targeting histone deacetylases for cancer...
TRANSCRIPT

Dr. Wilner Martínez-López
Epigenetics and Genomic Instability Laboratory
Instituto de Investigaciones Biológicas Clemente Estable (IIBCE)
Montevideo - Uruguay
Targeting histone deacetylases for
cancer therapy

Respuesta celular al daño inducido sobre el ADN
Detención del ciclo celular
Reparación del ADN
Apoptosis
Completo
Incompleto
Mutación
Aberraciones Cromosómicas
cáncer y enfermedades hereditarias
ADN

El ADN es empaquetado
por proteínas histónicas
y no histónicas en
cromatina.
La cromatina es un
material altamente
dinámico, que posee
información epigenética.

ESTRUCTURA Y FUNCIÓN DE
LA CROMATINA
Cromatina nucleosoma
estructura superior de la cromatina
regulación de la estructura de la cromatina
Código de histonas – mecanismos epigenéticos
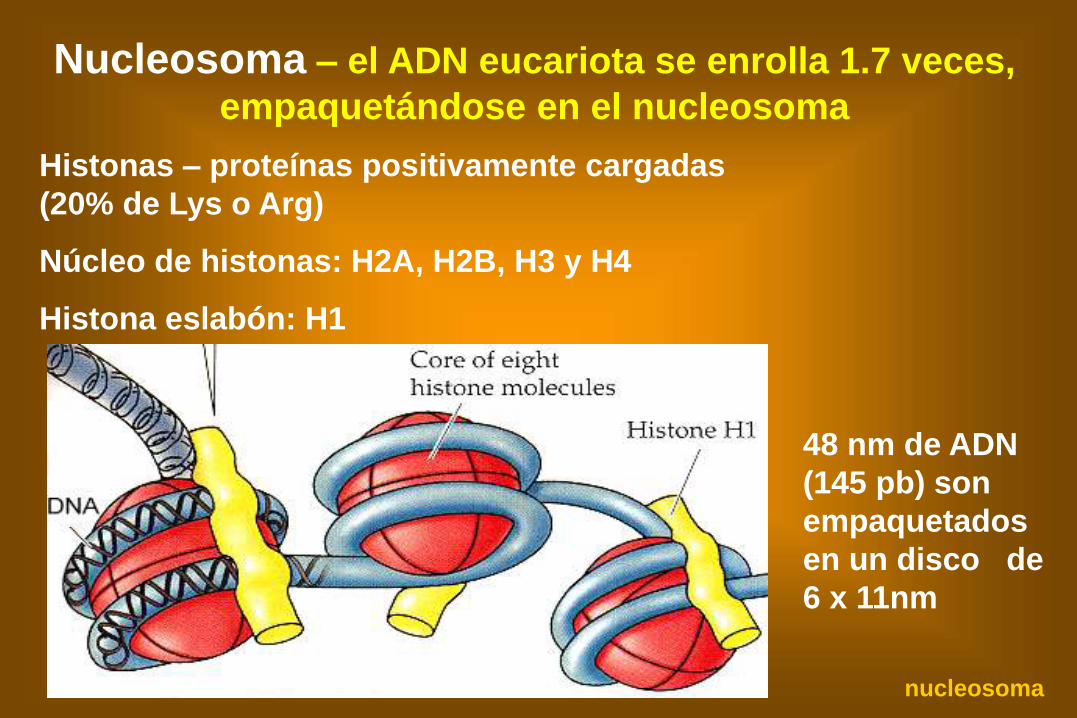
Nucleosoma – el ADN eucariota se enrolla 1.7 veces,
empaquetándose en el nucleosoma
Histonas – proteínas positivamente cargadas
(20% de Lys o Arg)
Núcleo de histonas: H2A, H2B, H3 y H4
Histona eslabón: H1
48 nm de ADN
(145 pb) son
empaquetados
en un disco de
6 x 11nm
nucleosoma

Dominios de plegamiento – 3
α-hélices separadas por lazos
sin estructura.
Ensamblado del
nucleosoma nucleosoma
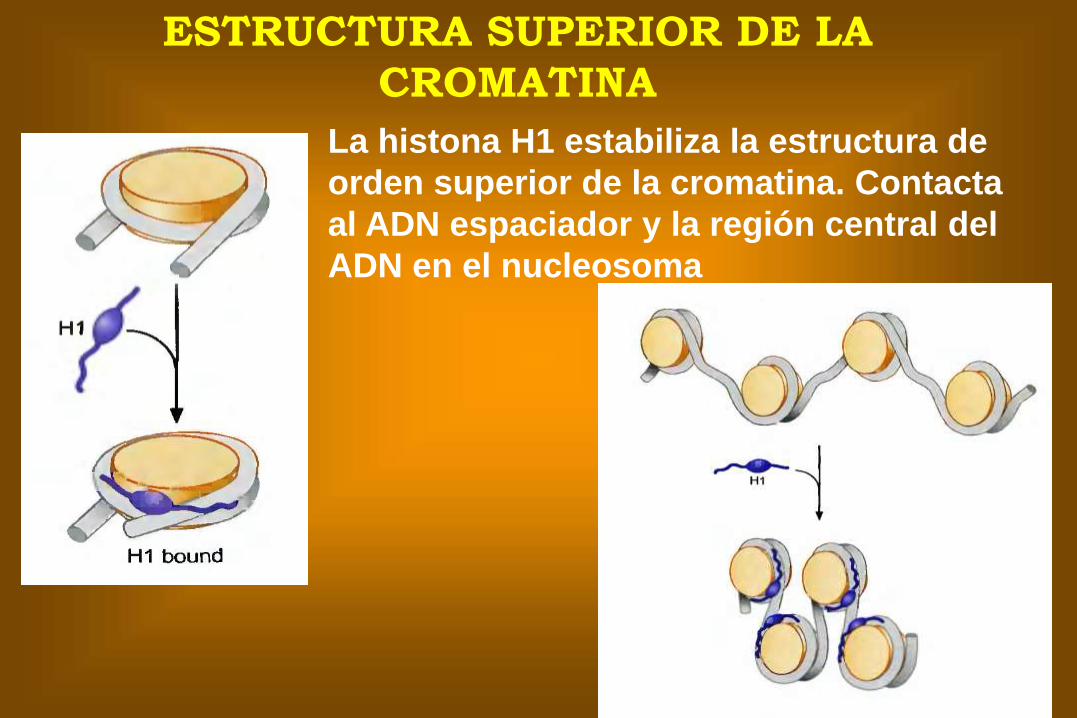
ESTRUCTURA SUPERIOR DE LA
CROMATINA
La histona H1 estabiliza la estructura de
orden superior de la cromatina. Contacta
al ADN espaciador y la región central del
ADN en el nucleosoma

Fibra de cromatina de 30 nm
Seis nucleosomas por
cada vuelta: Solenoide
Una vuelta completa:
1.2 Kb de ADN
H1
Condensinas
Topoisomerasas

Fibra de cromatina de 30 nm
se une formando lazos a un armazón proteico
Organización de la cromatina en interfase y en
cromosomas mitóticos
Cromosomas mitóticos
Scaffold
Interfase
Matriz nuclear

A- Información genética
Existen por lo menos dos formas de
información en el núcleo de la célula:
B- Información epigenética

EPIGENETICA
La epigenética es el estudio de los mecanismos
heredables que afectan el estado transcripcional de
un gene el cual no puede ser explicado por la
secuencia de ADN y que puede persistir por una o
más generaciones.

EPIGENETICA• Sistemas de metilación de ADN
• Remodelación de la cromatina mediante
modificaciones de las histonas:
metilación, fosforilación,
acetilación, ubiquitinación,
ADP ribosilación• Mecanismos de remodelación de la cromatina
dependientes de ATP:
ISWI, SWI/SNF2
• ARN no codificante
Metilación de ADN dirigida por ARN
Silenciamiento génico postranscripcional
ARN de interferencia

REGULACIÓN DE LA ESTRUCTURA DE LA
CROMATINA
La interacción ADN-octámero de histonas es
dinámica, permitiendo el acceso a regiones
particulares del ADN
Complejos remodeladores del
nucleosoma
Modificaciones covalentes de las
histonas

REMODELADORES DE LA CROMATINA
DEPENDIENTES DE ATP
switching (SWI)
and sucrose
non-fermenting
(SNF)
Imitation SWI

Clasificación según la identidad de la subunidad ATPasa:
SWI2/SNF2 (“switch sucrose non fermenting”)
ISWI (“imitation SWI”)
Regulación
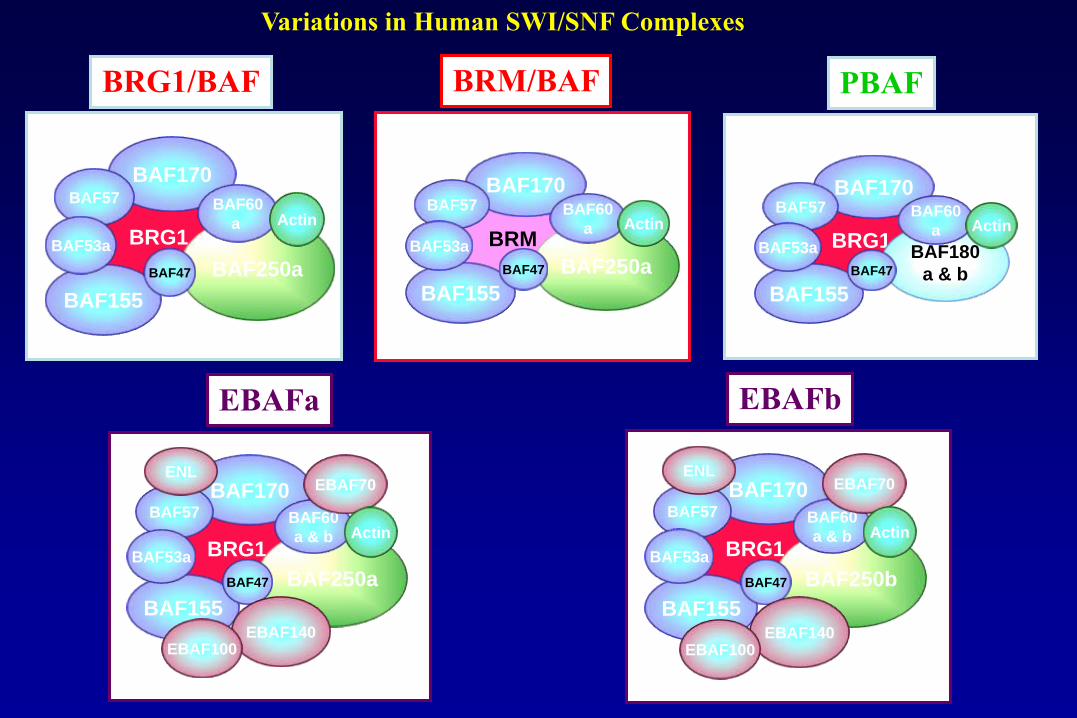
Variations in Human SWI/SNF Complexes
Actina
BRG1/BAF
BRG1
BAF155
BAF170BAF57
BAF53a
BAF250a
BAF60
a
BAF47
Actin
PBAF
BRG1
BAF155
BAF170BAF57
BAF53a BAF180
a & b
BAF60
a
BAF47
Actin
BRM/BAF
BRM
BAF155
BAF170BAF57
BAF53a
BAF250a
BAF60
a
BAF47
Actin
BRG1
BAF155
BAF170BAF57
BAF53a
BAF250b
BAF60
a & b
BAF47
EBAF70
EBAF140EBAF100
Actin
ENL
EBAFb
BRG1
BAF155
BAF170BAF57
BAF53a
BAF250a
BAF60
a & b
BAF47
EBAF70
EBAF140EBAF100
Actin
ENL
EBAFa

Regulación
Los remodeladores ATP
dependientes y las
modificaciones covalentes de
histonas trabajan juntas para
incrementar la accesibilidad al
ADN

Modificaciones covalentes – las
modificaciones en las colas N-
terminal de las histonas alteran
el acceso a la cromatina
Regulación
Acetilación
Deacetilación
Metilación
Fosforilación
Ubiquitinación
ADP-ribosilaciónLas colas de histonas están involucradas
en la formación de la fibra de 30nm
sus modificaciones afectan la estructura
de 30nm
Modificaciones
post-traduccionales

En general, el código genético incluye 4
nucleótidos, A G T y C, y el nivel de
complejidad está basado en la
combinación potencial de estas 4 bases.
Existe tambien un “código de histonas”
epigenético, el cual es mucho más
complejo y es parcialmente responsable
de la regulación funcional del código
genético en una célula en particular y
dentro de un ambiente determinado.
El código de histonas incluye:
Acetilación (mono-, di, and tri-)
Metilación (mono-, di-, and tri-)
Fosforilación
Ubiquitinación
La hipótesis del Código de Histonas

Hipótesis Código de Histonas
• Modificaciones de las histonas
• Variantes de histonas
• Enzimas responsables de las modificaciones de histonas
• Coordinación de la modificación de histonas
• Dominios de proteínas que se unen con las histonas modificadas

• Different combinations of histone modifications, especially located near or within a
gene’s promoter, may be VERY SPECIFIC to the transcriptional state of the gene
Peterson CL et al, Current Biology, 2004
Modificaciones de histonas
Hipótesis Código de Histonas


Modificaciones post-traduccionales de
las histonas

Methylated histones first described in 1964, Murray 1964 Biochemistry 3, 10-15
Hipótesis Código de HistonasEnzimas que proveen la metilación de las histonas

Metzger et al. (2005)
Shi et al., (2004).
Histona demetilasa LSD-1
Hipótesis Código de Histonas


H1
H2B
H2A
H3
H4
hélice
variable
conservada
La acetilación de histonas
es una modificación reversible
de los extremos N-terminal
del octámero de histonas.
Resultado:
• reducción de la unión al ADN
• desestabilización de la cromatinaN
Histona Peso molecular Numero de aminoácidos Contenido de aminoácidos básicos
LAS HISTONAS
son
proteínas básicas
altamente conservadas
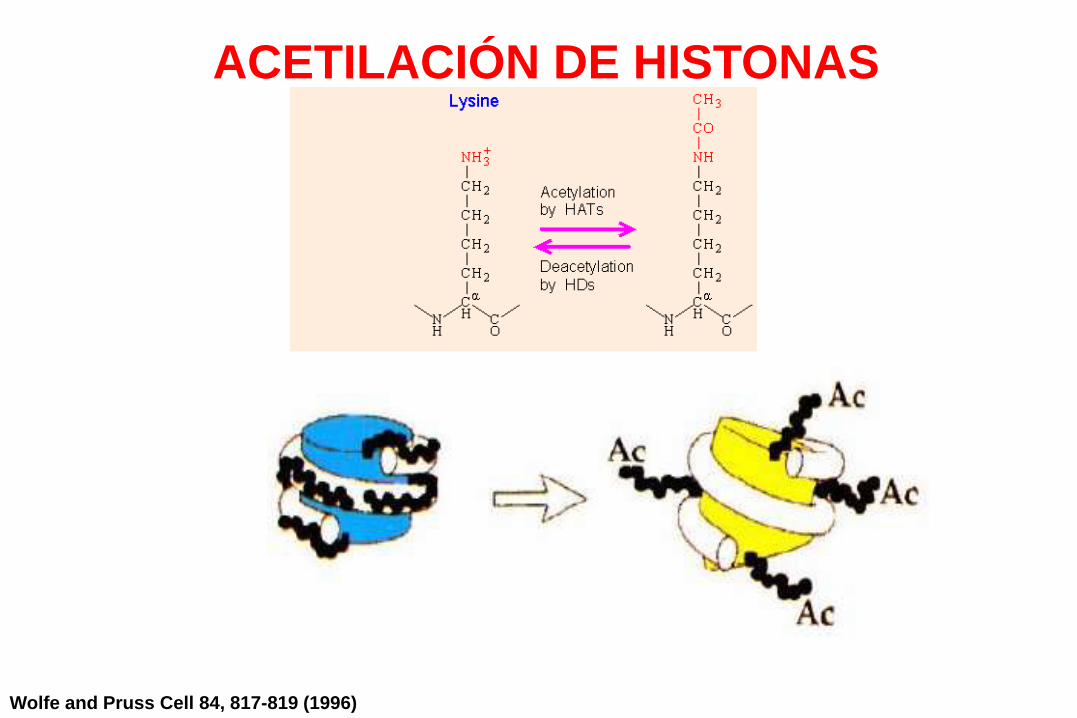
Wolfe and Pruss Cell 84, 817-819 (1996)
ACETILACIÓN DE HISTONAS
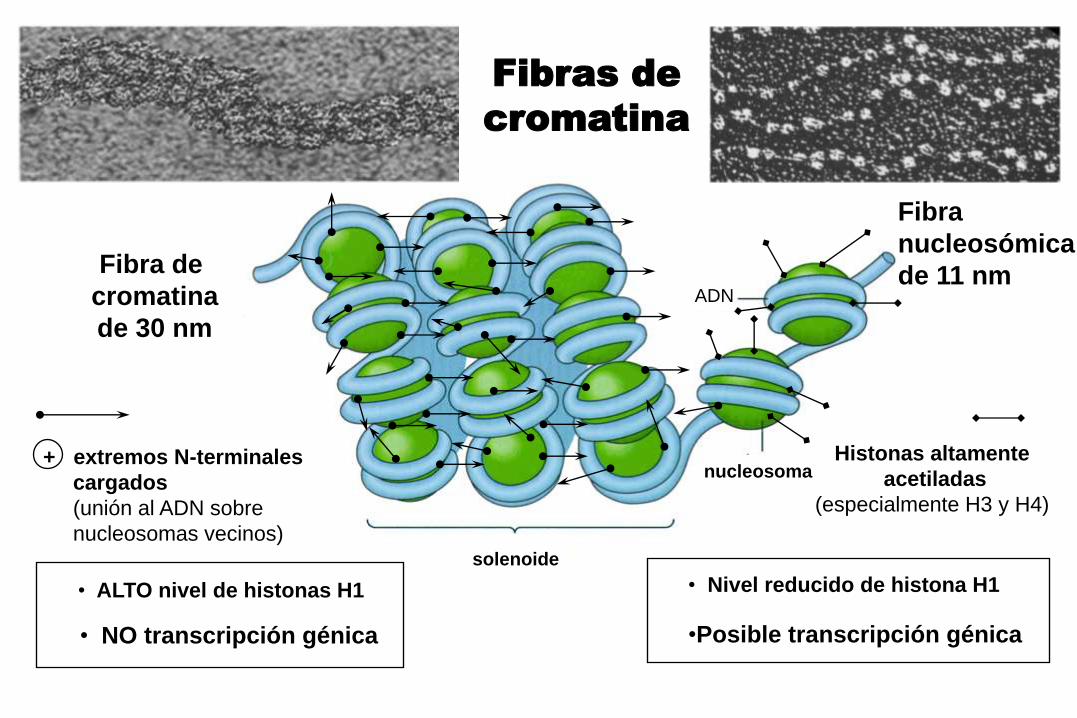
Fibras de
cromatina
+ extremos N-terminales
cargados
(unión al ADN sobre
nucleosomas vecinos)
Histonas altamente
acetiladas
(especialmente H3 y H4)
Fibra de
cromatina
de 30 nm
Fibra
nucleosómica
de 11 nm
• ALTO nivel de histonas H1 • Nivel reducido de histona H1
• NO transcripción génica •Posible transcripción génica
solenoide
nucleosoma
ADN


Variantes de histonas
K. Sharma 2005,NATURE REVIEWS
Hipótesis Código de Histonas

“Interplay” entre diferentes
mecanismos epigenéticos
Margueron ,Current Opinion in Genetics & Development (2005)

Histone Code Hypothesis
• As proposed by Allis and Strahl: “that multiple histone
modifications, acting in a combinatorial or sequential
fashion on one or multiple histone tails, specify unique
downstream functions”
Strahl, B.D. and Allis, C.D., Nature. 2000

35
Combinatorial histone modifications

Something is writing this code….
and
Something is reading this code…
Hipótesis Código de Histonas
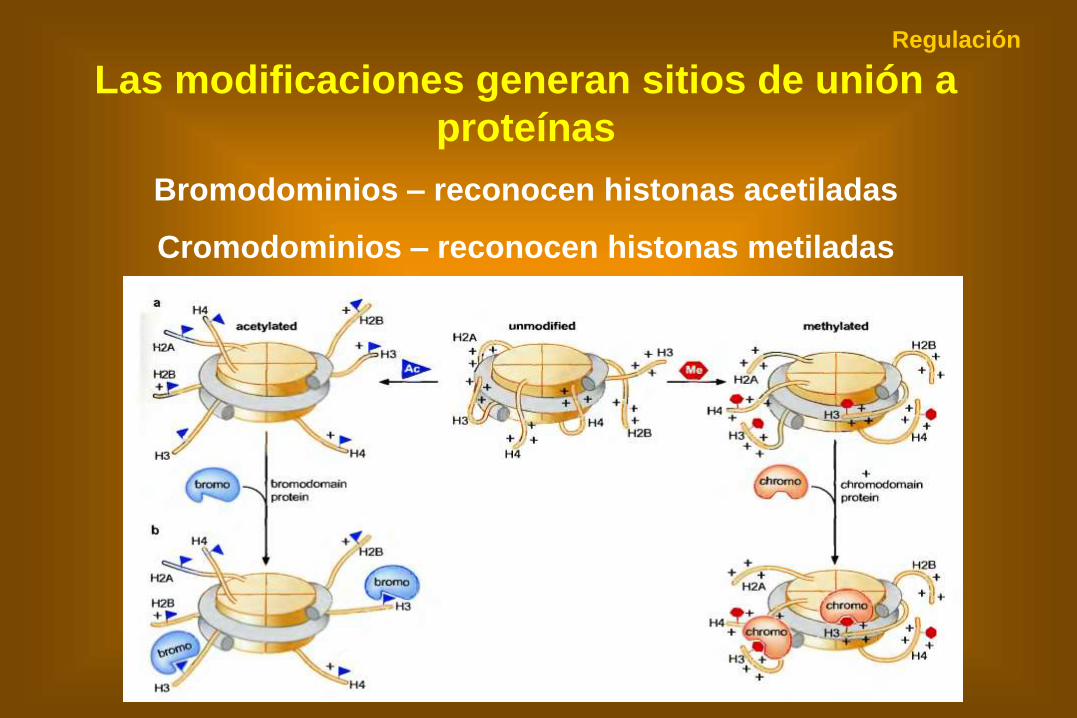
Las modificaciones generan sitios de unión a
proteínas
Bromodominios – reconocen histonas acetiladas
Cromodominios – reconocen histonas metiladas
Regulación
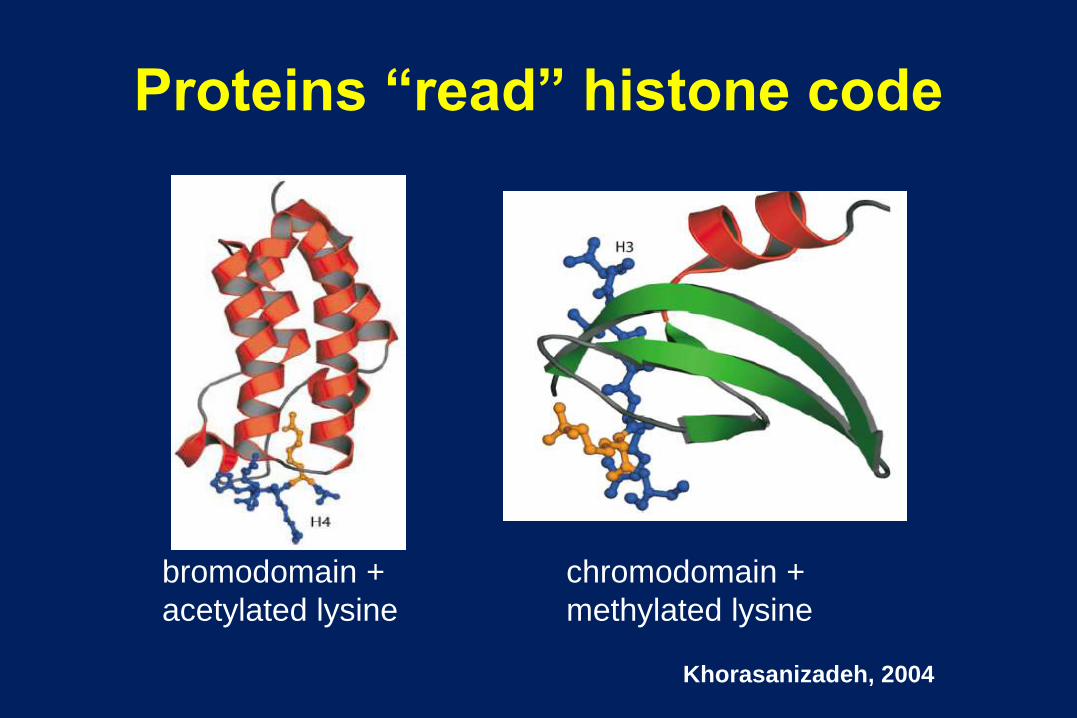
Proteins “read” histone code
bromodomain +
acetylated lysine
chromodomain +
methylated lysine
Khorasanizadeh, 2004

The combination of phosphorylation of
serine residue 10 and acetylation of a
lysine residue 14 on histone H3
correlate with active transcription.
Methylated lysine 9 and lysine 27 on
histone H3 are associated with
repressed genes.
Interdependencia entre modificaciones de histonas
Phosphorylation of serine residues 10 and 28 on histone H3
is a marker for chromosomal condensation.
Methylated lysine 4, 36 and 79 of H3 are associated with
“active” chromatin

Modificaciones de la superficie lateral
La energía de hidrólisis del ATP de los remodeladores de
histonas puede usarse para separar el ADN del octámero de
histonas y exponer sus aminoácidos laterales para su
modificación.
H3 Thr 118 se une por puente de
hidrógeno al esqueleto de ADN y
a H4 Arg45 . La fosforilación de
Thr 118 altera la interacción de H4
Arg45 con el surco menor,
permitiendo el deslizamiento del
octámero sobre el ADN
epigenesis

Protein code readers bind to specific sets of modifications leading to downstream effects such as gene transcription, DNA synthesis and repair.
Code Readers
Cellular Decisions
Modified Histones

Nuestro Epigenoma
• If epigenetic markers are dynamic and respond to environmental
influences, do they change over time?
– Evidence suggests the answer is yes.
• Twin studies have been highly informative for this question.

Author Statement
• “We found that, although twins are epigenetically indistinguishable
during the early years of life, older monozygous twins exhibited
remarkable differences in their overall content and genomic
distribution of 5-methylcytosine DNA and histone acetylation,
affecting their gene-expression portrait.”

Chromosomal Level
• Comparative genomic
hybridization for methylated
DNA
– Yellow = similar chromosome
methylation pattern between
twins.
– Red = regions of
hypomethylation in one twin
compared to the other.
– Green = regions of
hypermethylation in one twin
compared to the other.

Evidencias epidemiológicas sugieren que la exposición ambiental
temprana durante el desarrollo posee un rol en la susceptibilidad a
enfermedades a lo largo de nuestra vida. Algunos de estos efectos
podrían pasar a sucesivas generaciones
Las modificaciones epigenéticas constituirían un “link” entre el medio
ambiente y las alteraciones en la expresión génica que podrían
conducir a las enfermedades
Epigenetica y Medio Ambiente

Epigenética y CancerLos diferentes estados de la cromatina son esenciales para el
control génico
Es importante no solo encender los genes correctos sino tambienapagar aquellos que no deben ser expresados
Los componentes de la cromatina poseen un papel fundamentalen la decisión de estos procesos
El silenciamiento epigenético de genes constituye una nuevaherramienta para la terapia anti-neoplásica

Example: Replication errors
GENETIC
Altered DNA/mRNA/proteins
Altered DNA sequence
Historicamente se consideraba que el
cancer derivaba de cambios genéticos
XX
Oncogenesis
Tumor

Example: Replication errors
GENETIC EPIGENETIC
Example: Chromatin modification errors
Altered DNA/mRNA/proteins
Altered DNA sequence
Altered levels ofmRNA/proteins
Alteredchromatin structure
En las últimas décadas se ha demostrado que
los cambios epigenéticos son tambien
importantes en el desarrollo del cáncer
XX
Oncogenesis
Tumor
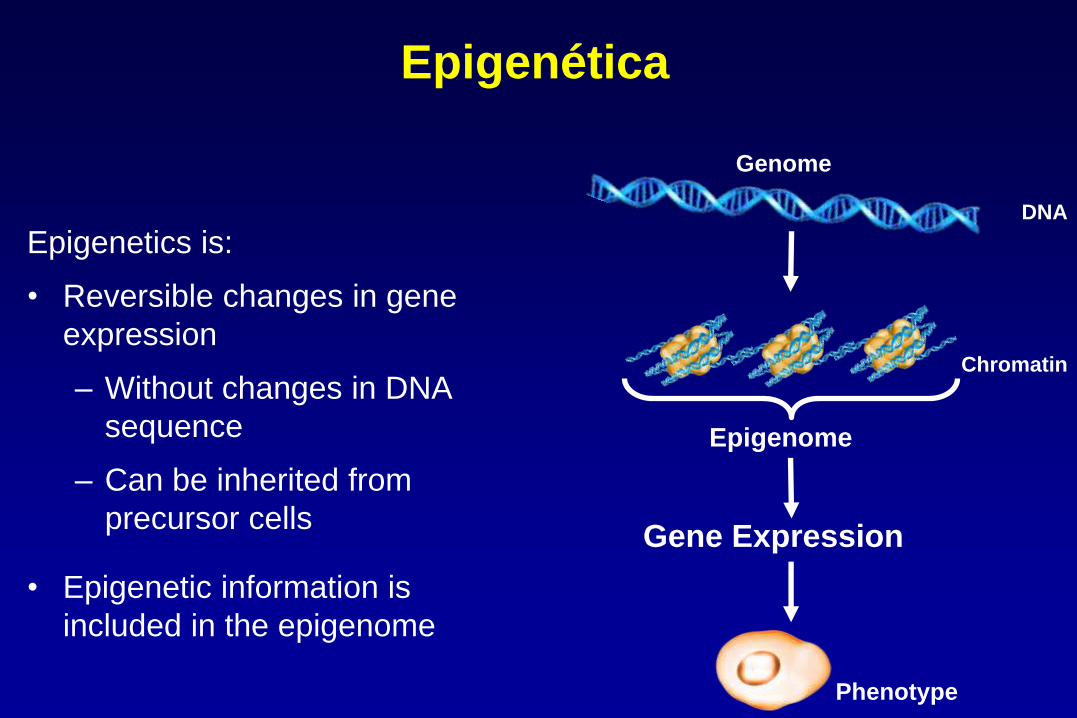
Epigenetics is:
• Reversible changes in gene
expression
– Without changes in DNA
sequence
– Can be inherited from
precursor cells
• Epigenetic information is
included in the epigenome
Genome
DNA
Gene Expression
Epigenome
Chromatin
Phenotype
Epigenética

Los mecanismos epigenéticos juegan un
importante papel en el desarrollo de una célula
normal como en una cancerosa
Epigenetic mechanisms can regulate genes involved in differentiation, cell cycle, and
cell survival
Deregulation of epigenetic mechanisms results in aberrant gene expression, which
can lead to cancer
Reversal of deregulated epigenetic changes is a rational strategy for targeting cancer
EPIGENETICS
Normal differentiated cells, e.g. embryonic
cells, hematopoetic cells
Malignant
progenitor cell Tumor
Normal epigenetic
mechanisms
Deregulated epigenetic
mechanisms

En el cáncer, cambios en la estructura de la
cromatina pueden silenciar Genes
Supresores de Tumores
Silencing of tumor suppressor genes, a major process in tumorigenesis, may result from epigenetic changes that condense chromatin structure
Cancer
cells
Tumor suppressor
gene expression
Tumor suppressor gene
expression
Normal
cellsAc
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
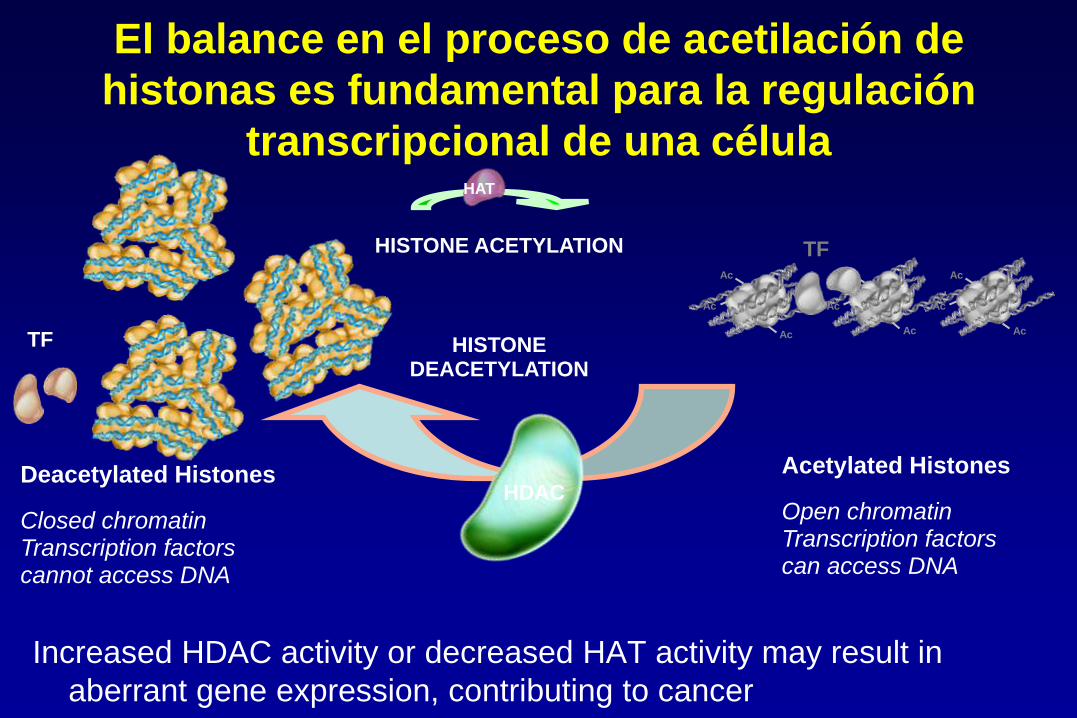
Increased HDAC activity or decreased HAT activity may result in
aberrant gene expression, contributing to cancer
HISTONE ACETYLATION
HISTONE DEACETYLATION
Acetylated Histones
Open chromatin Transcription factors can access DNA
Deacetylated Histones
Closed chromatin Transcription factors cannot access DNA
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
HDAC
HAT
TF
TF
El balance en el proceso de acetilación de
histonas es fundamental para la regulación
transcripcional de una célula
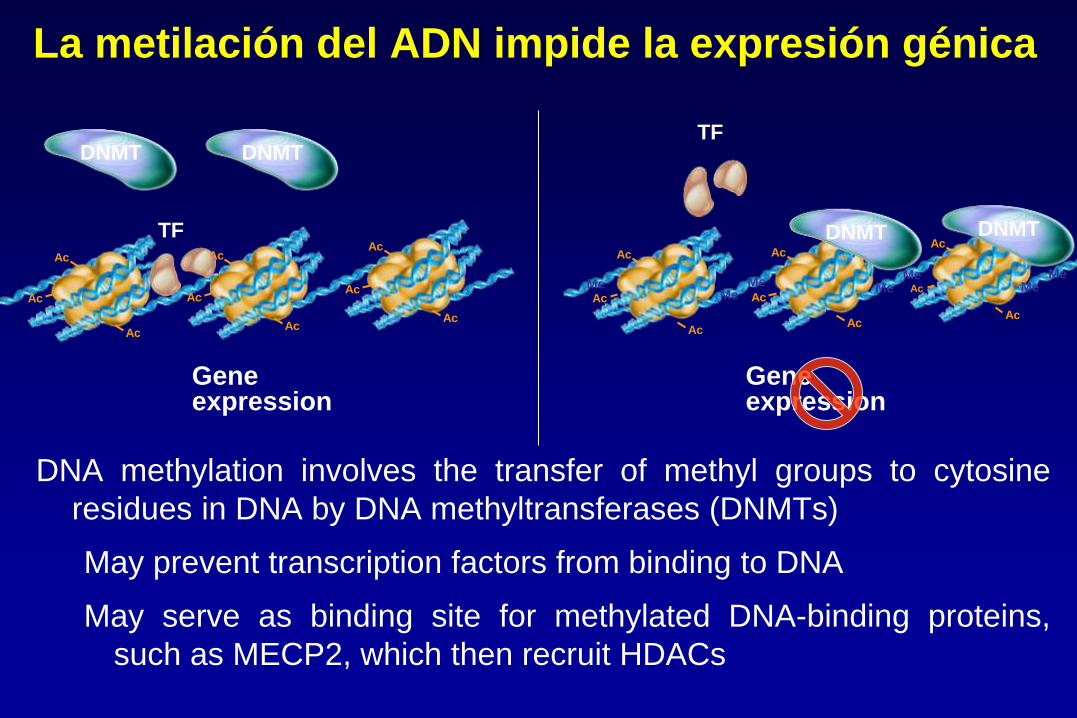
La metilación del ADN impide la expresión génica
DNA methylation involves the transfer of methyl groups to cytosine
residues in DNA by DNA methyltransferases (DNMTs)
May prevent transcription factors from binding to DNA
May serve as binding site for methylated DNA-binding proteins,
such as MECP2, which then recruit HDACs
MeMeMe Me
MeMe
Me
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
Ac
DNMT DNMT
DNMT DNMT
Geneexpression
Geneexpression
TF
TF

• Inhibition of HDAC activity
can restore the balance of
histone acetylation
• Targeting HDAC activity
may therefore allow
re-expression of silenced
genes to reverse
oncogenesis
La inhibición de HDACs puede revertir la
expresión génica alterada
Gene
expression
Ac Ac Ac
Differentiation
Growth control
Cell-cycle arrest
Apoptosis
Adhesion
Cell nucleus
HDACTF
DAC
Inhibitor

Since DNA methylation and
histone deacetylation can
co-operate to silence tumor
suppressors, inhibition of
both DNMT and HDAC
activities can synergize to
restore expression of
silenced genes
Una terapia combinada puede ser más eficiente
DAC
Inhibitor
Gene
expression
Ac
Ac Ac
Ac
Ac
Ac
Cell nucleus
HDAC
DNMT
DNMT
Inhibitor
TF
Differentiation
Growth control
Cell-cycle arrest
Apoptosis
Adhesion



Resumen
Los procesos epigenéticos regulan la expresión génica y conductacelular
Los cambios epigenéticos que conducen al silenciamiento de geneses central en la patogénesis de la enfermedades malignas, tantoen tumores sólidos como hematológicos
La deacetilación de histonas y la metilación del ADN representan dosmodificaciones epigenéticas con relevancia clínica en oncogénesis
Los blancos terapéuticos de la terapia epigenética pueden restaurarla expresión de genes que son críticos en el control del normalcrecimiento de la célula
Los inhibidores de HDACs muestran sinergismo con los agentesdemetilantes del ADN para inhibir el crecimiento tumoral

EPIGENETICA Y FARMACOLOGIA
• A diferencia de las patologías con base genética, lasrelacionadas con cambios epigenéticos sonreversibles.
• Las drogas epigenéticas pueden ser empleadas yasea para activar o silenciar genes relacionados conpatologías.
• Diferentes estados epigenéticos pueden condicionar larespuesta a drogas
• La farmacoepigenómica constituye un campoemergente.


Peedicayil 2006

Ensayo de terapias para incrementar la
sensibilidad al tratamiento quimioterápico en
líneas celulares derivadas de cáncer de vejiga
IIBCE-FMED
Tratamiento estándar: resección transuretral del tumor seguido de la
instilación intravesical de agentes de quimioterapia.
Recidiva: 70% con un 30% de neoplasias invasoras y mortalidad del
80% a 5 años (Swana et al., 1999).
Sobre-expresión de SURVIVINA, hecho asociado con el elevado
porcentaje de recidiva y su mal pronóstico en términos de sobrevida.
CARACTERISTICAS MAS SALIENTES

ESTRATAEGIA DE LA INVESTIGACION1. Estudiar el efecto de la inhibición de expresión de survivina
(mediante ARN de interferencia) en relación al tratamiento
quimioterápico que habitualmente se emplea en la terapia
anti-neoplásica en el cáncer de vejiga.
2. Analizar el efecto de la remodelación de la cromatina
(mediante cambios en el patrón de acetilación de histonas)
en la respuesta al tratamiento quimioterápico de líneas
celulares derivadas de cáncer de vejiga que expresan
diferentes niveles de survivina.
3. Asociación de ambas terapias con el fin de incrementar la
sensibilidad al tratamiento quimioterápico en las líneas
celulares derivadas de cáncer de vejiga.

Terapia con ARN de interferencia (ARNi)
anti-survivinaDiseño y sintesis de oligodeoxinucleótidos
a partir de 5 secuencias del mensajero de
survivina. Generación de un “shRNA”
(“short hairpin RNA”) clonado en el
plásmido pSilencer 1.0 para ser empleado
como ARN de interferencia.
Generación de un anticuerpo policlonal
específico contra todas las variantes de
“splicing” de survivina con el fin de poder
cuantificar su expresión en células
tumorales mediante la realización de
“western blots”.
Recuento de aberraciones cromosómicas y
porcentaje de células apoptóticas
Anticuerpo
comercial
Control
()
Anti-survivina
1/4000
Anti-survivina
1/8000
Anti-survivina
1/16000
ARNm de survivina
1619 bp
Exon I
Exon II
Exon III
Exon IV
shRNA_5 (100.0%)
shRNA_4 (100.0%)
shRNA_3 (100.0%)
shRNA_2 (100.0%)
shRNA_1 (100.0%)

Co-transfección con lipofectamina de los plásmidos con un vector
que produce la proteína fluorescente verde (pcEGFP, utilizada para
determinar el porcentaje de células transfectadas)

El tratamiento con tricostatina A, previo a la exposición a etopósido,
ellipticina, camptotecina, doxorubicina, cisplatino, 5-fluorouracilo o
ciclofosfamida, incrementa la muerte celular producida por estas
drogas (Kim et al. 2003).
Terapia cromatínica empleando
inhibidores de deacetilasas de histonas
(HDAC)
Datos preliminares obtenidos en nuestro laboratorio mostraron una
disminución en la sobrevida de células 253J (línea celular derivada
de un cáncer transicional de vejiga) expuestas a etopósido (5, 10 y 20
mM) en presencia de butirato de sodio (un inhibidor de deacetilasas
de histonas).
Recuento de aberraciones cromosómicas y porcentaje de células
apoptóticas

COULD THE INCREASE IN GENETIC DAMAGE OR
DECREASE IN DNA REPAIR BE RESPONSIBLE FOR
HDACi SENSITIZATION?
We studied the induction/removal of DNA damage
induced by anti-neoplasic agents, as well as cell cycle
progression in the presence or absence of HDACi.

Human and Chinese hamster cells were exposed to theHDACis sodium butyrate (NaB) or valproic acid (VA) beforeetoposide treatment or gamma rays exposure.
METHODOLOGY
DNA damage induction/removalComet assay – alkaline version
Cell survival
Clonogenic assay
Bladder cancer (253J) cell line
Chinese hamster (CHOK1) cell line
Sodium butyrate (NaB)(1 and 5 mM)
24 hours
Etoposide
( 20 mM, 1 hour)
Cell cycle progression
DNA content analysisby flow cytometry
G0-G1
S
G2-M
Nº
of
even
ts
Fluorescence
intensity

RESULTS and DISCUSSION
NaB (1 mM and 5 mM) + etoposide (20 µM) in CHOK1 and the 253J cell lines
Cell cycle arrest induced by the higher dose of NaB could explain the unevensensitivities to etoposide in terms of cell survival?
Cell survival
Figure 1 – Relative plate efficiency in 253J (a) and CHOK1 (b) cell lines after etoposide treatment, either with or without NaB pre-treatment
(1 and 5 mM). Pool data from at least two independent experiments are shown. Bars represent 95% confidence intervals.
CHOK1
0%
20%
40%
60%
80%
100%
120%
0 10 20 30 40 50
Etoposide (uM)
Rela
tive p
late
eff
icie
ncy
Eto
NaB 1 mM + Eto
NaB 5 mM +Eto
253J
0%
20%
40%
60%
80%
100%
0 10 20 30 40 50 60
Etoposide (uM)
Rela
tive p
late
eff
icie
ncy
NaB 5mM + Eto
Eto
NaB 1mM + Eto
253J CHOK1
a b

Cell cycle progression
Since cell cycle arrest produced by the higher dose of NaB could be involved inthe decrease of the sensitization effect to etoposide, we analysed its effect onDNA damage induction/removal.
Figure 2 – DNA content analysis evaluated by flow cytometry in 253J (a) and CHOK1 (b) cells treated with
etoposide with and without NaB (N) (1 or 5 mM) pre-treatment.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Control Eto NaB 1 NaB 5 N 1 + Eto N 5 + Eto
% p
hases o
f cell
cyc
le
253J% G2
%S
% G1
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Control Eto NaB 1 NaB 5 N 1 + Eto N 5 + Eto
% p
ha
ses
of c
ell
cycl
e
CHO K1% G2
%S
% G1
a
253J CHOK1
b
G0-G1
S
G2-
M
Nº
of
even
ts
Fluorescence
intensity

DNA damage induction
Therefore, in CHOK1 the higher dose of NaB applied in combination with etoposideproduced less DNA damage which was correlated with the higher cell survivalobserved.
Figure 3 – DNA damage induced by etoposide
(20 µM) with or without NaB (1 or 5 mM) pre-
treatment measured by comet assay (Tail
moment) in two bladder cancer cell lines: a)
253J, b) T24 and c) in hamster cell line CHOK1.
Pool data +/- SEM from at least two independent
experiments are shown.
253J
0
2
4
6
8
10
12
14
16
18
20
Control NaB 1 mM NaB 5 mM Etoposide NaB 1 mM +
Eto
NaB 5 mM +
Eto
Treatment
Tai
l Mom
ent
CHOK1
0
2
4
6
8
10
12
Control NaB 1 mM NaB 5 mM Etoposide NaB 1
mM+Eto
NaB 5
mM+EtoTreatment
Tai
l Mom
ent
T24
0
2
4
6
8
10
12
14
16
18
20
Control NaB 1 mM NaB 5 mM Etoposide NaB 1 mM +
Eto
NaB 5 mM +
Eto
Treatment
Tai
l Mom
ent
b
c
a
253J T24
CHOK1
*
*
*

DNA damage removal in human peripheral bloodlymphocytes H2AX foci inmunodetection
DNA damage induction/removal
(H2AX foci)
Valproic acid (VA) (0,7 mM)
24 hours
rays(0,5 Gy)
0
2
4
6
8
10
12
0 30 60 90 120 150 180 210Time (minutes)
Mean n
um
ber
of
H2A
X f
oci per
cell
0,5 Gy
VA+0,5 Gy
0,5 Gy + VA
Figure 5 – Mean number of -H2AX foci per cell with or without
valproic acid (0,7 mM) pre- (VA+0,5 Gy) or post-treatment (0,5 Gy
+ VA) at 30 minutes and 3 hours after 0,5 Gy irradiation. Pool
data from two independent experiments are shown. Bars represent
SD.

• Depending on the dose of HDACi applied, HDAC inhibition could leadto a higher sensitivity to anti-cancer agents.
CONCLUSIONS
• The sensitizing effect of NaB to anti-neoplasic agents could beexplained by an increase in genetic damage, which seems to beinversely related to cell cycle arrest induced by high doses ofHDACis.
•Depending on the cell type, the dose of HDACi should be carefullyadjusted when they are used as sensitizers to anticancer therapy inorder to increase treatment efficacy.

Desarrollo de terapias antineoplásicas sensibilizadoras
para células tumorales hipóxicas
Hipoxia tumoral
Los tumores sólidos se encuentran formados por una masa celular que
como resultado de la insuficiente neovascularización y las alteraciones de
la vasculatura tumoral resultan en un flujo sanguíneo muy irregular y
caótico, que lleva a la hipoxia tumoral reversible o aguda.
La hipoxia está confinada en el tumor sólido y por lo tanto representa un
blanco único que puede ser explotado para eliminar selectivamente las
células tumorales en los tumores sólidos.

Linker
Agrupamiento hidrofóbico
Grupo quelante de Zn+2
iHDACs
Estos compuestos tienen en común la
presencia de tres agrupamientos
esenciales para la interacción de los
mismos con las HDACs, que son un
grupo quelante de Zn+2 (i.e. ácido
carboxílico o ácido hidroxámico) una
cadena alifática como linker y un
agrupamiento hidrofóbico apolar (i.e.
agrupamiento fenilo).
El pre-tratamiento de algunas líneas celulares tumorales con inhibidores de
deacetilasas de histonas (iHDAC) produce un aumento en la sensibilidad a
agentes quimioterápicos

1) Determinación de la afinidad de los
compuestos por HDAC7 por estudio teórico
de docking para diseñar la síntesis de los
compuestos.
2) Síntesis de los compuestos.
3) Determinación de la LD50 de los compuestos
en las líneas celulares de cáncer en
condiciones de normoxia e hipoxia.
4) Determinación de la capacidad de actuar como
iHDACs de los compuestos a estudiar en
condiciones de normoxia e hipoxia. Efecto de
los iHDACs en la expresión de HDAC7 y HIF-1
en condiciones de hipoxia.
N
N
O
O
NH
N
NH
NHNH2
Agrupamiento quelante de Zinc
Agrupamientos quelantes de Zinc
NOH
NOH
R
LINKER
NH
C
O
LINKERSNH
S
OO
En condiciones de hipoxia, HIF-1 activa la transcripción de genes relacionados
con la supervivencia celular. Las HDACs del grupo II, 4, 6 y 7, modulan la
actividad de HIF-1 regulando la acetilación del mismo, de forma que su
inhibición generaría una disminución en la transcripción de estos genes.
De esta forma las HDACs 4, 6 y 7 constituyen un nuevo factor de regulación de
la transcripción de genes relacionados con la supervivencia del fenotipo
hipóxico. De esta manera el desarrollo de inhibidores de HDACs es un atractivo
blanco terapéutico para el tratamiento de células tumorales hipóxicas.
Objetivos del proyecto

PEDECIBA (Program for BasicSciences Development)
National Agency for Research andInnovation (ANII)
National Commission AgainstCancer (CHLCC – Uruguay)
ACKNOWLEDGEMENTS
IILA Fellowship - University ofTuscia (Italy) in Prof. Palitti’s lab.
Marie-Curie Program – EuropeanCommunity
IAEA Fellowship – Institute forRadioprotection and NuclearSecurity (IRSN), Fontenay AuxRoses (France) in Dr. Voisin’s lab.
Mag. Leticia Méndez Acuña
Dr. María Laura Lavaggi

Muchas gracias !!!
INSTITUTO DE INVESTIGACIONES
BIOLOGICAS CLEMENTE ESTABLE
MONTEVIDEO - URUGUAY
