB21, maintained a diploid chro- mosome number for at least 7.5 months); instead, like other murine cell lines, they eventually become hypotetraploid (Sherman, 197%). The reason for the failure of embryonal cells to persist in our blas- tocyst cultures is not clear, especially since they are maintained in blastocysts growing under the testis capsule (Stevens, 1970). At least part of the explanation may lie in the observation that em- bryonal cells derived from embryoid bodies (Jakob et al., 1973) grow poorly in NCTC-109 medium (Sherman, 19754). Blastocysts are now being cul- tured in other media to determine whether em- bryonal cell lines can be obtained.
The fact that a variety of different cell types de- velop in our blastocyst cultures raises a rather inter- esting question, namely whether all of these cell types arose at the beginning of the culture period from the embryonal-like ICM cells present, or whether a number of intermediate “oligopotential” cell types were generated from the ICM, each with the ability to give rise in turn to a restricted array of differentiated cells. The isolation of some clones from blastocyst culture MB2 which give rise to cells with a variety of morphologies, none resembling embryonal cells (Sherman, 1975~) appears to sup- port the latter alternative.
When cell cultures derived from embryoid bodies and blastocysts are compared, marked morphologi- cal resemblances are seen. The surface properties of cells derived from blastocysts and embryoid bodies also appear to be related. For example, anti- serum generated from embryoid body cells (Edidin et al., 1971) cross-reacts with all tested blastocyst derived cell lines (S. Rosenberg, M. I. Sherman, and M. Edidin, manuscript in preparation). It has been observed that serum from multiparous or pregnant mice contains antibodies which react with teratoma cells (Edidin, Gooding, and Johnson, 1974; Levine et al., 1975); such antiserum is also reactive with at least some blastocyst derived cell lines (J. D. Hall, A. J. Levine, and M. I. Sherman, unpublished observations). Finally, just as embryonal cells (Artzt et al., 1973) can evoke an antibody response when

Cell 348
injected into syngeneic mice, so too can at least two blastocyst-derived cell lines (Miller et al., 1975). The antiserum obtained is reactive with some blastocyst cell lines, but not others, and with some embryoid body derived cells, but not with embryonal cells.
Applications of the Blastocyst Culture Technique Mouse blastocysts can now be cultured easily and reproducibly. This will, hopefully, facilitate studies of the biochemistry of the early mouse embryo, par- ticularly during, and just after, implantation, times at which it is very difficult to retrieve embryos from the uterus. In fact, Salomon and Sherman (1975) have described an in vitro implantation system wherein blastocysts attach to cultured uterine cells and trophoblast outgrows in an invasive manner into the uterine monolayer. This may provide a model system for the study of cell invasiveness, a topic of interest in embryology and cancer re- search.
If our ability to selectively destroy only a part of the early embryo by antimetabolites or other agents can be extended to later stages and restricted to only certain cell types, a better understanding of the role of cell-cell interaction during various criti- cal periods in development could result. At the same time, it may also be possible to learn about developmental patterns by altering culture condi- tions For example, the blastocysts cultured by Hsu, as described above, can follow the gross morphol- ogy of in vivo development much more faithfully than those in our laboratory; furthermore, Hsu’s cul- ture conditions favor development of the embryo proper, whereas ours seem to provide an ideal envi- ronment for trophoblast and yolk sac development. A study of the differences in the culture conditions used by the two laboratories may pinpoint certain different factors essential for development of the embryonic and extraembryonic parts of the concep- tus. There have now been a whole variety of growth factors described, and, in some cases, highly puri- fied, by a number of investigators. These factors are often quite specific in the types of cells they stimulate. The blastocyst culture system would be an ideal one for testing the potency of these factors during early embryonic development.
Aside from the production of cell lines that might not otherwise be available, long term blastocyst cul- tures present opportunities for approaching a number of problems central to developmental and cell biology. For example, a variety of mutations in the mouse have now been described which are leth- al at some stage in embryonic development. The biochemical nature of these mutations cannot be easily studied in vivo because resorption or abor- tion inevitably results. Embryonic death may occur even if only one or a few cell types (for example,
only one of the three germ layers) are affected, or, as Bennett has proposed in the case of the T mutant locus (for a review, see Bennett, 1975) if the muta- tion affects the organization of cells within the em- bryo rather than lethality at the cellular level. If, in the case of the latter types of mutation, embryos can be removed from the demanding environment of the uterus and placed into culture where factors such as cell organization are not critical and where some cell types can go on to develop even if others fail to do so, they may provide cell lines which would facilitate investigation into the biochemical nature of these mutations. Along a similar line, a number of investigators (Tarkowski, Witkowska, and Nowicka, 1970; Graham, 1970; Komar, 1973) have found ways to produce parthenogenetic mouse em- bryos-that is, embryos which have not been fertil- ized. Some of these embryos are haploid, but none of them will continue in vivo past the first half of gestation, except for the spontaneous partheno- genones in LT/CH mice described by Stevens and Varnum (1974), which form ovarian teratomas. For the same reasons described above, haploid cell lines may be produced in culture, whereas haploid embryos would never survive in utero.
Finally, the observation that blastocyst derived cell lines, like teratoma cell lines, can generate an immune response when injected into syngeneic hosts (Miller et al., 1975) raises the possibility that a whole spectrum of antisera will be obtained from various blastocyst-derived cell lines, which, follow- ing adsorption with appropriate tissues, will be spe- cific for only one or a few embryonic cell types. Such antisera may make it possible to closely follow cell lineages during development.
Acknowledgments
I wish to thank Drs. K. Artzt, A. Levine, R. Pedersen. and D. Solter for allowing me to quote from manuscripts in preparation or in press, and Drs. K. Artzt, A. Levine, D. Salomon. A. Skalka, and L. Wudl for their comments on this manuscript.
Ansell, J. D., and Snow, M. H. L. (1975). In Early Development of Mammals, M. Balls and A. E. Wild, eds. (London: Cambridge University Press), in press.
Artzt, K., Dubois, P., Bennett, D., Condamine, H., Babinet, C., and Jacob, F. (1973). Proc. Nat. Acad. Sci. USA 70, 2988-2992.
Barlow, P. W.. and Sherman, M. I. (1972). J. Embryol. Exp. Morph. 27, 447-465.
Bell, K. E., and Sherman, M. I. (1973). Develop. Biol. 33, 38-47.
Bennett, D. (1975). In Early Development of Mammals, M. Balls and A. E. Wild, eds. (London: Cambridge University Press), in press.
Billington, W. D. (1965). J. Reprod. Fertil. 70, 343-352.
Billington, W. D., Graham, C. F., and McLaren, A. (1968). J. Em- bryol. Exp. Morph. 20, 391-400.

Mouse Blastocyst Cultures 349
Chew, N. J., and Sherman, M. I. (1975). Biol. Reprod. 12, 391-398.
Cole, R. J., and Paul, J. (1965). In Preimplantation Stages of Preg- nancy, G. E. Wolstenholme and M. O’Connor, eds. (Boston: Little, Brown & Co.). p. 82.
Damjanov, I., and Solter, D. (1974). Curr. Topics Pathol. 59, 69-130.
Edidin, M., Gooding, L. Ft., and Johnson, M. H. (1974). Karolinska Symp. Res. Methods Reprod. Endocrin. 7, 336-356.
Edidin. M., Patthey, H. L., McGuire, E. J., and Sheffield, W. D. (1971). In Embryonic and Fetal Antigens in Cancer, N. G. Anderson and J. H. Coggin, Jr., eds. (Oak Ridge: ORNL Press), p. 239.
Evans, V. J., Bryant, J. C., Kerr, H. A., and Schilling, E. L. (1964). Exp. Cell Res. 36, 439-474.
Fawcett, D. W., Wislocki, G. B., and Waldo, C. M. (1947). Amer. J. Anat. 81, 413-443.
Gardner, R. L. (1971). Adv. Biosciences 6, 279-296.
Gardner, R. L. (1972). J. Embryol. Exp. Morph. 28, 279-312.
Gardner, R. L., and Papaiannou. V. (1975). In Early Development of Mammals, M. Balls and A. E. Wild, eds. (London: Cambridge University Press), in press.
Gearhart, J. D., and Mintz, B. (1974). Proc. Nat. Acad. Sci. USA 71, 1734-t 738.
Goldstein, L. S., Spindle, A. I., and Pedersen, R. A. (1975). Radia- tion Res., in press
Graham, C. F. (1970). Nature 226, 165-167.
Graham, C. F. (1971). Symp. Sot. Exp. Biol. 25, 371-378.
Gwatkin, R. B. L. (1966). Ann. N. Y. Acad. Sci. 737, 79-90.
Hsu, Y.-C. (1971). Nature 237, 100-102.
HSU, Y.-C. (1972). Nature 239, 200-202.
Hsu, Y.-C. (1973). Develop. Biol. 33, 403-411.
Hsu, Y.-C., Baskar, J., Stevens, L. C., and Rash, J. E. (1974). J. Embryol. Exp. Morph. 31, 235-245.
Jakob, H., Boon, T., Gaillard, J., Nicolas, J.-F., and Jacob, F. (1973). Ann. Microbial. Inst. Pasteur 1248, 269-262.
Jenkinson. E. J., and Billington, W. D. (1974). J. Reprod. Fertil. 47, 403-412.
Kahan, B. W., and Ephrussi, B. (1970). J. Nat. Cancer Inst. 44, 1015-1036.
Kirby, D. R. S. (1962). J. Embryol. Exp. Morph. 10, 496-506.
Kirby, D. R. S. (1963). J. Anat. 97, 119-130.
Komar. A. (1973). J. Reprod. Fertil. 35, 433-443.
Lehman, J. M., Speers, W. C., Swartzendruber, D. E., and Pierce, G. B. (1974). J. Cell Physiol. 84, 13-26.
Levine, A. J.. Hall, J. D., Rifkin, D. and Teresky, A. K. (1975). In Teratomas and Differentiation, M. I. Sherman and D. Solter, eds. (New York: Academic Press), in press.
Martin, G. R. (1975). Cell 5, 229-243.
Martin, G. R., and Evans, M. J. (1974). Cell 2, 163-172.
Martin, G. R., and Evans, M. J. (1975). Proc. Nat. Acad. Sci. 72, 1441-1445.
Miller, R. A., Ruddle, F. H., and Sherman, M. I. (1975). In Teratomas and Differentiation, M. I. Sherman and D. Solter. eds. (New York: Academic Press), in press.
Mintz, B. (1964). J. Exp. Zool. 157, 273-292.
Mintz, 8. (1974). Ann. Rev. Genetics 8, 411-470.
Nicholas, J. S. (1942). J. Exp. Zool. 90, 41-71.
Nicolas, J. F., Dubois, P.. Jakob, H., Gaillard, J. and Jacob, F. (1975). Ann. Microbial. Inst. Pasteur. 126A, 3-19.
Pierce, G. B., Dixon, F. J., and Verney, E. L. (1960). Lab Invest. 9, 583-602.
Rosenthal, M. D., Wishnow, R. M., and Sato, G. H. (1970). J. Nat, Cancer Inst. 44, 1001-1014.
Rowinski, J., Solter, D., and Koprowski. H. (1975). J. Exp. Zool. 192. 133-142.
Runner, M. N. (1947). Anat. Rec. 98, 1-17.
Salomon, D. S., and Sherman, M. I. (1975). Exp. Cell Res. 90, 261-268.
Sherman, M. I. (1972a). Develop. Biol. 27, 337-350.
Sherman, M. I. (1972b). Exp. Cell Res. 75, 449-459.
Sherman, M. I. (1975a). In Third International Conference on Iso- zymes, 3, C. L. Markert, ed. (New York: Academic Press), p. 83.
Sherman, M. I. (1975b). In Early Development of Mammals, M. Balls and A. E. Wild, eds. (London: Cambridge University Press), p. 145.
Sherman, M. I. (1975~). Differention 3, in press
Sherman, M. I. (19756). In Teratomas and Differentiation, M. I. Sherman and D. Solter, eds. (New York: Academic Press), in press.
Sherman, M. I., and Atienza, S. 8. (1975). J. Embryol. Exp. Morph., in press.
Snell, G. D., and Stevens, L. C. (1966). In Biology of the Laboratory Mouse, E. L. Green, ed. (New York: McGraw-Hill), p. 205.
Snow, M. H. L. (1973a). J. Embryol. Exp. Morph. 29, 601-615
Snow, M. H. L. (1973b). In The Cell Cycle in Development and Differentiation, M. Balls and F. S. Billett, eds. (London: Cambridge University Press), p. 311.
Stevens, L. C. (1956). J. Nat. Cancer Inst. 20, 1257-1276.
Stevens, L. C. (1960). Develop. Biol. 2, 285-297.
Stevens, L. C. (1968). J. Embryol. Exp. Morph. 20, 329-341.
Stevens, L. C. (1970). Develop. Biol. 21, 364-382.
Stevens, L. C., and Varnum. D. S. (1974). Develop. Biol. 37, 369- 380.
Tarkowski, A. K., and Wroblewska, J. (1967). J. Embryol. Exp. Morph. 78, 155-180.
Tarkowski, A. K., Witkowska, A., and Nowicka, J. (1970). Nature 226, 162-l 65.
Teresky, E. K., Marsden, M., Kuff, E. L., and Levine, A. J. (1974). J. Cell. Physiol. 84, 319-332.
Todaro, G. J., and Green, H. (1963). J. Cell Biol. 17, 299-313.