task decision difficulty: effects on erps in a same-different letter classification task
TRANSCRIPT

ELSEVIER
BIOLOGICAL PSYCHOLOGY
Biological Psychology 38 (1994) 199-214
Task decision difficulty: effects on ERPs in a same-different letter classification task
Barbara Palmer a,*, Victoria Tepe Nasman a,*, Glenn F. Wilson b
a Logicon Technical Services, Inc., P.O. Box 317258, Dayton, OH, 45431-7258, USA b Wright-Patterson Air Force Base, OH, USA
Abstract
A “same-different” letter pair choice reaction time task was used to compare reaction time, performance accuracy, and P3 amplitude and latency at three levels of task difficulty. Stimulus set was held constant across tasks, and task difficulty was manipulated by instructions to the subject. Subjects delivered a button-press response to designate whether members of each letter pair matched or mismatched on the basis of their physical identity (low difficulty), name identity (medium difficulty), or category identity (vowels/ consonants, high difficulty). Task decision difficulty was confirmed by slower reaction times and reduced response accuracy. P3 amplitude was inversely related to task decision difficulty; relatively larger P3s were associated with matched (vs. mismatched) letter pairs. These findings evidence direct effects of task difficulty on P3 amplitude, possibly as the result of “equivoc- ation” related to more difficult task judgments.
Keywords: P300; Task difficulty
1. Introduction
The P300 (or P3) component is held to be a valid index of central information processing during task-related decision-making. Changes in P3 may help to de- scribe the effects of increasingly difficult stimulus discrimination and categoriza- tion judgments. To the extent that P3 is sensitive to the difficulty of cognitive tasks, its variation may provide a useful indicant of cognitive workload. In general, more difficult discriminations tend to produce smaller and later P3s (Hillyard & Kutas,
* Authorship order is random; B.P. and V.T.N. contributed equally.
0301-0511/94/$07.00 0 1994 Elsevier Science B.V. All rights reserved SSDZ 0301-0511(94)00953-U

200 B. Palmer et al. /Biological Psychology 38 (1994) 199-214
1983; Fitzgerald & Picton, 1983, 1984; Gevins & Cutillo, 1986; Taylor, 1988). Larger memory loads and more difficult mental rotation tasks are also associated with smaller P3s, longer P3 latencies, and slower reaction times (Gomer, Spicuzza & O’Donnell, 1976; Adams & Collins, 1978; Friedman, Vaughan & Erlenmeyer- Kimling, 1981; Sergeant, Geuze & van Winsum, 1987; Wilson, Swain & Davis, 1988; Mecklinger, Kramer & Strayer, 1992).
One possible explanation for such findings is that more difficult task decisions are likely to be associated with reduced levels of confidence or certainty. Decision confidence has been shown to influence P3 amplitude (Hillyard, Squires, Bauer & Lindsay, 1971; Squires, Squires & Hillyard, 1975; Ruchkin, Sutton, Kietzman & Silver, 1980; Sutton, Ruchkin, Munson, Kietzman & Hammer, 1982). Similarly, Johnson (1986) observes that P3 amplitude will tend to decrease with higher levels of post-decision uncertainty (“equivocation,” see Ruchkin & Sutton, 1978; John- son, 1986). In general, decisions made with a high degree of confidence will tend to evoke the largest P3s. Considered within the framework of Johnson’s triarchic model, effects of decision confidence may offer the most parsimonious explanation for reported effects of task difficulty on P3. This view would predict that P3 could serve as a reliable inverse index of task decision-making difficulty, providing that measures are taken to control against other variables which are known to exert a direct or indirect influence on the behavior of P3.
However, a variety of possible confounds present in earlier studies makes it difficult to conclude that changes in P3 are due specifically to effects of task difficulty, rather than to interactions with or among other variables. The most common paradigm for the study of P3 is the oddball paradigm, in which subjects are presented with a series of differentially probable stimulus types. The effects of event probability may interact with and tend to obscure task difficulty effects on P3 (see Polich, 1987). Variable stimulus sets may also be problematic because they introduce the possibility that certain stimuli are more meaningful than others, or that they may impose unique information processing demands. Finally, previous studies of task difficulty have generally employed tasks that require memory scanning or comparison operations. It is not known whether memory requirements exert common or unique effects on easy and difficult task-related judgments, but in either case they may tend to introduce confounding effects on encoding and classification processes (see Thatcher, 1977).
The purpose of the present study was to eliminate the potential confounds of earlier studies, and to observe directly the effects of task difficulty on P3. Task difficulty was manipulated directly by changing the task instructional set. Stimulus set was held constant, delivered in the context of an equiprobable “same-differ- ent” (match/mismatch) letter pair task (see Posner & Mitchell, 1967). By present- ing simultaneously in each trial the characters to be compared, this task eliminates memory requirements that are inherent to tasks which employ serial stimulus presentations.
The “same-different” letter pair task involves the presentation of a series of letter pairs, to which subjects respond by designating whether the members of each pair possess the same or different physical, nominal, or categorical (vowel vs.

B. Palmer et al. /Biological Psychology 38 (1994) 199-214 201
consonant) characteristics. Variations of this task have been used by other investi- gators to study the character and chronometry of stimulus identification, encoding and categorization processes (e.g., see Pachella & Miller, 1976). Target identifica- tion and detection studies that use such tasks have demonstrated that category judgments are relatively less automatic and occur more slowly than those based on physical or nominal stimulus features (Taylor, 1978; Prinz, 1980; Krueger, 1984). Despite the fact that the “same-different” task is particularly well-suited as a test of task difficulty effects on P3, we are aware of only one other study that has used such a task for this purpose. Poon, Thompson and Marsh (1976) compared P3s evoked by a simple reaction time task and a vowel/consonant category matching task, and found that category information processing produced larger late positivi- ties. However, simple reaction time tasks provide a very weak basis of comparison in studies of decision-making difficulty because they do not require subjects to discriminate within or among stimulus trials. As a result, simple reaction time tasks often produce no discernible endogenous response. (This was the case in the present study, where ERPs to simple reaction time tasks contained no clearly identifiable P3s.)
Here, the “same-different” task is used to compare ERPs and performance measures in three choice reaction time tasks, each of which required stimulus comparison, discrimination, and task-related decision-making. The objective of this study was to determine whether P3 is directly sensitive to the difficulty of task judgments, and to consider our findings within the theoretical framework by which P3 is presently studied and understood.
2. Methods
2.1. Subjects
Twelve (12) right-handed male college students served as subjects in the experiment. Subjects were paid in return for their participation. Each subject was tested in every condition.
2.2. Recording and averaging
The electroencephalogram (EEG) was monitored at 20 sites along the scalp using a lo/20 System (Jasper, 1958) electrode cap (Electra-Cap International, Inc.) referenced to linked mastoids. An electrode at Fpz served as ground. Electrode impedances were kept below 5 k0.
Data acquisition was managed with a Bio-Logic Brain Atlas (Bio-Logic, Inc.). Signals were amplified by 30 000 with 3 dB filter cutoffs set to pass signals between 0.1 and 30 Hz. Amplified signals were digitized on-line at a rate of 250 samples s-i. Trials containing amplitude values that exceeded +82 PV (98% A/D gain range) were considered artifact and were rejected on-line. ERPs were derived by averaging all trials on-line across a 1024 ms epoch, which included a 125 ms

202 B. Palmer et al. /Biological Psychology 38 (1994) 199-214
pre-stimulus baseline. Each ERP contained 15-25 artifact-free trials. Due to hardware limitations, it was not possible to eliminate error trials from the averaged ERPs.
2.3. Stimulus presentation
A series of letter pairs was presented in each condition. Each series contained letter pairs drawn from the same set of four letters (A, B, C, E, upper or lower case). Inherent differences between the three experimental tasks made it impossi- ble to match the three stimulus series on all possible factors (e.g. letter case), and match/mismatch trials were equated only along a single dimension for each task level. Because the three tasks differed in terms of their respective a priori matched and mismatched letter pair configuration probabilities, presentation probabilities were held constant for matched (50%) and mismatched (50%) letter pairs across trials within each task level and block. In the PHYSICAL and NAME conditions, matched pairs comprised 13% and 25% of all possible letter combinations, respectively; matched and mismatched pairs held equal a priori probabilities only in the CATEGORY task. Thus, fewer possible matched (vs. mismatched) letter pair configurations in both the PHYSICAL and NAME task conditions required that there be relatively more repetitions of matched pair stimuli in order to achieve equiprobable match/ mismatch presentations. The only condition in which the case of the letters played a defining role was in the PHYSICAL task, where a matched letter pair had to contain two letter characters that were identical in all physical respects. Finally, mismatched letter pairs in every task necessarily involved letters that were physically different, although not necessarily different in case. Example letter pairs presented in Table 1 illustrate that identical letter pairs could require different responses under different task levels.
Letter pair stimuli were presented as two characters, each 0.5 cm wide X 0.7 cm high, subtending a visual angle of less than 2 degrees on a monochrome monitor at
Table 1
“Same-different” task. Example match and mismatch letter pairs at each task difficulty level
Match AA aA ea CC-_-_-_7/-EC -__-,i,,
/ /
/ /
/ /
/ /
Mismatch SA’ .a?%’ m ~~~~~_~~ _____- x

B. Palmer et al. /Biological Psychology 38 f1994) 199-214 203
a viewing distance of 62 cm. Stimulus presentation was managed by a Commodore 64 computer, using a modified version of the Criterion Task Set linguistic process- ing task (Shingledecker, 1984, see also Posner & Mitchell, 1967). The order of stimulus presentation was randomly determined in each condition. Each letter pair stimulus remained on the viewing monitor until response issuance, limited by a maximum trial duration of 1.5 s. Letter pairs were presented at a 2.1 s inter-stimu- lus (onset-to-onset) interval; the inter-trial (offset-to-onset) interval varied with response issuance. Matched and mismatched pairs were equiprobable across presentations; 50 trials (25 matched and 25 mismatched letter pair presentations) were delivered in each condition. Separate ERPs were cdlected to match and mismatch trials.
2.4. Procedure
Subjects were instrumented for EEG, ECG and respiration monitoring (ECG and respiration data will be presented in a separate paper) and seated in a comfortabIe recliner facing the stimulus monitor. Subjects were fully instructed on the experimental protocol, and were asked to sit as still as possible and to minimize blinking and eye movement. Subject compliance and cooperation were pre-tested using a simple reaction time button-press task to 50 trials of word stimuli; during pre-testing, the experimenter monitored on-line EEG and provided feedback to the subjects about muscular and ocular artifacts.
During testing, subjects issued a button-press “yes” or “no” response after each presentation to indicate whether paired letters matched on the basis of their PHYSICAL (low difficulty), NAME (medium difficulty) or vowel/ consonant CATEGORY identity (high difficulty). Task order was counterbalanced. To exam- ine the possible effect of task practice on reaction time CRT), response accuracy, and ERPs, the task series was repeated in three blocks. Before and after testing, subjects performed a simple reaction time task using letter pair stimuli; this was done to identify any changes in RT due to fatigue over the course of the session.
2.5. Data analysis
Topographic brain maps were generated for baseline-adjusted ERPs. (Topo- graphic landscapes presented herein represent mapped voltage potential at 19 referenced electrode sites; inter-electrode values were derived by four-point linear interpolation.) A mid-latency component distributed over the fronto-central scalp was identified as P2. Also clearly identifiable in each condition was a later positivity whose latency and centro-parietal scalp distribution were consistent with P3.
ERPs were adjusted to baseline as determined by the average value of the 125 ms pre-stimulus recording period. Baseline-adjusted amplitude and latency values were then derived from nine (midline, lateral) recording sites using a Brain Mapping Utilities Program (Bio-Tech Interface Co.); this software identified and measured the most positive-going peak within a post-stimulus time window desig-
204 B. Palmer et al. /Biological Psychology 38 (1994) 199-214
Reaction Time Accuracy
P
_ 100 0
- 95 s
8 - 90 7
a c
- 85
.5 - - 80
1 2 3 1 2 3 Block Block
M physlcal
- name
. l category
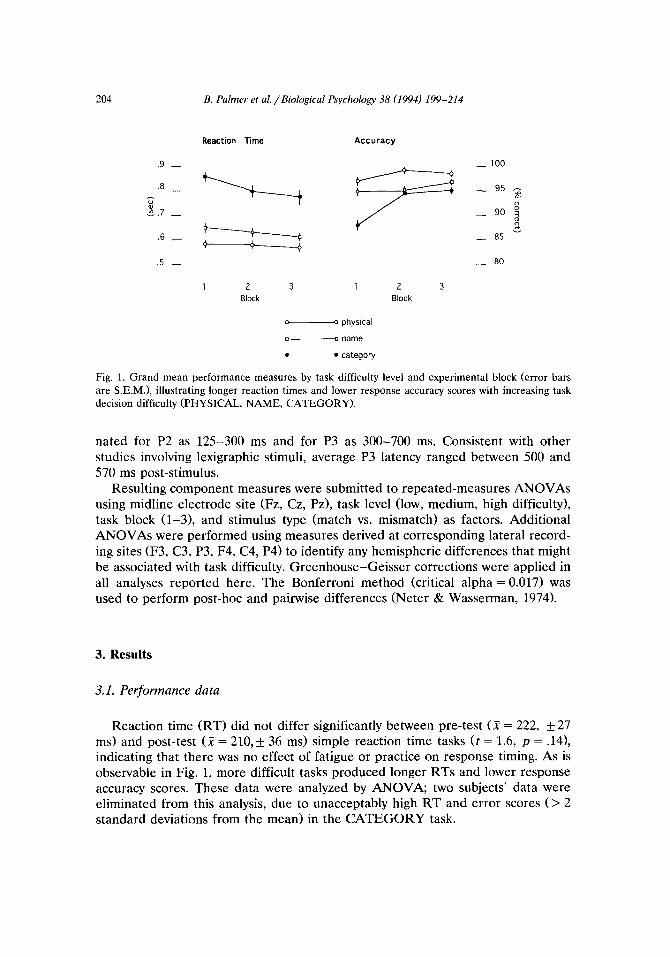
Fig. 1. Grand mean performance measures by task difficulty level and experimental block (error bars
are S.E.M.), illustrating longer reaction times and lower response accuracy scores with increasing task
decision difficulty (PHYSICAL, NAME, CATEGORY).
nated for P2 as 125-300 ms and for P3 as 300-700 ms. Consistent with other studies involving lexigraphic stimuli, average P3 latency ranged between 500 and 570 ms post-stimulus.
Resulting component measures were submitted to repeated-measures ANOVAs using midline electrode site (Fz, Cz, Pz), task level (low, medium, high difficulty), task block (l-3), and stimulus type (match vs. mismatch) as factors. Additional ANOVAs were performed using measures derived at corresponding lateral record- ing sites (F3, C3, P3, F4, C4, P4) to identify any hemispheric differences that might be associated with task difficulty. Greenhouse-Geisser corrections were applied in all analyses reported here. The Bonferroni method (critical alpha = 0.017) was used to perform post-hoc and pairwise differences (Neter & Wasserman, 1974).
3. Results
3.1. Performance data
Reaction time (RT) did not differ significantly between pre-test (X = 222, + 27 ms> and post-test (X = 210,* 36 ms) simple reaction time tasks (t = 1.6, p = .14), indicating that there was no effect of fatigue or practice on response timing. As is observable in Fig. 1, more difficult tasks produced longer RTs and lower response accuracy scores. These data were analyzed by ANOVA; two subjects’ data were eliminated from this analysis, due to unacceptably high RT and error scores (> 2 standard deviations from the mean) in the CATEGORY task.

B. Palmer et al. / B~~~ogica~ Psychology 38 f1994) 199%214 205
RTs increased significantly over PHYSICAL, NAME and CATEGORY task levels (F(2,18) = 140.7, p < .OOOl), confirming that task difficulty increased as task decision-making criteria became increasingly complex. There was a three-way interaction between task, stimulus and block (F(4,36) = 3.1, p < .05). The CATE- GORY task produced longer RTs than either the NAME (p < .OOl) or the PHYSICAL (p < .OOl> tasks; this was true for each of the three task blocks, and for both matched and mismatched letter pair presentations. However, RT de- creased with task practice only for mismatched letter pairs in the CATEGORY task (block 1 vs. block 3, p < .OOl).
There were main effects of both task difficult and task block on response accuracy; accuracy decreased with increasing task difficulty @X2,18) = 21.4, p <
,001) and increased with task practice (F(2,18) = 20.7, p < .OOll. Post-hoc tests showed that response accuracy scores were significantly reduced in the CATE- GORY (vs. PHYSICAL) task in each of the three experimental blocks. However, an interaction between task and match/mismatch stimulus type (F(2,18) = 10.5, p < .Ol> revealed that the effect of task difficulty on response accuracy was limited to matched letter presentations (PHYSICAL vs. NAME, p < .00.5; NAME vs. CATEGORY, p < .Ol; PHYSICAL vs. CATEGORY, p < .OOOS); matched letter pairs produced more errors than mismatched pairs in both the NAME and CATEGORY tasks (p < .012). Finally, an interaction between task condition and block (&X4,36) = 3.7, p < -05) revealed an effect of task practice on response accuracy only in the CATEGORY task (block 1 vs. block 2, and block 1 vs. block 3, p < .OOl).
3.2. P200
Overall ANOVAs were first performed to address possible effects of hemi- spheric (left vs. right) and/or recording site (frontal, central, parieta1) differences. There was no effect of hemisphere on P2 amplitude or latency. P2s recorded over the frontal scalp were consistently larger than those recorded over the central or parietal scalp. This was revealed as a main effect at lateral (F(2,22) = 7.96, p < .Ol> recording sites, and at midline sites as interactive with task (F(4,44) = 3.34, p < .OS> and with stimulus type and task block (F(4,44) = 3.49, p < .04). Lateral and midhne analyses also revealed main effects of experimental block on P2 amplitude (midline: F(2,22) = 4.94, p < .02) and Iatency (lateral: 1;(2,22) = 6.4,
p < .02; midline: F(2,22) = 9.3, p < .003). Separate analyses were performed at each recording site to examine further the
experimental effects of task, block, and stimulus type. At the vertex, P2 amplitude was found to increase over task blocks (F(2,22) = 7.1, p < -005); collapsed over tasks, P2 was enhanced in both the second and third task blocks relative to the first task block (p < ,005). Over the left frontal scalp (F(2,22) = 9.4, p < .Ol>, the second and third block tasks were also associated with shorter (p < .013) P2 latencies.
There was no effect of task difficulty or stimulus type on P2 at any site. A task X stimulus interaction on P2 amplitude was observed over the frontal scalp (at

206 B. Palmer et al. /Biological Psychology 38 (1994) 199-214
P3
Fig. 2. Grand mean (N = 12) ERPs to PHYSICAL, NAME, and CATEGORY tasks as recorded at
midline and lateral sites to 15-25 artifact-free trials. ERPs are collapsed across stimulus type
(match/mismatch) and block. In this and subsequent figures, stimulus onset and P3 are designated by
vertical hatched lines.
~3, ~(2,22)= 5.5, p < .OS; at Fz, F(2,22) = 4.3, p < .OS>, but no pairwise differ- ences reached significance in post-hoc testing.
3.3. P300
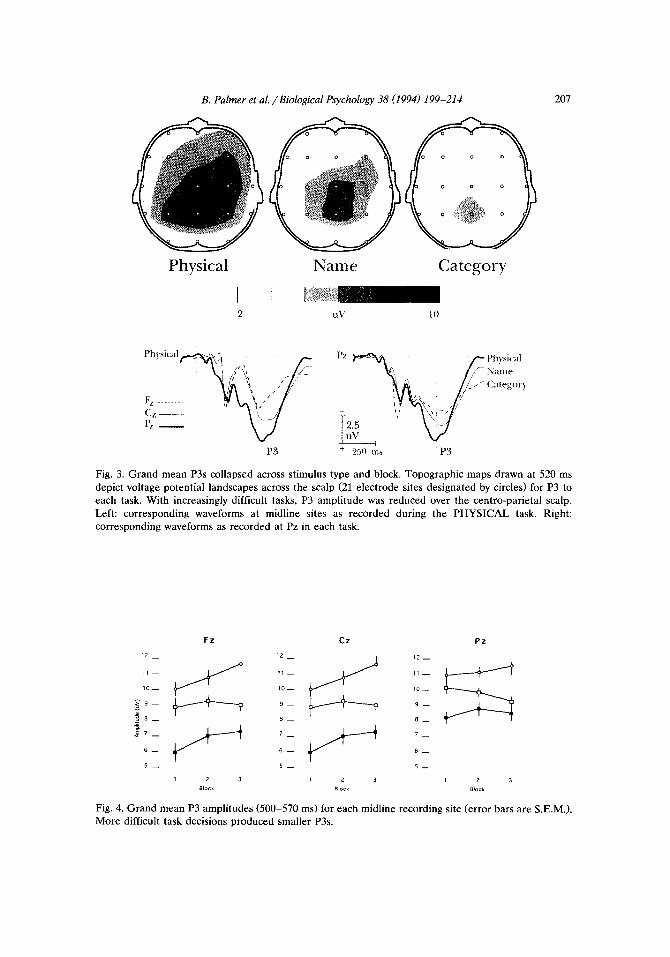
ERPs recorded at each site and to each task are shown in Fig. 2. Increased task difficulty produced an observable decrease in P3 amplitude over the midline, most pronounced over the centro-parietal scalp. Brain maps drawn for P3, collapsed across blocks and within each task, are presented in Fig. 3. Figs. 4 and 5 represent P3 amplitude means in each task level and block.
Overall ANOVAs were first performed to address possible effects of hemi- spheric (left vs. right) and/or recording site (frontal, central, parietal) differences. Lateral site analyses revealed that P3 amplitude was sensitive to complex interac- tive effects of hemisphere, stimulus, and recording site (F(4,44) = 4.4, p < .05) and hemisphere, task and recording site (F(4,44) = 3.3, p < .05); post-hoc tests re- vealed that P3 amplitude varied by hemisphere only at the central scalp (F(2,22) = 11.84, p < .0004), where P3 was enhanced (p < .017) at C4 only in response to matched letter pairs within the PHYSICAL and NAME tasks. Midline analyses revealed a main effect of recording site on P3 amplitude (F(2,22) = 7.03, p < .007), as well as interactive effects of stimulus and site (F(2,22) = 15.16, p < .0007) and
B. Palmer et al. /Biological Psychology 38 (1994) 199-214 207
Physical
I 2
Physical
Category
Fig. 3. Grand mean P3s collapsed across stimulus type and block. Topographic maps drawn at 520 ms
depict voltage potential landscapes across the scalp (21 electrode sites designated by circles) for P3 to each task. With increasingly difficult tasks, P3 amplitude was reduced over the centro-parietal scalp.
Left: corresponding waveforms at midline sites as recorded during the PHYSICAL task. Right: corresponding waveforms as recorded at Pz in each task.
5_ 5_ 5_
1 2 3 1 2 3 1 * 3
Block Block Block
Fig. 4. Grand mean P3 amplitudes (500-570 ms) for each midline recording site (error bars are S.E.M.). More difficult task decisions produced smaller P3s.
208 B. Palmer et al. /Biological Psychology 38 (1994) 199-214
8_ \
UV 4
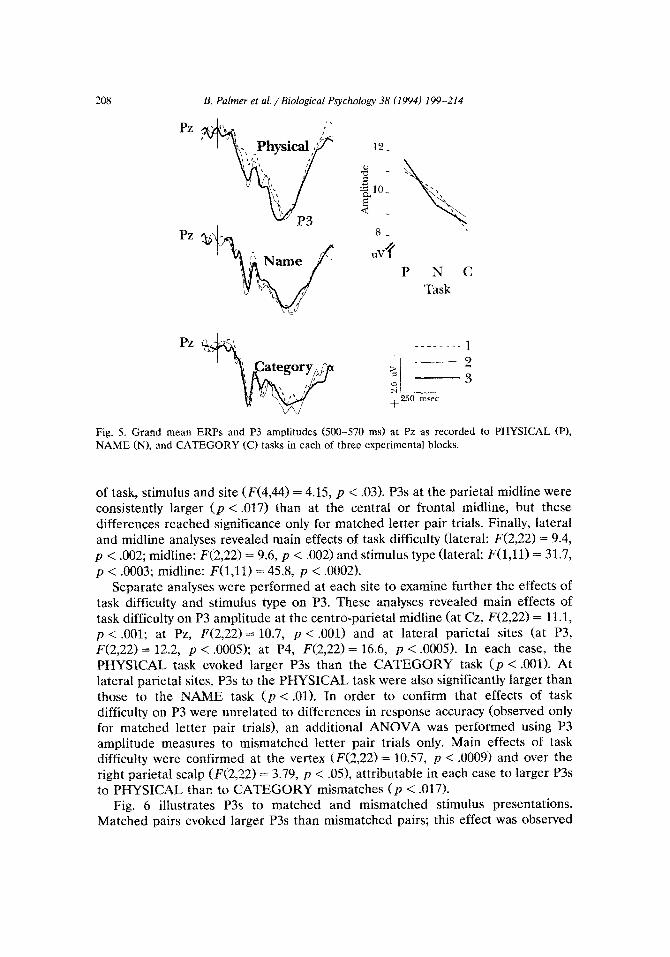
P N C Task
Fig. 5. Grand mean ERPs and P3 amplitudes (500-570 ms) at Pz as recorded NAME (N), and CATEGORY (C) tasks in each of three experimental blocks.
to PHYSICAL tP),
of task, stimulus and site (F(4,44) = 4.15, p < .03). P3s at the parietal midline were consistently larger (P < .017) than at the central or frontal midline, but these differences reached significance only for matched letter pair trials. Finally, lateral and midline analyses revealed main effects of task difficulty (lateral: P(2,22) = 9.4, p < .002; midline: F(2,22) = 9.6, p < .002) and stimulus type (lateral: F(l,ll) = 31.7, p < .0003; midline: F(l,ll) = 45.8, p < .0002).
Separate anaiyses were performed at each site to examine further the effects of task difficulty and stimulus type on P3. These analyses revealed main effects of task difficulty on P3 amplitude at the centro-parietal midline (at Cz, F(2,22) = 11.1, p < .OOl; at Pz, F(2,22) = 10.7, p < .OOl> and at lateral parietal sites (at P3, F(2,22) = 12.2, p < .OOOS>; at P4, F(2,22) = 16.6, p < .OOOS). In each case, the PHYSICAL task evoked larger P3s than the CATEGORY task (p < .OOl>. At lateral par&al sites, P3s to the PHYSICAL task were also significantly larger than those to the NAME task (p < .Ol). In order to confirm that effects of task difficulty on P3 were unrelated to differences in response accuracy (observed only for matched letter pair trials), an additional ANOVA was performed using P3 amplitude measures to mismatched letter pair trials only. Main effects of task difficulty were confirmed at the vertex (F(2,22) = 10.57, p < .0009) and over the right parietal scalp (F(2,22) = 3.79, p < .OS>, attributable in each case to larger P3s to PHYSICAL than to CATEGORY mismatches (I, < .017).
Fig. 6 illustrates P3s to matched and mismatched stimulus presentations. Matched pairs evoked larger P3s than mismatched pairs; this effect was observed

B. Palmer et al. /Biological Psychology 38 (1994) 199-214 209
Match
- 2
Mismatch
Fig. 6. Grand mean ERPs recorded at Pz to matched and mismatched stimuli in each task, collapsed
across blocks. Corresponding topographic maps drawn at 520 ms depict voltage potential landscapes
across the scalp for P3 to each stimulus type, collapsed across tasks. P3s to matched letter pairs were
larger than to mismatched letter pairs.
over the centro-parietal midline (at Cz, F(l,ll) = 41.0, p < .0005; at Pz, F(l,ll) = 48.5, p < .OOOS> and over the lateral parietal scalp (at P3, F(l,ll) = 34.3, p < .0005; at P4, F(l,ll) = 37.7, p < .OOOS). Effects of stimulus type on P3 amplitude were further confirmed by separate analysis of the third block dataset, where associated response accuracy measures were consistently high; main effects of stimulus type were observed over the vertex and across the parietal chain (p < .005), due in each case to larger P3s associated with matched letter presentations (p < .017).
4. Discussion
Increased RTs and error scores confirmed that PHYSICAL, NAME and CATEGORY levels of the same-different task involved increasingly difficult judgments. Practice effects on RT and response accuracy corresponded specifically with larger and earlier P2s, suggesting that faster RTs and improved response accuracy may have been related to improved sensory association processing.
The magnitude of P3 over the centro-parietal scalp was inversely related to the difficulty of task-related decision-making; PHYSICAL judgments evoked the largest

and CATEGORY judgments the smallest P3s. Because this experiment controlled for possible effects of stimulus probability, stimulus set variation, and nonspecific memory involvement, these results indicate a direct effect of task difficulty on P3.
It is unlikely that task effects on P3 were due to confounding effects of variation in response accuracy. Although response accuracy varied significantly by task level for matched letter pair trials only, there was no interaction between task level and stimulus type for P3. Task difficulty effects on P3 amplitude were confirmed by separate analysis of mismatched letter pair trials. In addition, although response accuracy improved with task practice, P3 demonstrated no corresponding sensitiv- ity to task practice. FinaIly, matched pairs produced the largest number of errors, and yet also produced the largest P3s, challenging the likelihood that ERPs containing more error trials might also contain smaller P3s.
The neurophysiological basis for task difficulty effects on P3 is not known. However, there are two possible explanations that should be addressed in future studies with access to ongoing EEG or single-trial ERP datasets. One possibility is that relatively more complex decisions require access to additional brain genera- tors, and thus produce phasic neural desynchronization (see Thatcher & John, 1977). Another possibility is that increased task decision difficulty leads to greater variability in the timing of mental operations that underlie P3 response production; in this case, P3 amplitude variation would be a secondary effect of increased latency variation among single-trial ERPs.
It is of interest to note that while RT varied with task difficult level, there was no effect of task difficulty on average P3 latency. The independence of stimulus and response processing time is well-established (Kutas, McCarthy & Donchin, 1977; Duncan-Johnson, 1981; Strayer, Karis, Coles & Donchin, 1984). Although longer latencies to relatively more difficult task discriminations have been reported by other investigators (McCarthy & Donchin, 1981; Goodin, Squires & Starr, 1983; Pfefferbaum, Ford, Johnson, Wenegrat & KopeIl, 1983; Magliero, Bashore, CoIes & Donchin, 1984; Johnson, Pfefferbaum & Kopell, 1985; Taylor, 1988), it has since been shown that P3 latency variation may be more directly related to direct or confound effects of stimulus complexity than to effects of task difficulty (Czigler & Szenthe, 1988). This explanation is also relatively more consistent with the notion that P3 reflects the duration of stimulus/input processing (Kutas et al., 1977; Duncan-Johnson, 1981; McCarthy & Donchin, 1981; Magliero et al., 1984).
Our use of a constant stimulus set and variable task instructions probably controlled against the possibility of P3 latency effects. Here, subjects viewed highly familiar alphabetic stimuli whose input processing is most likely automatized (Hasher & Zacks, 1979). If latency effects were a determinant contributor to the P3 amplitude effects observed here, they would be attributable to differences in the complexity of stimulus evaluation resulting only from differences in judgment criteria. Because letter pairs in every task level were drawn from the same set of four letters, latency variability difference between task levels could only be due to differences that relate directly to the fact of encoding a given stimulus on the basis of its physical, nominal or categorical identity. Although the absence of latency effects challenges the possibili~ that instructional set exerted any reliable influ-

B. Palmer et al. /Biological Psychology 38 (1994) 199-214 211
ence on stimulus input and evaluation processes, additional study is necessary to confirm direct effects on P3 amplitude by latency adjustments to single-trial waveforms.
Task difficulty effects on P3 amplitude can be considered within the context of existing theoretical frameworks concerning the behavior of the P3 component itself. Task difficulty must first be clearly distinguished from “task complexity,” considered by Johnson (1986) to be a factor that may affect stimulus meaning or significance. Within the framework of Johnson’s triarchic model, task complexity varies as a function of task response type (e.g. reaction time vs. counting vs. feedback), stimulus information load, and presumed effects of these variables on the extent of stimulus processing required for task performance. By this descrip- tion, more complex tasks generally produce relatively larger P3s (e.g. Chesney & Donchin, 1979). By contrast, we here defined task “difficult” in terms of decision-making criteria only, holding constant the parameters of task response (choice reaction time) and stimulus set (letters). This approach revealed a much different relationship than that established by varying task “complexity.”
The P3 amplitude changes observed here are more consistent with the inffuence of equivocation (Ruchkin & Sutton, 1978; Johnson, 1986). Equivocation refers to stimulus information loss that results from un~ertain~ about having judged an event correctly. According to this view, P3 amplitude varies as a function of information received, and the amount of information received is determined by (1) a priori expectation (subjective probability), and (2) a posteriori uncertainty (equiv- ocation). At a fixed level of subjective probability, equivocation should act as the primary determinant of P3 amplitude, which should in turn tend to decrease with increasing levels of post-decision uncertainty. Here, changes in P3 amplitude, RT, and response accuracy indicate a similar picture with respect to linguistic task decision difficulty.
An alternative framework is offered by Ullsperger, Gille and Metz (1987) and Ullsperger and Gille (19881, who consider P3 within the context of Adaptation- Level Theory. Specifically, Ullsperger and his colleagues have proposed that P3 amplitude varies as a function of the “mental distance” between an eliciting event and an internal “adaptation level” as originally conceived by Helson (1964). Ullsperger’s model may be useful in explaining sometimes apparently divergent results from studies that apply a variety of different task types. However, two characteristics of the present study make it difficult to interpret results within this framework. First, in Ullsperger’s view, stimulus probabili~ weighs heavily as a contributor to adaptation level; stimulus probability was eliminated as a factor in the present study. Second, because each task level was delivered separately, the question of P3 variation among difficulty levels within a single test cannot be addressed.
However, the Adaptation Level view may provide a useful context within which to consider the finding of enhanced P3s to matched (vs. mismatched) letter pairs. Matched pairs also produced the fastest RTs, consistent with earlier studies of “fast-same” effects for matching stimuli (Proctor, 1981; Farell, 1985; Czigler & Szenthe, 1988). In addition, error scores to matched stimuli were uniquely sensitive

212 B. Palmer et al. /Biological Psychology 38 (I 994) 199-214
to task difficulty level. These findings are consistent with the possibility of an adaptation level effect along the stimulus dimension itself, despite equiprobable stimulus type delivery. That is, matched letter pairs may have represented a relatively greater distance from an internal model or adaptation level. Enhanced late positivities to matched stimuli, in comparison to mismatched stimuli, have been reported by others (Posner, Klein, Summers & Buggie, 1973; Thatcher, 1977); Thatcher (1977) has proposed that this phenomenon may be due to a “representa- tional match” process associated with task-related expectancies and strategies. Although our match and mismatch trials were delivered at equal presentation probabilities, the relative overall subjective probability of a matched letter combi- nation may have been perceived by the subject as lower than that of a mismatch. In two of the three task levels, the number of matched-pair letter combinations possible a priori was, in fact, smaller than the number of possible mismatched pairs. It may be the case that subjects correctly or incorrectly perceived a lesser a priori likelihood of matched pairs; if so, lower subjective probability would tend to produce larger P3.s.
In summary, this study revealed a direct, inverse relationship between decision difficulty and P3 amplitude. Reduced P3s were associated with increasingly diffi- cult tasks in a paradigm that controlled for possible interactive effects of task response complexity, stimulus differences, memory requirements, and response probability. Although the precise neurophysiological mechanism for this effect is not yet clear, the absence of latency effects in this study suggests that P3 amplitude may have been affected directly by the difficulty of task-related decisions and the levels of uncertainty they provoked. Future research in this area should explore the role of subjective event probability as a variable that may interact with, obscure, or contribute to, the effects of task difficulty on P3. To the extent that real-world events are rarely perceived to occur at 50% probability, the likely effects of high versus low event expectancy should also be considered in any effort to apply P3 as an electrophysiologic index of mental workload.
Acknowledgments
This research was supported by a National Research Council-Armstrong Labo- ratory Research Associateship awarded to V.T.N. A preliminary report of this study was presented at the 1991 Annual Meeting of the Society for Psychophysio- logical Research.
References
Adams, N., & Collins, G.I. (1978). Late components of the visual evoked potential to search in long-term memory. Electroencephalography and Clinical Neurophysiology, 44, 147-156.
Chesney, G.L., & Donchin, E. (1979). Predictions, their confirmations, and the P300 component. Psychophysiology, 16, 174. (Abstract)

E. Palmer et al. /Biological Psychology 38 (1994) 199-214 213
Czigler, I., & Szenthe, A. (1988). Selection within fixation: Event-related potentials in a visual matching
task. ~nter~tio~l Journal of P~c~phys~olo~, 6, 39-49. Duncan-Johnson, C. (1981). P300 latency: A new metric of information processing. Psychophysiology,
18, 207-215. Farell, B. (1985). ‘Same’-‘different’ judgments: A review of current controversies in perceptual
comparison. Psychological Bulletin, 98, 419-456. Fitzgerald, P.G., & Picton, T.W. (1983). E vent-related potentials recorded during the discrimination of
improbable stimuli. Bjological P~chiat~, 17, 241-276. Fitzgerald, P.G., & Pi&on, T.W. (1984). Effects of probabiIi~ disc~minabiIi~ on the evoked potentials
to unpredictable stimuli. In: R. Karrer, J. Cohen & P. Tueting (Eds.), Brain and information: Euent-related potentials. Annals of the New York Academy of Sciences, 425, 199-203.
Friedman, D., Vaughan, H., & Erlenmeyer-Kimling, L. (19811. Multiple late positive potentials in two
visual discrimination tasks. Psychophysiology, 18, 635-649. Gevins, A.S., & CutiIIo, B.A. (1986). Signals of cognition. In: V.B. Mountcastle, F. Plum & S.R. Geiger
(Eds.), ~~andbook of physjo~o~, Vol. 5 (pp. 335-381). Bethesda, MD: American P~choIogicaI
Society. Gomer, FE., Spicuzza, R.L., & O’Donnell, R.D. (19761. Evoked potential correlates of visual item
recognition during memory-scanning tasks. Physiological Psychology, 4, 61-65. Goodin, D.S., Squires, K.C., & Starr, A. (1983). Variations in early and late event-related components
of the auditory evoked potential with task difficulty. Electroencephalography and Clinical Neurophys- iology, 55, 680-686.
Hasher, I., & Zacks, R.T. (19791. Automatic and effortful processes in memory. Journal of ~perimental P~cholo~: General, 103, 356-388.
Helson, H. (1964). Adaptation-level theory: An experimental and systematic approach to behavior. New York: Harper and Row.
Hillyard, .%A., & Kutas, M. (1983). Electrophysiology of cognitive processing. Annual Reuiew of Psychology, 34, 33-61.
Hillyard, S.A., Squires, K.C., Bauer, J.W., & Lindsay, P.H. (1971). Evoked potentiai correlates of
auditory signal detection. Science, 172, 1357-1360.
Jasper, H.H. (1958). Report of the committee on methods of clinical examination in electroencephalog-
raphy. Electroencephalography and Clinical Neurophysiology, IO, 370-375. Johnson, R. Jr. (19861. A triarchic model of P300 amplitude. Psychophysiology, 23, 367-384. Johnson, R. Jr., Pfefferbaum, A., & Kopell, B.S. (19851. P300 and long-term memory: Latency predicts
recognition performance. Psychophysiology, 22, 497-507. Krueger, L.E. (1984). The category effect in visuai search depends on physical rather than conceptual
differences. Perceptjon and P~chophysics, 35, 558-564. Kutas, M., McCarthy, G., & Donchin, E. (19771. Augmenting mental chronometry: The P300 as a
measure of stimulus evaluation time. Science, 197, 792-795. Magliero, A., Bashore, T.R., Coles, M.G.H., & Donchin, E. (1984). On the dependence of P300 latency
on stimulus evaluation processes. Psychophysiology, 21, 171-186. McCarthy, G., & Donchin, E. (1981). A metric for thought: A comparison of P300 latency and reaction
time. Science, 211, 77-80. Mecklinger, A., Kramer, A.F., & Strayer, D.L. (1992). Event related potentials and EEG components in
a semantic memory search task. P~chophysiolu~, 29, 104-119. Neter, J., & Wasserman, W. (1974). Applied linear statistical models. IL: Richard E. Irwin, Inc.
PacheIIa, R.G., & Miller, J.O. (1976). Stimulus probability and same-different classification. Perception & Psychophysics, 19, 29-34.
Pfefferbaum, A., Ford, J., Johnson, R.Jr., Wenegrat, B., & KopeII, B.S. (1983). Manipulation of P3
latency: Speed vs. accuracy instructions. Ele~troencephalography and Clinical Neurophysiology, 55, 188-197.
Polich, J. (1987). Task difficulty, probabiIi~, and inter-stimulus intervai as determinants of P3Ot_l from
auditory stimuli. Electroencephalography and Clinical Neurophysiology, 68, 31 l-320.
Poon, P.W., Thompson, L.W., & Marsh, G.R. (1976). Average evoked potential changes as a function of processing complexity. Psychophysiology, 13, 43-49.

214 B. Palmer et al. /Biological Psychology 38 (1994) 199-214
Posner, MI., Klein, R., Summers, J., & Buggie, S. (1973). On the selection of signals. Memory and Cognition, I, 2-12.
Posner, M.I., & Mitchell, R.F. (1967). Chronometric analysis of classification. Psychological Reuiew, 74, 392-409.
Prinz, W. (1980). Selectivity in character classification. In: R.S. Nickerson, Attention and performance VIII. Hillsdale, NJ: Lawrence Erlbaum Associates.
Proctor, R.W. (1981). A unified theory for matching task phenomena. Psychological Reuiew, 82, 116-149.
Ruchkin, D.S., & Sutton, S. (1978). Equivocation and P300 amplitude. In: D. Otto (Ed.), Multidisci- plinary perspectives in event-related brain potential research (pp. 175-177). Washington: U.S. Environ-
mental Protection Agency.
Ruchkin, D.S., Sutton, S., Kietzman, M.G., & Silver, K. (1980). Slow wave and P300 in signal detection. Electroencephalography and Clinical Neurophysiology, 50, 35-47.
Sergeant, J., Geuze, R., & van Winsum, W. (1987). Event-related desynchronization and the P300.
Psychophysiology, 24, 272-277. Shingledecker, C.A. (1984). A task battery for applied human performance assessment research.
AFAMRLTR-84-071. Air Force Aerospace Medical Research Laboratory, Aerospace Medical
Division. Squires, K.C., Squires, N.K., & Hillyard, S.A. (1975). Decision-related cortical potentials during an
auditory signal detection task with cued observation intervals. Journal of Experimental Psychology: Human Perception and Performance, 1, 268-279.
Strayer, D.L., Karis, D., Coles, M.G.H., & Donchin, E. (1984). P300 latency and reaction time are
dissociated in a Sternberg task. Psychophysiology, 21, 600. (Abstract)
Sutton, S., Ruchkin, D.S., Munson, R., Kietzman, M.L., & Hammer, M. (1982). Event-related poten- tials in a two-interval forced choice discrimination task. Perceptual Psychophysiology, 32, 360-374.
Taylor, D.A. (1978). Identification and categorization of letters and digits. Journal of Experimental Psychology: Human Perception and Performance, 4, 423-439.
Taylor, M.J. (1988). Developmental changes in ERPs to visual language stimuli. Biological Psychology, 26, 321-338.
Thatcher, R.W. (1977). Evoked-potential correlates of delayed letter-matching. Behauioral Biology, 26, 321-338.
Thatcher, R.W., & John, E.R. (1977). Foundations of cognitive processes: Functional neuroscience (Vol.
1). New York: Wiley. Ullsperger, P., Gille, H.G., & Metz, A.M. (lY87). The P300 as a metric in psychophysics of cognitive
processes. In: C. Barber & T. Blum (Eds.), Evoked potentials III: The third international evoked potential symposium. Borough-Green: Butterworth. (pp. 355-360)
Ullsperger, P., & Gille, H.G. (1988). The late positive component of the ERP and adaptation-level
theory. Biological Psychology, 26, 299-306. Wilson, G.F., Swain, R.A., & Davis, I. (1988). Human event-related potentials during spatial process-
ing: A topographic distribution. Society for Neuroscience Abstracts, 14, 1014.