telencephalic connections in the pacific hagfish …bi250b/2009/papers/wicht_northcutt...p2mc...
TRANSCRIPT

Telencephalic Connections in the PacificHagfish (Eptatretus stouti), With SpecialReference to the Thalamopallial System
HELMUT WICHT1* AND R. GLENN NORTHCUTT2
1Klinikum der Johann Wolfgang Goethe-Universitat, Dr. Senckenbergische Anatomie,Institut fur Anatomie II (Experimentelle Neurobiologie),
Theodor-Stern-Kai 7, 60590 Frankfurt, Federal Republic of Germany2Neurobiology Unit, Scripps Institution of Oceanography and Department of Neurosciences,
School of Medicine, University of California San Diego, La Jolla, California 9203-0201
ABSTRACTThe pallium of hagfishes (myxinoids) is unique: It consists of a superficial ‘‘cortical’’
mantle of gray matter which is subdivided into several layers and fields, but it is not clearwhether or how these subdivisions can be compared to those of other craniates, i.e., lampreysand gnathostomes. The pallium of hagfishes receives extensive secondary olfactory projec-tions (Wicht and Northcutt [1993] J. Comp. Neurol. 337:529–542), but there are noexperimental data on its nonolfactory connections. We therefore investigated the pallial anddorsal thalamic connections of the Pacific hagfish. Injections of tracers into the palliumlabeled many cells bilaterally in the olfactory bulbs. Other pallial afferents arise from thecontralateral pallium, the dorsal thalamic nuclei, the preoptic region, and the posteriortubercular nuclei. Descending pallial efferents reach the preoptic region, the dorsal thalamus,and the mesencephalic tectum but not the motor or premotor centers of the brainstem.Injections of tracers into the dorsal thalamus confirmed the presence of reciprocal thalamopal-lial connections. In addition, these injections revealed that there is no ‘‘preferred’’ pallialtarget for the ascending thalamic fibers; instead, ascending thalamic and secondary olfactoryprojections overlap throughout the pallium. The mesencephalic tectum and tegmentum,which receive afferents from a variety of sensory sources, are interconnected with the dorsalthalamus; thus, ascending nonolfactory sensory information may reach myxinoid pallia via atectal-thalamic-telencephalic route. A comparative analysis of pallial organization revealsthat the subdivisions of the pallium in gnathostomes (i.e., medial, dorsal, and lateral pallia)cannot be recognized with certainty in hagfishes. J. Comp. Neurol. 395:245–260, 1998.r 1998 Wiley-Liss, Inc.
Indexing terms: myxinoidea; phylogeny; forebrain; pallium; diencephalon
In amniotes, derivatives of the telencephalic pallium(i.e., the dorsal ventricular ridge in sauropsids and thevarious cortical fields in mammals) are generally regardedas the ‘‘highest’’ integrative centers of the entire neuraxis.This is probably due to the fact that they are, at least inhumans, responsible for the transformation of reflectorysensorimotor activities into the conscious events of percep-tion, recognition, decision, and action.
There is no direct line of pallial evolution ‘‘from fish toman.’’ The earlier scenario of pallial evolution held thatprimitive piscine pallia were entirely dominated by second-ary olfactory projections. An invasion of dorsal thalamicfibers into the pallium was thought to have occurred intetrapods, thus giving rise to the sensory pallial areascharacteristic of mammals. This scenario, however, had to
be rejected based on comparative data gathered in the lasttwo decades (reviewed in Northcutt, 1995). Ascendingsensory pathways reach the pallia of all gnathostomes( jawed craniates) investigated to date, and all gnathos-tomes display one or many pallial fields that receive no oronly sparse secondary olfactory afferents from the olfac-
Grant sponsor: NIH; Grant numbers: NS24699/DC01081; Grant sponsor:Dr. Senckenbergische Stiftung (Frankfurt/Main).
*Correspondence to: Helmut Wicht, Klinikum der Johann WolfgangGoethe-Universitat, Dr. Senckenbergische Anatomie, Institut fur AnatomieII (Experimentelle Neurobiologie), Theodor-Stern-Kai 7, 60590 Frankfurt,Federal Republic of Germany. E-mail: [email protected]
Received 23 September 1997; Revised 20 January 1998; Accepted 27January 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 395:245–260 (1998)
r 1998 WILEY-LISS, INC.

tory bulb. Thus, it is highly likely that the pallium of thecommon ancestor of all gnathostomes was not purelyolfactory but contained separate pallial fields that werededicated to the processing of olfactory and ascendingnonolfactory sensory information (Northcutt, 1995).
However, the history of pallial evolution before theoccurrence of gnathostomes is less clear. Recent studies injawless craniates (i.e., lampreys and hagfishes) have shedsome light on the very early evolution of the palliumamong craniates and have revealed substantial differencesbetween jawless craniates and gnathostomes. Experimen-tal studies of the secondary olfactory projections in lam-preys (petromyzontids; Northcutt and Puzdrowski, 1988)have shown that these projections are indeed very exten-sive and reach all pallial fields, as predicted by the earlierscenario of pallial evolution. However, it has also beenshown that the pallium of lampreys is not entirely olfac-tory, because there is a substantial, presumably sensory,input from diencephalic and mesencephalic cell groups toits medial subdivision (Polenova and Vesselkin, 1993;Northcutt and Wicht, 1997).
The pallium of the only other recent clade of jawlesscraniates, the hagfishes (myxinoids), is highly differenti-ated from a cytoarchitectural point of view (Jansen, 1930;Wicht and Northcutt, 1992). It consists of a superficial‘‘cortical’’ mantle of gray matter that is subdivided intovarious strata and fields, but it is not clear whether andhow these subdivisions correspond to the medial, lateral,and dorsal subdivisions of the pallium of other craniates.Similar to the situation in lampreys, the pallium in
myxinoids is dominated by secondary olfactory projections(Wicht and Northcutt, 1993). However, there are no dataon its nonolfactory connections. It is therefore not clearwhether nonolfactory and, in particular, ascending tha-lamic projections occur in myxinoids; nor is it clear whetherthese projections, if they exist, have a particular pallialfield as their target. In order to answer these questions, weanalyzed the connections of the pallium and the thalamusof the Pacific hagfish, Eptatretus stouti, by means oftract-tracing techniques.
MATERIALS AND METHODS
Twenty-six adult and subadult Pacific hagfishes (Eptatre-tus stouti, body length between 250 and 450 mm) wereused for this study. All animals were treated according tothe guidelines outlined in the National Institutes of HealthGuide for the Care and Use of Laboratory Animals. Theanimals were caught in baited offshore traps near theScripps Institution of Oceanography (La Jolla, CA) andtransferred into aquaria with running sea water at 10oC.Prior to any surgical manipulation the animals wereanesthetized with MS 222 (tricaine-methanesulfonate,diluted 1:10,000 in sea water).
The in vitro modification of the DiI technique (Godementet al., 1987) was used in three cases. In short, theanesthetized animals were transcardially perfused with0.1 M phosphate-buffer (PB) at pH 7.4, followed by afreshly prepared solution of 4% paraformaldehyde (PFA)
Abbreviations
’a’ nucleus ‘‘a’’ of Kusunokiaol octavolateral areaap area praetectalisBO olfactory bulbBOg glomerular layer of BOBOm mitral layer of BOBOp periglomerular layer of BOcg central mesencephalic graycoh habenular commissurecoib interbulbar commissurecopo postoptic commissurectp commissure of the posterior tubercular areadcn dorsal column nucleiDi diencephalonfai internal arcuate fibersfr fasciculus retroflexusfrs superior reticular formationfri inferior reticular formationgs posterior (‘‘saccular’’) octaval gangliongu anterior (‘‘utricular’’) octaval gangliongV trigeminal ganglionHa habenulaHYinf infundibular hypothalamic nucleusinp interpeduncular nucleuslb bulbar lemniscusmIX,X glossopharyngeal and vagal motor nucleusMes mesencephalonmV trigeminal motor nucleusnIX,X glossopharyngeal and vagal nervenVII facial nerveNCd dorsal nucleus of central prosencephalic complexNCm medial nucleus of central prosencephalic complexNCvl ventrolateral nucleus of central prosencephalic complexnflm nucleus of the medial longitudinal fasciclenII optic nervenl olfactory nerve(s)nlla nervus lateralis aP1 pallial layer 1
P2 pallial layer 2P2co compact part of pallial layer 2P2l lateral part of pallial layer 2P2mc magnocellular part of pallial layer 2P2pc parvocellular part of pallial layer 2P3 pallial layer 3P4l lateral part of pallial layer 4P4m medial part of pallial layer 4P5 pallial layer 5POe external preoptic nucleusPOim intermediate preoptic nucleusPOp periventricular preoptic nucleusra nuclei of the raphesIX,X sensory nucleus (lobe) of the glossopharyngeal and vagal
nervessc spinal cordsco subcommissural organsp1 first spinal nervespocc spinooccipital nervest striatumsV sensory nucleus of the trigeminusTel telencephalonTHa anterior nucleus of the thalamusTHdi diffuse nucleus of the thalamusTHe external nucleus of the thalamusTHi internal nucleus of the thalamusTHpco paracommissural nucleus of the thalamusTHsh subhabenular nucleus of the thalamusTHt triangular nucleus of the thalamustm mesencephalic tectumTPl lateral nucleus of the posterior tubercular areaTPm medial nucleus of the posterior tubercular areatrolp deep part of the lateral olfactory tracttrols superficial part of the lateral olfactory tracttrpht palliohabenular and -thalamic tracttrtb tectobulbar tractvmes mesencephalic ventricle
246 H. WICHT AND R.G. NORTHCUTT

in PB. The fixed brains were dissected from the skulls, andthe pallium (two cases) and dorsal thalamus (one case)were injected with minute crystals of DiI (MolecularProbes, Eugene, OR). The brains were stored in PFA in PBat 40°C for 2 weeks. Afterwards, they were cut on aVibratome in the transverse plane at 50 µm, counter-stained with m-phenylene-diamine (Quinn and Weber,1988), and viewed under fluorescent light.
In eighteen cases, biotinylated dextran-amines (BDA,Molecular Probes) were used as tracers (Veenman et al.,1992). The animals were anesthetized, the dorsal aspect ofthe brain was exposed and BDA dried onto the tips of veryfine insect pins was applied to the pallium (seven cases),the pallium and the striatum (two cases), the pallium andthe central prosencephalic complex (two cases), and thedorsal thalamus (seven cases). After survival periods of 6to 9 days, the animals were reanesthetized, perfusedtranscardially with PB followed by PFA in PB, which alsocontained 1.8% L-lysine and 0.2% sodium-iodate. Thebrains were imbedded in gelatine, and transverse frozensections were cut at 40 µm on a sliding microtome.Free-floating sections were incubated with an avidin-biotinylated horseradish peroxidase complex (ABC-HRP)prepared and diluted according to the instructions of themanufacturer (Vectastain, Vector Laboratories, Burlin-game, CA). The reactive sites were visualized by usingAdam’s (1981) protocol for the detection of HRP activity.Afterwards, the sections were mounted on slides, dehy-drated, and coverslipped. Some series were counterstainedwith neutral red. The in vivo HRP technique was used infive cases for large injections into the hemispheres. Theprotocol is described in Wicht and Northcutt (1993).
In general, the BDA method provided the most consis-tent and satisfactory results in both retrograde and antero-grade connections, and the majority of the data reportedhere stems from our BDA material. The HRP techniqueproved to be much less sensitive. Only restricted antero-grade and retrograde transport was revealed and thetechnique required the injection of large amounts of tracer,thus producing very large injection sites, often coveringthe entire telencephalon. The DiI method was very sensi-tive with respect to anterograde labeling, but it alsoresulted in very large injection sites that were difficult todelineate from surrounding structures.
RESULTS
We will first present the results of pallial and largertelencephalic injections, followed by those of thalamicinjections. As noted, the BDA method provided the mostconsistent data and most results are based on BDA experi-ments. Throughout this paper, we will use our own termi-nology (Wicht and Northcutt, 1992) for the diencephalicand telencephalic cell groups. The terminology for themesencephalon and rhombencephalon is from Jansen(1930), with modifications as suggested by Ronan (1989)and Ronan and Northcutt (1990). Figure 1 is a dorsal viewof the brain of a Pacific hagfish indicating the levels of thetransverse sections shown in Figures 2 and 6, which chartthe results of telencephalic and thalamic injections, respec-tively.
Injections into the pallium
Injection sites. The pallium of hagfishes is a complex,layered structure that covers the dorsolateral aspect of the
telencephalic hemispheres (Jansen, 1930; Wicht and North-cutt, 1992). It consists of five alternating cellular andfibrous layers at midtelencephalic levels (Fig. 3A); thecellular layers can again be subdivided into various subdi-visions (Fig. 2B-D; see also Wicht and Northcutt, 1992).Single injections of BDA resulted in relatively small injec-tion sites that covered only small parts of the pallium.Usually, two injections (one lateral and one more medial,Fig. 2B,C) were required to achieve satisfactory injectionsites. Such injection sites (Fig. 2B) typically included allpallial layers at midtelencephalic levels, sparing onlysegments of pallial layer 2: the most posterior regions (Fig.2D), parts of the most medial, compact region (P2co), andparts of the most ventral, magnocellular region (P2mc).Due to the close proximity of the pallium to the underlyingstriatum, the larger injections also affected small parts ofthis latter structure (Fig. 2B). However, we also performeda number of smaller injections, that did not involve thestriatum, and the results of these injections were consis-tent with those observed after the larger injections.
Intra- and interhemispheric connections. As re-ported earlier (Wicht and Northcutt, 1993), hagfishes havean extensive bilateral secondary olfactory projection viathe lateral olfactory tract to the pallium. The lateralolfactory tract consists of a superficial component and adeep component; the latter occupies pallial layer 5, theformer pallial layer 1 (Fig. 3B). Numerous secondaryolfactory fibers decussate in the interbulbar and habenu-lar commissures and thus give rise to the contralaterallateral olfactory tract which also splits into a deep tractand a superficial tract. Therefore, our pallial injectionsalways affected the secondary olfactory system. Retro-gradely, large numbers of neurons were labeled in theolfactory bulbs, both ipsilateral and contralateral to theinjection site (Fig. 2A). The neurons were typically bipolar(Fig. 4A) and concentrated in the mitral and periglomeru-lar layers of the olfactory bulb. Anterogradely, the deepand the superficial lateral olfactory tracts were labeledbilaterally (Fig. 2B-D).
Pallial injections also revealed a large number of labeledstructures that were not seen after injections into theolfactory bulb, however. These structures therefore repre-sent intrahemispheric and interhemispheric afferents andefferents of the pallium. A few retrogradely labeled cellswere observed in the lateral parts (P2l, P2mc, P4l) of thecontralateral pallium, but a large number of contralater-ally projecting cells was found in the medial subdivisionsof the pallium, i.e., in the compact part of pallial layer 2(P2co) and in the medial part of pallial layer 4 (P4m, Figs.2B-D; 4C). Their axons decussate in the interbulbar andhabenular commissures (Fig. 2D,E), but they could not bedistinguished from the secondary olfactory fibers decussat-ing in these commissures. Similarly, the exact sites oftermination of the commissural palliopallial projectionscould not be established due to the simultaneous labelingof the extensive secondary olfactory tracts.
The striatum ipsilateral to the injection site containedrelatively few labeled cells (Fig. 2B,C), but was traversedby a strong fiber tract that appeared to arise from theanterior part of the pallium and to descend into thepreoptic region (Fig. 2B,C). There, a densely labeled andsharply circumscribed neuropil and numerous labeledperikarya were found adjacent to the tract in the external
TELENCEPHALIC CONNECTIONS IN HAGFISHES 247

preoptic nucleus (Figs. 2B-D; 4B). Further caudally, in thediencephalon (Fig. 2D), single decussating fibers were seento split off from the tract and enter the postoptic commis-sure (Fig. 2D). Again more caudally (Fig. 2D,F), the fibertract became more diffuse and ascended into the dorsalthalamus.
No labeled structures were revealed in the medial andthe ventrolateral subnuclei of the central prosencephaliccomplex, even after large pallial injections. However, theipsilateral dorsal nucleus contained numerous retro-gradely labeled cells and dense terminal labeling at itsdorsal border (Fig. 2D).
Diencephalic and mesencephalic connections. Alarge number of retrogradely labeled cells was observed inthe ipsilateral external and internal nuclei of the dorsalthalamus (Figs. 2F,G; 4D). A few cells were also seencontralaterally in these nuclei and ipsilaterally in theanterior thalamic nucleus (Fig. 2D). The medial and thelateral nuclei of the posterior tubercular area (Figs. 2G;5A,C) contained heavily labeled cells both ipsilaterally andcontralaterally; occasionally, single-labeled cells were seenin the anterior parts of the central mesencephalic gray(Figs. 2H; 5B). There were no labeled cells caudal to thelevel shown in Figure 2H; in particular, pallial injections
produced no labeling in the locus coeruleus or in the nucleiof the raphe (but see below, section on pallial/striatalinjections).
Both the diencephalon and mesencephalon receive heavyprojections from the telencephalic pallium. As seen inFigure 2E-H, the entire dorsal superficial aspect of bothregions was covered by numerous labeled fibers of verythin caliber. A diffuse terminal-like, punctate labeling wasevident throughout these dorsal regions in the diencepha-lon and mesencephalon (Figs. 4D; 5A, arrowheads). In
Fig. 2. A-H: Drawings of rostral (A) to caudal (H) transversesections at the levels indicated in Figure 1, charting the results of aninjection of biotinylated dextran amines into the pallium. The injec-tion site is visible in B and C. The blackened areas in C represent thetracks of the needles used to inject the tracer; the shaded area in B andC represents the extent of a typical injection site. Throughout allpanels, retrogradely labeled cell bodies are symbolized by triangles,fibers by thin lines, and terminal fields by fine stippling. Circles in E,F, and H represent labeled cells that occurred after larger injectionsthat also involved the striatum. The arrowheads in C point to aterminal field ventral to the magnocellular part of pallial layer 2; thisfield originates from fibers of the deep part of the lateral olfactorytract. For abbreviations, see list. Scale bar 5 0.5 mm.
Fig. 1. Dorsal view of the brain of the Pacific hagfish, Eptatretus stouti. The vertical lines indicate thelevels of individual rostral (A) to caudal (H) transverse sections in Figures 2 and 6, respectively. Forabbreviations, see list. Scale bar 5 1 mm.
248 H. WICHT AND R.G. NORTHCUTT

Figure 2
TELENCEPHALIC CONNECTIONS IN HAGFISHES 249

Figure 3
250 H. WICHT AND R.G. NORTHCUTT

some areas, these terminals condensed to form sharplycircumscribed neuropils: one in the ventrolateral part ofthe habenula (Fig. 2D), one over the ipsilateral internalthalamic nucleus (Fig. 2E), and a bilateral neuropil continu-ing from the ventral part of the external thalamic nucleus(Fig. 2F) all the way into the lateroventral part of themesencephalic tectum (Figs. 2H; 5A, arrows). Anothercircumscribed terminal field occurred immediately dorsalto the cerebral aqueduct and the subcommissural organ inthe pretectal and tectal regions (Figs. 2B, 5B; the subcom-missural organ of myxinoids is very elongated and extendswell into the mesencephalon; Adam, 1963). In contrast toan earlier claim, based on preliminary DiI-experiments(Wicht and Northcutt, 1992), we did not observe a correla-tion between the position of these neuropils and thelocation of the pallial injection sites in the present study.The labeled neuropils occupied the same positions aftersmall injections of BDA into different (i.e., medial vs.lateral) regions of the pallium; only the intensity of thestaining varied with the size of the injection site.
It is, of course, possible that some of the terminalformations in the di- and mesencephalon actually arisefrom collaterals of thalamic neurons which also project tothe pallium. However, the distinctive and circumscribedterminal fields described above were only observed afterpallial, but not after thalamic injections; thus, they prob-ably represent pallial, and not thalamic efferents.
Due to the reciprocity of the connections between thepallium and the diencephalon and mesencephalon, it wasnot possible to identify the projection routes of the indi-vidual projection systems with certainty. However, twomain channels of communication appear to exist: oneventral route and one dorsal route. The ventral route is viathe tract descending from the anterior part of the palliuminto the preoptic area (see above); fibers of that tract jointhe postoptic commissure, with which they travel caudallytowards the dorsal thalamus, the posterior tuberculararea, and the mesencephalic tectum (Fig. 2E-G). Thedorsal communication route consists of fibers that collectat the posterior margin of the pallium, rostral to the
habenular commissure (Fig. 2D); arranged in a conspicu-ous radial pattern, these fibers give rise to the circum-scribed labeled neuropils in the dorsal nucleus of thecentral prosencephalic complex (Fig. 2D), in the habenula(Fig. 2D), and in the internal thalamic nucleus (Fig. 2E).Some of the fibers of the dorsal communication routedecussate in the ventral part of the habenular commissure(Fig. 2E), in a pattern similar to that of Jansen’s (1930)palliohabenular and palliothalamic tracts.
Injections into the pallium and the striatum
In two cases, the injection sites were centered in thestriatum, at the approximate level shown in Figure 2B.The overlying pallium was also affected by these injec-tions, but the central prosencephalic complex medial to thestriatum was unaffected. In these cases, we observed apattern of retrograde and anterograde labeling virtuallyidentical to that one described for the restricted pallialinjections but with additional retrograde labeling. Labeledcells were seen in the ipsilateral diffuse and intracommis-sural nuclei of the ventral thalamus (circles in Fig. 2E,F).Other labeled perikarya (circles in Fig. 2H) were observedin the ipsilateral mesencephalic tectum, the superiornuclei of the raphe, and in a region medial and ventral tothe octavolateral area that might correspond to the locuscoeruleus of other craniates.
Injections into the pallium and the centralprosencephalic complex
The additional projections revealed by these injectionswere mainly reciprocal connections with the infundibularnucleus of the hypothalamus. This finding confirms theobservations of Amemiya and Northcutt (1996) on theconnections of the central prosencephalic complex.
Injections into the thalamus
Injection sites. The thalamic injection sites coveredthe central part of the external and internal dorsal tha-lamic nuclei in about half of our injections (Fig. 6E,F). Inseveral other cases, either the pretectal area or the habenu-lar commissure was affected by spread of the tracer. Thesecases were carefully analyzed and compared to the morerestricted injections and only those connections that werealso revealed by the more restricted injections are reportedhere. In addition, we also performed very small injectionsthat were restricted to the external thalamic nucleus. Thegeneral labeling was very weak in these cases but consis-tent with the results from the other injections.
Connections with the rhomb- and mesencephalon.
Retrogradely labeled cells were seen in the ipsilateral andcontralateral inferior reticular formations (Fig. 6H), aswell as in the ipsilateral superior reticular formation (Fig.2G). Very occasionally, single labeled cells were seen in theipsilateral sensory nucleus of the trigeminus (Figs. 6G,7C). Numerous retrogradely labeled cells occurred bilater-ally in the central mesencephalic gray, among the fibers ofthe bulbar lemniscus and the tectobulbar tract and in themesencephalic tectum (Figs. 6C, 7A). The pretectal areawas consistently seen to contain labeled cells; a few labeledneurons were also observed in the ipsilateral medialnucleus of the posterior tubercular area. Terminal-like,punctate labeling was evident throughout most of the
Fig. 3. Photomicrographs of different preparations of pallial sec-tions from Eptatretus stouti. All are from approximately the sametransverse level and have the same orientation (the pial surface is tothe top) and magnification. A: Cresyl violet preparation, cut fromparaffin at 10 µm. Note the alternating layers of gray and whitematter (P1-P5). B: Results of a horseradish peroxidase injection intothe ipsilateral olfactory bulb, counterstained with methylene green.The superficial layer of white matter (P1) is largely occupied by thesuperficial part of the lateral olfactory tract (trols); the deep palliallayer (P5) contains the deep part of the lateral olfactory tract (trolp;Wicht and Northcutt, 1993). C: Results of a large injection of biotinyl-ated dextran-amines (BDA) into the ipsilateral diencephalon. Numer-ous varicose fibers (some are indicated by arrowheads) are visible inall pallial layers; a large number of retrogradely labeled cells is alsovisible, particularly in pallial layers 2 and 4. Pallial layer 3 is mainlytaken up by fiber bundles (arrows) which will coalesce more caudallyto form the palliohabenular tract and palliothalamic tract (see Fig.2D). D: Results of a small injection of BDA into the ipsilateral dorsalthalamus. Compared to the larger injection shown in C, the number ofretrogradely labeled pallial neurons and the intensity of their labelingis much reduced, and the distinct fiber bundles in pallial layer 3 arenot visible. Due to the less intense retrograde labeling of pallial cells,however, the photomicrograph shows the evenly distributed system ofvaricose fibers and terminal-like puncta (some are indicated byarrowheads) more clearly than in C. For abbreviations, see list. Scalebar 5 200 µm.
TELENCEPHALIC CONNECTIONS IN HAGFISHES 251

tectal area (Fig. 6G). The inferior and superior reticularformations contained numerous coarse and winding fibers(Figs. 6G,H, 7B), but it was impossible to discern whetherthey were ascending axons of reticular neurons, or whetherthey were descending tectal or thalamic fibers.
Intradiencephalic connections. In the immediate vi-cinity of the injection sites, many labeled cells were foundin the external and internal thalamic nuclei and amongthe fibers of the postoptic commissure (in the intracommis-sural nucleus of Wicht and Northcutt, 1992). There wereno labeled cells in the triangular nucleus (Fig. 6E) and onlya few cells in the diffuse nucleus of the ventral thalamus.Contralaterally, a smaller number of cells was observed inthe same areas. A few labeled cells were seen in theipsilateral infundibular nucleus of the hypothalamus andin the ventral and posterior part of the habenula (Fig. 6C,D).
Connections with the hemispheres. A distinct fibertract left the injection sites ventrally, joined the postopticcommissure and ran forward with this commissure to the
preoptic region (Fig. 6C-E). There, a large number of fibersdecussated in the postoptic commissure. Ipsilaterally andcontralaterally, the fibers continued rostrally and gave riseto terminal formations in the external preoptic nuclei (Fig.6A-C). Numerous retrogradely labeled cells were foundintermingled between those fibers and terminals (Fig.6A-C). A few fibers continued even further rostrally andentered the striatum; this projection was more pronouncedipsilaterally than contralaterally. A few labeled cells alsooccurred in the ipsilateral striatum (Fig. 6A).
Numerous labeled structures were observed in the pal-lium. Labeled perikarya were seen in the cellular layers,more ipsilaterally than contralaterally and more rostrallythan caudally (Fig. 6 A-C). The distribution of labeled cellswithin the various subfields of the pallial layers was nothomogeneous. Most cells were seen in the parvicellularand the lateral parts of pallial layer 2 and in the lateralpart of pallial layer 4 (Fig. 3D). The compact and themagnocellular parts of pallial layer 2 contained almost nolabeled cells. Large numbers of varicose fibers and terminal-
Fig. 4. Photomicrographs of transverse sections through variousforebrain regions after injections of biotinylated dextran-amines intothe pallium. Dorsal and lateral directions in B and D are indicated by dand l, respectively. A: Retrogradely labeled cells in the mitral layer(BOm) of the ipsilateral olfactory bulb. B: Two retrogradely labeledcells in the ipsilateral external preoptic nucleus (at the approximatelevel shown in Fig. 2C), immediately adjacent to a heavily labeledterminal field. Also note the terminal-like punctate labeling through-out the neuropil. Neutral red counterstain. C: The medial regions ofthe contralateral hemisphere. Labeled fibers decussate in the interbul-
bar commissure (coib), and a large number of retrogradely labeledcells occurs in the medial part of pallial layer 4 (P4m) and in thecompact part of pallial layer 2 (P2co). Note that the dorsal part of thecentral prosencephalic complex (NCd) is entirely devoid of labeling.D: Labeled cells in the ipsilateral internal (THi) and external (THe)nuclei of the dorsal thalamus (at the approximate level shown in Fig.2F). The terminal-like punctate labeling is far heavier dorsally (in theexternal nucleus) than ventrally (in the internal nucleus). For abbre-viations, see list. Scale bars 5 500 µm in A, 50 µm in B, 200 µm in C,100 µm in D.
252 H. WICHT AND R.G. NORTHCUTT

like puncta were distributed evenly and diffusely through-out all pallial layers (Figs. 3D; 6A-C). In cases of relativelylarge injections that covered most of the dorsal thalamus,a distinct labeled fiber system was seen in the pallium. Atmidtelencephalic levels (Figs. 3C, 6B) the fibers formedsmall bundles that ran in pallial layer 3; more posteriorly(Fig. 6C) the fibers arranged in the radial pattern typical ofthe palliohabenular and palliothalamic tract (see above).Some of the fibers decussated in the ventral part of thehabenular commissure (Fig. 6D).
Notably, there were no labeled structures in any of thenuclei of the central prosencephalic complex followinginjections of tracers into the thalamus.
Brief summary of the results
In summary, our experiments confirm the presence ofextensive bilateral secondary olfactory projections to thepallium in myxinoids. Retrogradely labeled neurons in theolfactory bulb were higher in number and density than inany other brain area after pallial injections. Labeling alsorevealed that the pallium is heavily and reciprocallyinterconnected with the contralateral pallium, the preop-tic area, and the dorsal thalamus. The posterior tuberculararea is another source of ascending projections to thepallium. After large injections that also involved thestriatum, additional retrogradely labeled cells were seenin the mesencephalic tectum, in the nuclei of the raphe,and in an area possibly homologous to the locus coeruleus.Descending pallial projections reach the dorsal thalamus,the pretectum and the tectum, but not the motor orpremotor centers of the brainstem. There are no connec-tions between the pallium and the ventromedial andmedial subnuclei of the central prosencephalic complex;the dorsal nucleus, on the other hand, shows reciprocalconnections with the overlying pallium.
Injections of tracers into the dorsal thalamic nuclei (i.e.,the external and internal nuclei of the thalamus) labeled alarge number of cells in the pallium, as well as an evenlydistributed system of terminal structures in all palliallayers and fields. No connections were observed betweenthese dorsal thalamic nuclei and the central prosence-phalic complex. Other reciprocal connections were estab-lished between the dorsal thalamic nuclei and the stria-tum, the preoptic area, the mesencephalic tectum, and thereticular formation. Apart from rare, single, labeled cellsin the trigeminal sensory nucleus, there was no evidence
Fig. 5. Photomicrographs from transverse sections through thepretectum and mesencephalon after injections of biotinylated dextran-amines into the pallium. A: Pretectum and transitional region be-tween the di- and mesencephalon; the level of the section is approxi-mately halfway between the sections shown in Figure 2G and H.Bilaterally, there is a heavily labeled discrete terminal field (arrows)that extends from the mid-diencephalon into the mesencephalon (seeFig. 2F-H). The arrowheads indicate a more diffuse, dorsal field ofterminals ipsilateral to the injection site, which also extends from thediencephalic into the mesencephalic roof (see Fig. 2F-H). Some labeledcells are visible in the medial (TPm) and in the lateral (TPl) nuclei ofthe posterior tubercular area, mainly ipsilateral to the injection site(see C for a higher power photomicrograph). The dark appearance ofthe ependyma around the mesencephalic ventricle and the darkstaining of the large neurons in the nucleus of the medial longitudinalfascicle is due to the neutral red counterstain (see B and C). B: Centralregion of the mesencephalon, at the approximate level of the trans-verse section shown in Figure 2H. The cell-dense subcommissuralorgan (sco) surrounding the circular mesencephalic ventricle appearsdark due to the neutral red counterstain. Some terminal-like punctaare visible immediately dorsal to the sco (arrow). An intensely labeledneuron is visible laterally in the central mesencephalic gray.C: Posterior tubercular area, slightly caudal to the level shown in A.Again, the dark appearance of the large neurons of the nucleus of themedial longitudinal fascicle (nflm) is due to the neutral red counter-stain. Several intensely labeled cells are visible in the medial nucleusof the posterior tubercular area (TPm), fewer in the lateral nucleus(TPl). For abbreviations, see list. Scale bars 5 500 µm in A, 100 µm inB, 200 µm in C.
TELENCEPHALIC CONNECTIONS IN HAGFISHES 253
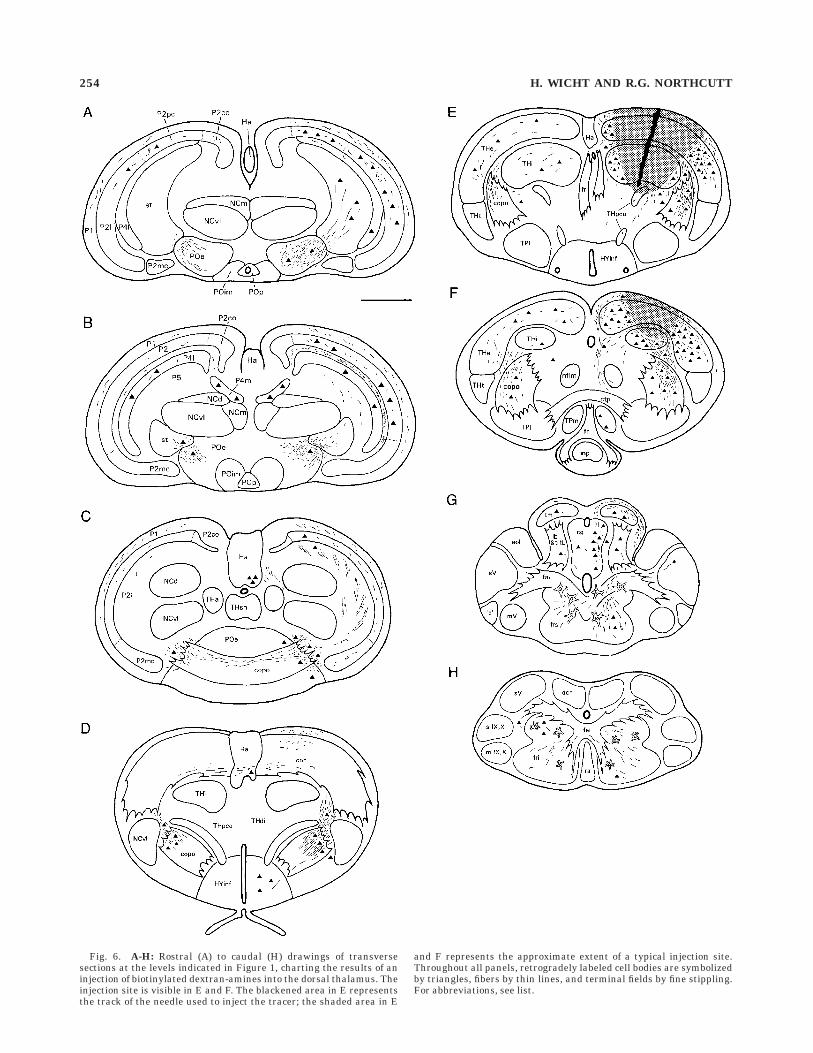
Fig. 6. A-H: Rostral (A) to caudal (H) drawings of transversesections at the levels indicated in Figure 1, charting the results of aninjection of biotinylated dextran-amines into the dorsal thalamus. Theinjection site is visible in E and F. The blackened area in E representsthe track of the needle used to inject the tracer; the shaded area in E
and F represents the approximate extent of a typical injection site.Throughout all panels, retrogradely labeled cell bodies are symbolizedby triangles, fibers by thin lines, and terminal fields by fine stippling.For abbreviations, see list.
254 H. WICHT AND R.G. NORTHCUTT

for an ascending spinal or bulbar lemniscal pathway to thedorsal thalamus.
DISCUSSION
The discussion is divided into two major sections. In thefirst, we discuss the present data in the context of homolo-gies in the brains of myxinoids and other craniates. In thesecond, we compare hagfishes, lampreys, and gnathos-tomes with regard to their pallial organization.
Problems of myxinoid neuroanatomy
Interpretations of myxinoid neuroanatomy have longbeen confounded by controversies regarding the homology
of individual brain areas and nuclei (summary and re-views in Wicht and Northcutt, 1992; Ronan and Northcutt,1998). Some of the questions are finally resolved by thepresent data; in some other cases we can at least rule outsome of the competing hypotheses.
Pallial vs. nonpallial character of the central prosen-
cephalic complex. There is a longstanding debate aboutthe extent of the pallium in myxinoids. Jansen (1930) andothers thought that the pallium was very extensive andthat it included our (Wicht and Northcutt, 1992) centralprosencephalic complex, which was believed by Jansen tobe a homologue of the medial pallium. More recently, anumber of studies (Kusunoki et al., 1981; Wicht andNorthcutt, 1992, 1993, 1994) have shown that the central
Fig. 7. Photomicrographs from transverse sections, counter-stained with neutral red, through various brain regions after injec-tions of biotinylated dextran-amines into the dorsal thalamus. A: Themesencephalic tectum, at the approximate level shown in Figure 6G.Numerous retrogradely labeled cells are visible in the ipsilateraltectum (tm) and in the central gray laterally adjacent to the mesence-phalic ventricle (vmes). B: The mesencephalic tegmentum and thesuperior reticular formation at a level slightly more caudal than thatshown in A. Again, numerous labeled cells can be seen in the centralgray (cg); many of these cells are interspersed between the (unlabeled)
fiber bundles of the bulbar lemniscus (lb) and the tecto-bulbar tract(trtb). A large number of relatively coarse and winding fibers occurs inthe superior reticular formation (frs). C: The octavolateral area (aol)and the sensory nucleus of the trigeminus (sV) at the approximatelevel shown in Figure 6G and ipsilateral to the injection site. A singleretrogradely labeled neuron and its axon (arrows) can be seen in themedial part of the sensory nucleus of the trigeminus. Dorsal andlateral directions are indicated by d and l, respectively. For abbrevia-tions, see list. All scale bars 5 200 µm.
TELENCEPHALIC CONNECTIONS IN HAGFISHES 255

prosencephalic complex is not a homogeneous nucleus; itconsists of at least three nuclei (medial, ventrolateral, anddorsal; see Figs. 2B or 6B) that differ with respect to theircytoarchitecture, and chemoarchitecture, and connectiv-ity.
As regards the medial and ventrolateral nuclei of thecentral prosencephalic complex, Jansen’s (1930) hypoth-esis can be rejected based on present and previous data.The medial pallium is the main pallial target of ascendingthalamic projections in many gnathostomes (Northcutt,1995) and in lampreys (Polenova and Vesselkin, 1993;Northcutt and Wicht, 1997), but none of the nuclei of thecentral prosencephalic complex receives any ascendingthalamic fibers in hagfishes. Amemiya and Northcutt(1996), who injected tracers directly into the medial andventrolateral subnuclei of that complex, also found noconnections with the dorsal thalamic nuclei. Similarly, thehistochemical, topographical, and embryological data(Wicht and Northcutt, 1992, 1993, 1994; Wicht and Tusch,1998) all deny the pallial identity of those nuclei, and thehypothetical homology with the medial pallium must berejected.
There are various other hypotheses regarding the na-ture of these nuclei, ranging from a homology with thedorsal thalamus (Crosby and Schnitzlein, 1974) to ahomology with the striatum (Wachtler, 1974, 1975, 1983)or even the thalamic eminence (Kuhlenbeck, 1977). Obvi-ously, the present data rule out the first of these hypoth-eses completely, as there are no projections ascending fromthe medial and ventrolateral nuclei of the central prosence-phalic complex to the pallium. Similarly, the absence ofany projection from the thalamus to the central prosence-phalic complex is not compatible with an interpretation ofthat complex as the striatum, since the striatum is a majortarget of such projections in gnathostomes (Northcutt,1995). A homology with the thalamic eminence (which waspostulated on purely topological grounds by Kuhlenbeck in1977) can currently not be ruled out completely. Some ofthe known connections of the ventrolateral and medialsubnuclei of the central prosencephalic complex (recipro-cal connections with the preoptic area, reciprocal connec-tions with the habenula; see Amemiya and Northcutt,1996) match those of the thalamic eminence of axolotls, asdescribed by Krug et al. (1993), who performed the onlyexperimental study on the connections of that area in avertebrate. On the other hand, the sheer size of the centralprosencephalic complex in hagfishes speaks against ahomology with the thalamic eminence. In gnathostomesand lampreys, the thalamic eminence is sometimes con-spicuous but is always a relatively small structure at thejunction of the telencephalon and the diencephalon(Holmgren, 1922; Heier, 1948; Herrick, 1948; Smeetset al., 1983). Thus, even if the two structures werehomologous, the relatively large size of the central prosen-cephalic complex of hagfishes must be rated as an apomor-phy.
The dorsal nucleus of the central prosencephalic com-plex poses a special problem. In contrast to the medial andventrolateral subnuclei, it does receive secondary olfactoryinput (Wicht and Northcutt, 1993), and it has reciprocalconnections with the pallium (present study), criteria
compatible with (but not sufficient for) interpreting thedorsal nucleus as the medial pallium (Wicht and North-cutt, 1993), an identity also supported by some of itschemoarchitectural features (Wicht and Northcutt, 1994).On the other hand, the present study has shown that thisnucleus lacks a number of connectional features that aretypical of the medial pallium, including commissural pal-lial connections and ascending thalamic projection. Thus,our previous suggestion (Wicht and Northcutt, 1993, 1994),that the dorsal part of the central prosencephalic complexmight be homologous to the medial pallium should beviewed with caution.
In summary, none of the hypotheses that homologize thecentral prosencephalic complex or its individual nucleiwith any known structure in the forebrain of other crani-ates is really convincing in light of currently availabledata. The reason for this may be simple: the centralprosencephalic complex may have no homologue in thebrain of any other craniates and may represent a derived,apomorphic feature of hagfishes (see Wicht and Northcutt,1992).
Location of the posterior tubercular area. Jansen(1930) identified our (Wicht and Northcutt, 1992; see Fig.2E,F in present study) triangular thalamic nucleus as thenucleus of the posterior tuberculum, while he regarded ourposterior tubercular nuclei (Fig. 2G in present study) asparts of the interpeduncular nucleus. In practically allgnathostomes, the posterior tubercular area is a source ofan ascending projection to the telencephalon (Northcutt,1995). We have rejected Jansen’s interpretation based ontopographical (Wicht and Northcutt, 1994) and neurochemi-cal (Wicht and Northcutt, 1994) arguments. The currentconnectional data, i.e., that ascending telencephalic projec-tions arise from our posterior tubercular area but not fromthe triangular nucleus, clearly favor our interpretation.
Dorsal vs. ventral thalamus. It is difficult to delin-eate a dorsal thalamus from a ventral thalamus in hag-fishes solely by cytoarchitecture and topography (Wichtand Northcutt, 1992). The current data show that at leastthe external and the internal nuclei of the thalamus can beregarded as dorsal thalamic nuclei, because they harborthe majority of the diencephalic cells that project to thetelencephalon. The status of most of the other thalamicnuclei of the diencephalon in myxinoids, however, remainsuncertain.
Multiple pallial fields in hagfishes. In previous stud-ies (Wicht and Northcutt, 1992, 1994) we identified severalsubfields in the laminated pallium of hagfishes based oncytoarchitectural and immunohistochemical criteria. Sev-eral of these fields also have unique connectional proper-ties. For example, the compact part of pallial layer 2 andthe medial part of pallial layer 4 are the main source ofcommissural pallial connections; the parvicellular andlateral parts of pallial layer 2 and the lateral part of palliallayer 3 contain the majority of the pallial projectionneurons. On the other hand, none of the many pallial fieldsis a ‘‘preferred’’ target for either secondary olfactory (Wichtand Northcutt, 1992) or ascending thalamic afferents(present study), with the possible exception of the magno-cellular part of pallial layer 2, which is pierced by the deeppart of the lateral olfactory tract and lies in close vicinity toa heavy terminal field formed by that tract (Wicht and
256 H. WICHT AND R.G. NORTHCUTT

Northcutt, 1993; arrows in Fig. 2C of present study). It isalso reached, however, by a sparse number of ascendingthalamic fibers (Fig. 6C).
In our opinion, the available data on pallial organizationin hagfishes do not allow a direct comparison between themedial/dorsal/lateral pallial fields of gnathostomes andthe diverse pallial fields in hagfishes. It is, of course,possible to subdivide the pallium of myxinoids into medial,dorsal, and lateral fields based solely on topographicalcriteria and to declare those regions homologous to therespective fields of gnathostomes (Crosby and Schnitzlein,1974), but this attempt denies and ignores the manyunique characters of the pallium in myxinoids that webelieve preclude a direct homology with the pallial fields ofgnathostomes: the lamination of the pallium (Jansen,1930; Wicht and Northcutt, 1992, 1994); the presence of adeep part of the lateral olfactory tract (Wicht and North-cutt, 1993); the absence of a particular and well-definedtarget area for the ascending thalamic projections (presentstudy); and the apparently complete overlap betweenascending thalamic projections. If there is no basis for a‘‘one-to-one’’ comparison between the pallial fields of myxi-noids and those of other craniates, two alternate hypoth-eses must be considered: (1) the pallium of myxinoids, as awhole, is a homologue of all three pallial areas of gnathos-tomes, or (2) the pallium of myxinoids is a homologue ofonly one or two of these areas.
We think that the second hypothesis is more likely valid,because the pallium of myxinoids combines charactersthat are typical of the medial and lateral pallia of gnathos-tomes. There, the medial pallium is the major target ofthalamic projections, it is interconnected with the fornixsystem, and it is the main source of commissural and longdescending pallial projections; the lateral pallium is theprincipal target of the secondary olfactory fibers (North-cutt, 1995). A dorsal pallium that receives no or only sparsesecondary olfactory projections as in gnathostomes, cannotbe recognized in myxinoids.
In summary, the pallium of hagfishes has multiplesubdivisions with differential connections which probablyreflect different functional roles. None of these subdivi-sions, however, appears to have a direct homologue inother craniates.
Hagfishes, lampreys, and gnathostomes, andthe evolution of telencephalic connections
The most widely accepted hypothesis (Løvtrup, 1977;Janvier, 1981; see Janvier, 1996; Wicht and Nieuwenhuys,1997 for reviews) regarding the taxonomic relationshipsamong hagfishes, lampreys, and gnathostomes is illus-trated in Figure 8. Using this cladogram and the cladisticmethodology for determining plesiomorphic vs. apomor-phic character states (i.e., phylogentically primitive vs.derived states; see Northcutt, 1995, for a detailed descrip-tion of the method and its limitations), a morphotype of thepallium in craniates, its divisions and connections can bereconstructed. The data for gnathostomes have recentlybeen reviewed and a morphotype of their forebrain hasbeen reconstructed (Northcutt, 1995). We will thus usethis gnathostome morphotype and compare it to the condi-tions observed in lampreys and hagfishes in order to avoid
lengthy repetitions and discussions of gnathostomian char-acter sets.
Unfortunately, the morphotype of the diencephalon andtelencephalon of craniates cannot be reconstructed basedon an out-group comparison with cephalochordates, thepresumed sister group (Fig. 8) of craniates. Cephalochor-dates do possess a homologue of the diencephalon (Lacalliet al., 1994; Lacalli, 1996 a,b), but it shows few if any of themorphological features (nuclear centers and axonal tracts)typical of the diencephalon in adult craniates (Fritzsch,1996). The pineal organ is a notable exception (Lacalli,1996a). A telencephalon appears to be entirely missing incephalochordates (Lacalli, 1996b; Northcutt, 1996). There-fore, the determination of plesiomorphic character statesof the diencephalon and telencephalon of craniates has torely on comparison of the three extant craniate taxa and onthe cladogram shown in Figure 8. Assuming that this
Fig. 8. A cladogram (in the lower part of the figure) showing thetaxonomic interrelationships of major recent chordate taxa, andintroducing the systematic terms (in the upper part of the figure) forthe major monophyletic groups of chordates. The cladogram is used todetermine the plesiomorphic states of forebrain characters amongcraniates and to reconstruct the morphotypes at different taxonomicallevels (see text). The boxes in the lower part that partially overlap thestem lines of the various monophyletic groups contain the keycharacters of those morphotypes.
TELENCEPHALIC CONNECTIONS IN HAGFISHES 257

cladogram is valid,1 any character that is present in allthree taxa, or in hagfishes and lampreys, or in hagfishesand gnathostomes, is probably plesiomorphic for the entirecraniate clade. Characters shared by lampreys and gnatho-stomes, but not by hagfishes, are plesiomorphic for myopte-rygians, but their polarity at the level of craniates cannotbe established.
Secondary olfactory system. Extensive bilateral sec-ondary olfactory projections to many, if not all, pallial andsubpallial areas are a shared character of lampreys (North-cutt and Puzdrowski, 1988) and hagfishes (Wicht andNorthcutt, 1993) and therefore must be regarded as aplesiomorphic character of craniates. The large pallialfields that are devoid of such input or receive only sparseolfactory projections (Northcutt, 1995) must be regardedas an evolutionary novelty that arose in the stem line ofgnathostomes.
Thalamopallial projections and ascending sensory
pathways. Thalamopallial projections occur in all threegroups of craniates (Polenova and Vesselkin, 1993; North-cutt and Wicht, 1997; present study) and are a plesiomor-phic craniate feature.
The organization of the ascending spinal and bulbarsensory pathways to the thalamus, however, differs be-tween lampreys and hagfishes on one hand and many (butnot all) gnathostomes on the other. In hagfishes (Ronanand Northcutt, 1990; present study) and lampreys (Ronanand Northcutt, 1990; Polenova and Vesselkin, 1993) nei-ther the spinal nor the bulbar lemnisci ascend to thethalamus. Among gnathostomes, spinal lemniscal path-ways to the thalamus have been found in all tetrapods; butonly in two nontetrapod species (see Munoz et al., 1997, fora recent overview). The absence of a spinothalamic lemnis-cal system is therefore a plesiomorphic feature of crani-ates, and this character probably evolved independently insome nontetrapods and in the stem line of tetrapods(Ronan and Northcutt, 1990; Munoz et al., 1994, 1997).
In hagfishes, the tectum is a major target of the ascend-ing bulbar (Amemyia, 1983; Ronan, 1988) and spinal(Ronan and Northcutt, 1990) lemnisci. In lampreys (Ronan
and Northcutt, 1990), the spinal lemniscus does not ap-pear to ascend to the tectum but terminates in themesencephalic tegmentum. Similar to the situation inhagfishes, however, the tectum of lampreys receives ascend-ing projections from the sensory and the reticular nuclei ofthe rhombencephalon via the bulbar lemniscus (M. Ronanand R.G. Northcutt, unpublished observations). The tec-tum and tegmentum, in turn, are interconnected with thedorsal thalamus in both groups (Polenova and Vesselkin,1993; present study); thus, ascending spinal and bulbarafferents to the pallium may use a tectal/tegmental-thalamotelencephalic route as their final pathway.
In most gnathostomes (Wicht and Himstedt, 1986, 1988;Northcutt, 1995) two visual pathways to the telencephaloncan be distinguished. A retinothalamotelencephalic path-way typically relays in the anterior thalamic nucleuswhich, in turn, projects to many targets in the telencepha-lon, including pallial fields. A similar situation is found inlampreys, where the retinal fibers project to a lateral cellplate in the dorsal thalamus (‘‘the corpus geniculatumlaterale’’) which displays ascending projections to themedial pallium (Polenova and Vesselkin, 1993; Northcuttand Wicht, 1997). Hagfishes also have an anterior retinore-cipient (Wicht and Northcutt, 1990) nucleus in theirrostral diencephalon, which also gives rise to an ascendingpallial projection, even though it is very weak (presentstudy), perhaps due to the reduced state (Fernholm andHolmberg, 1975) of their eyes and visual system. At anyrate, a retinothalamotelencephalic visual system appearsto be a primitive craniate character. The same is true forthe second ascending visual pathway, which reaches thetelencephalon via a retinotectothalamo-telencephalic routein gnathostomes (Northcutt, 1995). A retinotectal projec-tion is present in lampreys (Kennedy and Rubinson, 1977;Kosareva, 1980; Vesselkin et al., 1980) and hagfishes(Kusunoki and Amemiya, 1993; Wicht and Northcutt,1990), and the tectum projects to the dorsal thalamus inboth groups (see above).
Projections from the posterior tubercular area to the
pallium. Neurons in the posterior tubercular area projectto the telencephalon in all gnathostomes investigated todate. Their major target area is the striatum (Northcutt,1995), but projections to the pallium have also beenreported in sharks (Squalus acanthias, Smeets and North-cutt, 1987). Notably, no tuberculopallial projections havebeen observed in lampreys (Polenova and Vesselkin, 1993;Northcutt and Wicht, 1997), but it is possible that lam-preys possess a tuberculostriatal projection. The presenceof a tuberculopallial connection in hagfishes (presentstudy) indicates that this character is a plesiomorphiccraniate feature that was lost in lampreys.
Connections with the preoptic and infundibular ar-
eas. In both lampreys (Polenova and Vesselkin, 1993;Northcutt and Wicht, 1997) and hagfishes (present study),there are substantial reciprocal connections between thepreoptic area and the pallium. In lampreys, these connec-tions extend caudally into the infundibular area of thehypothalamus. Similar connections with the medial pal-lium have been reported in all gnathostomes investigatedto date (Northcutt, 1995); thus, a palliopreoptic projectionsystem (which probably corresponds to the mammalianpostcommissural fornix) is an ancient trait of the prosen-cephalon of craniates.
1This cladogram has indeed been challenged based on molecular andmorphological data. Yalden (1985) and, more recently, Mallat (1996) havebrought forward a number of arguments in favor of a presumed monophylyof hagfishes and lampreys (thus resurrecting the old taxon ‘‘Agnatha/Cyclostomata’’). They report a large number of similarities in the feedingapparatus and in the organization of the premandibular region of both taxa.These similarities are indeed striking, but because it could not be estab-lished whether they represent symplesiomorphies or synapomorphies, theyare of no relevance for the identification of monophyletic groups among thethree craniate taxa. Molecular evidence in favor of a presumed monophylyof hagfishes and lampreys stems from a comparative analysis of the 18SrRNA in hagfishes, lampreys, and a number of gnathostomes, with cephalo-chordates and tunicates as out-groups (Stock and Whitt, 1992). However,the study of Stock and Whitt has also shown that the 18S rRNA ofmyxinoids shows a rate of base-substitutions that is twice as high as that ofany other chordate, and it may thus be questioned whether this molecule isa particularly useful tool for the study of myxinoid relationships. Morerecently, Suzuki et al. (1995) performed a similar analysis by using thesequence of the vasotocin cDNA of lampreys, hagfishes, and variousgnathostomes. Their analysis grouped lampreys together with gnathos-tomes, thus supporting the cladogram used in the present study. It must bementioned, however, that Suzuki et al. (1995) used the cDNA sequence of amolluscan (the pond snail, Lymnaea stagnalis) vasotocin-homologue to roottheir cladogram. Because molluscs certainly do not form the sister group ofcraniates, and because data on the vasotocin-homologues of non-chordatedeuterostomes are currently not available, these data should also be viewedwith caution.
258 H. WICHT AND R.G. NORTHCUTT

Efferent pallial projections. In hagfishes, there aredistinct descending pallial projections to the roof of thediencephalon, the pretectum, and the tectum. The projec-tions to the tegmentum are weak, and we did not observeany palliofugal fibers caudal to the mesencephalic-rhombencephalic boundary. A very similar pattern is seenin lampreys after injections of tracers into the pallium(Polenova and Vesselkin, 1993; Northcutt and Wicht,1997), even though the projection to the mesencephalictegmentum appears to be somewhat more intense than inhagfishes. Pallial efferents are restricted to the telence-phalic hemisphere and the preoptic region in most gnatho-stomes. Long descending projections to the di- and mesen-cephalic roof occur in anuran amphibians (Neary, 1990;Northcutt and Ronan, 1992) and long descending projec-tions to the motor centers of the medulla oblongata andspinal cord are characteristic of mammals. The presence ofpalliothalamic and palliotectal fibers in hagfishes andlampreys indicates that this is the plesiomorphic characterstate of pallial efferents among craniates.
Pallial fields. Three (medial, dorsal, and lateral) or,possibly, four pallial fields were present in the last commonancestor of gnathostomes (Northcutt, 1995). In lampreys,three pallial fields can be identified based on topographi-cal, cytoarchitectural, and connectional criteria (Northcuttand Puzdrowski, 1988; Polenova and Vesselkin, 1993;Northcutt and Wicht, 1997): (3) a large lateral field whichis the main recipient of the secondary olfactory projectionand is probably homologous to the lateral pallium ofgnathostomes, (2) a smaller medial field which receivessomewhat fewer secondary olfactory fibers but is the maintarget of thalamic projections and the main source ofdescending pallial fibers and probably homologous to themedial pallium of gnathostomes, and (3) a third pallialsubdivision in the form of the so-called ‘‘subhippocampallobe’’ (Heier, 1948), an area that might be homologous tothe dorsal pallium of gnathostomes (Northcutt and Puzd-rowski, 1988). The ventral part of that lobe is traversedand innervated by secondary olfactory fibers, but thedorsal part, which receives some thalamic afferents, ap-pears to receive no secondary olfactory projections (Pole-nova and Vesselkin, 1993). Thus, lampreys and gnathos-tomes share a pallial bauplan at least grossly similar.
As noted above, it is not possible to compare the multiplepallial fields of hagfishes directly to those of either gnatho-stomes or lampreys, and it is therefore not possible todetermine whether the myopterygian (lampreys and gna-thostomes) or the myxinoid pattern of pallial organizationis primitive for craniates. Unfortunately, with only lam-preys and hagfishes as the sole survivors of a once flourish-ing radiation of jawless craniates, we will never be able todetermine with certainty whether a third, yet unknownpattern of pallial organization existed in the commonancestor of hagfishes, lampreys, and gnathostomes, orwhether the condition seen in hagfishes or that seen inlampreys is truly primitive for our clade, the craniates.
ACKNOWLEDGMENTS
Ron McConnaughey caught the hagfishes used in thisstudy and also helped to maintain the animals: Manythanks, Ron, also for sharing our enthusiasm for slimyeels. As usual, Mary Sue Northcutt assisted throughout allphases of the research and the preparation of the manu-
script and her help is greatly appreciated. Barbara Ruppprovided moral, editorial, graphical, and photographicalsupport to H.W. This study was supported by NIH grantsto R.G.N. (NS24699/DC01081) and by travel grants fromthe Dr. Senckenbergische Stiftung (Frankfurt/Main) toH.W.
LITERATURE CITED
Adam, H. (1963) Brain ventricles, ependyma, and related structures. In A.Brodal and R. Fange (eds): The Biology of Myxine. Oslo: Universitetsfor-laget, pp. 137-149.
Adams, J.C. (1981) Heavy metal intensification of DAB-based HRP-reaction product. J. Histochem. Cytochem. 29:775.
Amemiya, F. (1983) Afferent connections of the tectum mesencephali in thehagfish Eptatretus burgeri: An HRP-study. J. Hirnforsch. 24:225-236.
Amemiya, F. and R.G. Northcutt (1996) Afferent and efferent connections ofthe central prosencephalic nucleus in the Pacific hagfish. Brain Behav.Evol. 47:149-155.
Crosby, E.C. and H.N. Schnitzlein (1974) The comparative anatomy of thetelencephalon of the hagfish, Myxine glutinosa. J. Hirnforsch. 15:211-236.
Fernholm, B. and K. Holmberg (1975) The eyes in three genera of hagfish(Eptatretus, Paramyxine, and Myxine): A case of degenerative evolution.Vision Res. 15:253-259.
Fritzsch, B. (1996) Similarities and differences in lancelet and craniatenervous systems. Israel J. Zool. 42 (Suppl.):147-160.
Godement, P., J. Vanselow, S. Thanos, and F. Bonhoeffer (1987) A study indeveloping visual systems with a new method of staining neurones andtheir processes in fixed tissue. Development 101: 697-713.
Heier, P. (1948) Fundamental principles in the structure of the brain. Astudy of the brain of Petromyzon fluviatilis. Acta anatomica 5 (Suppl.VIII):1-213.
Herrick, C.J. (1948) The Brain of the Tiger Salamander. Chicago: TheUniversity of Chicago Press.
Holmgren, N. (1922) Points of view concerning forebrain morphology inlower vertebrates. J. Comp. Neurol. 34:391-495.
Jansen, J. (1930) The brain of Myxine glutinosa. J. Comp. Neurol. 49:359-507.
Janvier, P. (1981) The phylogeny of the craniata, with particular referenceto fossil ‘agnathans’. J. Vert. Paleontol. 1:121-159.
Janvier, P. (1996) Early Vertebrates. Oxford: Claredon Press.Kennedy, M.C. and K. Rubinson (1977) Retinal projections in larval,
transforming, and adult sea lamprey, Petromyzon marinus. J. Comp.Neurol. 171:465-480.
Kosareva, A.A. (1980) Retinal projections in lamprey (Lampetra fluviatilis).J. Hirnforsch. 21:243-225.
Krug L., H. Wicht, and R.G. Northcutt (1993) Connections of the thalamiceminence of the axolotl, Ambystoma mexicanum. Neurosci. Lett. 149:145-148.
Kuhlenbeck, H. (1977) The Central Nervous System of Vertebrates. Vol. 5/I.Derivatives of the Prosencephalon: Diencephalon and Telencephalon.Basel: Karger.
Kusunoki, T., T. Kadota, and R. Kishida (1981) Chemoarchitectonics of theforebrain of the hagfish, Eptatretus burgeri. J. Hirnforsch. 22:285-298.
Kusunoki, T. and F. Amemiya (1983) Retinal projections in the hagfish,Eptatretus burgeri. Brain Res. 262:295-298.
Lacalli T.C. (1996a) Frontal eye circuitry, rostral sensory pathways andbrain organization in amphioxus larvae: Evidence from 3D reconstruc-tions. Phil. Trans. Roy. Soc. London B 351:243-263.
Lacalli, T.C. (1996b) Landmarks and subdomains in the larval brain ofBranchiostoma: Vertebrate homologs and invertebrate antecendents.Israel J. Zool. 42 (Suppl.):131-146.
Lacalli, T.C., N.D. Holland, and J.E. West (1994) Landmarks in the anteriornervous system of amphioxus larvae. Phil. Trans. Roy. Soc. London B344:165-185.
Løvtrup, S. (1977) The Phylogeny of Vertebrata. London: Wiley.Mallat, J. (1996) Ventilation and the origin of jawed vertebrates: A new
mouth. Zool. J. Linn. Soc. 117:329-404.Munoz, A., M. Munoz, A. Gonzalez, and H.J. ten Donkelaar (1994)
Spinothalamic projections in amphibians as revealed with anterogradetracing techniques. Neurosci. Lett. 171:81-84.
TELENCEPHALIC CONNECTIONS IN HAGFISHES 259

Munoz, A., M. Munoz, A. Gonzalez, and H.J. ten Donkelaar (1997) Spinalascending pathways in amphibians: Cells of origin and main targets. J.Comp. Neurol. 378:205-228.
Neary, T. (1990) The pallium of anuran amphibians. In A.Peters andE.Jones (eds): Cerebral Cortex, Vol. 8A, New York: Plenum Press, pp.107-138.
Northcutt, R.G. (1995) The forebrain of gnathostomes: In search of amorphotype. Brain Behav. Evol. 46:275-318.
Northcutt, R.G. (1996) The origin of craniates: Neural crest, neurogenicplacodes, and homeobox genes. Israel. J. Zool. 42 (Suppl.):273-313.
Northcutt, R.G. and M.C. Ronan (1992) Afferent and efferent connections ofthe bullfrog medial pallium. Brain Behav. Evol. 40: 1-16.
Northcutt, R.G. and R. Puzdrowski (1988) Projections of the olfactory bulband the nervus terminalis in the Silver lamprey. Brain Behav. Evol.32:96-107.
Northcutt, R.G. and H. Wicht (1997) Afferent and efferent connections ofthe lateral and medial pallia of the Silver lamprey. Brain Behav. Evol.49:1-19.
Polenova, O.A. and N.P. Vesselkin (1993) Olfactory and non-olfactoryprojections in the River lamprey (Lampetra fluviatilis) telencephalon. J.Hirnforsch. 34:261-279.
Quinn, B. and E. Weber (1988) m-Phenylenediamine: A novel fluorescentNissl-like stain for neuroanatomy. Soc. Neurosci. Abstr. 14:547.
Ronan, M. (1988) The sensory trigeminal tract of Pacific hagfish. Primaryafferent projections and neurons of the tract nucleus. Brain Behav.Evol. 32:169-180.
Ronan, M. (1989) Origin of descending spinal projections in petromyzontidand myxinoid agnathans. J. Comp. Neurol. 281:54-68.
Ronan, M. and R.G. Northcutt (1990) Projections ascending from the spinalcord to the brain in petromyzontid and myxinoid agnathans. J. Comp.Neurol. 291:491-508.
Ronan, M. and R.G. Northcutt (1998) The central nervous system ofhagfishes. In J.M Jørgensen, J.P. Lomholt, R.E. Weber, and H. Malte(eds): The Biology of Hagfishes. London: Chapman and Hall, pp.451-479.
Smeets, W.J.A.J. and R.G. Northcutt (1987) At least one thalamotelence-phalic pathway in cartilaginous fishes projects to the medial pallium.Neurosci. Lett. 78:277-282.
Smeets, W.J.A.J., R. Nieuwenhuys, and B.L. Roberts (1983) The CentralNervous System of Cartilaginous Fishes, Berlin: Springer.
Stock, D.W. and G.S. Whitt (1992) Evidence from 18S ribosomal RNAsequences that lampreys and hagfishes form a natural group. Science257:787-789.
Suzuki, M., K. Kubokawa, H. Hagasawa, and A. Urano (1995) Sequenceanalysis of vasotocin cDNAs of the lamprey, Lampetra japonica, and thehagfish, Eptatretus burgeri: Evolution of cyclostome vasotocin precur-sors. J. Molec. Endocrinol. 14:67-77.
Veenman, C.L., A. Reiner, and M.G. Honig (1992) Biotinylated dextranamine as an anterograde tracer for single- and double-labelling studies.J. Neurosci. Meth. 41:239-254.
Vesselkin, N.P., T.V. Ermakova, J. Reperant, A.A. Kosareva, and N.B.Kenigfest, (1980) The retinofugal and retinopetal systems in Lampetrafluviatilis. An experimental study using radioautographic and HRPmethods. Brain Res. 195:453-460.
Wachtler, K. (1974) The distribution of acetylcholinesterase in the cyclo-stome brain. I. Lampetra planeri. Cell Tissue Res. 152: 259-270.
Wachtler, K. (1975) The distribution of acetylcholinesterase in the cyclo-stome brain. II. Myxine glutinosa. Cell Tissue Res. 159: 109-120.
Wachtler, K. (1983) The acetylcholine-system in the brain of cyclostomeswith special reference to the telencephalon. J. Hirnforsch. 24:63-70.
Wicht, H. and W. Himstedt (1986) Two thalamo-telencephalic pathways in aurodele, Triturus alpestris. Neurosci. Lett. 68: 90-94.
Wicht, H. and W. Himstedt (1988) Topologic and connectional analysis ofthe dorsal thalamus of Triturus alpestris (Amphibia, Urodela, Salaman-dridae). J. Comp. Neurol. 267:545-561.
Wicht, H. and R.G. Northcutt (1990) Retinofugal and retinopetal projec-tions in the Pacific hagfish, Eptatretus stouti. Brain Behav. Evol.36:315-328.
Wicht, H. and R.G. Northcutt (1992) The forebrain of the Pacific hagfish: Acladistic reconstruction of the ancestral craniate forebrain. BrainBehav. Evol. 40:25-64.
Wicht, H. and R.G. Northcutt (1993) Secondary olfactory projections andpallial topography in the Pacific hagfish, Eptatretus stouti. J. Comp.Neurol. 337:529-542.
Wicht, H. and R.G. Northcutt (1994) An immunohistochemical study of thetelencephalon and the diencephalon in a myxinoid jawless fish, thePacific hagfish, Eptatretus stouti. Brain Behav. Evol. 43:140-161.
Wicht, H. and U. Tusch (1998) Ontogeny of the head and nervous system ofmyxinoids. In J.M. Jørgensen, J.P. Lomholt, R.E. Weber, and H. Malte(eds): The Biology of Hagfishes. London: Chapman and Hall, pp.429-448.
Wicht, H. and R. Nieuwenhuys (1997) Hagfishes (Myxinoidea) In R.Nieuwenhuys, H.J. ten Donkelaar, C. Nicholson (eds): The CentralNervous System of Vertebrates, Vol. 1. Heidelberg: Springer, pp.497-549.
Yalden, D.W. (1985) Feeding mechanisms as evidence for cyclostomemonophyly. Zool. J. Linn. Soc. 84:291-300.
260 H. WICHT AND R.G. NORTHCUTT