the appearance of acetylcholinesterase in the
TRANSCRIPT

THE APPEARANCE OF ACETYLCHOLINESTERASE
IN THE MYOTOME OF THE EMBRYONIC RABBIT
An Electron Microscope Cytochemical
and Biochemical Study
VIRGINIA M . TENNYSON, MIRO BRZIN, and PAUL SLOTWINER
From the Departments of Pathology, Division of Neuropathology, and Neurology, College ofPhysicians and Surgeons of Columbia University, New York 10032, and the Departments ofNeurology and Neuropathology, Mt. Sinai School of Medicine of Yeshiva University, NewYork 10029. Dr. Brzin's present address is the Institute of Pathophysiology, University ofLjubljana, Ljubljana, Yugoslavia.
ABSTRACT
Acetylcholinesterase (AChE) activity has been studied in the myoblast of skeletal muscle ofthe 9-13 day fetal rabbit . Cytochemical activity is present in the nuclear envelope and theendoplasmic reticulum, including its derivatives the subsurface reticulum and the sarco-plasmic reticulum . End product is also found in the Golgi complex of the more differentiatedmyoblasts . The formation of reticulum-bound acetylcholinesterase in the myoblast appearsto be independent of nerve-muscle contact, since the enzyme is present before the outgrowthof the spinal nerve . The nerve lacks cytochemical end product until the myoblast is welldifferentiated. Possible mechanisms of spontaneous muscle contraction have been discussed .A second type of myotomal cell, which exhibits a poorly localized end product of AChEactivity, has been described . The ready solubility of the enzyme or diffusibility of its endproduct suggests that the enzyme may be a lyoesterase. This cell may be the precursor of themorphologically undifferentiated cell which is closely apposed to the myotubes in later stagesof skeletal muscle development . Biochemical studies show a significant increase in AChEactivity in the dermomyotome by day 12, when many of the myoblasts are well differentiatedand the second type of myotomal cell is prominent . Cytochemical studies have indicatedthat many of the cells in the sample lack reaction product of enzymic activity, whereas othersare very active. Biochemical values, therefore, reflect the amount of enzyme in the dermo-myotome as a whole, but give little information on the enzymic content of individual cells .
INTRODUCTION
An initial formation of cholinesterase (ChE)' nervation (Nachmansohn, 1938, 1939, 1940 ;occurs in developing skeletal muscle before in- Kupfer and Koelle, 1951 ; Zacks, 1954; Mumen-
'Abbreviations: ACh-Che, acetylcholine-cholinesterase ; BuThCh, butyrylthiocholine iodide ; ChE, cholin-AChE, acetylcholinesterase ; AThCh, acetylthio- esterase ; iso-OMPA, tetrais propyl pyrophosphor-choline iodide ; BuChE, butyrylcholinesterase; tetramide .
THE JOURNAL OF CELL BIOLOGY • VOLUME 51, 1971 . pages 703-721
703
on January 2, 2019jcb.rupress.org Downloaded from http://doi.org/10.1083/jcb.51.3.703Published Online: 1 December, 1971 | Supp Info:

thaler and Engel, 1961 ; Karczmar, 1963) . Theenzyme appears in embryonic amphibian muscledeprived of its nerve supply (Shen, 1958), as wellas in nerveless muscle cells grown in tissue culture(Engel, 1961) . Histochemical studies have shownthat myogenic ChE is located diffusely in thesarcoplasm of uninucleated myoblasts, and isfound in greater quantities in the early pluri-nucleated myotubes (Bonichon, 1957 ; Gerebtzoff,1957, 1959 ; Mumenthaler and Engel, 1961 ;Filogamo, 1964 ; Gabella, 1964 ; Veneroni andMurray, 1969). A perinuclear localization ofenzymic activity has been reported (Mumenthalerand Engel, 1961 ; Engel, 1961) as well as depositionof reaction product within the nuclei at later stagesof development (Kupfer and Koelle, 1951) .
Electron microscope cytochemical studies havebeen done on the appearance of ChE during theformation of the motor end plate (Hirano, 1967 ;Lentz, 1969), but detailed information on theintracellular localization of this enzyme duringmyoblast differentiation is lacking . A preliminarystudy from this laboratory (Slotwiner and Tenny-son, 1969) reported that acetylcholinesterase(AChE) activity is localized in the nuclear en-velope and in elements of the endoplasmic retic-ulum of the myoblast . These results are reported infull here . In addition, a second type of myotomalcell containing AChE activity is also described .This cell might be the precursor of the morpho-logically undifferentiated cell which accompaniesthe myotube, and might also be related to themuscle satellite cell (Mauro, 1961) . Biochemicalstudies were done on myotomes at different stagesof development to estimate the amount of enzymepresent. Spinal nerves adjacent to the myotomeswere examined cytochemically as a frame ofreference for a discussion of the induction of theenzyme in skeletal muscle .
MATERIALS AND METHODS
Fixation and Cytochemical Procedure
Brachial and thoracic dermomyotomes of 84Dutch rabbit embryos from day 9 to day 13 of gesta-tion were used . At least one cytochemical procedurewas performed on multiple sections from each em-bryo. In control studies either adjacent sections of thesame embryo or similar areas from littermates wereused depending on the amount of tissue available ata given age . Specimens of peripheral nerve adjacentto some of the myotomes were also examined . Detailsof the fixation and cytochemical procedure have been
7 04
THE JOURNAL OF CELL BIOLOGY • VOLUME 51, 1971
described elsewhere (Tennyson, 1970; Tennyson andBrzin, 1970) . The embryos were fixed by perfusionwith 1 0/0 glutaraldehyde (Sabatini et al., 1963) or amixture of aldehydes in a phosphate buffer. Someembryos were perfused with saline before fixation toremove blood cells, since preliminary studies showedthat these cells contained ChE activity . After fixation,the tissue was washed in several changes of buffer forat least 1 hr. Specimens were cut freehand into 0 .2mm blocks or were embedded in agar and cut at60,u on a Smith-Farquhar microchopper . The latterprocedure permitted a more satisfactory penetrationof substrates and inhibitors into the tissue, and re-sulted in a consistent deposition of reaction product inthe areas illustrated in Fig . 1 . Cytochemical studieswere also done on unfixed tissue, but cellular preserva-tion was so poor that the results will not be em-phasized .INCUBATION MEDIUM : The copper thiocholine
technique for the localization of AChE activity(Koelle and Friedenwald, 1949) was used as modifiedby Brzin et al. (1966) . The specimens were incubatedat pH 6 for 5-50 min at 4 °C in a freshly preparedmedium containing 3.46 X 10 -3 M acetylthiocholineiodide (AThCh) (Sigma Chemical Co ., St. Louis,Mo.), 0 .04 M glycine, 0.008 M copper sulfate, 0.12 Msodium chloride, and 0 .03 M magnesium sulfate in0.02 M sodium hydrogen maleate buffer . Before incu-bation the tissue was preincubated for 30-60 min inthe copper glycinate medium lacking the substrate,but containing the appropriate inhibitor . After incu-bation, the tissue was washed briefly in cold distilledwater and postfixed in a freshly prepared solution ofcold 3% potassium permanganate for 30 min. Thetissue was rapidly dehydrated and embedded inDurcupan (International Chemical & Nuclear Corp.,Burbank, Calif.) .
The Koelle-Friedenwald technique (1949), includ-ing copper sulfide conversion of the end product, wasdone on whole dermomyotomes and nerve, and thehistochemical reaction was examined with the dis-secting microscope .CONTROL STUDIES : To determine the effect of
inhibitors on AChE activity, the following substanceswere added to the preincubation and incubationmedium : BW 284 C 51 (1 ,5-bis-[4-allyl-dimethyl-ammoniumphenyl]pentan - 3 - one - dibromide) 2 X10-4 M (Burroughs Welcome & Co ., Tuckahoe,N. Y.), an inhibitor of AChE, and eserine (physostig-mine sulfate I X 10-4 M (K & K Lab., Plainview,N. Y.), an inhibitor of ChE in general . To eliminatethe possibility of hydrolysis of AThCh by butyryl-cholinesterase (BuChE), tetraisopropyl pyrophos-phortetramide (iso-OMPA) 2 X 10-4 M (PierceChemical Co., Rockford, Ill.), an inhibitor of BuChE,was used in most experiments . To determine whethersites of nonspecific ChE activity were present, butyryl-thiocholine iodide (BuThCh) 3 .1 X 10-3 M (K & K

Labs.), with or without the inhibitor BW 284 C 51,was used . To determine whether nonspecific bindingoccurs between the tissue and the components ofthe medium, specimens were incubated in themedium lacking substrate for 85 min .
Since the use of saturated sodium sulfate has beenrecommended for the elimination of diffusion artifactin ChE studies (Koelle, 1951), this substance was triedin some experiments . Concentrations varying from0.1 to 1 .5 cc of a saturated sodium sulfate solution per5 cc of solution were used either in the incubationmedium, the fixative, or in a saline perfusion beforefixation .
Semites and peripheral nerve adjacent to the areasexamined cytochemically were postfixed with 2%Os04 (Palade, 1952) in a phosphate buffer to studyorganelles not optimally preserved by potassiumpermanganate.
Thick sections for phase microscopy and thin sec-tions for electron microscopy were cut with glass ordiamond knives on a Proter-Blum (Ivan Sorvall, Inc .Norwalk, Conn .) microtome. Thin sections wereplaced on 200-mesh copper grids, blotted dry, andthen stained with uranyl acetate for 5 min (Watson,1958), and with lead citrate for 30 min (Reynolds,1963). Electron micrographs were taken with aSiemens Elmiskop I .
Quantitative Procedure
The embryos were perfused with saline to removeall blood before dissection. Tissue from 57 embryoswas isolated in cold saline by freehand technique withthe aid of a dissecting microscope. Four or five somitesin a row were collected from each side of the embryosduring days 9 and 10. From days 11 to 13, individualdermomyotomes were large enough so that they couldbe dissected free of ectoderm and much of the inter-segmental connective tissue . Individual dermomyo-tomes were then pooled into groups of 3-6 per sample .Immediately after isolation the specimens were frozenand then dried . The dry weight of the specimens(measured on a torsion electromagnetic balancehaving a sensitivity of •0 .04 °g) was between 2 and80 °g, depending on the stage of development andnumber of pooled specimens .
The radiomimetric assay for AChE described byMcCaman et al . (1968) was followed using acetyl-l-I4 C-choline iodide (The Radiochemical Centre,Amersham, England), except that the volumesthroughout the procedure were doubled and theenzymic reaction was performed at 19 µC. Specimensweighing less than 20 jig were introduced directly intothe reaction tube and hydrated in Ringer's solutionfor at least 2 hr . Then the tissue was dispersed with afine glass needle with the aid of a dissecting micro-scope. Samples weighing more than 20 °g were eithercut into smaller pieces, each of which was suspended
as described above, or were homogenized in anappropriate volume of Ringer's solution (5-50 °l)using small glass homogenizers . Samples of 5 °l ofhomogenate were used for the ChE determination .
OBSERVATIONS
Phase microscope sections and a brief descriptionof osmium-fixed material will be given for eachdevelopmental period as a frame of reference forthe cytochemical study . The ultrastructuralchanges taking place during embryonic develop-ment of the myoblast of skeletal muscle in therabbit are very similar to those occurring in otherspecies (Bergman, 1962 ; Hay, 1963, 1968 ; Allenand Pepe, 1965 ; Dessouky and Hibbs, 1965 ;
Heuson-Stiennon, 1965 ; Przybylski and Blumberg,1966 ; Fischman, 1967 ; Auber, 1969), and thusthey will not be illustrated in detail here . Ribo-somes, filaments, and cell junctions, moreover,are not optimally visualized in permanganate-treated specimens, which are used in most of thisstudy .GROSS OBSERVATIONS OF THE HISTO-
C H E M I C A L REACTION : The distribution ofAChE activity in the dermomyotome from days 10to 13, as seen by gross observation, is illustrated inthe diagram in Fig . 1 . The enzyme-active area of
11 day 12 day
13 day
FIGURE 1 Diagram showing the distribution of AChEactivity in two adjacent dermomyotomes and the inter-segmental septum (arrow) viewed from the lateralsurface . Dorsal (top of diagram) . The dermatomal cells(unstippled) lack end product. The areas of the myo-tome containing the heaviest amount of reactionproduct (stippled) can be seen through the dermatomeby transillumination . The precipitate of AChE activityis present in the ventral portion of the intersegmentalseptum at day 11 (dense stipple) and throughout mostof the septum during day 12 . A dense precipitate isfound along the margin of the dermomyotome andscattered within the septum at day 13 . A branch ofthe spinal nerve running through the septum duringthis period exhibits end product.
TENNYSON ET AL . Acetyleholinesterase in Myotome of Embryonic Rabbit
705

the myoblast is more extensive when viewed fromthe surface of the myotome. The dermatome (un-stippled) and dorsal portion of the myotome lackcytochemical end product. At day 10 the ventralportion of the myotome exhibits a light histo-chemical reaction which increases somewhat inintensity and extends throughout the midportionof the myotome during day 11 . In the interseg-mental septum (arrow) a deeper staining precipi-tate is present in the ventral portion (densestipple) . The histochemical end product occupiesa larger area of the myotome by day 12 and theprecipitate is denser. An intense reaction extendsthroughout most of the septum . The enzyme-activearea is concentrated in the ventral regions of themyotome during day 13, but end product extendsdorsally in a narrow central zone . A dense reactionis present at the edge of the myotome borderingthe septum, in areas within the septum, and alongthe nerve .
Myoblasts
NONFILAMENTED MYOBLAST : The ovoid orpyramidal cells of the somite exhibit very littlemorphologic differentiation during day 9 of gesta-tion . They are filled with polyribosomes, but have
only a few rough-surfaced cisternae of the endo-plasmic reticulum and scattered mitochondria.The Golgi complex is small and consists of flat-tened sacs and vesicles . The cells are joined apicallyby primitive desmosome-like junctions (maculaadherens diminuta, Hay, 1968), and they rest on athin basal lamina .
Only some of the myoblasts have AChE activity .They are usually located in the presumptive myo-tomal region (Fig. 2, inset, arrow), but a few othersare scattered throughout the somite . A patchydistribution of AChE activity is present in thenuclear envelope (Fig . 2, Ne) and in the endo-plasmic reticulum (Er) . Rarely, a sclerotomal cellexhibits a cisterna containing end product sur-rounding a moderately dense droplet . Althoughonly a small proportion of the cells in the somiteexhibit AChE activity at this period, a few are seenin every section .EARLY FILAMENTED MYOBLAST : The ros-
tro-caudally oriented cells in the middle and ven-tral portions of the myotome (Fig. 3, between thearrows) differentiate into elongated spindle-shaped myoblasts during day 10. Patches of thick-150 A and thin -60 A filaments in associationwith one another are found in areas containingpolyribosomes . Randomly oriented intermediate-
FIGURE 2 Cytochemical end product is found in the nuclear envelope (Ne) and endoplasmic reticulum(Er) of some cells in the somite at day 9 of gestation. The mitochondria (M) are ovoid and contain fewcristae . AThCh + iso-OMPA, 75 min. Scale marker : 1 A. X 11,200 . Inset: phase micrograph of theneural tube and somite at day 9 of gestation . Most of the enzyme-active cells are found in the presumptivemyotomal region (arrow) . 19 somite embryo . Scale marker : 10 u. X 150 .
FIGURE 3 Phase micrograph of the neural tube and dermomyotomal plate before the formation ofthe spinal nerve . The cranio-caudally oriented myoblasts in the middle and ventral portion of the myotome(between arrows) are elongated cells with cigar-shaped nuclei and prominent nucleoli. Although someneuroblasts are differentiating in the ventral horn (crossed arrow), only a few short ventral root axonscould be found in a sequential section. Bipolar neuroblasts are present in the presumptive dorsal rootganglion, but their processes are short and do not form a peripheral root . Dermatosne (d) . Day 10 ofgestation, 29 somite embryo . Scale marker : 10 µ . X 150 .
FIGURE 4 Cross-section through the perinuclear region of myoblasts from the ventral region of themyotome at day 10. Cytochemical end product is present in the nuclear envelope (Ne) and endoplasmicreticulum (Er) . Enzyme-active cisternae are closely apposed to the cell surface (arrow) . The Golgi com-plex (G) rarely contains end product at this stage . Vacuoles (V) and ovoid mitochondria (M) with fewcristae are present. Scale marker : 1 µ . X 10,500 . Inset: cisternae of the endoplasmic reticulum containingend product (arrow) approximate the surface membrane . AThCh + iso-OMPA, 60 min . Scale marker :1 µ . X 20,800 .
FIGURE 5 Longitudinal section of the distal prolongations of myoblasts from a 10% day embryo .Filaments (F) are forming in the cytoplasm . Cytochemical end product is present in the endoplasmicreticulum (Er), which is located close to the filaments and at the cell surface (arrow) . The constrictedmitochondrion (M) with few cristae may be undergoing division . AThCh + iso-OMPA, 40 min. Scalemarker : 1 u . X 12,400.
706
THE JOURNAL OF CELL BIOLOGY • VOLUME 51, 1971

707

sized filaments -100 A (Ishikawa et al., 1968),microtubules, and glycogen granules are conspic-uous in ground substance of low density . Thecisternae of the endoplasmic reticulum are studdedwith ribosomes over much of the surface, but aconsiderable surface area lacks them . Primitivedesmosomal-like junctions are seen betweenmyoblasts .The few myoblasts having AChE activity at the
beginning of day 10 are located ventrally (Fig . 3,lower arrow), but as their numbers increase, theyare found as far as the middle zone (upper arrow) .Neither the dermatomal cells (d) nor cells in thedorsal edge of the myotome contain end product .Initially, most of the enzymic activity of the myo-blast is found in the perinuclear region, i.e ., inportions of the nuclear envelope (Fig . 4, Ne) and inthe endoplasmic reticulum (Er) . As the myoblastselongate and form more filaments (Fig . 5, F),narrow channels of endoplasmic reticulum (Er)containing end product are found in the distalprolongations of the cell . The enzyme-activereticulum is often closely approximated to thesurface of the myoblast (Fig . 4, arrow and inset ;Fig . 5, arrow) . Study of osmium-fixed tissue fromadjacent somites confirms that the subsurface
708 THE JOURNAL OF CELL BIOLOGY • VOLUME 51, 1971
reticulum at this stage is derived from ribosome-studded cisternae of the endoplasmic reticulum(Fig. 6, arrow) rather than from invaginations ofthe surface membrane . The Golgi complex (Fig .4, G) rarely contains end product during the earlystage. Pinocytotic vesicles are also devoid ofAChE activity.
Enzyme-active myoblasts are distributedthroughout most of the myotome by day 11 (Fig .7, ventral to arrow) and by day 12, with the excep-tion of the dorsal edge . The latter area and thedermatome (d) lack cytochemical end product .Filaments (Fig . 8, F) are still irregularly disposedin the myoblast, but a larger number of thick andthin filaments assume a parallel orientation withrespect to one another and become associated withZ-line material (arrow) . The flattened reticulumin the perinuclear region has attached ribosomesin osmium-fixed tissue . The subsurface reticulumand other channels in the distal prolongations ofthe myoblast have large areas devoid of ribosomes.The distal tip of the myoblast which extends intothe intersegmental septum often has collections ofsmall and large ovoid bodies (Fig . 9, arrow),ranging from 500 to 1500 A in diameter, and ir-regular elongate profiles (crossed arrow) . Both of
FIGURE 6 Subsurface reticulum has a few ribosomes (arrow) attached to its cytoplasmic surface . Thesurface apposed to the cell membrane (crossed arrow) is smooth . Nucleus (N) . Day 10 of gestation, 29somite embryo. Osmium postfixation . Scale marker : 0.1 µ . X 58,000 .
FIGURE 7 Phase micrograph of the neural tube, dorsal root ganglion and dermomyotome just after theoutgrowth of the spinal nerve. The myoblasts (ventral to arrow) show considerable differentiation andenzyme activity. The dorsal edge of the myotome is relatively undifferentiated . Dermatome (d) . Thespinal nerve (Ax) runs about 20 u away from the tip of the ventral edge of the myotome . Day 11 ofgestation . Scale marker : 10 g . X 150 .
FIGURE 8 A greatly elongated myoblast at day 12 of gestation shows a partial alignment of filaments inassociation with Z-line material (arrow) . Some filaments (F) are still randomly oriented . The flattenedcisternae (Er), containing cytochemical end product, probably correspond to ribosome-studded endo-plasmic reticulum . Nucleus (N) . AThCh 50 min. Scale marker : 1 A. X 11,600 .
FIGURE 9 Dense ovoid (arrow) and elongate bodies (crossed arrow) fill the tip of the myoblast whichprojects into the intersegmental septum . There is an increased density of the cell membrane and an under-coating of finely filamentous material (*) . A basal lamina coats the exterior . Day 11 of gestation. Osmiumpostfixation . Scale marker : 0 .1 A . X 40,000 .
FIGURE 10 Invaginations of the cell membrane (crossed arrow) lacking end product often have elongatedconnections to the surface . Enzyme-active subsurface reticulum (arrow) is present in the adjacent fila-mentous myoblast. Day 12 of gestation . AThCh, 50 min. Scale marker : 0.1 g . X 60,000 .
FIGURE 11 A considerable amount of end product (arrow) is present in the nuclear envelope of thedeeply indented myoblast nucleus (N), and in a few cisternae in the cytoplasm. Bundles of filaments (F)are cut in cross section. Scale marker : 1 µ . X 21,600. Inset: cytochemical end product is in narrow channels,some of which are closely apposed to the surface membrane of the peripheral pole of the myoblast . Day12 of gestation. AThCh, 50 min. X 41,600.

709

these structures contain a moderately densematerial, but a clear zone is often seen beneath themembrane of the larger ovoid bodies . Filamentousmaterial (*) undercoats the surface membrane .Invaginations of the cell membrane to form cave-olae (Fig . 10, crossed arrow) are common . Some ofthem exhibit elongated connections with thesurface.
The nucleus (Figs . 8 and 11, N) of these greatlyelongated myoblasts frequently exhibits deep in-foldings. The nuclear envelope often containsmore end product of AChE activity (Fig. 11,arrow) than at earlier periods . Cytochemical endproduct can be seen from time to time in the Golgicomplex of the more differentiated myoblasts .Enzyme-active reticulum is found in the centralcytoplasm (Fig . 8, Er) and beneath the cell mem-brane (Fig. 10, arrow ; Fig . 11, inset) . Caveolaeinvaginating from the cell membrane (Fig . 10,crossed arrow) lack end product of AChE activityas do many other vesicles and saccules of unknownorigin .LATE MYOBLAST - EARLY SARCOMERE
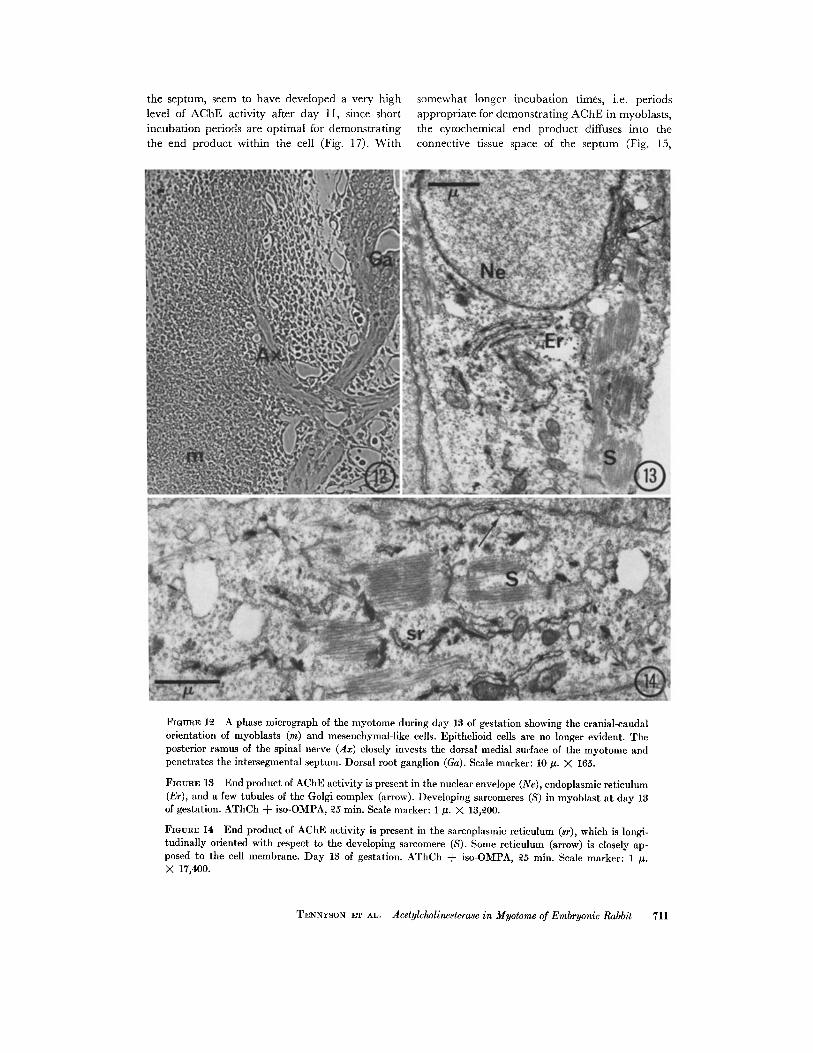
STAGE : The myotome consists of greatly elon-gated spindle-shaped myoblasts (Fig. 12, m) andother mesodermal cells by day 13 . Very little, ifany, dermatomal epithelium or epithelioid ele-ments of the dorsal edge are evident, but there aremany mesodermal cells between the myoblastswhich lack end product of AChE activity . Othermesenchymal-like cells which contain the enzymewill be discussed in the next section. The more ad-vanced myoblasts show early sarcomere formation(Figs. 13 and 14, S) .
The elongated nuclei of the myoblasts are fre-quently indented ; and they have end product ofAChE activity in the nuclear envelope (Fig . 13,Ne) . A few tubules of the Golgi complex (arrow)also exhibit end product at this stage . As in earlierstages, the endoplasmic reticulum (Er) in the peri-nuclear region, as well as the reticulum beneaththe surface membrane of more distal parts of thecell (Fig. 14, arrow), have cytochemical activity .It is likely that portions of the enzyme-activereticulum which are aligned parallel to the sarco-meres have begun to differentiate into sarco-plasmic reticulum (sr), since corresponding chan-nels in osmium-fixed tissue are smooth surfaced .As in the preceding stage, caveolae lacking endproduct are common, but there is little or no evi-dence of extensive T-system formation as yet .
71 0 THE JOURNAL OF CELL BIOLOGY . VOLUME 51, 1971
A Second Type of Myotomal Cell
A cell type distinct from the myoblast is recog-nizable in the myotome as early as day 10 of gesta-tion, but is more common by day 11 . It is firstfound bordering the intersegmental septum closeto the ventral margin of the myotome (Fig . 15,arrows). In the more differentiated myotome, it ispresent along the length of the septum and alsobetween the myoblasts in deeper portions of themyotome, but it appears to be absent from thedorsal edge (D) of the myotome proper . Morpho-logically, this cell resembles the relatively un-differentiated mesenchymal cell described byothers (Bergman, 1962; Hay, 1963 ; Allen andPepe, 1965; Dessouky and Hibbs, 1965 ; Przybylskiand Blumberg, 1966) . It has a large ovoid nucleus(Fig. 16, N), prominent nucleolus, and scantperikaryal cytoplasm. There are few organelles,but sometimes a centriole is present in a Golgiregion . The cytoplasm lacks filaments, but is filledwith polyribosomes in osmium-fixed tissue . Theendoplasmic reticulum is sparse . Junctions arenot obvious in permanganate-fixed tissue in whichthis cell can most readily be identified . The cellvaries from early undifferentiated mesoderm andmesenchymal cells elsewhere in that it exhibitsAChE activity . The distribution of the enzyme,however, is different from the membrane-boundpattern seen in the nonfilamented and filamentedmyoblasts. The end product in the second type ofmyotomal cell is scattered throughout the nucleusand cytoplasm in a random manner (arrows),suggesting that the enzyme or the end product maybe loosely bound in these cells .
Near the end of day 10 and the early part of day11, small groups of free cells, which resemble thesecond type of myotomal cell, are found in theventral portion of the intersegmental septum (Fig .15,*) close to the walls of the blood vessel (Ba) .During the latter part of this period, similar cellsare found deeper within the septum (arrowhead),and by day 12 they are spread throughout theseptum. These cells have large nuclei (Fig . 17),scant cytoplasm, and few organelles . Cytochemicalend product (arrows) is scattered throughout thenucleus and cytoplasm . It is common to findrandom end product in neighboring cells in mitosis .As will be pointed out in the discussion, it is likelythat the mesenchymal-like cell containing AChEactivity originates from the second type of myo-tomal cell .
The second type of myotomal cell and its prob-able derivative, the enzyme-containing cell free in
the septum, seem to have developed a very high somewhat longer incubation times, i.e . periodslevel of AChE activity after day 11, since short appropriate for demonstrating AChE in myoblasts,incubation periods are optimal for demonstrating the cytochemical end product diffuses into thethe end product within the cell (Fig. 17) . With connective tissue space of the septum (Fig . 15,
FIGURE 12 A phase micrograph of the myotome during day 13 of gestation showing the cranial-caudalorientation of myoblasts (m) and mesenchymal-like cells . Epithelioid cells are no longer evident . Theposterior ramus of the spinal nerve (Ax) closely invests the dorsal medial surface of the myotome andpenetrates the intersegmental septum . Dorsal root ganglion (Ga) . Scale marker : 10 µ. X 165 .
FIGURE 13 End product of AChE activity is present in the nuclear envelope (Ne), endoplasmic reticulum(Er), and a few tubules of the Golgi complex (arrow) . Developing sarcomeres (S) in myoblast at day 13of gestation. AThCh + iso-OMPA, 25 min. Scale marker : 1 µ. X 13,200 .
FIGURE 14 End product of AChE activity is present in the sarcoplasmic reticulum (sr), which is longi-tudinally oriented with respect to the developing sarcomere (S) . Some reticulum (arrow) is closely ap-posed to the cell membrane. Day 13 of gestation . AThCh + iso-OMPA, 25 min. Scale marker : 1 µ .X 17,400.
TENNYSON ET AL . Acetylcholinesterase in Myotome of Embryonic Rabbit
711

crossed arrow) and into the distal processes of themyoblasts bordering it. The second type of myo-tomal cell contains somewhat more endoplasmicreticulum by day 13 (Fig . 18, Er), but the cyto-chemical end product (arrows) remains dispersed .The cells at this stage are most commonly foundadjacent to the distal poles of the myoblasts border-ing the intersegmental septum and in groupswithin the septum, as well as within the myotomeproper adjacent to myoblasts . A diffuse distribu-tion of end product of AChE activity is also seenin other embryonic cells, such as developing redblood cells (Fig . 19) and Schwann cells (Fig. 20,Sc), differentiating along the spinal nerve after day12 of gestation. AChE has been found biochem-ically in human red blood cells (Richter and Croft,1942 ; Bellhorn et al ., 1970) .
Spinal Nerve
None of the cells of the neural tube at day 9(Fig. 2, inset) have differentiated into neuroblasts .At day 10, i .e . the 29 somite stage, neuroblasts arepresent in the ventrolateral neural tube (Fig . 3,crossed arrow), but only occasionally are thereneurites which extend into the mesenchyme . Theseaxons are about 20 .s in length at most, and do notjoin the short cytoplasmic prolongations of theloosely arranged spindle-shaped neuroblasts, whichare forming the presumptive dorsal root ganglion .After the middle of day 10, i.e . the 34 somite stage,the motor axonal processes are longer and they
7 1 2
ramify within the mesenchyme just medial to themyotome. Some axons have been traced to within10-20 e of the central area of the myotome, i .e . thenuclear zone of the longitudinally oriented myo-blasts, but then the axons turn and run ventrally,forming a distinct motor root by day 11 (Fig . 7,Ax) . No axonal-myoblast contacts have been ob-served. The peripheral axons of the dorsal rootganglia are rudimentary until day 11 ;4. They jointhe motor root, forming a distinct spinal nerve byday I l ) . Some axons wander in the mesenchymeclose to the myotome, but do not appear to enter it .A few axons of the posterior ramus enter the medialentrance of the intersegmental septum at clay12.4. A well-formed nerve (Fig . 12, Ax) penetratesthe medial and dorsal edges of the myotome, aswell as the septum from day 1232 to 13 . Presump-tive Schwann cells separate the nerve from thebulk of the myotomal cells, but it is not knownwhether scattered exploratory nerve-muscle con-tacts occur or not. Most of the cells in the dorsaledge of the myotome during this period resemblemesenchymal cells and lack cytochemical endproduct .A detailed account of the development of AChE
in the spinal nerve will be the subject of anotherpaper, but the following data will be given sincecontamination of the myotomal sample by AChE-containing nerve may affect the biochemicalresults . There is very little AChE activity in theearly nerve fibers (Fig. 21, Ax) or Schwann cells
FIGURE 15 Phase micrograph of a longitudinal section of portions of two myotomes and the inter-segmental septum at day 11% of gestation. Elongated filament-containing distal processes of the myo-blasts (m) are evident . A second type of myotomal cell (arrows) border the ventral portion of the septum.These cells, as well as free cells (*) which appear first near the entrance of the capillary (Bv) and laterwithin the septum (arrowhead), are the source of the cytochemical end product (crossed arrow) in theconnective tissue area. No end product is present in the dorsal edge of the myotome (D), or in the dorsalportion of the septum or around the distal extension of the capillary (Br') . AThCh + iso-OMPA, 20 min.Scale marker : 10 A. X 400.
FIGURE 16 A second type of myotomal cell having a large nucleus (N), scant cytoplasm, and few or-ganelles borders the extracellular space (Ex) close to the ventral portion of the intersegmental septum .Cytochemical end product (arrows) is randomly distributed throughout the nucleus and cytoplasm ofthese cells. In the filamented (F) myoblast, end product is most frequently associated with the reticulum(Er) . Day 10% of gestation. AThCh + iso-OMPA, 40 min. Scale marker : 1 µ . X 8700 .
FIGURE 17 A group of cells within the intersegmental septum that closely resemble the AChE-contain-ing second type of myotomal cell. Cytochemical end product (arrows) is scattered throughout the nucleusand cytoplasm . Day 11% of gestation . AThCh + iso-OMPA, 10 min . Scale marker : 1 is . X 7200 .
FIGURE 18 Mesenchymal-like cells containing random end product (arrows) of AChE are present inthe intersegmental septum at day 13 . Elongated channels of endoplasmic reticulum (Er) are evident .AThCh + iso-OMPA, 25 min. Scale marker : 1 µ . X 14,400 .
THE JOURNAL OF CELL BIOLOGY . VOLUME 51 .1971

713

which course past the myotome during day 11(Fig . 7, Ax) . Only occasionally has end productbeen seen, and that was found in the agranularreticulum of growth cones . When enzymic activitydevelops in the spinal nerve, it seems to appear ina distal to proximal direction, as was found in thedorsal root (Tennyson and Brzin, 1970) . TheSchwann cells exhibit a significant amount of cyto-chemical end product by day 12 ;2 (Fig . 20, Sc),
7 1 4
but the reaction is diffuse . AChE activity can beseen at the surfaces of some axons (Fig . 22, arrows)at day 13, but the Schwann cells are more con-sistently active . It is likely that the axons andSchwann cells containing AChE activity in thespinal nerve are derived from the sensory nerve .The peripheral nerve of the dorsal root ganglionexhibits AChE activity at day 13, but the anteriorroot of the spinal cord does not .
FIGURE 19 Developing red blood cells also show a random distribution of end product of AChE activity(arrows) . Day 11 of gestation. AThCh + iso-OMPA, 45 min. Scale marker : 1 ,u. X 6300.
FIGURE 20 Random end product (arrows) is present in the nucleus and cytoplasm of many of theSchwann cells (Sc), in sensory nerve fibers joining the spinal nerve at day 12% . The axons (Ax) lessfrequently contain cytochemical activity. AThCh + iso-OMPA, 15 min. Scale marker : 1 ,u. X 18,600 .
FIGURE 21 Axons (Ax) of the spinal nerve passing within 20 µ of the somite at day 11 lack cytochemicalend product. AThCh + iso-OMPA, 20 min. Scale marker : 1 ~u . X 15,900 .
FIGURE 22 Some axons of the posterior ramus of the spinal nerve, which are closely associated with themyotome at day 13, have some end product of AChE activity at their surfaces (arrows) . Most of the axonslack end product. AThCh + iso-OMPA, 30 min. Scale marker : 1 µ. X 13,500 .
THE JOURNAL OF CELL BIOLOGY • VOLUME 51, 1971

Control Studies
myotomal cell, and spinal nerve reacted similarlyat all ages studied . The myoblast of the 13 day
The specificity of the enzyme was examined fetus was selected to illustrate the control studiescytochemically in specimens from day 9 through since it consistently contains end product afterday 13 of gestation. Myoblasts, the second type of incubation in AThCh and it is readily recognized
FIGURES 23-25 show the effect of inhibitors on AChE activity in the reticulum of elongated processesof the late myoblast at day 13 of gestation . Scale marker : 1 .s. X 17,400.
FIGURE 23 Cytochemical activity is inhibited by treatment with BW 284 C 51 . AThCh + BW 284 C 51,25 min.
FIGURE 24 Cytochemical activity is inhibited by treatment with eserine . AThCh + eserine, 25 min .
FIGURE 25 Cytochemical activity is inhibited after incubation in BuThCh + BW 284 C 51 for 25 min .
TENNYSON ET AL . Acetyleholinesterase in Myotome of Embryonic Rabbit
715

7 16
TABLE IAChE Activity of the Dermomyotome Expressed as µm ACh X I0-4/hr per Dry Weight of Tissue (+SD)
* n = number of experiments .
by its filaments . The second type of myotomal cell,on the other hand, can only be distinguished froma mesenchymal cell by the presence of end productof AChE activity. The statement on the effect ofinhibitors on these cells, therefore, is based on theabsence of end product in mesenchymal-like cellsin the specific areas of the myotome and inter-segmental septum where these cells are normallyfound .
There is little or no effect on enzymic activitywhen specimens are treated with iso-OMPA (2 X10 -4), an inhibitor of nonspecific ChE . The dis-tribution and the relative number of deposits ofcytochemical end product (Fig. 14) is the same asin experiments with AThCh without inhibitors .There was inhibition of cytochemical activitywhen the tissue was incubated in AThCh in thepresence of either BW 284 C 51 (2 X 10-4 M), aninhibitor of AChE (Fig . 23), or eserine (1 X 10 -4M), an inhibitor of both specific and nonspecificChE (Fig. 24) . Preincubation for 1 hr of 60 k sec-tions in the medium without substrate, but con-taining the inhibitors, is necessary to effect com-plete inhibition of the enzyme . Cytochemical endproduct was occasionally observed when speci-mens were incubated with BuThCh (3 .1 X 10-3M) as substrate, but these deposits are abolished bythe use of BW 284 C 51 (Fig . 25) . There was noprecipitate if the tissue was incubated in the coppermaleate buffer medium without the substrate .
Attempts made to alter the diffuse distributionof end product in the second type of myotomal cellhad no effect on its localization. The use of highconcentrations of sodium sulfate in (a) a saline orbuffer perfusate before fixation, (b) in the perfusingfixative itself, or (c) in the incubation mediumacused tissue shrinkage but did not confine the endproduct to membranes . The random distributionof end product is present in both unfixed andfixed tissue . It is not present if AThCh is omittedfrom the incubation medium, and thus the pre-cipitate cannot be due to a nonenzymatic bindingof copper (Langley and Landon, 1969) .
Gestation day
0.31 f 0.07
1 .13 f 0.38
2 .69 f 0 .54
10 .29 f 0 .80
14 .52 f 1 .81(n - 7)*
(n = 7)
(n = 8)
(n = 5)
(n = 5)
THE JOURNAL OF CELL BIOLOGY . VOLUME 51, 1971
12
13
Quantitative Studies
Chemical measurements for AChE activity insomites taken from 16 to 19 somite embryos at day9 of gestation are low (Table I) . Enzymic activityhas increased almost fourfold by day 10 and almostninefold by day 11 . A sharp rise in AChE activityoccurs during day 12, and it reaches a peak at day13, when enzymic activity is almost 50-fold that ofday 9 .
DISCUSSION
AChE activity has been demonstrated in thenuclear envelope and in elements of the endo-plasmic reticulum of the myoblast of the rabbitembryo by use of the substrate AThCh in con-junction with selective inhibitors . Although it isnot possible to distinguish with certainty betweenrough and smooth reticulum in permanganate-treated specimens, study of osmium-fixed tissueshows that most of the reticulum during earlymyoblast development is studded with ribosomes .In later stages some of the reticulum is devoid ofribosomes, particularly that closely associated withdeveloping sarcomeres . Although the Golgi com-plex begins to show a small amount of cytochem-ical end product in later stages, the preponderanceof AChE activity first in the endoplasmic reticulumthen in areas known to contain smooth reticulumsupports the evidence showing that the sarco-plasmic reticulum is derived from the endoplasmicreticulum (Ezerman and Ishikawa, 1967) . Itshould be pointed out that cytochemical end prod-uct is abundant in the honeycomb structuresclearly identifiable as sarcoplasmic reticulum inmyotubes (unpublished observations) . Study ofosmium-fixed tissue also shows that the enzyme-containing subsurface reticulum is derived fromthe rough surfaced reticulum rather than from theT system .Ezerman and Ishikawa (1967) and Ishikawa
(1968) have presented evidence showing that T-tubule formation begins in vitro in the myotube

stage. There is an inpocketing of the sarcolemmato form a caveola, which resembles a micropino-cytotic vesicle having a tunnel-like connection tothe surface . Then secondary caveolae extend fromthe inpocketing and the structure becomesbranched. A well-formed, branching T system isnot apparent in the present study of myoblastsbefore fusion, although there are numerous vesiclesof unknown origin in the cytoplasm . Caveolae arepresent at all stages, so it is likely that many ofthem are pinocytotic vesicles. Caveolae are en-countered which exhibit an elongated neck sugges-tive of T-tubule formation. End product has notbeen found in any of the caveolae . A study of laterstages of development is necessary, therefore, todetermine whether the T system has AChE activity .
Second Type of Myotomal Cell
A second type of myotomal cell which exhibits apoorly localized cytochemical reaction for AChEhas been described in the intersegmental septumand along the medial border of the myotome withthe septum. It is derived from the mesodermalsomite, as is the nonfilamented myoblast, but itdifferentiates slightly later . The second type ofmyotomal cell differs from the nonfilamentedmyoblast in that it lacks primitive desmosomal-likejunctions, exhibits a random end product of AChEactivity, and initially appears only in a localizedpart of the myotome, i .e ., along the ventral borderof the septum. Similar cells containing randomend product are found along the surfaces of myo-tubes at later stages (unpublished observations)and also in dystrophic muscle (Miranda, 1971) .Fibroblasts lack the reaction product. The secondtype of myotomal cell, therefore, may be theprecursor of the cell associated with myotubes andreferred to as the satellite cell (Muir et al ., 1965 ;Ishikawa, 1966 ; Church, 1969) or undifferent-iated cell (Kelly and Zacks, 1969), and mayultimately be related to the satellite cell of maturemuscle (Mauro, 1961) .
It is not possible to state with the present tech-nique the precise intracellular localization in vivoof AChE in the second type of myotomal cell .Attempts made to limit diffusion artifact were un-successful . Koelle and coworkers (1970) have dis-cussed the possibility that differences with respectto solubility or diffusibility of this enzyme at motorend plates as opposed to autonomic ganglia, mightbe explained by the recent findings of isoenzymesof AChE (Bernsohn et al ., 1962; Eränko et al .,1964 ; Ecobichon and Israel, 1967) .
Histochemical studies (Eränko et al ., 1964 ;Koelle et al ., 1970) have dealt with the question oftissue-bound esterases vs. diffusible esterases.Eränko et al . (1964) have distinguished desmo-esterases, which are tightly bound and are demon-strable by histochemistry, and lyoesterases, whichare readily solubilized in fresh tissue sections, butare immobilized by formalin fixation . The randomcytochemical end product in the second type ofmyotomal cell, the developing red blood cell, andthe developing Schwann cell in this study and
in a study of the dorsal root (Tennyson andBrzin, 1970) suggests that the enzyme in thesecells may be closely related to a lyoesterase . Al-
though a certain amount of the enzyme is retainedby fixation, the solubility of the enzyme or thediffusibility of its end product is so great that itsultrastructural localization may be meaningless .The electron microscope does show, however, thatthis AChE-containing cell from the myotome isdistinct from the myoblast in which the enzymeappears to be more tightly bound to membranes .
It is probable that the enzyme-active mesenchy-mal cells in the septum are derived from the secondtype of myotomal cell. Although they aggregatearound the endothelial cells, it is unlikely that theyoriginate from the wall of the blood vessel . Peri-capillary cells around the same blood vessel in thedorsal portion of the septum lack the end productduring the early stage . Later, after the second typeof myotomal cell differentiates along the dorsalmargins of the septum, the enzyme-active mesen-chymal cell can be found in the adjacent inter-segmental area .
Chemical Measurements
The cytochemical results as diagrammed inFig. 1 have shown that AChE is not distributeduniformly throughout the tissue studied . Thebiochemical measurements, therefore, representthe enzymic activity in the dermomyotome as awhole, but give little information concerning theamount of enzyme within the individual cells .At day 10, there is a considerable amount ofcytochemical activity in a few myoblasts in themiddle and ventral myotome, but the rest of themyotome and the dermatome are inactive . The
values for day 10 are rather low because the sampleconsists of a sparse population of AChE-contain-ing myoblasts and many inactive dermatomal cells .The marked increase in biochemical values duringdays 11, 12, and 13 is in part due to the largernumber of differentiated myoblasts, but probably
TENNYSON ET AL. Aeetyleholinesterase in Myotome of Embryonic Rabbit
717

even more to the second type of myotomal cell,which shows intense histochemical activity . Thespinal nerve may also have contributed to themeasurements of AChE activity in the myotomeat day 13 . Although care was taken to removeall of the spinal nerve visible to the eye, it ispossible that microscope nerve fibers and sheathcells, which now contain AChE activity, mayhave been adherent to the sample measuredbiochemically.
Nerve-Muscle Interaction
A direct contact between nerve and musclehas been postulated as a necessary prerequisitefor the formation of AChE at the neuromuscularjunction (Mumenthaler and Engel, 1961 ; Ze-lenra, 1962 ; Lentz, 1969), but differentiation ofmuscle is not directly dependent on innervation(Zelena, 1962) . The myoblast, moreover, canform intracellular AChE at very early stages ofdevelopment in the absence of nerves as indicatedby light microscope studies (Zacks, 1954 ;Bonichon, 1957 ; Shen, 1958 ; Engel, 1961 ; Mu-menthaler and Engel, 1961), and as confirmedhere both cytochemically and biochemically .Even after nerve outgrowth, the formation ofACNE within elements of the endoplasmic reticu-lum of the myoblast appears to be independent ofdirect nerve-muscle contact . There is a consider-able amount of enzymic activity in the myotomeby day 11, but the spinal nerve coursing within10-20 µ of it does not appear to enter the myotomeproper. The nerve itself does not exhibit AChEat this time, nor were motor end plates en-countered. Using silver impregnation, Romanes(1941) noted that a short posterior primary ramusfirst penetrated the dorsal musculature of therabbit during day 12, but no definite neuromuscu-lar endings were seen until day 19 . He noted,however, that reflex contraction on stimulationof the snout could be elicited by day 16l. andsuggested that the earliest endings might not beimpregnated by silver .
Visintini and Levi-Montalcini (1939) havedemonstrated fine argentophilic nerve fibersdirected toward the medial surface of the myotomeof the chick embryo of 68-96 hr . This period wouldcorrespond roughly to days 103 -12 in the rabbitembryo. Since motor end plates are not presentat the end of day 4 in the chick, Visintini andLevi-Montalcini (1939) postulated the existenceof provisional contacts between terminal boutons
7 1 8
TuE JOURNAL OF CELL BIOLOGY • VOLUME 51, 1971
and myoblasts to explain spontaneous movementsin myoblasts. They also showed inhibition ofthese movements by curare . Hamburger (1963)has confirmed the observations concerning curari-zation and assumes that the earliest motility inthe embryo is neurogenic . Engel (1961) has pointedout that the presence of spontaneous movementsin young embryos does not necessarily demon-strate functional neuromuscular transmission ofstimuli, since Li et al . (1959) have found strongand rhythmic spontaneous contractions of chickembryo skeletal muscle in tissue culture in theabsence of innervation . Hagopian et al . (1970)have shown that AChE is present in the reticulumof spontaneously contracting cardiac musclebefore the formation of T tubules or the ingrowthof nerve fibers. It was suggested that an acetyl-choline (ACh)-ChE system of myogenic originmay be involved in ionic events underlying spon-taneous contractions of cardiac muscle . Skeletalmyoblasts exhibit similar conditions in earlystages, i .e ., AChE is present in the reticulumbefore the formation of T tubules and beforeneuromuscular contacts are made. An ACh-ChE system of myogenic origin may be postulated
contractions in skeletal muscle,can demonstrate ACh within
for spontaneousif future studiesthese cells .
On the other hand, if ACh is present withinearly axons, a neurogenic origin for spontaneouscontraction may be possible even in the absenceof direct nerve-muscle contact. ACh may bereleased from early neurites and diffuse to themyotome . This possibility has been suggested fora later stage in myotube development (Kupferand Koelle, 1951) . Burt (1968) has shown thatcholine acetyltransferase, the enzyme necessaryfor ACh synthesis, is present in the chick spinalcord after the middle of day 3 . The onset ofspontaneous activity in the chick occurs at thistime (Hamburger et al., 1965) . Although thereis AChE activity in the cell bodies of the motorhorn of the chick, this enzyme seems to be absentfrom the ventral root fibers during their earlyoutgrowth (Gerebtzoff, 1959) . The latter ob-servation was confirmed in the present study ofthe rabbit embryo. Since there is little or no AChEin the early neurites to inactivate ACh and theaxons are incompletely ensheathed, there wouldbe no barrier to the diffusion of ACh 10-20 kthrough the mesenchyme to the neighboringmyotome. The AChE-containing subsurface re-

ticulum in the myoblast might serve as a siteof inactivation of ACh . This reticulum closelyresembles subsurface cisternae in the neu-ron (Rosenbluth, 1962) . Rosenbluth (1962) hassuggested that the close apposition of thesecisternae to the cell membrane may change theproperties of the neuronal plasmalemma, possiblyaltering its permeability and affecting electro-physiological behavior in these regions . A similarmechanism may operate in the myoblast beforethe formation of an extensive T system . AChEin the subsurface reticulum may play a role inspontaneous contraction of the myoblast as partof an ACh-receptor protein-AChE system. Nach-mansohn (1970) has suggested that ACh mayinduce a conformational change in the receptorprotein thereby releasing previously boundcalcium ions, which are involved in the excit-ability of muscle . The AChE rapidly hydrolyzesACh, thereby permitting the return of the re-ceptor protein to its original conformation andreestablishing the barrier for ion movements(Nachmansohn, 1970) .
However tenable the theory of free diffusion ofACh from early neurites to myotomes may be,one must still take into account the recent evi-dence from electrophysiological studies of de-veloping neuromuscular junctions in culture(Robbins and Yonezawa, 1971) . This investiga-tion indicates that chemical transmission beginsabout the time when nerve-muscle contacts arefirst visible by light microscopy . Stimulation ofspinal cord fragments or nerve fibers results inexcitatory junctional potentials in innervatedmyotubes. No response is detected in noninner-vated myotubes, even though they are sensitiveto iontophoretically applied ACh . These dataseem to show that free diffusion of ACh does notoccur through the culture at this stage after stimu-lation . The absence of free diffusion of ACh canmost easily be explained by assuming that thenerve fibers in the culture have matured, andare probably ensheathed and have AChE activityat their junctions. The study by Robbins andYonezawa (1971), which depends on the responseof motor end plates to demonstrate ACh release,cannot be expected to provide information on amuch earlier stage of axonal-myoblast interaction .On the other hand, their evidence that nonin-nervated myotubes are sensitive to iontophoreti-cally applied ACh is pertinent to the previousdiscussion of a possible mechanism for spontaneouscontraction involving AChE-containing subsurface
reticulum and ACh diffusing from sparselysheathed, AChE-free neurites . AChE-containingsubsurface reticulum and sarcoplasmic Ieticulumare prominent in early myotubes, but decrease inlater myotube stages before motor innervation(unpublished observations) . This may suggestthat the mechanism for spontaneous activity isno longer operational and has been replaced byanother.
The authors wish to express their gratitude for theexcellent technical assistance of Miss Mary Budinin-kas and Dr. Domingo Espiritu and the secretarialhelp of Miss Judy Eyrich . The management of theelectron microscope by Mr. Moshe Rosen, and thephase-contrast microscopy done by Mrs. Ida Nathanare gratefully acknowledged .
This work was supported in part by the MuscularDystrophy Associations of America, the Clinical Re-search Center for Parkinson's and Allied Diseases,NS 05184, and the Boris Kidri6 Foundation, Yugo-slavia .
Received for publication 26 April 1971, and in revised form8 June 1971 .
REFERENCES
ALLEN, E . R., and F . A. PEPE . 1965 . Ultrastructure ofdeveloping muscle cells in the chick embryo . Amer.J. Anat. 116 :115.
AUBER, J. 1969 . La myofibrillogenése du muscle strié.II . Vertébrés . J. Microsc. 8 :367 .
BELLHORN, M. B., O. O . BLUMENFELD, and P. M .GALLOP. 1970. Acetyleholinesterase of the humanerythrocyte membrane . Biochem . Biophys. Res.Commun. 39:267 .
BERGMAN, R. A. 1962. Observations on the morpho-genesis of rat skeletal muscle . Bull. Johns HopkinsHosp. 110 :187 .
BERNSOHN, J ., K . D . BARRON, and A. R. HESS. 1962.Multiple nature of acetylcholinesterase in nervetissue . Nature (London) . 195 :285 .
BONICHON, A. 1957. Localisation de l'acétylcholin-estérase dans les muscles striés au cours du dévelop-pement chez l'embryon de poulet. Ann. Histochim .2:301 .
BRzIN, M., V . M . TENNYSON, and P . E. DUFFY. 1966 .Acetylcholinesterase in frog sympathetic and dorsalroot ganglia. A study by electron microscope cyto-chemistry and microgasometric analysis with themagnetic diver . J. Cell Biol. 31 :215 .
BURT, A. M. 1968 . Acetylcholinesterase and cholineacetyltransferase activity in the developing chickspinal cord. J. Exp . Zool. 169 :107 .
CHURCH, J. C. T . 1969 . Satellite cells and myogenesis ;a study in the fruit-bat web . J. Anat. 105 :419 .
TENNYSON ET AL. Acetylcholinesterqo in Myotome of Embryonic Rabbit
719

DESSOUKY, D. A., and R . G. HIBBS. 1965 . An electronmicroscope study of the development of the somaticmuscle of the chick embryo. Amer . J. Anat. 116 :523 .
ECOBICHON, D . J ., and Y. ISRAEL . 1967, Characteriza-tion of the esterases from electric tissue of electro-phorus by starch-gel electrophoresis. Can. J.Biochem . 45:1099 .
ENGEL, W. K. 1961 . Cytological localization ofcholinesterase in cultured skeletal muscle cells . J.Histochem . Cytochem. 9 :66.
ERÄNKO, O ., M. HÄRKÖNEN, A. KoKKo, and L .RÄISÄNEN . 1964. Histochemical and starch gelelectrophoretic characterization of desmo- andlyo-esterases in the sympathetic and spinal gangliaof the rat. J. Histochem . Cytochem. 12 :570 .
EZERMAN, E. B., and H . ISHIKAWA. 1967 . Differentia-tion of the sarcoplasmic reticulum and T system indeveloping chick skeletal muscle in vitro. J. CellBiol. 35 :405 .
FILOGAMO, G. 1964. Le développement de l'activitéacétylcholinestérase dans les muscles des mam-mifères . Acta. Anat . 59 :382 .
FISCHMAN, D. A. 1967. An electron microscope studyof myofibril formation in embryonic chick skeletalmuscle . J. Cell Biol . 32 :557 .
GABELLA, G. 1964 . Le développement de l'activitéAChE dans les muscles, chez les poissons et lesamphibiens . Acta . Anat. 59:382.
GEREBTZOFF, M. A. 1957 . Appareil cholinestérasiquemusculo- tendineux : structure, développement,effet de la dénervation et de la téntomie . ActaPhysiol . Pharmacol. Neer. 6 :419.
GEREBTZOFF, M . A. 1959. Morphogenetic study ofacetylcholinesterase in nervous system and skeletalmuscle . In Cholinesterases. Pergamon Press, Inc .,Elmsford, N . Y . 36.
HAGOPIAN, M ., V. M. TENNYSON, and D . SPIRO . 1970 .Cytochemical localization of cholinesterase in em-bryonic rabbit cardiac muscle. J. Histochem. Cyto-chem . 18 :38 .
HAMBURGER, V. 1963. Some aspects of the em-bryology of behavior . Quart . Rev. Biol. 38:342.
HAMBURGER, V., M. BALABAN, R . OPPENHEIM, andE. WENGER. 1965. Periodic motility of normal andspinal chick embryos between 8 and 17 days ofincubation . J. Exp. Zool. 159 :1 .
HAY, E . D. 1963 . The fine structure of differentiatingmuscle in the salamander tail . Z. Zellforsch . Mikrosk .Anat. 59 :6 .
HAY, E. D. 1968 . Organization and fine structure ofepithelium and mesenchyme in the developingchick embryo . In Epithelial-Mesenchymal Inter-actions . R. Fleischmajer, editor . The Williams &Wilkins Co ., Baltimore. 31 .
HEUSON-STIENNON, J . A. 1965. Morphogénese de lacellule musculaire striée au microscope électroniqueI. Formation des structures fibrillaires. J. Microsc .4 :657 .
7 2 0 THE JOURNAL OF CELL BIOLOGY . VOLUME 51, 1971
HIRANO, H . 1967 . Ultrastructural study on the mor-phogenesis of the neuromuscular junction in theskeletal muscle of the chick . Z . Zellforsch . Mikrosk.Anat. 79:198 .
ISHIKAwA, H. 1966 . Electron microscopic observationsof satellite cells with special reference to the devel-opment of mammalian skeletal muscles . Z. Anat.Entwicklungsgesch. 125 :43 .
ISHIKAWA, H. 1968 . Formation of elaborate networksof T-system tubules in cultured skeletal muscle withspecial reference to the T-system formation. J. Cell38 :51 .
ISHIKAWA, H., R. BISCHOFF, and H . HOLTZER. 1968 .Mitosis and intermediate-sized filaments in de-veloping skeletal muscle. J. Cell Biol. 38 :538 .
KARCZMAR, A . G . 1963 . Ontogenesis of cholinesterasesB. Ontogenesis of ChE's in the vertebrate muscleand heart. In Handbuch der ExperimentellenPharmakologie. G. B. Koelle, editor. Springer-Verlag KG ., Berlin . 15 :129 .
KELLY, A. M., and S . I . ZACKS . 1969. The histo-genesis of rat intercostal muscle . J. Cell Biol. 42 :135.
KOELLE, G . B. 1951 . The elimination of enzymaticdiffusion artifacts in the histochemical localizationof cholinesterases and a survey of their cellular dis-tributions. J. Pharmacol . 103:153.
KOELLE, G. B., and J. S. FRIEDENWALD. 1949. Ahistochemical method for localizing cholinesteraseactivity. Proc. Soc . Exp. Biol. Med. 70 :617 .
KOELLE, W. A., K. S. HOSSAINI, P . AKBARZADEH,and G. B. KOELLE . 1970 . Histochemical evidenceand consequences of the occurrence of isoenzymes ofacetylcholinesterase. J. Histochem. Cytochem . 18 :812 .
KUPFER, C., and G . B. KOELLE . 1951 . A histochem-ical study of cholinesterase during formation of themotor end plate of the albino rat . J. Exp. Zool.116 :397 .
LANGLEY, O. K., and D. M. LANDON. 1969 . Copperbinding at nodes of Ranvier : a new electron histo-chemical technique for the demonstration ofpolyanions . J. Histochem . Cytochem . 17 :66 .
LENTZ, T. L. 1969 . Development of the neuromuscu-lar junction . I. Cytological and cytochemicalstudies on the neuromuscular junction of differen-tiating muscle in the regenerating limb of the newtTriturus. J. Cell Biol. 42 :431 .
Li, C.-L ., K . ENGEL, and I. KLATZO . 1959. Someproperties of cultured chick skeletal muscle withparticular reference to fibrillation potentials . J.Cell Comp . Physiol. 53 :421 .
MAURO, A. 1961 . Satellite cell of skeletal musclefibers . J. Cell Biol. 9 :493.
MCCAMAN, M. W., L. R. ToMEY, and R . E. Mc-CAMAN. 1968. Radiomimetric assay of acetyl-cholinesterase activity in sabmicrogram amounts oftissue . Life Sei . 7 (Pt. 11) :233 .
MIRANDA, A. F. 1971 . Ultracytochemical studies of

cholinesterase (ChE) in muscle of normal anddystrophic mice. Anat . Rec. 169 :381 .
MUIR, A. R., A. H. M. KANJI, and D . ALLBROOK .1965. The structure of satellite cells in skeletalmuscle . J. Anat . 99 :435 .
MUMENTHALER, M., and W. K . ENGEL, 1961 . Cyto-logical localization of cholinesterase in developingchick embryo skeletal muscle. Acta Anat . 47 :274 .
NACHMANSOHN, D . 1938 . Changements de la cholin-estél ase dans le muscle strié . C. R . Soc. Biol. 128 :599.
NACHMANSOHN, D . 1939. Choline esterase in voluntarymuscle . J. Physiol. (London) . 95 :29 .
NACHMANSOHN, D . 1940. Choline esterase in brain andspinal cord of sheep embryos. J. Neurophysiol .3 :396.
NACHMANSOHN, D . 1970 . Proteins in excitable mem-branes . Science (Washington) . 168 :1059.
PALADE, G . E . 1952 . A study of fixation for electronmicroscopy. J. Exp. Med. 95 :285 .
PRZYBYLSKI, R . J., and J. M. BLUMBERG . 1966 .Ultrastructural aspects of myogenesis in the chick .Lab . Invest. 15 :836.
REYNOLDS, E . S. 1963 . The use of lead citrate at highpH as an electron-opaque stain in electron micros-copy . J. Cell Biol. 17:208.
RICHTER, D ., and P. G. CROFT . 1942 . Blood esterases.Biochem . J . 36 :746 .
ROBBINS, N ., and T. YONEZAWA. 1971 . Developingneuromuscular junctions : first signs of chemicaltransmission during formation in tissue culture .Science (Washington) . 172 :395 .
ROMANES, G. J. 1941 . The development and signific-ance of the cell columns in the ventral horn of thecervical and upper thoracic spinal cord of therabbit . J. Anat . 76 :112 .
ROSENBLUTH, J . 1962 . Subsurface cisterns and theirrelationship to the neuronal plasma membrane . J.Cell Biol. 13 :405.
SABATINI, D. D., K. BENSCH, and R. J. BARRNETT.1963 . Cytochemistry and electron microscopy . Thepreservation of ultrastructule and enzymatic ac-tivity by aldehyde fixation . J. Cell Biol. 17 :19.
SHEN, S. C . 1958 . Changes in enzymatic patternsduring development . In A Symposium on theChemical Basis of Development. W. D. McElroyand B. Glass, editors. The Johns Hopkins Press,Baltimore . 416.
SLOTWINER, P ., and V. M. TENNYSON . 1969 . Thedevelopment of acetylcholinesterase activity withinskeletal muscle of the embryonic rabbit . Anat . Rec .163 :326.
TENNYSON, V. M . 1970 . The fine structure of theaxon and growth cone of the dorsal root neuro-blast of the rabbit embryo . J. Cell Biol . 44 :62 .
TENNYSON, V. M., and M . BRZIN . 1970 . The appear-ance of acetylcholinesterase in the dorsal rootneuroblast of the rabbit embryo . A study by elec-tron microscope cytochemistry and microgaso-metric analysis with the magnetic diver . J. CellBiol . 46 :64 .
VENERONI, G ., and M. R. MURRAY. 1969. Formationde novo and development of neuromuscular junc-tions in vitro. J. Embryol. Exp . Morphol. 21 :369 .
VISINTINI, F ., and R. LEVI-MONTALCINI . 1939. 7 .Relazione tra differenziazione strutturale e funzion-ale de centri e delle vie nervose nell'embrione dipolio . Schweiz . Arch. Neurol . Psychiat . 44 :119 .
WATSON, M. L . 1958 . Staining of tissue sections forelectron microscopy with heavy metals . J. Biophysic .Biochem . Cytol. 4 :475 .
ZACKS, S . I . 1954. Esterases in the early chick embryo .Anat. Rec . 118 :509 .
ZELENA, J . 1962 . The effect of denervation on muscledevelopment. In The Denervated Muscle . E .Gutmann, editor. Academia, Prague. 103 .
TENNYSON ET AL. Acetylcholinesterase in Myotome of Embryonic Rabbit
721