synergism between curdlan and gm-csf confers … journal of immunology synergism between curdlan and...
TRANSCRIPT

of April 13, 2017.This information is current as
Dendritic CellsConfers a Strong Inflammatory Signature to Synergism between Curdlan and GM-CSF
Christiane RuedlSiu Kwan Sze, Ottavio Beretta, Alessandra Mortellaro and Lin Min, Siti Aminah Bte Mohammad Isa, Wee Nih Fam,
ol.1101755http://www.jimmunol.org/content/early/2012/01/16/jimmun
published online 16 January 2012J Immunol
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2012 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Synergism between Curdlan and GM-CSF Confers a StrongInflammatory Signature to Dendritic Cells
Lin Min,* Siti Aminah Bte Mohammad Isa ,* Wee Nih Fam,* Siu Kwan Sze,*
Ottavio Beretta,† Alessandra Mortellaro,† and Christiane Ruedl*
A simultaneous engagement of different pathogen recognition receptors provides a tailor-made adaptive immunity for an efficient
defense against distinct pathogens. For example, cross-talk of TLR and C-type lectin signaling effectively shapes distinct gene ex-
pression patterns by integrating the signals at the level of NF-kB. In this study, we extend this principle to a strong synergism
between the dectin-1 agonist curdlan and an inflammatory growth factor, GM-CSF. Both together act in synergy in inducing
a strong inflammatory signature that converts immature dendritic cells (DCs) to potent effector DCs. A variety of cytokines (IL-
1b, IL-6, TNF-a, IL-2, and IL-12p70), costimulatory molecules (CD80, CD86, CD40, and CD70), chemokines (CXCL1, CXCL2,
CXCL3, CCL12, CCL17), as well as receptors and molecules involved in fugal recognition and immunity such as Mincle, dectin-1,
dectin-2, and pentraxin 3 are strongly upregulated in DC treated simultaneously with curdlan and GM-CSF. The synergistic effect
of both stimuli resulted in strong IkBa phosphorylation, its rapid degradation, and enhanced nuclear translocation of all NF-kB
subunits. We further identified MAPK ERK as one possible integration site of both signals, because its phosphorylation was
clearly augmented when curdlan was coapplied with GM-CSF. Our data demonstrate that the immunomodulatory activity of
curdlan requires an additional signal provided by GM-CSF to successfully initiate a robust b-glucan–specific cytokine and
chemokine response. The integration of both signals clearly prime and tailor a more effective innate and adaptive response
against invading microbes and fungi. The Journal of Immunology, 2012, 188: 000–000.
Dendritic cells (DCs) are the key regulators of immuneresponses linking innate and adaptive components of theimmune system. As sentinels of the mammalian immune
system, they are among the first to detect, capture, and processinvading infectious agents for the scrutiny of T cells (1). Hence, theoptimal collaboration of DCs and T cells is absolutely crucial forsuccessful immunity.The hallmark of a DC life is a complex maturation process,
which involves the upregulation of MHC and costimulatorymolecules and secretion of several proinflammatory cytokines.This phenotypic shift dramatically increases their immunostimu-latory potential, rendering them the most potent APCs of theimmune system (2). Therefore, mature DCs can be considered assupervaccines that are indeed being developed for this purpose tofight against infections and tumors.A variety of stimuli control and trigger efficiently DC matura-
tion. Pathogen-induced DC activation is mediated via TLR-dependent as well as TLR-independent pathways (3–5). Matura-tion is also promoted by proinflammatory cytokines, endogenous
danger signals, chemokines/chemokine receptor triggering, as wellas by feedback signals from cells of the innate and acquired im-mune system (2). Distinct combinations of various stimuli willdifferentially affect DC migratory behavior, their capacity to ini-tiate T cell activation, and, ultimately, T cell polarization (6).Recently, we identified GM-CSF as a potent DC activator (7),
providing a new face for an old factor with a well-known support-ing role in DC development and recruitment (8). Signaling throughthe GM-CSFR in ex vivo-purified DCs upregulates the expressionof costimulatory molecules more efficiently than any tested TLRagonists alone. Combined with a variety of microbial stimuli, GM-CSF supports the formation of potent effector DCs capable in se-creting a variety of proinflammatory cytokines that guide the dif-ferentiation of T cells during the immune response (7).In this study, we analyzed in particular the effect of GM-CSF
in combination with curdlan on the maturation process of DCs.Curdlan, a linear nonionic homopolymer of D-glucose with (1–3)glucisidic linkages obtained from Alcaligenes faecalis, hasgained attention from the pharmaceutical industry due to its im-munomodulatory function as well as its suitability as a proteindrug delivery vehicle (9, 10). Its receptor, dectin-1 (or Clec7a),a member of the C-type lectin family, is expressed on macro-phages and myeloid-related DCs and, when engaged, controlsfungal infections (11) by activating several intracellular signalingpathways, like NF-kB, MAPK, and NFAT (5). Independently ofdectin-1’s physiological role in antifungal response, DCs stim-ulated through dectin-1 influence many arms of the adaptive im-munity, including Th CD4+ T cells, cytotoxic CD8+ T cells, andB cells (12, 13). Dectin-1 signaling in human monocyte-derivedDCs triggers NF-kB activation through two independent sig-naling pathways, one through Syk and the second through thekinase Raf-1. Both signaling pathways converge at the level ofNF-kB activation and regulation to control antifungal adaptiveimmunity (14).
*School of Biological Sciences, Nanyang Technological University, Singapore 637551;and †Singapore Immunology Network, A*Star, Singapore 138648
Received for publication June 14, 2011. Accepted for publication December 14,2011.
This work was supported by an Academic Research Fund Tier 1 Grant RG 48/10 (toC.R.).
The microarray data presented in this article have been deposited in the GeneExpression Omnibus (http://www.ncbi.nlm.nih.gov/geo/) under accession numberGSE32986.
Address correspondence and reprint requests to Prof. Christiane Ruedl, School ofBiological Sciences, Nanyang Technological University, 60 Nanyang Drive, Singa-pore 637551. E-mail address: [email protected]
Abbreviations used in this article: BM, bone marrow; DC, dendritic cell; FLT3L,FLT3 ligand; PRR, pattern recognition receptor.
Copyright� 2012 by The American Association of Immunologists, Inc. 0022-1767/12/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1101755
Published January 16, 2012, doi:10.4049/jimmunol.1101755 by guest on A
pril 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from

Our transcriptional profiling revealed robust synergistic effectbetween curdlan and GM-CSF. In fact, the integration of bothsignals strongly affected the DC maturation program, resulting inan increased upregulation of several costimulatory molecules, ina potent proinflammatory cytokine and chemokine release as wellas in a superior capacity in polarizing Th cells. How curdlan andGM-CSF signaling pathways integrate to synergistically enhanceproinflammatory cytokine secretion is still elusive. In this study, weshow that both signaling pathways, when initiated simultaneously,result in enhanced IkBa degradation and increased NF-kB nucleartranslocation. We furthermore identified the MAPK kinase ERKas one of the possible targets activated upon GM-CSF and curdlancostimulation. These results show that GM-CSF intensifies andsustains the curdlan-mediated DC activation and that the inte-gration of both signals allows a more effective response to po-tential invading pathogens than each ligand alone.
Materials and MethodsMice
C57BL/6 mice were obtained from The Jackson Laboratory (Bar Harbor,ME), bred, and maintained in the animal facility of the Nanyang Tech-nological University under specific pathogen-free conditions. All animalexperiments were carried out within institutional guidelines.
Reagents
Curdlan was purchased from Wako and dissolved in 0.15 M NaOH ata concentration of 10 mg/ml and further diluted in the culture medium to thefinal required concentration (300 to 1 mg/ml). ATP was purchased fromInvivoGen (San Diego, CA). The following Abs were used for Westernblotting: anti–IL-1b Ab (R&D Systems), anti-NLRP3 (Alexis), anti–caspase-1 (Santa Cruz Biotechnology), and anti–caspase-11 (BioLegend,San Diego, CA). IL-1b, IL-12p70, IL-2, TNF-a, and IL-6 ELISAs, rGM-CSF, anti–IL-17, anti–IFN-g, anti–IL-22, and all secondary Abs werepurchased from BioLegend.
Generation of bone marrow-derived DCs and bone marrow-derived macrophages
FLT3 ligand (FLT3L)-derived bone marrow (BM) DCs were generatedby incubating freshly prepared BM cells for 8 d in IMDM medium sup-plemented with 100 ng/ml FLT3L, respectively. CD11b+ cells were thenpurified by magnetic separation (Miltenyi Biotec, Bergisch Gladbach,Germany). Macrophages were generated from BM cells using L929 cell-conditioned medium (30%).
RNA preparation
CD11b+ fractions of FLT3L-generated BM DCs (3 to 4 3 106) werestimulated for 4 h with 100 or 1 mg/ml curdlan in presence or absence of5 ng/ml GM-CSF. One untreated control group and a GM-CSF–stimulatedgroup were included (total of six groups). Three independent samples foreach group were prepared. Total RNA was extracted using the Pure LinkRNA Isolation Kit (Invitrogen) and used for cRNA preparation accordingto the manufacturer’s instructions (Affymetrix, Santa Clara, CA).
Array hybridization and analysis
Biotinylated cRNA was hybridized to the Affymetrix GeneChip MouseGenome 430 2.0 arrays (Affymetrix). Microarray analysis was performedusing R/Bioconductor (15). Probe set intensity signals were obtained bya GC robust multiarray average background adjustment, normalized bya quantile method (16), and, to discard the genes with the lowest vari-ability, filtered by an interquartile method with an interquartile range of0.25. A statistically significant set of differentially expressed genes wasdefined by both a linear model (17), setting the adjusted p value of the falsediscovery rate to 0.05 (18), and a |log2(Ratio)| . 2 among each comparedconditions, respectively, represented on the y- and x-axis in a Volcano plot.To describe the overall response of each treatment versus untreated DCs,a hierarchical clustering was performed through the MultiExperimentViewer software (19). Using an Euclidean distance as a dissimilaritymeasure and an average linkage as linkage method, each heat map’s col-ored bar represents the log2(Ratio) between each signal and the mediansignal of the probe set. Microarray data have been deposited to GeneExpression Omnibus with the accession number GSE32986 for unstimu-
lated, GM-CSF, curdlan (1 and 100 mg/ml), and curdlan/GM-CSF–stim-ulated FLT3L-derived BM DCs, respectively (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE32986).
Flow cytometry analysis
Cells were distributed into 96-well plates and stimulated overnight witha different concentration of curdlan in the presence or absence of GM-CSF(5 ng/ml). Cells were washed and double stained with allophycocyanin-labeled anti-CD11b and PE-labeled anti-CD80, anti-CD86, anti-CD40,anti-CD70, and anti–dectin-1, respectively. Mean fluorescence intensitywas assessed by flow cytometry (FACSCalibur; BD Biosciences).
In vitro Th polarization
Naive T cells were isolated by magnetic cell separation following themanufacturer’s instruction (Miltenyi Biotec). A total of 105 cells wereplated into a 96-well anti-CD3/CD28 precoated wells together with 2 3104 DCs, which were 24 h stimulated with curdlan alone or in the presenceof GM-CSF and 1 ng/ml TGF-b. Four days later, cell supernatant wasanalyzed for released IL-17, IL-22, and IFN-g by ELISA. Cells werecollected, stained with anti-CD4, subsequently fixed with 2% parafor-maldehyde, and permeabilized with 0.5% saponine. Cells were thenstained with anti–IL-17, anti–IFN-g, or anti–IL-22 and analyzed by flowcytometry (FACSCalibur; BD Biosciences).
Cytokine measurements
Cells were distributed into 96-well plates and stimulated overnight withcurdlan in absence or in absence of GM-CSF. The next day, cell supernatantswere analyzed for IL-12, IL-6, TNF-a, IL-1b, and IL-2 by ELISA fol-lowing the manufacturer’s instructions. In the case of Il-1b, an additional1-h incubation with 5 mM ATP was included.
Detection of Syk and MEK 1/2 phosphorylation
Phosphorylated Syk was measured by flow cytometry after 5 min stimu-lation, respectively. Untreated CD11c+CD11b+ FLT3L-generated DCs orDCs treated with 5 ng/ml GM-CSF, 100 mg/ml curdlan, or a curdlan/GM-CSF mix were fixed and permeabilized with 0.1% saponine solution. Cellswere stained with rabbit anti–phospho-Syk (Tyr525/526; Cell SignalingTechnology) followed by goat anti-rabbit FITC (Southern BiotechnologyAssociates). MEK1/2 cells were permeabilized with cold 100% methanolfor 10 min and stained for 30 min with anti–phospho-MEK1/2 (Ser221; CellSignaling Technology).
Intracellular detection of pro–IL-1b, NLRP3, caspase-1, andcaspase-11
CD11b+ FLT3L-derived DCs incubated overnight untreated or with 5 ng/ml GM-CSF, 100 mg/ml curdlan, or the combination of 5 ng/ml GM-CSFand 100 mg/ml curdlan, respectively. For detection of the pro–IL-1b form,caspase-1, caspase-11, and NLRP3, cells were lysed in buffer containing1% Triton X-100 supplemented with complete protease inhibitor mixture(Roche, Basel, Switzerland). Lysates were resolved by SDS-PAGE andtransferred onto nitrocellulose membranes. The blots were developed bychemiluminescence according to the manufacturer’s instructions (Perkin-Elmer, Waltham, MA).
Western blot
For biochemical analysis, purified CD11c+CD11b+ FLT3L-derived DCs(3 to 4 3 106) were seeded in 24-well plates and cultured overnightin FLT3L-containing medium before stimulation with curdlan (100 mg/ml)and GM-CSF (5 ng/ml) alone or in combination. Because biochemicaldetection of MEK phosphorylation demands a large number of cells, L929-derived BM macrophage cells were used instead of purified CD11b+ DCs.Like CD11b+ DCs, L929-derived BM macrophages express GM-CSFR aswell as dectin-1. Upon treatment (5 min for STAT5 and JAK2, 15 min forMEK1/2, 20 min for ERK, p38, and JNK, and 2 h for IkBa), cells werelysed for 10 min in a buffer containing 50 mM Tris (pH 7.5), 0.27 MSucrose, 0.1 mM EGTA, 1 mM sodium orthovanadate, 50 mM sodiumfluoride, 5 mM sodium pyrophosphate, Roche protease inhibitor mixture,0.1% 2-ME, and 1% Triton X-100. Cell extracts were quantified by pro-tein assay (Bio-Rad). Fixed amounts of total protein (20 mg for non-phosphorylated proteins or 40 mg for phosphorylated proteins) were re-solved by SDS-PAGE and electroblotted to nitrocellulose membranes.The following Abs were used to visualize total or phosphorylated IkBa,STAT5, JAK2, MEK1/2, p38, ERK, and JNK: mouse anti-tubulin, goatanti-ERK, and rabbit anti–phospho-ERK (Santa Cruz Biotechnology);rabbit anti–IkB-a, mouse anti–phospho-IkBa, rabbit anti-JNK, rabbit anti-
2 SYNERGY BETWEEN GM-CSFR AND DECTIN-1
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

p38, rabbit anti–phospho-JAK2, rabbit anti–phospho-STAT5, rabbit anti–phospho-p38, rabbit anti-MEK1/2, and anti–phospho-MEK1/2 (Cell Sig-naling Technology); and rabbit anti–phospho-JNK (Invitrogen). The blotswere developed by chemiluminescence as described above.
Nuclear translocation of NF-kB subunits
To measure nuclear NF-kB subunit p65, p50, p52, RelB, and c-Rel levels,FLT3L-derived CD11b+ DCs were kept untreated or stimulated for 20 hwith 5 ng/ml GM-CSF, 100 mg/ml curdlan, or the combination of 5 ng/mlGM-CSF and 100 mg/ml curdlan, respectively. NF-kB DNA binding ac-tivity of p65, p50, p52, and RelB was determined using a DNA-bindingELISA kit (TransAM NF-kB transcription factor kit; Active Motif,Carlsbad, CA) using equal amounts of nuclear extracts. c-Rel and USF-2as an internal standard were monitored by Western blotting.
MEK1/2 inhibition
A total of 5 3 105 purified FLT3L-derived CD11b+ DCs were seeded into96-well flat-bottom plates in 200 ml complete medium and pretreated with10 and 1 mM MEK1/2 inhibitor U0126 (Sigma-Aldrich). One hour later,different stimuli and their combinations were added to the cells (5 ng/mlGM-CSF, 100 mg/ml curdlan, or combination of curdlan/GM-CSF). Afterovernight incubation, release of TNF-a, IL-1b, and IL-6 was measured byELISA. In the case of IL-1b, cells were additionally stimulated for 1 hwith ATP before the collection of the supernatant.
ResultsGene expression profile in FLT3L-derived CD11b+ DCs uponstimulation with curdlan alone or in combination withGM-CSF
DNA microarray analysis was performed to investigate the dif-ferential effect of curdlan versus GM-CSF and their combinationson DC physiology. At first, we analyzed the gene expression profileinduced at early time points upon DC stimulation. Therefore,purified FLT3L-derived CD11b+ DCs were processed for analysisafter a short 4-h stimulation with two concentrations of curdlan (1and 100 mg/ml), alone or in combination with GM-CSF (5 ng/ml).As shown in the Volcano plot, the obtained gene expressionanalysis revealed clear differences in the transcriptome upondistinct treatments (Fig. 1). A large number of genes were up-regulated by .2-fold; in particular, 100 mg/ml curdlan combinedwith GM-CSF modulated the largest number (871), followed by1 mg/ml curdlan combined with GM-CSF (571). When 100 and1 mg/ml curdlan were tested alone, clearly smaller number of geneswas upregulated (394 and 63, respectively), whereas GM-CSFtreatment alone revealed an upregulation of 400 genes. Clearly,GM-CSF signaling had strong synergistic and diversifying effecton transcriptome induced in conventional FLT3L-derived BMDCs by curdlan.A summary of genes differentially expressed or modulated in
maturing DC compared with nonstimulated cells are listed in Table
I. The highest upregulated genes (.10-fold) in the early phase ofDC activation in response to the combination of curdlan/GM-CSF include several proinflammatory cytokines (e.g., IL-1a, IL-1b,TNF-a, IL-6, IL-12p35, and IL-2), many chemokines (e.g.,CXCL2, CCL17, CXCL3, CCL12), and several costimulatorymolecules (e.g., CD80, CD86, CD40, and CD70). Furthermore,a series of C-type lectins, such as Clec5a (MDL-1), Clec4e(Mincle), and Clec4n (dectin-2), recently identified as signalingfungal pattern recognition receptors (PRRs), were differentiallymodulated. Furthermore, pentraxin 3, a potent antifungal mole-cule, was also differently expressed in DCs in response to thecombination GM-CSF and curdlan. Furthermore, our results showa GM-CSF–mediated increase of TLR2 (Table I), a PRR describedto collaborate with dectin-1 for optimal induction of proin-flammatory cytokine and chemokine response (20, 21). Theseresults clearly demonstrate that curdlan and GM-CSF can potentlyact in synergy inducing in DCs an inflammatory signature, whichconverts immature DCs to potent effector DCs.
Curdlan and GM-CSF enhance the expression of costimulatorymolecules and synergistically boost secretion ofproinflammatory cytokines in conventional CD11c+CD11b+
DCs
Purified FLT3L-derived CD11c+CD11b+ DCs were analyzed byflow cytometry for expression pattern of CD80, CD86, CD40, andCD70 after 24 h stimulation with 100 mg/ml curdlan alone or incombination with GM-CSF. Curdlan, GM-CSF, and the combi-nation of both stimuli augmented the expression of all four co-stimulatory receptors when compared with the expression level onunstimulated DCs. Confirming the gene array data, the combina-tion curdlan/GM-CSF was the most potent stimulus as shown inFig. 2A.We next focused on the validation of the upregulated cytokines
such as IL-12p70, IL-6, TNF-a, and IL-2. Despite the fact thatGM-CSF was capable in upregulating the expression of costim-ulatory molecules on DCs, the growth factor alone was unable toinduce subsequent proinflammatory cytokine release. In fact, IL-12p70, TNF-a, IL-6, or IL-2 could not be detected as shown inFig. 2B. In contrast, GM-CSF combined with curdlan synergisti-cally augmented the production of IL-12p70, TNF-a, and IL-2when compared with the secretion induced by curdlan alone.IL-6 secretion was only slightly affected by the curdlan/GM-CSFcombination at this late stimulation time point of 48 h (Fig. 2B).These results show that GM-CSF intensifies and sustains the
curdlan-mediated DC activation and that the integration of bothsignals allows a more effective response to potential invadingpathogens than each ligand alone.
FIGURE 1. Affymetrix GeneChip platform (Affy-
metrix) was used for global transcriptional analysis in
DCs in response to different concentrations of curdlan
(1 and 100 mg/ml), GM-CSF, and their combinations
with respect to unstimulated DCs. Four hours upon
stimulation, mRNA of triplicates was processed and
analyzed. Volcano plots of obtained microarray data of
the different experimental groups. On x-axis, the log2
(Ratio) of stimulated DCs versus control DCs (unsti-
mulated), in which green and red lines represent the
threshold for |log2(Ratio)|.2. On y-axis, the 2log2(p
value) resulting from the comparison of the two con-
ditions, in which the blue line represents the threshold
for log2(p) . 2. In summary, the x-axis represents an
intensity measure of the treatment, whereas the y-axis
estimates the reliability of this measure.
The Journal of Immunology 3
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

GM-CSF synergizes curdlan-induced pro–IL-1b synthesis butdoes not affect the inflammasome components
An efficient IL-1b secretion depends on an NF-kB–dependentsynthesis of pro–IL-1b followed by a required danger signal, likeATP, which mediates the activation of the caspase-1 and subse-quently the cytokine release. To clarify the role of GM-CSF in IL-1b synthesis and secretion, DCs were stimulated for 2, 4, and 6 hwith curdlan in the absence or in the presence of GM-CSF and IL-1b was assessed at the level of mRNA. As already observed in thetranscriptome analysis (Table I), both low (1 mg/ml) and high (100mg/ml) curdlan concentrations combined with GM-CSF resultedin the strongest increase of IL-1b mRNA when compared withcurdlan alone (Fig. 3A). Also at the intracellular protein level,Western blot analysis showed five times higher amounts of thepro–IL-1b form in cells stimulated with the combination of cur-dlan and GM-CSF as compared with cells treated with curdlanalone (Fig. 3B). Together with pro–IL-1b, we measured the con-tent of the inflammasome components, such as NLRP3, caspase-1,and caspase-11 in untreated and differently stimulated cells. Weobserved that caspase-1 protein levels were similar in all testedconditions, whereas NLRP3 and caspase-11 levels were increasedupon stimulation, but without any obvious difference betweencurdlan and curdlan/GM-CSF treatments. To test the effect ofGM-CSF on the IL-1b production in a danger situation, westimulated FLTL3-derived DC in the presence of ATP with cur-dlan alone or in combination with GM-CSF. Single stimuli (GM-CSF or curdlan) did not induce any significant cytokine release,but their combination dramatically enhanced in a dose-dependentmanner the secretion of IL-1b (Fig. 3C). We ruled out the role ofGM-CSF as a danger signal itself because it always needs topartner with ATP to boosts the IL-1b release (data not shown).The absolute requirement of ATP as a danger signal suggests thatdectin-1 engagement alone in FLT3L-derived myeloid DCs isunable to trigger a strong reactive oxygen species formation (22),
which was described to directly activate the NRLP3 inflamma-some in macrophages (23).Taken together, these results indicate that DCs require GM-CSF
for increased pro–IL-1b synthesis in response to curdlan.
Curdlan/GM-CSF–activated DCs strongly support Th1 andTh17 polarization
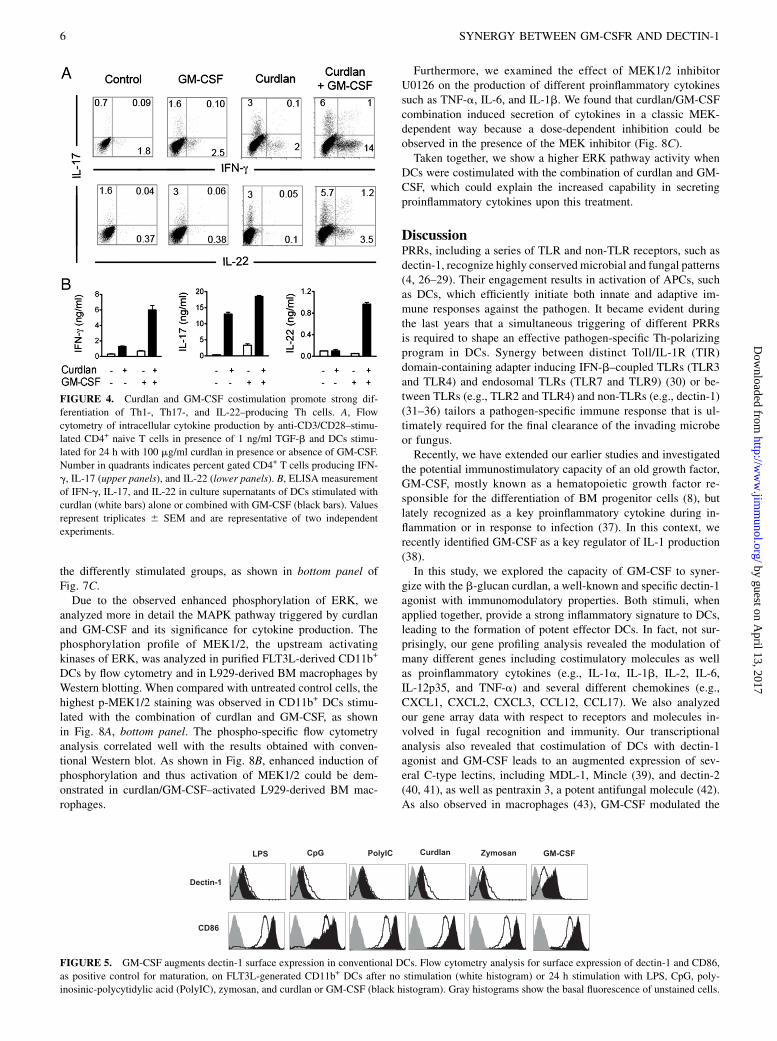
Recent in vivo and in vitro experiments have shown that dectin-1agonists, including curdlan, promote Th1 as well as Th17 immu-nity (12). Therefore, we studied the role of GM-CSF in supportingthe formation of these two different types of Th subsets. Follow-ing an Ag-independent polarization protocol, we stimulated naiveT cells on anti-CD3/CD28 plates in the presence or absence of DCsactivated via curdlan and GM-CSF, alone or in combination. Whenanalyzed by flow cytometry, curdlan/GM-CSF–activated DCs weremore potent in polarizing both Th1 as well as Th17 Th subsets whencompared with DCs that were stimulated with curdlan alone (Fig.4A, upper panel). Interestingly, the combination of stimuli enhancedalso the formation of IL-17+/IL-22+ double-producing Th cells aswell as a single secreting IL-22+ subset (24, 25) as shown in Fig. 4A,lower panel. The enhanced production of IFN-g, IL-17, and IL-22was not only observed at a single-cell level by intracellular staining,but was also confirmed by ELISA (Fig. 4B).These data suggest that the observed enhanced production of
proinflammatory cytokines upon costimulation of curdlan andGM-CSF by DCs generally support a stronger polarization on Thcells. However, as demonstrated by our gene array, the inability ofcurdlan/GM-CSF to prime p19 mRNA synthesis as well as toinduce secretion of bioactive IL-23 in conventional myeloid DCs(data not shown) indicate that this type of DCs does prime but notsustain the further Th17 differentiation.
GM-CSF augments dectin-1 surface expression in DCs
Next, we analyzed whether the basal level of the known curdlan-specific receptor dectin-1 could be further augmented by different
Table I. Overview of a selection of different genes strongly upregulated by the combination GM-CSFand 100 or 1 mg/ml curdlan, respectively
Genes GM-CSF AloneCurdlan Alone
(1 mg/ml)Curdlan Alone(100 mg/ml)
GM-CSF +Curdlan(1 mg/ml)
GM-CSF +Curdlan
(100 mg/ml)
ChemokinesCXCL1 (MSGA-a) 0 12 44 22 75CXCL2 (MIP-2 a) 3 41 140 100 321CXCL3 (MIP-2 b) 1 4 25 14 93CCL12 (MCP-1) 100 0 4 110 112CCL17 (MCP-3) 65 2 15 94 242
CytokinesIL-1b 10 13 40 81 84IL-1a 0 2 7 10 38IL-2 0 0 0 2 20TNF-a 0 0 23 18 49IL-6 2 2 6 10 60IL-12p35 0 0 2 0 8
Costimulatory receptorsCD40 3 3 13 9 21CD70 0 0 5 0 10CD80 7 0 3 14 18CD86 2 0 3 2 5
Fungal-related molecules,TLR, and C-type lectinsMincle (Clec4e) 0 5 42 10 50MDL-1 (Clec5a) 0 0 2 0 4Dectin-2 (Clec4n) 3 0 2 4 6Pentraxin 3 0 0 6 5 33TLR2 3 4 9 6 9
Fold induction is represented.
4 SYNERGY BETWEEN GM-CSFR AND DECTIN-1
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

TLR and non-TLR agonists or by GM-CSF itself. LPS, CpG,polyinosinic-polycytidylic acid, zymosan, curdlan, and GM-CSFwere tested on FLT3L-derived CD11c+CD11b+ DCs. As expected,all stimuli induced maturation of DCs visualized by the upregu-lation of CD86; however, dectin-1 was highly upregulated only byGM-CSF treatment, as shown in Fig. 5.
Synergy between curdlan and GM-CSF results in strong IkBaphosphorylation, its rapid degradation, and nucleartranslocation of different NF-kB subunits
We next analyzed the effect of our different stimuli on the phos-phorylation and degradation of IkBa by stimulating FLT3L-generated CD11b+ DCs for 2 h with curdlan and GM-CSF aloneor in combination. As shown in Fig. 6A, costimulation with bothstimuli resulted in a slight enhanced IkBa phosphorylation andIkBa degradation when compared with cells stimulated withcurdlan or GM-CSF alone. Furthermore, we observed that co-stimulation of DCs supported augmented nuclear translocation ofdifferent NF-kB subunits. In fact, p50, p52, p65, RelB, as well asc-Rel were clearly detectable in nuclear extracts of DCs cotreatedwith curdlan and GM-CSF (Fig. 6B), results that are consistentwith the observed increased proinflammatory cytokine and che-mokine response.
Analysis of GM-CSF– and curdlan-induced intracellularsignaling pathways
We next sought to clarify the integration point of curdlan and GM-CSF intracellular signals, which leads to this NF-kB activation and
as a consequence to a stronger cytokine and chemokine secretion.First, we analyzed the signaling pathway induced by engagementof the GM-CSFR. An equal phosphorylation pattern of STAT5 andJAK2 could be observed in the DCs treated with GM-CSF alone orin combination with curdlan (Fig. 7A), whereas the unstimulatedcells as well as curdlan-stimulated cells showed none. We nextfocused on the dectin-1–mediated NF-kB canonical pathwaycomprising the kinase Syk as one the most upstream elements.Both curdlan and curdlan/GM-CSF induced rapid and equal Sykphosphorylation in DCs, whereas control and GM-CSF–activatedcells did not (Fig. 7B), which excludes Syk as an integration pointof both signals. Finally, we analyzed the phosphorylation patternof the MAPKs p38, ERK, and JNK because these kinases areknown to regulate expression of genes involved in the innate im-mune response such as proinflammatory cytokines. Stimulationof CD11b+ FLT3L-derived DCs with GM-CSF–induced phos-phorylation of two MAPKs, JNK and ERK, whereas no increaseabove the basal level of phosphorylation was observed in curdlan-treated DCs. In the case of JNK, the phosphorylation pattern couldnot be further augmented by cotreatment with curdlan/GM-CSF,whereas this was the case for ERK (Fig. 7C). No apparent dif-ferences in the phosphorylation of p38 was observed between
FIGURE 2. Curdlan and GM-CSF enhances the expression of costim-
ulatory molecules and synergistically boosts secretion of proinflammatory
cytokines. A, FLT3L-generated CD11b+ DCs were cultured for 24 h with
100 mg/ml curdlan in absence (white bars) or presence of 5 ng/ml GM-
CSF (black bars). CD86, CD80, CD40, and CD70 upregulation was vi-
sualized by FACS analysis. B, FLT3L-generated CD11c+CD11b+ DCs
were stimulated with different curdlan concentrations ranging from 300 to
1 mg/ml in absence (white circles) or presence of 5 ng/ml GM-CSF (black
circles). IL-12p70, TNF-a, IL-6 (48-h stimulation), and IL-2 (24-h
stimulation) were measured by ELISA. Results show nanograms per
milliliter released cytokines and are representative of three independent
experiments.
FIGURE 3. Pro–IL-1b synthesis is synergistically augmented upon
curdlan and GM-CSF costimulation. FLT3L-generated CD11c+CD11b+
DCs were cultured for 2, 4, 6, and 24 h with 100 or 1 mg/ml curdlan in
absence (white bars) or presence of 5 ng/ml GM-CSF (black bars). A, IL-
1b gene expression was assessed by semiquantitative RT-PCR analysis
after 2, 4, and 6 h upon stimulation. All samples were normalized with the
internal control gene HPRT. B, Western blot analysis of pro–IL-1b,
NLRP3, caspase-1, and caspase-11 in FLT3L-derived CD11b+ DCs. Cells
were left untreated or stimulated overnight with 100 mg/ml curdlan, 5 ng/
ml GM-CSF, or a combination of both, and cell lysates were subsequently
prepared and separated on a 10% SDS-PAGE gel. C, IL-1b ELISA after
stimulation with different curdlan concentrations in absence (white circles)
or in presence of GM-CSF (black circles). Additional 1-h incubation with
5 mM ATP was included after 24 h stimulation. Measured cytokines of
triplets are represented in picograms per milliliter, and results are repre-
sentative of two separate experiments.
The Journal of Immunology 5
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from
the differently stimulated groups, as shown in bottom panel ofFig. 7C.Due to the observed enhanced phosphorylation of ERK, we
analyzed more in detail the MAPK pathway triggered by curdlanand GM-CSF and its significance for cytokine production. Thephosphorylation profile of MEK1/2, the upstream activatingkinases of ERK, was analyzed in purified FLT3L-derived CD11b+
DCs by flow cytometry and in L929-derived BM macrophages byWestern blotting. When compared with untreated control cells, thehighest p-MEK1/2 staining was observed in CD11b+ DCs stimu-lated with the combination of curdlan and GM-CSF, as shownin Fig. 8A, bottom panel. The phospho-specific flow cytometryanalysis correlated well with the results obtained with conven-tional Western blot. As shown in Fig. 8B, enhanced induction ofphosphorylation and thus activation of MEK1/2 could be dem-onstrated in curdlan/GM-CSF–activated L929-derived BM mac-rophages.
Furthermore, we examined the effect of MEK1/2 inhibitorU0126 on the production of different proinflammatory cytokinessuch as TNF-a, IL-6, and IL-1b. We found that curdlan/GM-CSFcombination induced secretion of cytokines in a classic MEK-dependent way because a dose-dependent inhibition could beobserved in the presence of the MEK inhibitor (Fig. 8C).Taken together, we show a higher ERK pathway activity when
DCs were costimulated with the combination of curdlan and GM-CSF, which could explain the increased capability in secretingproinflammatory cytokines upon this treatment.
DiscussionPRRs, including a series of TLR and non-TLR receptors, such asdectin-1, recognize highly conserved microbial and fungal patterns(4, 26–29). Their engagement results in activation of APCs, suchas DCs, which efficiently initiate both innate and adaptive im-mune responses against the pathogen. It became evident duringthe last years that a simultaneous triggering of different PRRsis required to shape an effective pathogen-specific Th-polarizingprogram in DCs. Synergy between distinct Toll/IL-1R (TIR)domain-containing adapter inducing IFN-b–coupled TLRs (TLR3and TLR4) and endosomal TLRs (TLR7 and TLR9) (30) or be-tween TLRs (e.g., TLR2 and TLR4) and non-TLRs (e.g., dectin-1)(31–36) tailors a pathogen-specific immune response that is ul-timately required for the final clearance of the invading microbeor fungus.Recently, we have extended our earlier studies and investigated
the potential immunostimulatory capacity of an old growth factor,GM-CSF, mostly known as a hematopoietic growth factor re-sponsible for the differentiation of BM progenitor cells (8), butlately recognized as a key proinflammatory cytokine during in-flammation or in response to infection (37). In this context, werecently identified GM-CSF as a key regulator of IL-1 production(38).In this study, we explored the capacity of GM-CSF to syner-
gize with the b-glucan curdlan, a well-known and specific dectin-1agonist with immunomodulatory properties. Both stimuli, whenapplied together, provide a strong inflammatory signature to DCs,leading to the formation of potent effector DCs. In fact, not sur-prisingly, our gene profiling analysis revealed the modulation ofmany different genes including costimulatory molecules as wellas proinflammatory cytokines (e.g., IL-1a, IL-1b, IL-2, IL-6,IL-12p35, and TNF-a) and several different chemokines (e.g.,CXCL1, CXCL2, CXCL3, CCL12, CCL17). We also analyzedour gene array data with respect to receptors and molecules in-volved in fugal recognition and immunity. Our transcriptionalanalysis also revealed that costimulation of DCs with dectin-1agonist and GM-CSF leads to an augmented expression of sev-eral C-type lectins, including MDL-1, Mincle (39), and dectin-2(40, 41), as well as pentraxin 3, a potent antifungal molecule (42).As also observed in macrophages (43), GM-CSF modulated the
FIGURE 4. Curdlan and GM-CSF costimulation promote strong dif-
ferentiation of Th1-, Th17-, and IL-22–producing Th cells. A, Flow
cytometry of intracellular cytokine production by anti-CD3/CD28–stimu-
lated CD4+ naive T cells in presence of 1 ng/ml TGF-b and DCs stimu-
lated for 24 h with 100 mg/ml curdlan in presence or absence of GM-CSF.
Number in quadrants indicates percent gated CD4+ T cells producing IFN-
g, IL-17 (upper panels), and IL-22 (lower panels). B, ELISA measurement
of IFN-g, IL-17, and IL-22 in culture supernatants of DCs stimulated with
curdlan (white bars) alone or combined with GM-CSF (black bars). Values
represent triplicates 6 SEM and are representative of two independent
experiments.
FIGURE 5. GM-CSF augments dectin-1 surface expression in conventional DCs. Flow cytometry analysis for surface expression of dectin-1 and CD86,
as positive control for maturation, on FLT3L-generated CD11b+ DCs after no stimulation (white histogram) or 24 h stimulation with LPS, CpG, poly-
inosinic-polycytidylic acid (PolyIC), zymosan, and curdlan or GM-CSF (black histogram). Gray histograms show the basal fluorescence of unstained cells.
6 SYNERGY BETWEEN GM-CSFR AND DECTIN-1
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

dectin-1 as well as TLR2 expression in FLT3L-derived DCs, in-creasing the capability for fungal recognition and ability to initiatean effective antifungal response.This synergistic DC stimulation results in the generation of
strong effector DCs with augmented capacity in priming effi-ciently Th1-, Th17-, and IL-22–secreting CD4+ Th cells and inshaping a highly specialized antifungal immune response. Theseresults are in accordance with a previous work of Rosas et al.(44), which demonstrated the crucial effect of GM-CSF in cel-
lular programming of a proinflammatory dectin-1–mediated re-sponse in macrophages. Using both curdlan and glucan micro-particles, the authors showed that macrophages do not mounta significant proinflammatory cytokine response upon dectin-1engagement, which can be dramatically boosted via GM-CSFcostimulation (44). They speculated about a GM-CSF–medi-ated alteration of the downstream signaling components of theSyk/ CARD9 and NF-kB pathways, but the exact mechanism ofaction was not clarified. Goodridge et al. (45) also recently re-ported the capacity of GM-CSF as well as of IFN-g to efficientlyamplify dectin-1–CARD9-induced TNF-a production in BM-derived macrophages. In the same study, they observed thatdistinct myeloid cells differentially signal through the dectin-1–CARD9–Bcl10 to induce NF-kB activation and TNF-a produc-tion. Although dectin-1 signals via CARD9 and BCL10 in GM-CSF–derived BM DCs, this pathway does not activate NF-kB
FIGURE 6. Enhanced IkBa phosphorylation, IkBa degradation, and
NF-kB nuclear translocation upon costimulation of conventional DCs with
curdlan/GM-CSF combination. A, FLT3L-generated CD11b+ DCs were
stimulated with GM-CSF, curdlan, and their combination for 2 h and as-
sayed by Western blot for IkBa phosphorylation and IkBa degradation.
Tubulin was used as internal control to standardize the samples. B, Nuclear
recruitment of NF-kB subunits in FLT3L-derived CD11b+ DCs was ana-
lyzed 24 h after stimulation by DNA-binding ELISA (p50, p52, RelB, and
p65) and Western blot (c-Rel). USF-2 was used as internal control to
standardize the samples. Values represent triplicates 6 SEM and were
confirmed in two independent experiments.
FIGURE 7. Curdlan/GM-CSF costimulation augments activation of
ERK. A, Western blot analysis of activated phosphorylated forms of JAK2
and STAT5. Two different curdlan concentrations were tested, referred to
as low (1 mg/ml) and high (100 mg/ml), respectively. Tubulin was used as
internal control to standardize the samples. B, Flow cytometry analysis of
Syk activation upon different combinations of stimulation. Cells were fixed
and permeabilized 30 min upon treatment and intracellular staining with
rabbit anti–phospho-Syk and FITC-labeled goat anti-rabbit, respectively.
C, Western blot analysis of activated phosphorylated forms of MAPK p38,
ERK, and JNK. After 20 min stimulation, cells were lysed and immuno-
blotted for active p38, ERK, and JNK and total p38, ERK, and JNK as
internal controls. The results are representative of at least two independent
experiments.
The Journal of Immunology 7
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

and subsequent TNF-a release in BM-derived macrophages aswell as in FLT3L-derived DCs. In our study, we confirm theinability of FLT3L-derived DCs to secrete high amounts ofproinflammatory cytokines when stimulated with curdlan alone,and we further demonstrate that these cells require GM-CSF toefficiently activate the NF-kB pathway and induce the subse-quent proinflammatory cytokine response as previously reportedfor the BM-derived macrophages (45).In this work, to elucidate the molecular mechanisms underlying
this observed synergistic effect between the two different stimuli,we focused on the analysis of known signaling pathways initiatedupon the curdlan-specific receptor dectin-1 as well as GM-CSFRengagement. No cross-talk between these two different receptorshas been described so far. It is well documented that GM-CSFactivates the Jak2/STAT5, Ras/Raf/MAPK, and PI3K/Akt path-ways through its heterodimeric receptor composed by a majorbinding unit, GM-CSFRa, and a major signaling unit, GM-CSFRbc
(8, 46, 47). Curdlan initiates via dectin-1 binding a NF-kB ca-nonical pathway comprising the kinase Syk and the adapter pro-teins Card9, Bcl10, and Malt1 (12, 48) as well as an NF-kBnoncanonical Raf-1–dependent signaling pathway (14). In humanimmature DCs, both pathways converge at the level of NF-kBactivation by translocating NF-kB subunits c-Rel and p65 in thecase of the canonical pathway or the RelB-p52 subunit in the caseof the noncanonical pathway (14). We found that curdlan com-bined with GM-CSF enhanced dramatically the nuclear translo-
cation of all NF-kB subunits tested, including p50, p52, p65,RelB, and c-Rel.We next focused on JAK-2/STAT5 and, in particular, on the
MAPK signaling pathway because the latest was described to betriggered by GM-CSF as well as by dectin-1 agonists (12, 49–51).As expected, we could not see any differences in the GM-CSF–induced STAT5/JAK2 pathway regardless the presence or ab-sence of curdlan. In the case of the canonical pathway of dectin-1 signaling, we also could not observe any difference of Sykphosphorylation when GM-CSF was coapplied to curdlan.However, one of the MAPKs, ERK, showed a clearly augmentedphosphorylation pattern in response to both stimuli when com-pared in response to single stimuli. The same enhanced phos-phorylation was observed for the upstream activating kinasesMEK1/2. In contrast, the phosphorylation of other two MAPKs,JNK and p38, were not further enhanced by the combination ofthe stimuli. In disagreement with other published data (12, 51),we could not detect any significant ERK, JNK, and p38 phos-phorylation differences upon stimulation of CD11b+ FLT3L-derived DCs with curdlan alone. Indeed, the observed MAPKphosphorylation in previous studies might be explained by theuse of different type of DCs in those biochemical experiments.In fact, curdlan-dependent MAPK phosphorylation could be ob-tained only when GM-CSF–derived BM DCs were used and/orGM-CSF was included in the culture medium during the stimu-lation period (12, 51). In our experiment, we used naive conven-tional CD11b+ FLT3L-derived DCs that express GM-CSFR andare reactive against the growth factor, which was not used for theirderivation.ERK signaling cascade is not only restricted to the regulation
of cellular processes, such as proliferation, differentiation, and on-cogenic differentiation, but also in recent years, it has become clearthat it also controls proinflammatory cytokine production. ERK,once phosphorylated, can directly activate transcription factorsupon translocation into the nucleus or, in contrast, phosphorylatea variety of cytoplasmic targets. Several MAPK kinase kinases,such as MEKK1, MEKK2, and MEKK3, have been identified toactivate NF-kB through IkBa (52–54), which we show in thisstudy is strongly phosphorylated and downregulated upon co-stimulation of curdlan and GM-CSF.Furthermore, several studies have recently reported about the
contribution of ERK in initiating the production of proinflam-matory cytokines upon dectin-1 engagement. For example, zy-mosan mediates a Syk-dependent ERK production of IL-2 andIL-10 in GM-CSF–derived DCs. This suggests that not only theknown canonical pathway through Syk/Card9/NF-KB, but alsoan independent pathway through Syk/ERK is controlling the ex-pression of these cytokines (50). Furthermore, Brereton et al. (55)demonstrated that ERK activation via dectin-1 is also controllingIL-23 and IL-1b production because inhibition of MEK1/2, thekinase upstream of ERK, suppressed both cytokines and, as aconsequence, reduced the ability of DCs to promote Th1 and Th17responses. In this study, we further show that inhibition of MEK1/2 suppressed clearly IL-1b as well as TNF-a and IL-6 uponcurdlan and GM-CSF costimulation.In summary, recognition of fungal pathogens involves a co-
ordinated interplay of multiple receptors including several TLRsas well as C-type lectins. In this study, we have shown thata growth factor, GM-CSF, can provide in the presence of curdlana potent inflammatory signature to conventional DCs, whichensures a robust cytokine and chemokine response and subse-quent Th cell polarization. Our results strongly suggest thatinflammation caused by fungal invasion is requisite for primingand tailoring an effective host innate and adaptive response. Our
FIGURE 8. Curdlan/GM-CSF–mediated proinflammatory cytokine
release is regulated by the MAPK/ERK pathway. A, FLT3L-derived
DCs were stained with allophycocyanin-labeled anti-CD11b and stimu-
lated with 5 ng/ml GM-CSF, 100 mg/ml curdlan, or combination of cur-
dlan/GM-CSF, respectively. After 20 min, cells were processed, stained
with anti–phospho-MEK1/2, and analyzed by flow cytometry. Control
unstimulated CD11b+ cells are represented as gray filled histogram,
whereas the stimulated cells are represented with open histograms. B,
L929-derived BM macrophages were stimulated with 5 ng/ml GM-CSF,
100 mg/ml curdlan, or combination of curdlan/GM-CSF. Cell lysate was
prepared and analyzed by Western blotting with Abs against total MEK1/2
and against the Ser221-phosphorylated form of MEK1/2. C, Inhibition of
MEK1/2 in FLT3L CD11b+-derived DCs suppresses TNF-a, IL-6, and
IL-1b secretion. Purified FLT3L-derived CD11b+ DCs were pretreated
with 10 and 1 mM MEK1/2 inhibitor U0126. One hour later, different
stimuli and their combinations were added for 16 h to the cells (without
stimulation [white circles], 5 ng/ml GM-CSF [black circles], 100 mg/ml
curdlan [white squares], or combination of curdlan/GM-CSF [black
squares]). The release of TNF-a, IL-1b, and IL-6 was measured by
ELISA. Each point represents the mean of three samples, and error bars
represent SDs. The results were confirmed in two independent experi-
ments.
8 SYNERGY BETWEEN GM-CSFR AND DECTIN-1
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

findings might be relevant for the design of novel adjuvantformulations, which are more effective in priming a strong anti-fungal inflammatory response.
AcknowledgmentsWe thank Dr. Klaus Karjalainen for critical reading of the manuscript and
Dr. Peter Cheung for valuable technical advice. We also thank Josephine
Lum and Yolanda Aphrilia Setiagani for technical support.
DisclosuresThe authors have no financial conflicts of interest.
References1. Banchereau, J., and R. M. Steinman. 1998. Dendritic cells and the control of
immunity. Nature 392: 245–252.2. Macagno, A., G. Napolitani, A. Lanzavecchia, and F. Sallusto. 2007. Duration,
combination and timing: the signal integration model of dendritic cell activation.Trends Immunol. 28: 227–233.
3. Kumar, H., T. Kawai, and S. Akira. 2009. Pathogen recognition in the innateimmune response. Biochem. J. 420: 1–16.
4. Kawai, T., and S. Akira. 2010. The role of pattern-recognition receptors in innateimmunity: update on Toll-like receptors. Nat. Immunol. 11: 373–384.
5. Osorio, F., and C. Reis e Sousa. 2011. Myeloid C-type lectin receptors inpathogen recognition and host defense. Immunity 34: 651–664.
6. Kawai, T., and S. Akira. 2011. Toll-like receptors and their crosstalk with otherinnate receptors in infection and immunity. Immunity 34: 637–650.
7. Min, L., S. A. Mohammad Isa, W. Shuai, C. B. Piang, F. W. Nih, M. Kotaka, andC. Ruedl. 2010. Cutting edge: Granulocyte-macrophage colony-stimulatingfactor is the major CD8+ T cell-derived licensing factor for dendritic cell acti-vation. J. Immunol. 184: 4625–4629.
8. Hamilton, J. A., and G. P. Anderson. 2004. GM-CSF Biology. Growth Factors22: 225–231.
9. Castro, G. R., B. Panilaitis, E. Bora, and D. L. Kaplan. 2007. Controlled releasebiopolymers for enhancing the immune response. Mol. Pharm. 4: 33–46.
10. Mocanu, G., D. Mihai, M. Moscovici, L. Picton, and D. LeCerf. 2009. Curdlanmicrospheres. Synthesis, characterization and interaction with proteins(enzymes, vaccines). Int. J. Biol. Macromol. 44: 215–221.
11. Taylor, P. R., S. V. Tsoni, J. A. Willment, K. M. Dennehy, M. Rosas, H. Findon,K. Haynes, C. Steele, M. Botto, S. Gordon, and G. D. Brown. 2007. Dectin-1 isrequired for beta-glucan recognition and control of fungal infection. Nat.Immunol. 8: 31–38.
12. LeibundGut-Landmann, S., O. Gross, M. J. Robinson, F. Osorio, E. C. Slack,S. V. Tsoni, E. Schweighoffer, V. Tybulewicz, G. D. Brown, J. Ruland, and C. Reise Sousa. 2007. Syk- and CARD9-dependent coupling of innate immunity to theinduction of T helper cells that produce interleukin 17. Nat. Immunol. 8: 630–638.
13. Leibundgut-Landmann, S., F. Osorio, G. D. Brown, and C. Reis e Sousa. 2008.Stimulation of dendritic cells via the dectin-1/Syk pathway allows priming ofcytotoxic T-cell responses. Blood 112: 4971–4980.
14. Gringhuis, S. I., J. den Dunnen, M. Litjens, M. van der Vlist, B. Wevers,S. C. Bruijns, and T. B. Geijtenbeek. 2009. Dectin-1 directs T helper cell dif-ferentiation by controlling noncanonical NF-kappaB activation through Raf-1and Syk. Nat. Immunol. 10: 203–213.
15. Gentleman, R. C., V. J. Carey, D. M. Bates, B. Bolstad, M. Dettling, S. Dudoit,B. Ellis, L. Gautier, Y. Ge, J. Gentry, et al. 2004. Bioconductor: open softwaredevelopment for computational biology and bioinformatics. Genome Biol. 5: R80.
16. Bolstad, B. M., R. A. Irizarry, M. Astrand, and T. P. Speed. 2003. A comparisonof normalization methods for high density oligonucleotide array data based onvariance and bias. Bioinformatics 19: 185–193.
17. Smyth, G. K. 2004. Linear models and empirical bayes methods for assessingdifferential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol.3: Article3.
18. Reiner, A., D. Yekutieli, and Y. Benjamini. 2003. Identifying differentiallyexpressed genes using false discovery rate controlling procedures. Bio-informatics 19: 368–375.
19. Saeed, A. I., N. K. Bhagabati, J. C. Braisted, W. Liang, V. Sharov, E. A. Howe,J. Li, M. Thiagarajan, J. A. White, and J. Quackenbush. 2006. TM4 microarraysoftware suite. Methods Enzymol. 411: 134–193.
20. Brown, G. D. 2006. Dectin-1: a signalling non-TLR pattern-recognition receptor.Nat. Rev. Immunol. 6: 33–43.
21. Dennehy, K. M., and G. D. Brown. 2007. The role of the beta-glucan receptorDectin-1 in control of fungal infection. J. Leukoc. Biol. 82: 253–258.
22. Underhill, D. M., E. Rossnagle, C. A. Lowell, and R. M. Simmons. 2005. Dectin-1 activates Syk tyrosine kinase in a dynamic subset of macrophages for reactiveoxygen production. Blood 106: 2543–2550.
23. Gross, O., H. Poeck, M. Bscheider, C. Dostert, N. Hannesschlager, S. Endres,G. Hartmann, A. Tardivel, E. Schweighoffer, V. Tybulewicz, et al. 2009. Sykkinase signalling couples to the Nlrp3 inflammasome for anti-fungal host de-fence. Nature 459: 433–436.
24. Fujita, H., K. E. Nograles, T. Kikuchi, J. Gonzalez, J. A. Carucci, and J. G.Krueger.2009. Human Langerhans cells induce distinct IL-22-producing CD4+ T cellslacking IL-17 production. Proc. Natl. Acad. Sci. USA 106: 21795–21800.
25. Zenewicz, L. A., and R. A. Flavell. 2011. Recent advances in IL-22 biology. Int.Immunol. 23: 159–163.
26. Brown, G. D., and S. Gordon. 2003. Fungal beta-glucans and mammalian im-munity. Immunity 19: 311–315.
27. Iwasaki, A., and R. Medzhitov. 2004. Toll-like receptor control of the adaptiveimmune responses. Nat. Immunol. 5: 987–995.
28. Akira, S., and K. Takeda. 2004. Toll-like receptor signalling. Nat. Rev. Immunol.4: 499–511.
29. Creagh, E. M., and L. A. O’Neill. 2006. TLRs, NLRs and RLRs: a trinity ofpathogen sensors that co-operate in innate immunity. Trends Immunol. 27: 352–357.
30. Napolitani, G., A. Rinaldi, F. Bertoni, F. Sallusto, and A. Lanzavecchia. 2005.Selected Toll-like receptor agonist combinations synergistically trigger a Thelper type 1-polarizing program in dendritic cells. Nat. Immunol. 6: 769–776.
31. Gantner, B. N., R. M. Simmons, S. J. Canavera, S. Akira, and D. M. Underhill.2003. Collaborative induction of inflammatory responses by dectin-1 and Toll-like receptor 2. J. Exp. Med. 197: 1107–1117.
32. Yadav, M., and J. S. Schorey. 2006. The beta-glucan receptor dectin-1 functionstogether with TLR2 to mediate macrophage activation by mycobacteria. Blood108: 3168–3175.
33. Netea, M. G., N. A. Gow, C. A. Munro, S. Bates, C. Collins, G. Ferwerda,R. P. Hobson, G. Bertram, H. B. Hughes, T. Jansen, et al. 2006. Immune sensingof Candida albicans requires cooperative recognition of mannans and glucans bylectin and Toll-like receptors. J. Clin. Invest. 116: 1642–1650.
34. Goodridge, H. S., and D. M. Underhill. 2008. Fungal recognition by TLR2 andDectin-1. Handb. Exp. Pharmacol. 183: 87–109.
35. Ferwerda, G., F. Meyer-Wentrup, B. J. Kullberg, M. G. Netea, and G. J. Adema.2008. Dectin-1 synergizes with TLR2 and TLR4 for cytokine production inhuman primary monocytes and macrophages. Cell. Microbiol. 10: 2058–2066.
36. Dennehy, K. M., G. Ferwerda, I. Faro-Trindade, E. Pyz, J. A. Willment,P. R. Taylor, A. Kerrigan, S. V. Tsoni, S. Gordon, F. Meyer-Wentrup, et al. 2008.Syk kinase is required for collaborative cytokine production induced throughDectin-1 and Toll-like receptors. Eur. J. Immunol. 38: 500–506.
37. Hamilton, J. A. 2002. GM-CSF in inflammation and autoimmunity. TrendsImmunol. 23: 403–408.
38. Khameneh, H. J., S. A. Isa, L. Min, F. W. Nih, and C. Ruedl. 2011. GM-CSFsignalling boosts dramatically IL-1 production. PLoS ONE 6: e23025.
39. Willment, J. A., and G. D. Brown. 2008. C-type lectin receptors in antifungalimmunity. Trends Microbiol. 16: 27–32.
40. Robinson, M. J., F. Osorio, M. Rosas, R. P. Freitas, E. Schweighoffer, O. Gross,J. S. Verbeek, J. Ruland, V. Tybulewicz, G. D. Brown, et al. 2009. Dectin-2 isa Syk-coupled pattern recognition receptor crucial for Th17 responses to fungalinfection. J. Exp. Med. 206: 2037–2051.
41. Saijo, S., S. Ikeda, K. Yamabe, S. Kakuta, H. Ishigame, A. Akitsu, N. Fujikado,T. Kusaka, S. Kubo, S. H. Chung, et al. 2010. Dectin-2 recognition of alpha-mannans and induction of Th17 cell differentiation is essential for host defenseagainst Candida albicans. Immunity 32: 681–691.
42. Garlanda, C., E. Hirsch, S. Bozza, A. Salustri, M. De Acetis, R. Nota,A. Maccagno, F. Riva, B. Bottazzi, G. Peri, et al. 2002. Non-redundant role ofthe long pentraxin PTX3 in anti-fungal innate immune response. Nature 420:182–186.
43. Willment, J. A., H. H. Lin, D. M. Reid, P. R. Taylor, D. L. Williams, S. Y. Wong,S. Gordon, and G. D. Brown. 2003. Dectin-1 expression and function are en-hanced on alternatively activated and GM-CSF-treated macrophages and arenegatively regulated by IL-10, dexamethasone, and lipopolysaccharide. J.Immunol. 171: 4569–4573.
44. Rosas, M., K. Liddiard, M. Kimberg, I. Faro-Trindade, J. U. McDonald,D. L. Williams, G. D. Brown, and P. R. Taylor. 2008. The induction of in-flammation by dectin-1 in vivo is dependent on myeloid cell programming andthe progression of phagocytosis. J. Immunol. 181: 3549–3557.
45. Goodridge, H. S., T. Shimada, A. J. Wolf, Y. M. Hsu, C. A. Becker, X. Lin, andD. M. Underhill. 2009. Differential use of CARD9 by dectin-1 in macrophagesand dendritic cells. J. Immunol. 182: 1146–1154.
46. Hansen, G., T. R. Hercus, B. J. McClure, F. C. Stomski, M. Dottore, J. Powell,H. Ramshaw, J. M. Woodcock, Y. Xu, M. Guthridge, et al. 2008. The structure ofthe GM-CSF receptor complex reveals a distinct mode of cytokine receptoractivation. Cell 134: 496–507.
47. Hercus, T. R., D. Thomas, M. A. Guthridge, P. G. Ekert, J. King-Scott,M. W. Parker, and A. F. Lopez. 2009. The granulocyte-macrophage colony-stimulating factor receptor: linking its structure to cell signaling and its role indisease. Blood 114: 1289–1298.
48. Gross, O., A. Gewies, K. Finger, M. Schafer, T. Sparwasser, C. Peschel,I. Forster, and J. Ruland. 2006. Card9 controls a non-TLR signalling pathway forinnate anti-fungal immunity. Nature 442: 651–656.
49. Suzuki, K., M. Hino, F. Hato, N. Tatsumi, and S. Kitagawa. 1999. Cytokine-specific activation of distinct mitogen-activated protein kinase subtype cascadesin human neutrophils stimulated by granulocyte colony-stimulating factor,granulocyte-macrophage colony-stimulating factor, and tumor necrosis factor-alpha. Blood 93: 341–349.
50. Slack, E. C., M. J. Robinson, P. Hernanz-Falcon, G. D. Brown, D. L. Williams,E. Schweighoffer, V. L. Tybulewicz, and C. Reis e Sousa. 2007. Syk-dependentERK activation regulates IL-2 and IL-10 production by DC stimulated withzymosan. Eur. J. Immunol. 37: 1600–1612.
51. Hernanz-Falcon, P., O. Joffre, D. L. Williams, and C. Reis e Sousa. 2009. In-ternalization of Dectin-1 terminates induction of inflammatory responses. Eur. J.Immunol. 39: 507–513.
The Journal of Immunology 9
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

52. Zhao, Q., and F. S. Lee. 1999. Mitogen-activated protein kinase/ERK kinasekinases 2 and 3 activate nuclear factor-kappaB through IkappaB kinase-alphaand IkappaB kinase-beta. J. Biol. Chem. 274: 8355–8358.
53. Lee, F. S., J. Hagler, Z. J. Chen, and T. Maniatis. 1997. Activation of the IkappaBalpha kinase complex by MEKK1, a kinase of the JNK pathway. Cell 88: 213–222.
54. Chen, B. C., and W. W. Lin. 2001. PKC- and ERK-dependent activation of Ikappa B kinase by lipopolysaccharide in macrophages: enhancement by P2Yreceptor-mediated CaMK activation. Br. J. Pharmacol. 134: 1055–1065.
55. Brereton, C. F., C. E. Sutton, S. J. Lalor, E. C. Lavelle, and K. H. Mills. 2009.Inhibition of ERK MAPK suppresses IL-23- and IL-1-driven IL-17 productionand attenuates autoimmune disease. J. Immunol. 183: 1715–1723.
10 SYNERGY BETWEEN GM-CSFR AND DECTIN-1
by guest on April 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from