symposium tropical botany: principles and practice || inflorescence structure in primitive...
TRANSCRIPT

Inflorescence Structure in Primitive AngiospermsAuthor(s): Focko WeberlingSource: Taxon, Vol. 37, No. 3, Symposium Tropical Botany: Principles and Practice (Aug.,1988), pp. 657-690Published by: International Association for Plant Taxonomy (IAPT)Stable URL: http://www.jstor.org/stable/1221107 .
Accessed: 15/09/2013 18:32
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
International Association for Plant Taxonomy (IAPT) is collaborating with JSTOR to digitize, preserve andextend access to Taxon.
http://www.jstor.org
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON 37(3): 657-690. AUGUST 1988
INFLORESCENCE STRUCTURE IN PRIMITIVE ANGIOSPERMS
Focko Weberling'
Summary The inflorescences of Magnoliales exhibit a wide range of diverse forms. Monotelic inflorescences
prevail by far. Panicles or thyrses terminating the flowering shoots are relatively rare, mostly the main axis shows a proliferating growth, then bearing "axillary" inflorescences only. By reduction of the ramification of the flower bearing axis, botryoids or even further reduced inflorescences can be formed. Solitary flowers terminating leafy shoots are also found in several families of the Magnoliales (as well as in Dilleniales). Highly specialized long shoot-short shoot systems are not rare (Calycanthaceae, Schisandraceae, Winteraceae, Eupteleaceae, Tetracentraceae, Cercidiphyllaceae, Eucommiaceae, My- risticaceae), and even cauliflory occurs. Progression to an inflorescence of the polytelic type is relatively rare and occurs especially in some genera of the Lauraceae and apparently generally in Hernandiaceae and probably in members of Myristicaceae.
The question, how inflorescences may have evolved and what might be or might have been the primitive form of an inflorescence has been answered in different ways. Since we have been dealing with the morphology of inflorescences in angiosperms for many years we were often asked this question (Weberling, 1982, 1983a) and thus we may discuss it here with special reference to the so-called primitive angiosperms. In our terminology we follow Troll (1964/1969), Troll and Weber (1957), and Weberling (1982, 1983b).
Leaving aside the old assumption of de Candolle (1827), that flowers are axillary branches, we find mainly two different opinions, which were expressed by Parkin (1914) and Pilger (1921). The suggestions of Parkin can be traced back to St. Hilaire (1840, p. 276). We may quote them from his summary (Parkin, 1914, pp. 559-560) without going too far into details: "From a comparative study it seems highly probable that flowers were originally borne on the plant singly, each terminal to a leafy shoot ...." "From such a shoot (or shoots) ... all inflorescences, as well as the solitary axillary flower have probably arisen." "The first flower-cluster ... is formed by two or more axils below the terminal flower pushing out lateral floral shoots." Looking for evidence to confirm his assumption Parkin (1914, p. 521) asked: "Do plants still exist exhibiting this early mode of flower-bearing?" and then stated, that "the most conspicuous examples of solitary terminal flowers occur in those families-e.g., the Magnoliaceae, Ranunculaceae, etc.-which on other grounds are now generally regarded as primitive." Accordingly, the solitary terminal flowers, which we find in Liriodendron, in all species of Magnolia and Talauma could be regarded as primitive as well as in certain species of Paeonia (Paeoniaceae), Dillenia indica L., and Hibbertia cuneiformis (Labill.) Gilg (Dilleniaceae) within the Dilleniales, the second nucleus of so-called "primitive" angiosperms. Nevertheless Parkin (1914) bore in mind that solitary terminal flowers may also arise "through the reduction of an inflorescence to a single flower" and that "we must be on our guard in assuming primitiveness in every case." For the cases of Liriodendron, Magnolia, and some similar examples, however, "reduction seems extremely unlikely" to him.
The formation of lateral flowers alongside the terminal one, their increase in number, and the "reduction of all the foliage leaves into bracts" as the successive steps of the evolution of an "apical inflorescence" may be illustrated by Fig. 1, generalizing a series of examples given by Parkin (1914). The first of his examples was Calycanthus occidentalis
1 Abt. Spezielle Botanik, University of Ulm, Germany.
657
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
x I -- Z " I rv v l
Fig. 1. Diagrams to illustrate the hypothesis of Parkin on inflorescence evolution (I-V), and to explain the formation of proliferating inflorescences (VI) and the differentiation of the shoot system into long shoots and short shoots which either develop synchronously with the long shoot and flower within the same season (VII) or develop and flower in the next season (VIII).
Hook. & Am., in which "lateral floral shoots are produced sometimes from the axils of the uppermost pair of foliage leaves, the three flowers ... thus forming a simple dichasial cyme," which he regarded to be the "first step in the evolution of inflorescences."
If we follow the mostly adopted opinion regarding the flower as a shoot or a terminal part of a shoot bearing leaves which serve for sexual reproduction and which accordingly are transformed, then the view of Parkin (1914) (which was adopted by Takhtajan, 1959, pp. 110-118) seems to be quite convenient at first sight. This view, however, presupposes the flowers as elements already existing at the starting point of inflorescence evolution and being available for the formation of primitive inflorescences. There does not exist much doubt, however, that the first formation of inflorescences, that is to say, of flower-bearing systems, must be intimately connected with the phylogeny of the flower itself. Although the remark of Rickett (1944, p. 215) that "The inflorescence may be older than the flower" appears to be somewhat exaggerated, we must admit that we do not know very much about the successive stages of flower evolution.
It was Pilger (1921, 1922), who pointed out, that the primary aspect must be the vegetative ramification and that the ramification of the inflorescence should be considered as being derived from the mode of branching within the vegetative zone. Thus to him the "leafy panicle" appeared as the "most primitive form of the inflorescence in angiosperms" (Pilger,
658
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
II II I -,
Fig. 2. I, Liriodendron tulipifera L., two shoot generations (1962, 1963) with solitary terminal flowers (orig. W. Troll, unpublished); II, Michelia champaca L. var. alba, branch with uniflorous short shoots.
1922, p. 21). At least we may follow Celakovsky (1892) in regarding the panicle as the most generalized inflorescence, as a central type from which all the other forms of inflo- rescences can be derived. This does not necessarily imply a phylogenetical interpretation.
It may therefore be an idle question to ask for "the primitive inflorescence." This, however, does not imply that we must refrain from any evaluation of inflorescences as relatively primitive or more advanced in their characters. Thus a comparative study of inflorescence-structures in Magnoliales (and Dilleniales) will in any case yield data, which can be useful for systematic conclusions and which might also elucidate evolutionary tendencies, even if the fundamental question, what might have been the primitive inflo- rescence remains unsolved.
The Magnoliaceae have been mentioned already because of the frequent occurrence of solitary terminal flowers. On further consideration we can confirm this for Liriodendron as well as for all the genera of the Magnolieae (Nooteboom, 1985), namely Manglietia BI., Pachylarnax Dandy, Kmeria (Pierre) Dandy, and Magnolia L. with all the genera included into this genus by Nooteboom (1985), as for example, Talauma Juss., Alcimandra Dandy, and Aromadendron Bl. On the other hand the Michelieae with Elmerrillia Dandy and Michelia L. (incl. Paramichelia H. H. Hu and Tsoongiodendron W. Y. Chun) are charac- terized by lateral flowers. As we see from the example of Michelia champaca L. (Fig. 211) these flowers, which originate in the axils of foliage leaves are often preceded by one or two foliage leaves. Thus these uniflorous partial inflorescences may indicate a first step of a transition to a long shoot-short shoot system (macroblast-brachyblast system) as a spe- cialization of the branching system. This interpretation was also given by Nooteboom (1985, p. 74), who even transferred it to the terminal flowers of the Magnolieae: "in the other genera the brachyblast or the pedicle with the flower is terminal." Although this interpretation might be acceptable for short seasonal shoots bearing quite a few foliage leaves or bracts only, it seems doubtful to apply the term brachyblast if these shoots develop a greater number of foliage leaves before they end in a terminal flower, and all the less when these shoots bear the buds for the new shoot generations continuing the branching system, as is shown by the diagram of Liriodendron tulipifera (Fig. 21).
A branching system similar to that of Michelia champaca is known for Degeneria vitiensis
659
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
I II Fig. 3. I-II, Eupomatia laurina R. Br. Previous year's long shoot with uniflorous short shoots
(Weberling 7545, N Queensland, ULM).
Bailey and Smith (1942) with the difference that the lateral axes ending in solitary flowers bear two or three scales. In Austrobaileya scandens C. T. White the number of scales is
greater (Fig. 81). In Idiospermum australiense (Diels) T. S. Blake, now regarded as repre- sentative of a separate family Idiospermaceae (Blake, 1972), the lateral flowers arising from the axils of foliage leaves are preceded by "2-4 distant pairs of... decussate bracts" (Blake, 1972, p. 5), sometimes a secondary flower arising from the axil of one of these bracts can be observed. According to the interpretation of Bailey and Smith (1942, p. 360) "The fact, that the pedicels of Degeneria are bracteate near the middle may indicate that the inflo- rescence is reduced from a more complex structure and that the supposed pedicel is partially peduncular in origin." Whereas in Degeneria and Austrobaileya subtending foliage leaves are still present during anthesis, we always found them fallen off in Eupomatia laurina R. Br. (Figs. 3, 131); they were only present in the distal part of the main axis. The flowers are preceded by about six bracteose leaves in median-distichous position. They are certainly able to protect the flower bud in young stages or in a period of dormancy. We found these flower buds unfolding in axes which had completely lost their foliage leaves. This, however, is not always the case (see also: Curtis's Bot. Mag. 81, t. 4848, 1855; Williams, 1979).
660
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
Fig. 4. Illicium anisatum L. (I. religiosum Siebold & Zucc.): I, flowering branch; II, uniflorous
partial inflorescence. (After K6hler, 1923.)
Eupomatia bennettii F. Muell. usually has solitary flowers terminating leafy shoots with a
great number of foliage leaves (cf. Uphof, 1 959a, p. 175 and fig. 41 A, p. 174). In some cases, however, the number of foliage leaves preceding the terminal flower is very small (2 or 3), whereas on the other hand in E. laurina one or two of the scalelike leaves can be replaced by foliage leaves. Thus we may have here another example of transition towards the formation of a macroblast-brachyblast system.
Uniflorous partial inflorescences, arising from the axils of persistent foliage leaves of a main axis which continues vegetative growth, are also characteristic for the Illiciaceae. Illicium anisatum L. (Fig. 4) may serve as an example. The singular flowers are preceded by a series of scalelike leaves, including the two prophylls (Fig. 411). According to the
description in Curtis's Botanical Magazine (69, t. 3965, 1843) a second flower can arise, probably from the axil of one of the bracts, and the same is reported for many other species (Smith, 1947), where the number of flowers can even increase to three or four as in I. griffithii Hook. f. & Thoms. and I. leiophyllum A. C. Smith. A similar series of scales can be found in other species such as L simonsii Maxim. Not rarely the bracts are confined to the base of the "peduncle" (I. griffithii, I. parviflorum Michx., I. macranthum A. C. Smith, I. manipurense Watt.). Their number is often reduced to four or two as in I. mexicanum
661
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
I II tIl Fig. 5. Flowering branches of (I) Schisandra propinqua (Wall.) Baill., and (II) Kadsura (roxburgh-
iana Am.) heteroclita (Roxb.) Craib. (After King, 1891.)
A. C. Smith and I. floridanum Ellis. In the latter especially the "peduncles" are compar- atively long and as in several other species their subtending foliage leaves form one or more pseudoverticils of three or four leaves toward the apex of the branchlets.
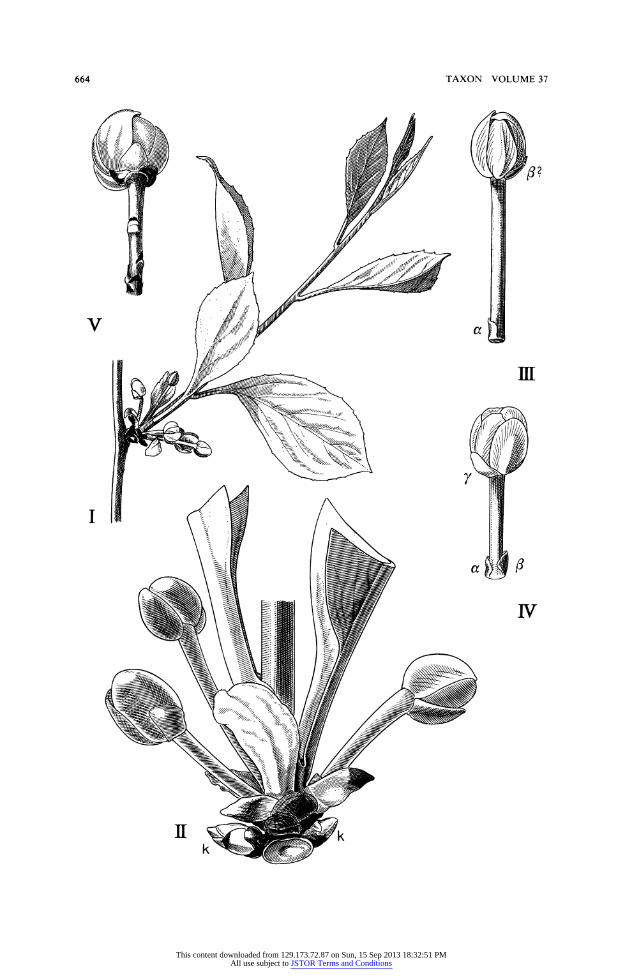
The Schisandraceae is another family well known for solitary flowers in the axils of foliage leaves as is exemplified by Fig. 51 and II for Schisandra and Kadsura, the only two genera of the family. The description of the family given by Smith (1947, p. 82) is somewhat more complicated in this point: "flowers variously arranged, most often solitary and axillary to foliage leaves on ultimate branchlets or in axils of fugacious bracts near the base of ultimate shoots, often paired, sometimes aggregated and glomerulate on ultimate shoots or on main branches, pedicellate, .. . pedicels ebracteolate to several bracteolate ...." In Schisandra axillaris Hook. f. & Thoms. (Fig. 61) and S. propinqua (Wall.) Baill. (Figs. 51, 611) as well as in Kadsura heteroclita (Roxb.) Craib (Fig. 511), K. japonica (L.) Dunal, K. lanceolata King, and other species the axes ending in flowers originate in the axils of foliage leaves of the "ultimate branchlets" (Smith, 1947). As can be seen from Figs. 51 and 6 the foliation of these branchlets starts with numerous scalelike leaves at the very base of the axis, which probably serve as bud scales. In Schisandra chinensis (Turcz.) Baill. (sect. Maximowiczia) the flowers are situated in "axils of fugacious bracts toward base of annual shoots" (Smith, 1947, p. 95), which are equivalent to short shoots on the previous year's long shoots, but often continue their growth, proliferating to long, frondose shoots as is shown in Fig. 71 and II. The two lowermost scales, the prophylls, bear vigorous buds in their axils. Proliferation, however, also can take place on short shoots which remain short, as is the case in S. repanda (Siebold & Zucc.) A. C. Smith. The brachyblasts of this vine bear their flowers in the axils of scalelike leaves which probably serve as bud scales for a while. They are followed by a number of foliage leaves which again are replaced by scales. It can be seen from the leaf scars of older brachyblasts, that this leaf arrangement can be repeated for several vegetation periods. Sometimes, however, these shoots develop into
662
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
I
II
Fig. 6. Diagrams of flower-bearing axillary shoots of (I) Schisandra axillaris Hook. f. & Thoms. (Brinkma 874a, L) and (II) S. propinqua (Wall.) Baill. (D.'Alleizette 67, cult., L.)
663
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions
TAXON VOLUME 37
i
V
I
Il
aI
IV
664
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
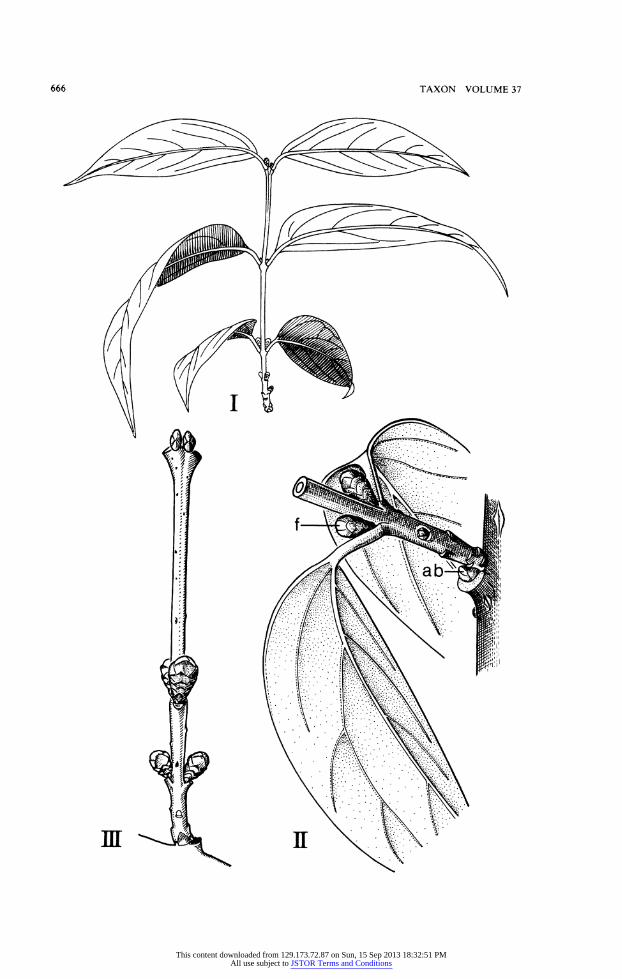
I II Fig. 8. I,Austrobaileyascandens C. T. White (Endress& Gray 4265, M); II, Calycanthusoccidentalis
Hook. & Am., flowering shoot. (II, Orig. W. Troll.)
long shoots. There are also transitions between the species bearing their flowers in the axils of foliage leaves and those where they arise from the axils of scales. Here they develop flowers in the basal parts of the branchlets from the axils of bud scales as well as from the axils of foliage leaves (Schisandra elongata Hook. f. & Thoms.). In S. chinensis the "ped- icels" of the flowers are described as "ebracteolate or with a single obscure bracteole near the base" (Smith, 1947, p. 137). They can even be provided with three bracteoles (Fig. 7IV) two prophylls (a, X) at the base and a third bracteole being integrated into the perianth (-y). The description mentioned before could, however, refer to the case shown in Fig. 7111, but here the second prophyll (/) may appear immediately below the perianth. It has also been found (Troll, notices), that secondary flowers arose from the axils of the two prophylls. The number of bracteoles is often restricted to one or two, but can be increased considerably, as in Schisandra axillaris. The same applies to species of Kadsura, e.g., K. japonica (L.) Dunal (Fig. 7V), K. heteroclita (Fig. 511), and K. hainanensis Merr. An occurrence of more than a single flower, as mentioned before, has been found in specimens of Schisandra axillaris, S. propinqua, and also in Kadsura and is reported for a great number of species of both genera, as in the descriptions of Schisandra sect. Sphaerostema ("occasionally in clusters of 3-5") and of the genus Kadsura ("flowers solitary or rarely paired or rarely 2- 4-glomerulate"), that is to say 2-4-flowered partial inflorescences can occur. In Kadsura
Fig. 7. I-IV, Schisandra chinensis (Turcz.) Baill. I, axillary flowering shoot; II, basis in detail, k prophyllary buds; III, flower with prophyll (s), a, df; and IV, with an additional bract (y); V, Kadsura japonica (L.) Dunal, uniflorous partial inflorescence with bracts. (Orig. W. Troll.)
665
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions
TAXON VOLUME 37
1J/
Ii
666
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
cauliflora B1. the solitary flowers provided with several bracteoles arise on old branches from the axils of leaves which are fallen off long ago.
In Calycanthaceae, Calycanthus occidentalis (Fig. 811) has already been mentioned as the first of Parkin's examples for the formation of a primitive inflorescence by the devel- opment of lateral floral shoots in addition to the solitary terminal flower. At first sight C. floridus L. (Fig. 10IV), the species next to C. occidentalis (cf. Nicely, 1965), seems to be in this way further progressed by the increased number of floral branches. However, the foliage leaves from the axils of which they have arisen are lost during the winter before the flowering time-the lateral floral branches are this years brachyblasts originating from a previous years macroblast. The short shoots are accompanied by phylloscopic accessory buds (inserted between the insertion of the short shoot and the subtending leaf), which in Fig. O1IV have already developed into accessory shoots. It is not rare, that this shrub flowers once more in the late summer. Then the proleptic-anthetic (Miiller-Doblies and Weberling, 1984) flowering branches, although being in the position of lateral brachyblasts, can develop more than ten pairs of foliage leaves. In Chimonanthes praecox (L.) Link the specialization of the uniflorous brachyblasts has still gone further in so far as their foliation is restricted to many pairs of bud scales, whereas the foliage leaves are confined to the first year's macroblasts or younger parts of the older macroblasts (Figs. 9, 10). As in Calycanthus floridus there are accessory buds below the insertion of the branches (Fig. 911 and III).
There can be no doubt, that all the inflorescences of the families mentioned before represent variations of the monotelic type. Even if the growth of the main axis is indefinite as in case of proliferating inflorescences (e.g., Illicium) the lateral flower bearing axes, the paracladia, reveal the monotelic character by ending in a terminal flower and, at least partly, bearing more leaves than the two prophylls only, thus comprising the ability for paniculoid or thyrsoidal branching. [Although the terms 'thyrse' and 'thyrsic' have been in use for indeterminate (open) systems as well as for terminate (closed) inflorescences of this ramification type, we follow a suggestion made by Briggs and Johnson (1979) using the term thyrse (thyrsic) for indeterminate and the term thyrsoid (thyrsoidal) for determinate systems. The noun thus is comparable with the term 'botryoid' (Troll) which designates an inflorescence which resembles a raceme, but ends in a terminal flower.]
In Winteraceae the flowering branches are terminated by the inflorescence in Bubbia, Belliolum, Exospermum, and Zygogynum, now all to be included in Zygogynum (Vink, 1985). The richest inflorescences are doubtless found within the former genus Bubbia. Their ramification is thyrsoid-paniculate. However, since the paracladia of first order are usually arranged in pseudoverticils and their hypopodia are relatively long, the inflorescence could be taken for a double-umbel at first sight (Fig. 1 1IV), and the distal part of the inflorescence has often been interpreted as a "terminal partial inflorescence." A very rich ramification can be found in Zygogynum howeanum (F. Muell.) Vink (Bubbia howeana (F. Muell.) van Tieghem), where the lowermost paracladia can show a thyrsoidal ramification themselves as shown by Fig. 1 lIVa (van Baalgooy 1111, L, according to Vink, 1983); the same can apply to Z. calothyrsum (Diels) Vink and Z. semecarpoides (F. Muell.) Vink (Endress & Gray 4212, L). This high degree of ramification can even be exceeded by the inflorescences of Z. amplexicaule (Parm.) Vink (Vink, pers. comm.). In Z. queenslandianum (Vink) Vink and Z. semecarpoides (F. Muell.) Vink var. whiteanum (A. C. Smith) Vink they are "dis- tinctly reduced" and "in other species again they always consist of solitary flowers, e.g .....
Fig. 9. Chimonanthus praecox (L.) Link. I, leafy long shoot with buds of flower-bearing short shoots (Sept. 1958); II, base of a long shoot with buds of flower-bearing short shoots (f) and accessory bud (ab) below the insertion of the shoot (Sept. 1958); III, long shoot with buds of flower-bearing short shoots (Nov. 1958). (Orig. W. Troll.)
667
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
I
II
m X Fig. 10. I-III, Chimonanthus praecox (L.) Link, buds of uniflorous short shoots at the nodes of a
previous year's long shoot, III immediately before anthesis; IV, Calycanthus floridus L., uniflorous short shoots on a previous year's long shoot; ab = accessory branches. (I-III, Orig. W. Troll; IV, after Baillon, 1869.)
B. pauciflora" (Vink, 1985, p. 43). The inflorescences of Belliolum can have a similar appearance as shown for Bubbia howeana, e.g., the inflorescences of B. crassifolium van Tiegh. (Schlechter 15348, New Caledonia, M), where the lower pseudoverticil comprises six cymes with long hypopodia. In other species (e.g., B. burttianum A. C. Smith) the
668
r
I ..? ... ...
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
I
1 I I IV I IVa Fig. 11. Diagrams of inflorescences of I, Drimys angustifolia Miers (Reitz & Klein 6982, Serra do
Orat6rio, Brasil, M); II, Zygogynum pomiferum Baill., according to data given by Vink; III, Pseu- dowintera axillaris (J. R. & G. Forst.) Dandy, generalized diagram; in I-III the intemodes of the flower- bearing parts which partly remain short are all drawn as elongated; IV, Zygogynum howeanum (F. Muell.) Vink (Hoogland & Craven 11049, TNG, CANB); IVa, lowermost pc of van Balgooy 1111, according to data given by Vink.
inflorescence is less ramified and the flowers within the cymes are somewhat loosely ar- ranged because of the elongation of all subsequent axes. In B. gracile A. C. Smith (Schodde 3718, Bougainville, CANB) the inflorescences are reduced to few-flowered umbel-like botryoids ('sciadioids') as in Zygogynum (Fig. 11II) and Exospermum (cf. Vink, 1977, 1983), and in Zygogynum vinkii Simpson the inflorescence is reduced to its terminal flower (Vink, 1985, p. 43). In all these genera, the continuation of the branching system is achieved by axillary buds "of a higher or the highest cataphyll (bract) below the flowers" and thus is sympodial, "Sometimes two axillary buds develop" (Vink, 1977).
In contrast to the genera described before, Drimys is characterized by the fact that the main axis of the flowering system, instead of ending in a terminal flower, grows out and returns to vegetative growth. This phenomenon is called proliferation, and we have made use of this term already in describing inflorescences like that of Illicium as "proliferating inflorescences." In Drimys the change from reproductive to vegetative development be- comes more obvious because in place of the bracts which subtend the floriferous branches, the paracladia, foliage leaves are formed again (Fig. 121) which also can subtend vegetative
669
,It ^ ^- ^-*^'^^~~~~~~~~~~
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
f , I I I Fig. 12. Drimys winteri J. R. & G. Forst., proliferating inflorescences, in II the terminal leafy bud
is just to be seen in the centre of the inflorescence. (Orig. Troll.)
branches. (For a comparative histology of inflorescence apex and vegetative apex, see Gifford, 1950, 1951, and Tucker, 1959.) In Drimys winteri J. R. & G. Forst. (Fig. 121 and II) the lowest paracladia consist of three (four or even more) lateral flowers and end in a terminal flower, thus revealing the monotelic character of the inflorescence. Since the lateral flowers and their subtending bracts are inserted more or less at the same level an umbel- like shape of the paracladia results; they are sciadioids. Towards the apex the paracladia are reduced to triadic, dyadic and uniflorous branches (Fig. 13III-VI), the latter being very numerous. In other species the inflorescence is less ramified, as in Drimys angustifolia Miers (Fig. 13II), and in a considerable number of species they are raceme-like only (Fig. 11I). In such cases we can find descriptions like that of Drimys insipida (R. Br.) Pilger: "flowers single, numerous, congested around the growing point of branchlets, at length pseudo-lateral" (A. C. Smith, 1943, p. 125), which means that because of the continuous growth of the axis after the development of the flowers these obviously appear in lateral position on a longer leafy axis. In one aberrant case Vink (1970) found the inflorescence of D. insipida "to be provided with a terminal flower instead of with the usual vegetative apex."
A very peculiar situation is found in D. piperita Hook. f. (sect. Tasmannia) and has given rise to a particular investigation by Vink (1970). Whereas the other species of the section (D. lanceolata, D. purpurascens, D. stipitata, and D. insipida) are characterized by an inflorescence consisting of "solitary flowers," D. piperita bears "either a solitary flower or a 'cluster' of flowers" in the leaf-axils (Vink, 1970, p. 251). "The flowers are arranged
670
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
E
Fig. 13. I, Eupomatia laurina R. Br., uniflorous brachyblasts on an elder macroblast (Weberling 7545, N Queensland); II, Drimys angustifolia Miers, apex of a flowering shoot at the beginning of anthesis (Reitz & Klein 6982, Brasil, M); III-VI, Drimys winteri J. R. & G. Forst., series ofparacladia, showing reduction until uniflory is reached the single flowered paracladia of 2nd order which are marked "x" originate from the axils of the likewise marked (x) bracts.
in 1-3 rows parallel to the base of the bract" (Fig. 14IX). The "initiation of flower primordia starts with the central flower and is followed by two adjacent flowers, one on each side in the adaxial row" (Fig. 14111). "The next pair of primordia can be situated either laterally in the same row or in the second row in positions alternating .. ." (p. 254/255, see Fig. 14IV-VII, VIII), even a third, abaxial row can be formed. According to Vink these axial
partial inflorescences "represent reduced branching systems of which axes have merged into the cushion and the bracteoles are lost" (p. 255). For the basic arrangement of the flowers two solutions are envisaged by Vink: 1) "a primary cyme, the lateral branches of which produce cymes of second and third order" (see Fig. 141), or 2) "a main axis with a terminal flower and up to four lateral flowers or lateral branches arranged in a 2/5 spiral with reduction on the adaxial side. This situation is far more probable if compared with that in sect. Drimys and Pseudowintera, and even with that in Bubbia" (see Fig. 1411). This
interpretation, which is based on thorough investigations, is in good accordance with the
general view of inflorescence morphology in Winteraceae. The same applies to Vink's
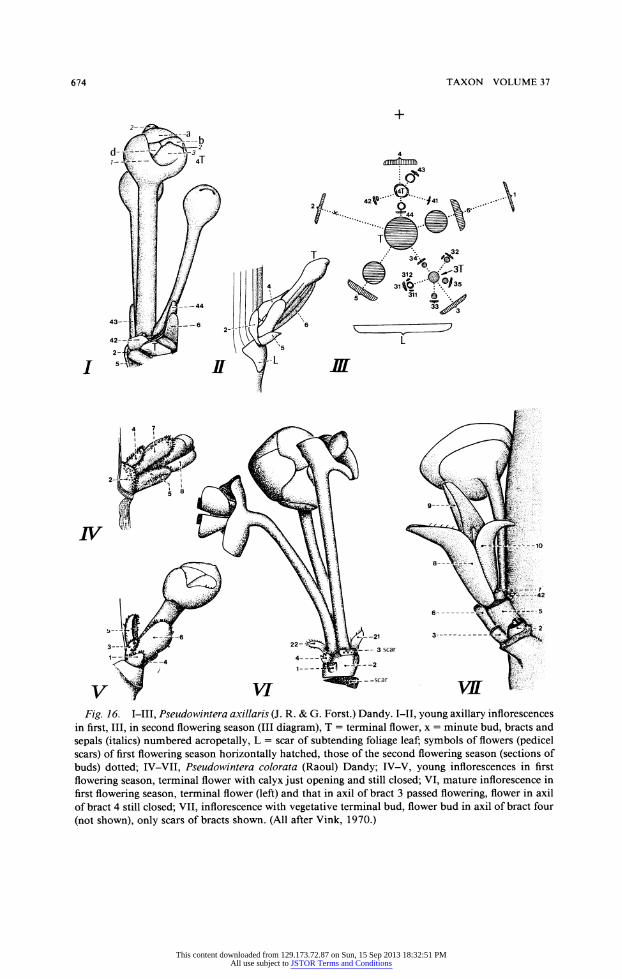
interpretation of the species of Pseudowintera Dandy. These species, P. axillaris (J. R. & G. Forst.) Dandy (Fig. 15), P. colorata (Raoul) Dandy, and P. traversii (Buchanan) Dandy, differ from Drimys by the position and by the structure of their partial inflorescences. The
position "is axillary at its inception, and flowers arise from a lateral position often on branchlets of several years' growth"; this statement of Smith (1943, p. 155) means that we cannot interpret the axillary flowering systems as paracladia of a proliferating panicle as in Drimys winteri. Vink (1970, p. 228) regarded them as short shoots. Their "bracts are
very small and caducous; they are arranged not in pairs, ... but in a 2/5 spiral, .. ." (Fig. 1611 and III), "In the first flowering season flowerbuds develop in the terminal position
671
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
C C- A ', o "o /
x x x xI II
VIm
IV CIV
VI 2
VIII 3 IX Fig. 14. Drimys piperita Hook. f. I-II, two solutions for the basic arrangement of the flowers in
the partial inflorescences; III-VIII, stages of the ontogeny of a partial inflorescence; IX, apical view of terminal bud after removal of bracts, vegetatively proliferating apex in the centre. (After Vink, 1970.)
and (usually) in the axils of the upper bracts. The size of the flower buds increases acrop- etally" (Fig. 161). "In the following flowering season the buds in the axils of the lower bracts develop into secondary inflorescences which repeat the organization of the primary one just described." These are the systems of "smaller" flowers (31-35 and 41-44 with their terminal flowers) in the axils of bract three and four in Fig. 16III. Thus, even in the "primitive" family of Winteraceae we find on the one hand highly specialized 'syndesmic' partial inflorescences as in Drimys piperita, in which all the axes have merged into a cushion, whereas in the other species of the section Tasmannia perennial short shoots are formed which are able to flower during several seasons.
A very broad spectrum of varied inflorescence forms is represented in the Monimiaceae, of which a brief survey may be given here. In doing so we may not forget that most of the Monimiaceae have monoecious or dioecious flowers and that the female inflorescences mostly contain fewer flowers than the male ones; it also occurs that male and female inflorescences are strikingly different, as in Ephippiandra myrtoidea Decne., where the male flowers are arranged in uniflorous partial inflorescences in the axils of foliage leaves, whereas the female plant bears a solitary terminal flower at the end of a leafy shoot (cf. Perkins and Gilg, 1901, fig. 9, p. 51). In Hennecartia omphalandra Poisson, as pointed out by
672
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
Fig. 15. Pseudowintera axillaris (J. R. & G. Forst.) Dandy habit. (After Vink, 1970.)
Perkins and Gilg (1901, p. 4 and p. 72, fig. 19A and F), the male partial inflorescences are botryoids (not racemes) originating in the axils of foliage leaves, whereas the female flowers are solitary and axillary. [For an explanation of the different positions of male and female flowers in mixed (partial) inflorescences, see Perkins and Gilg, 1901, p. 4.]
We have to mention first taxa in which terminal panicles are formed, as is the case in
673
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions
TAXON VOLUME 37
+
2N ' -
' (nnr]i>2 d- 4_mntm4
6.. ..
...~ 2 .6. . A;
?3
.. ? . ~. I ..31
. ' 5
32
43-- ~li~; W yi*'""^ s:;.':.^1
III ij , **"" 34T e 2
"? ^ ^^y^^ ^""?35
3T--
4 . ...31
42---s a
4 7'
_IVIA
T4 - - -
Fig. 16. I-III, Pseudowintera axillaris (J. R. & G. Forst.) Dandy. I-II, young axillary inflorescences in first, III, in second flowering season (III diagram), T = terminal flower, x = minute bud, bracts and sepals (italics) numbered acropetally, L = scar of subtending foliage leaf; symbols of flowers (pedicel scars) of first flowering season horizontally hatched, those of the second flowering season (sections of
buds) dotted; IV-VII, Pseudowintera colorata (Raoul) Dandy; IV-V, young inflorescences in first flowering season, terminal flower with calyx just opening and still closed; VI, mature inflorescence in first flowering season, terminal flower (left) and that in axil of bract 3 passed flowering, flower in axil of bract 4 still closed; VII, inflorescence with vegetative terminal bud, flower bud in axil of bract four (not shown), only scars of bracts shown. (All after Vink, 1970.)
674
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
E
V VI VII VII
x
^1~~~~~~~t m
,.R1
:~~~~
.9
I /I nI I Fig. 17. Inflorescence diagrams of I, Palmeria scandens F. v. Muell. (Schodde 3573, New South
Wales, CANB); II, Hedycarya angustifolia A. Cunn. (Adams 2473, New South Wales, CANB); III, Mollinediafloribunda Tul. (Weberling 6377, Rio de Janeiro, ULM); IV, Steganthera macooraia (Bailey) Endress (after Endress, 1979); V-VIII, partial inflorescences (brachyblasts) of Wilkiea macrophylla (A. Cunn.) A. DC. (Schodde 3359, N Queensland, CANB).
Peumus boldo Mol. (Fig. 191) and in some species of Kibara Endl. [K. coriacea (B1.) Tul.]. In Palmeria scandens F. Muell. (Fig. 171) a discontinuity within the branching system of the inflorescence can be observed, resulting in the formation of a terminal and several lateral botryoid-like parts, whereas in other species (Pullen 8004, Papua, CANB) a contin- uous increase of the ramification of the paracladia in basipetal direction can be found. The majority of the Monimiaceae show a proliferating growth of the inflorescence axis (see also Perkins and Gilg, 1901, p. 3; Perkins, 1925). In this respect Hedycarya angustifolia A. Cunn. (Fig. 1711) differs from Palmeria scandens in that instead of ending in a botryoid and being closed by a terminal flower, the apex of the inflorescence axis remains indeter- minate and continues vegetative growth. The same applies to the inflorescences of Hortonia angustifolia Trimen. with the difference that the axillary, flower-bearing axes can end in a terminal bud instead of a terminal flower, thus forming raceme-like partial inflorescences (see also fig. 1 in Perkins and Gilg, 1901). In Mollinedia floribunda Tul. (Fig. 17III) and
675
r
eA
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
many other species of this genus the axillary partial inflorescences, which otherwise often form panicles (M. gilgiana Perk., M. calodonta Perk.), are reduced to triadic branches, originating in the axils of bracts on a somewhat lignified main axis which after the pro- duction of several pairs of bracts and axillary partial inflorescences turns over to bring forth foliage leaves. The floral region of the main axis is preceded by some pairs of sterile scalelike leaves which probably serve as bud scales for a period of dormancy.
A further reduction of the axillary partial inflorescences can be observed in Steganthera macooraia (Bailey) Endress (Endress, 1979). This species is described as having "single flowers instead of cymes in the axils of leaves or bracts on leaf shoots" (Endress, 1979, p. 316), whereas the genus is described by Perkins and Gilg (1901, p. 53) as having "Flores in cymas racemiformes paucifloras vel in paniculas multifloras dispositi." Figure 17IV, which is reproduced from Endress (1979), shows that the sequence of foliage leaves and bracts with vegetative buds in their axils, and bracts with uniflorous partial inflorescences is repeated during the life time of a shoot. Another form of reduction takes place in the axillary partial inflorescences of Siparuna, a genus which according to Schodde (1970) should be placed in a separate family Siparunaceae together with Bracteanthus Ducke and Glossocalyx Benth. For Hortonia angustifolia Trimen. we have mentioned already that the raceme-like partial inflorescences can end in a bud instead of a terminal flower. In relation to these raceme-like, flower-bearing branches the partial inflorescences of Siparuna cease the formation of primary lateral flowers after the first pair and end in a terminal bud, which can grow out forming a vegetative shoot. This reduction is compensated, however, by the formation of monochasial branching systems in place of the single flowers, as may be seen here from Siparuna apiosyce (Mart.) A. DC. (Fig. 18). Although these monochasia can be very rich in flowers, nothing is reported in literature, whether they are scorpioid or helicoid cymes. The latter seems to be probable to us, although the material investigated was too young to permit a clear decision. In other species such as S. guianensis Aubl., we found the two flower-bearing branches of the axillary, partial inflorescence to be branched di- chasially first thus bearing two monochasia, which here appeared to be scorpioid cymes, although sometimes interrupted by irregular branching indicated by few-flowered clusters. As can be inferred from the figures of Tambourissa species given by Tulasne (1855/1856 pls. XXVI, XXVII), there are also solitary, terminal flowers which could result from a reduction of a terminal botryoid, whereas in T. religiosa (Tul.) A. DC. the (mixed male and female) flowers are arranged in botryoids terminating a leafy shoot. Tambourissa purpurea (Tul.) A. DC. has a female solitary terminal flower. This can also be the case in T. purpurea. In both species the innovation shoots arise from the axils of the foliage leaves immediately below the terminal flower or botryoid. In Wilkiea macrophylla (A. Cunn.) A. DC., bracteose botryoids (Fig. 17VI and VII) are found in the axils of foliaceous leaves or on older branches in axils of leaves which are fallen off, and sometimes even in terminal position. They can bear triadic branches or even botryoids in their proximal part, which arise from the axils of one or more pairs of basal scalelike leaves (including the prophylls), which otherwise can remain empty (or even function as bud scales?). This observation certainly points to the fact that the botryoids are derived from panicles as they are found in terminal position in W. wardellii (F. Muell.) Perk. It is not rare, however, that a cluster of numerous botryoids arises from an older leaf axil, but we could not yet detect the rule of their arrangement, since we had only herbarium material at our disposition. To some extent accessory buds seem to be involved in the formation of the clusters. (Phylloscopic accessory flowers also can occur within the botryoids.) A similar situation can be found in some other taxa such as Kibara serrulata (Blume) Perk. (For further notices on inflorescences in Monimiaceae see Wagner, 1922, 1942).
Several genera of the Monimiaceae, which formerly were included into the subfamily Atherospermoideae (Pax, 1891 a; Perkins and Gilg, 1901), are now regarded as a separate family Atherospermataceae by Schodde (1969). The inflorescences of these genera, Daph-
676
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
41
I
I
/ /
I 1 //
I I I'I
I I
I I i
I I I I
I I
i i
/ I
/ /
/ ' /J
II
I Fig. 18. Siparuna apiosyce (Mart.) A. DC. (Citriosma apiosyce Mart. ex Tul.). I, node of a flowering
shoot with two partial inflorescences, apical part of the main axis removed (Weberling 6456, Cor- covado, Rio de Janeiro, ULM); II, diagram to explain the ramification of the partial inflorescences.
677
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
-A0 --oA
*I 4
^J.
X~IV ImI
II
Fig. 19. Inflorescence diagrams of I, Peumus boldo Mol. (diagram according to Pax, 1901, fig. 65A); II, Atherosperma moschatum Labill. (diagram according to Pax, 1901, fig. 68A); III, Doryophora sassafras Endl. (Weberling 7408, New South Wales, ULM); IV, Piptocalyx moorei Oliv. (Schodde 5119, New South Wales, CANB).
nandra, Nemuaron, Laurelia, Laureliopsis (see Schodde, 1983), Dryadodaphne, Atheros- perma, and Doryophora are monotelic, probably without exception. As far as could be seen from the literature and the scarce material, most of them have a proliferating main axis. The monotelic character of the inflorescences is, however, usually revealed by the axillary partial inflorescences, the paracladia. Their ramification ranges from paniculate-thyrsoidal systems (e.g., Daphnandra) to botryoids (e.g., Nemuaron) and even uniflorous paracladia in Atherosperma (Fig. 1911). Often (phylloscopic) accessory flowering branches or flowers can be found, e.g., in Daphnandra or in Doryophora sassafras Endl. In the latter the accessory branches below the triadic paracladia form triadic flowering branches of similar shape, as can be seen from Fig. 19111. These accessory flowering branches start their anthesis slightly later than the regular paracladia. There are also accessory buds below the large leafy branches. Figure 19111 also shows that the main axis of the entire branching system, as well as the leafy branches, have a proliferating growth. This becomes even more obvious in the main axis which after the production of axillary triadic paracladia returns to the formation of leafy axillary branches. Regarding the triadic paracladia we can discover two supernumerary bracts being inserted between the lateral flowers and the terminal one. These supernumerary bracts might indicate that the triadic paracladia are derived from botryoids by reduction.
In Trimeniaceae, formerly included in the Monimiaceae, Piptocalyx moorei Oliv. (Fig. 19IV) has paniculate inflorescences terminating the shoot and resembling those of Palmeria in their ramification. According to the figure given by Gilg and Schlechter (1919, fig. 2),
678
I *S ,
^
4
izi
ek _k
Z
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
P. macrurus Gilg & Schlechter appears to be similar. For Trimenia papuana Ridl. the terminal position of the inflorescence may be inferred from the description given by Endress and Sampson (1983, p. 449)-"The paniculate inflorescences are produced in leaf axils and at the end of leafy shoots." Trimenia neocaledonica Baker f., is described to have "fewer-flowered botryoids," which, however, also occur in T. papuana. Amborella tricho- poda Baill., formerly included in the Monimiaceae, now regarded to represent a separate family Amborellaceae, has paniculate, partial inflorescences (i.e., with a terminal flower) in the axils of alternate foliage leaves (cf. Money et al., 1950, p. 388; Perkins, 1925, fig. 6A and D) and thus corresponds well to the characters found in Monimiaceae.
In the Trochodendraceae the monotelic character of the inflorescence is clearly seen. Trochodendron aralioides Siebold & Zucc., has botryoids in the sense of Troll (1964, p. 52), i.e., a terminal flower preceded by numerous single lateral flowers, uniflorous para- cladia, thus forming what sometimes is misleadingly named a "raceme with a terminal flower." According to Nast and Bailey (1945), the basal paracladia sometimes bear flowers of tertiary order which means that one or both of the prophylls can be fertile. This reveals that the inflorescence is a reduced panicle, which should correctly be called a botryoid (instead of botrys or raceme). Below the terminal flower a number of sterile bracts ("Zwi- schenblitter," Troll, 1964, p. 15, referring to Nordhagen, 1937, p. 12, "metaxyphylls" Briggs and Johnson, 1979, p. 244) can occur. According to Nast and Bailey the terminal flower of the botryoid can be missing. Then a raceme-like truncate synflorescence is formed, which is not discernible from a polytelic raceme. The botryoids terminate the main axis of the shoot. The continuation of the branching system takes place from the axils of the leaves immediately below the inflorescence, thus a sympodial branching system is formed. [After the completion of this manuscript P. K. Endress published results on "Floral struc- ture, systematics, and phylogeny in Trochodendrales" (Ann. Missouri Bot. Gard. 73: 297- 324. 1986). This publication also contains data on inflorescence morphology, which, how- ever, do not differ from the data given here.]
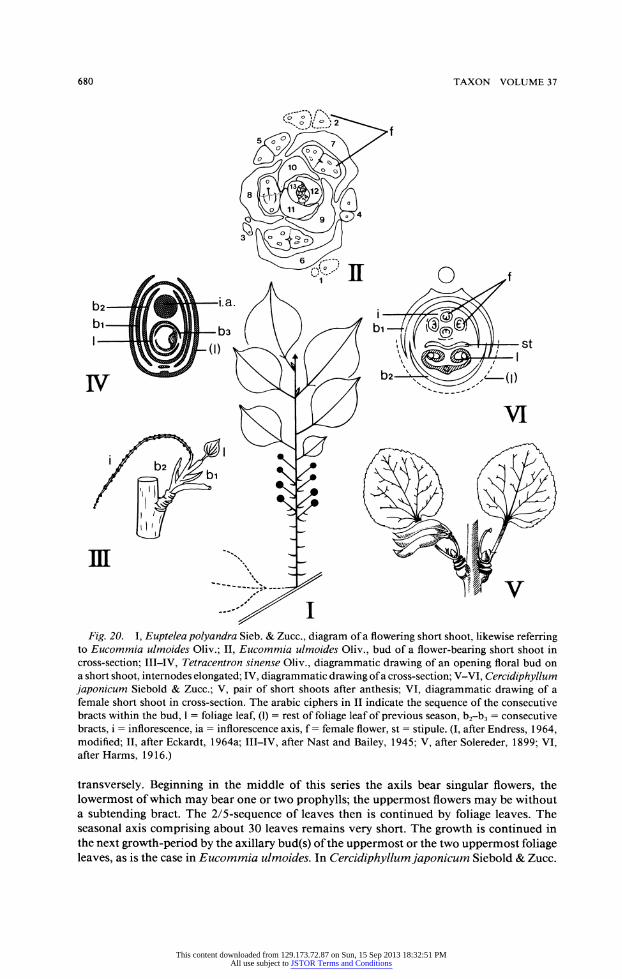
In Euptelea Siebold & Zucc. (Eupteleaceae), Tetracentron Oliv. (Tetracentraceae), and Cercidiphyllum Siebold & Zucc. (Cercidiphyllaceae), all three formerly included in the Trochodendraceae), as well as in Eucommia Oliv. (Eucommiaceae), the formation of in- florescences is confined to short shoots which originate from a previous year's long shoot (Harms, 1897, 1930; Endress, 1986, etc.). In these genera the short shoots develop the inflorescence in a terminal position and their subsequent growth is sympodial. The archi- tecture of the male and female short shoots of Eucommia ulmoides Oliv. is described by Eckardt (1964a, p. 28) as follows (Fig. 201 and II); "The flower buds begin with some 6- 8 sterile scales, which are succeeded on incipient shoots by a varying number (from 1-6- 8) of floriferous bracts of transitional nature and these are followed by a varying number of leaves." These inflorescences may "be compared with an open raceme." However, "Fertile shoots of both sexes show a flower-bearing part only at the base, which is succeeded by an extensive leaf-bearing prolongation." This proliferating growth is limited, however, to the production of a small number of foliage leaves only. The uppermost axillary bud(s) continue the development of the short shoot in the next season. Eckardt (1964a, p. 27, see also 1964b) also stated "Vegetative as well as flower-bearing shoots of Eucommia are sympodial, the uppermost functioning bud formed at the end of the growing season being pseudoterminal." Eckardt (1957, 1964b) already pointed to the high degree of resemblance between the flower-bearing short shoots of Eucommia and Euptelea. This was confirmed by a thorough investigation of Euptelea made by Endress (1969) in which the structure of the flower-bearing short shoots was nearly the same (see also Nast and Bailey, 1946). Thus the diagram in Fig. 201, which was outlined in slight modification of a diagram delineated by Endress for Euptelea, likewise refers to Euptelea polyandra Siebold & Zucc. as to Eucommia. The foliation of the axis starts with about 20 scalelike leaves, the first of which are rather small and relatively broad, especially the two prophylls which are arranged
679
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions
TAXON VOLUME 37
lf
IV ?4 b2 st
VI
bi
11f
Fig. 20. I, Euptelea polyandra Sieb. & Zucc., diagram of a flowering short shoot, likewise referring to Eucommia ulmoides Oliv.; II, Eucommia ulmoides Oliv., bud of a flower-bearing short shoot in cross-section; III-IV, Tetracentron sinense Oliv., diagrammatic drawing of an opening floral bud on a short shoot, interodes elongated; IV, diagrammatic drawing of a cross-section; V-VI, Cercidiphyllum japonicum Siebold & Zucc.; V, pair of short shoots after anthesis; VI, diagrammatic drawing of a female short shoot in cross-section. The arabic ciphers in II indicate the sequence of the consecutive bracts within the bud, 1 = foliage leaf, (1) = rest of foliage leaf of previous season, b2-b3 = consecutive bracts, i = inflorescence, ia = inflorescence axis, f= female flower, st = stipule. (I, after Endress, 1964, modified; II, after Eckardt, 1964a; III-IV, after Nast and Bailey, 1945; V, after Solereder, 1899; VI, after Harms, 1916.)
transversely. Beginning in the middle of this series the axils bear singular flowers, the lowermost of which may bear one or two prophylls; the uppermost flowers may be without a subtending bract. The 2/5-sequence of leaves then is continued by foliage leaves. The seasonal axis comprising about 30 leaves remains very short. The growth is continued in the next growth-period by the axillary bud(s) of the uppermost or the two uppermost foliage leaves, as is the case in Eucommia ulmoides. In Cercidiphyllum japonicum Siebold & Zucc.
680
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
the foliation of the short shoots begins with three cataphylls in a median-distichous series, which is continued by a single foliage leaf in abaxial position (Fig. 20V). This follows from investigations by Solereder (1899) and Harms (1916; see also Harms, 1897). In short shoots remaining vegetative or even growing out later to form long shoots, the foliation is continued by foliage leaves with (more or less) decussate or helical phyllotaxis. In the male or female flower-bearing short shoots, the main axis ends in a raceme-like inflorescence consisting of one to three pairs of perianthless female (Fig. 20VI) or one or two of perianthless male flowers. At least the lowermost of these flowers originates in the axils of small bracts. The apex of the inflorescence remains open. Thus we may conclude that we have to deal with the main florescence of a polytelic system, although coflorescences, the presence of which would permit a clear decision, are missing. In the next season the growth of the short shoot is continued by the axillary bud of the one foliage leaf developed in the last season, as is the case in vegetative short shoots; thus several-years-old short shoots can be formed, which in each season develop only one foliage leaf in abaxial position and, when fertile, end in a few-flowered botrys (Fig. 21). The large number of scars covering the surface of their thickened woody base (Fig. 21) indicates that the age of the short shoots can be considerably high.
In Tetracentron sinense Oliv. a seasonal growth unit of a fertile short shoot comprises two bud scales in median position enclosed by "the stipular flanges" of a withered leaf (Fig. 20IV) and preceding a terminal inflorescence (Fig. 20III). The uppermost scale bears an axillary bud comprising a (third) scale and a foliage leaf in transversal position. This bud shows a proleptic-ananthic development in so far as the scale (which falls off very early) and the foliage leaf are unfolded in the year before the following two scales and the terminal inflorescence come to unfold. The "development of the short axis ... crowds the inflorescence into an apparently lateral position." "The inflorescence is a spike with nu- merous sessile flowers, each subtended by a minute bract .... The flowers tend to be arranged in false whorls. Four decussate bracts are usually present at the apex of the axis, but occasionally the rhachis may terminate in a flower" (Nast and Bailey, 1945, p. 268). Thus the inflorescence is truly monotelic or truncate and in this way demonstrating a transition between the primitive monotelic and the derived polytelic type.
A few remarks on some other families may close this comprehensive survey. The An- nonaceae have been investigated by Hoppe and Weberling in connection with the inter- national, multidisciplinary project arranged by the Department of Systematic Botany of the University of Utrecht. The results, which will be published within a short time do not essentially diverge from those given before by Fries (1919, 1941, 1959). Although the main axis of the inflorescence never ends in a terminal flowering system but persists in the formation of foliage leaves, and of only "axillary inflorescences," the often paniculate or
thyrsoidal structure of these paracladia indicates that the inflorescences are monotelic and in this respect primitive; at least they are clearly derived from monotelic systems. A well known peculiarity of the axillary, flower-bearing systems is their median-distichous phyl- lotaxis, a character which is shared with other families of the Magnoliales as for example Eupomatiaceae. Even in cases of extreme reduction the number of these mostly bracteous leaves indicates the derivation from a richer branched, partial inflorescence. Not rarely a sympodial mode of ramification can be found, but we never met any indication of transition to the polytelic type.
In Himantandraceae the "flowering shoots" are described by van Royen (1962, p. 127) as "axillary, with one terminal flower and 2 or 3, alternating, caducous, sometimes folia- ceous bracts (2 prophylls in ? transversal position and an additional bract), with obscure, axillary buds which rarely develop into a flowering axis" (see also Croft, 1978, fig. 64). Bailey et al. (1943, p. 197) also report, "that in exceptional instances, one of the buds develops a subsidiary flowering axis bearing two scales and an apical flower." We confirmed these data during our studies. Diels (1919, p. 126) regarded the flowering shoots as brach-
681
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
Fig. 21. Cercidiphyllum japonicum Siebold & Zucc. I, node with flower-bearing short shoots after anthesis; II, detail.
yblasts equivalent to those of Michelia (see also Endress, 1977, who also compares Gal- bulimima with Eupomatiaceae and Degeneriaceae). Their subtending foliaceous leaves may fall off at the time of anthesis.
The inflorescences of the Lauraceae have been treated in a survey by Weberling (1985). They are rarely terminal, sometimes pseudoterminal and usually originate from the axils of frondose or bracteose leaves of a proliferating (auxotelic) main axis. In many genera the
682
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
ramification of these axillary flowering systems is thyrso-paniculate. The distal parts of their main axis and their larger branches sometimes form more or less extended botryoids. In some taxa botryoidal or dibotryoidal flowering systems seem to be a more or less constant character (e.g., Urbanodendron verrucosum). In Sassafras, Laurus, Lindera, and also species of Neolitsea the (mostly) botryoidal flowering systems originate from the axils of cataphylls of axillary or terminal brachyblasts which end in a terminal bud. This terminal bud unfolds its leaves synchronously with the botryoids or some time later or even can remain arrested. Whereas all these inflorescences are of monotelic character, the genera Cassytha, and (partly?) Mezilaurus are characterized by polytelic flowering systems with botric or spike- like or umbellate florescences. The results on the genus Litsea reported by Weberling (1985, pp. 406-410) which were based on herbarium material need to be revised, since the flower- shaped elements in Litsea confusa (Figs. 8, 9), L. calophyllantha (Fig. 10) and L. chinensis (Fig. 11 and II) proved to be pseudanthia, i.e., partial inflorescences which are surrounded by an involucre and thus appear as flowers! Since these partial inflorescences probably bear terminal flowers (and then they are botryoids) the inflorescences of the Litsea-species investigated must be regarded as truncate monotelic inflorescences.
In the possibly related Hernandiaceae the inflorescences are polytelic throughout. This became evident after an analysis by Kubitzki (1969) in connection with a monographic treatment of this family. Commonly the inflorescences are described as axillary (see Pax, 1891b). This means, that the main axis of the entire flowering system bears foliaceous leaves and continues its growth, whereas the flower bearing branches arise from the axils of the foliaceous leaves (Fig. 24V). As described by Kubitzki the basic ramification type of these flower bearing branches is a double (or multiple) thyrse. This may be explained here by using some species of the genus Hernandia as examples. In H. guianensis Aubl. and some related species of the subgen. Hernandia (H. obovata 0. C. Schmidt and H. ovigera Vieill.), pleiothyrses (multiple thyrses) as shown in Fig. 241 are formed, in which the thyrsic ramification systems are composed of three-flowered, cymose, partial inflores- cences (symbolized by circles marked with a cross). Each of these umbel-shaped cymes is surrounded by an involucre of four bracts (Fig. 251; see also Pax, 1891b, fig. 79B). As demonstrated by the transverse and vertical diagrams in Fig. 2411 and III, the two outer bracts of the involucre are the prophylls of the cyme which is terminated by the primary flower. In the axil of one of these prophylls a lateral flower is formed which is also preceded by a pair of prophylls, one of which bears a tertiary flower in its axil (marked by two concentric circles). This tertiary flower is also provided with a pair of prophylls. Since the hypopodium (the internode preceding the prophylls) of the secondary flower, remains undeveloped, the prophylls of the secondary flower together with the prophylls of the entire cyme form a false whorl consisting of four bracteose leaves, the involucre. It may be added that the hypopodium of the tertiary flower also remains short. Kubitzki (1969, p. 86) described this three-flowered cyme as a scorpioid cyme. The decision, however, whether the ramification is that of a scorpioid or of a helicoid cyme can at the earliest be made with the occurrence of a fourth flower. Nevertheless scorpioid cymes do occur at least in Illigera Bl. Of major interest is the fact that the main axis of the thyrses and also those of the double and triple thyrses do not end in a terminal flower. Thus the flower bearing system shown in Fig. 241 and IV can be regarded as a polytelic inflorescence which ends in a (rudimentary) terminal bud instead of a main florescence, and therefore, must be called a truncate, polytelic synflorescence in the sense of Troll (1964, p. 100). It consists of paracladia only which either end in a thyrsic coflorescence or bear thyrsic paracladia of higher order. The umbel-shaped, three-flowered parts composing the thyrses are cymose partial florescences. The whole flower-bearing system in Fig. 241, however, is a paracladium itself, forming a part of a proliferating, frondose, polytelic flowering system as shown in Fig. 24V.
As Kubitzki (1966) has shown, these paracladia (Fig. 241) can be modified in different
683
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
Fig. 22. I, Knema furfuracea (Hook. f. & Thoms.) Warb.; II, Myristica bivalvis Hook. f. (After King, 1891.)
ways, e.g., by a lesser degree of ramification resulting in dithyrsic or even monothyrsic systems (H. voyroni Jumelle, H. moerenhoutiana Guillem.), or by different proportions of their interodes. If the frondose main axis of the whole flowering system remains short within the paracladia-bearing zone, these can be crowded in a broad corymb-like inflores- cence, which later is left behind by the further growing tip of its main axis. As could be confirmed by a revision of herbarium material, the thyrsic flowering systems of the genera Illigera Bl., Gyrocarpus Jacq., and Sparattanthelium Mart. also do not end in terminal flowers and can be regarded as polytelic inflorescences. As mentioned already, Kubitzki found the cymes of Illigera forming scorpioid cymes, mostly double-scorpioids whereas the cymose partial florescences of Gyrocarpus are characterized by high degree of dichasial branching in which the terminal (primary, secondary, etc.) flowers are missing with the exception of the ultimate branchlets. For further details see the descriptions of Kubitzki (1966) who refrained from giving a typological classification of the Herandiaceae inflo- rescences.
Fig. 23. Horsfieldia spec. (Darbyshire 1020, Papua, CANB), branching pattern of a paniculate partial inflorescence.
684
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988 685
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
I
) I
m
/r
IV I ? V Fig. 24. I-III, Hernandia subgen. Hernandia. I, vertical diagram of the entire flowering system,
the circles marked with a cross symbolize cymose partial inflorescences, shown in II and III by a transversal (II) and a vertical (III) diagram, p = primary flower, the interrupted lines designate inter- nodes which in reality remain undeveloped. (After Kubitzki, 1969); IV, Hernandia peltata Meissn., trithyrsic paracladium (Docters v. Leeuwen 1964, Java, U); V, H. guianensis Aubl., flowering plant (P. J. Maas 5562, Guayana, U).
686
1
4 ill,-e
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
t p Fig. 25. I, Hernandia guianensis Aubl., thyrsic paracladium with umbel-shaped cymose partial
florescences surrounded by an 'involucre' (A. J. M. Leeuwenberg 11781, Fr. Guiana, U); II, Knema pectinata Warb., part of a flowering branch with flower-bearing short shoot (Beccari 1601, Borneo, M).
Gomortega keule (Baillon) Molina, the only representative of the Gomortegaceae, has axillary and terminal(?) botryoids (see also Rodriguez et al., 1983, pl. 31). These are reported as "panicles" by Buchheim (in Syllabus 1964). By an increase of ramification of the low- ermost branches the botryoids could indeed easily be transformed into panicles.
Although in Myristicaceae the shape of the inflorescences was already used by de Candolle for classification (cf. Eichler, 1875/1878, and Prantl, 1891) these still need further inves- tigation. The inflorescences are always described as axillary. This means that the main axis of the flowering shoot produces foliage leaves only and can bear partial inflorescences in the axils of these leaves, but after the production of a smaller or greater number of axillary partial inflorescences continues its growth as a completely vegetative shoot (Fig. 2211). The axillary partial inflorescences are often thyrsoid-paniculate as in Horsfieldia (Fig. 23), but their ramification can be varied in different ways, or a reduction to few-flowered systems can take place. However, for reliable analyses of their ramification pattern, herbarium material was generally not suitable. In many cases even the occurrence of a terminal flower could not be confirmed by studying herbarium material. Although we feel certain to have found terminal flowers in Virola carinata (Benth.) Warb. (Lleras et al. P 17486, Amazonas, M), we could not confirm their existence in several other genera. In Iryanthera ulei Warb. and I. macrophylla (Benth.) Warb., the head-like flower clusters composing the panicle- shaped inflorescences were found to be indeterminate, the same applies to Compsoneura debilis (A. DC.) Warb. This would mean that these taxa have inflorescences of the polytelic type. A very peculiar ramification of the axillary partial inflorescences is found in Myristica fragrans Houtt. Probably the statement of Prantl (1891, p. 40) and Uphof(1959b, p. 181) that the inflorescences can be cymose, but simply forked without a terminal flower, refers to cases of this kind. The ramification of the two "pronges" (Gabeliste) of the forked inflorescences, however, appears to be racemose. A further complication is the occurrence of a "superposed" bract (in abaxial position) explained by Eichler (1875/1878, p. 144) by the suggestion of a fusion of two originally lateral prophylls. Long shoot-short shoot differentiation of the branching system and cauliflory are also reported for the Myristicaceae, especially in Knema. In this genus the inflorescences consist "of a short woody brachyblast
687
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
up to 17 mm long, of slow unlimited growth, and covered with the scars of fallen pedicels and bracts. It is simple or 2- or 3-furcate, usually appearing as one or more lumpy warts." "They are never distinctly ramified or even paniculate as in a large part of Myristica or in all members of Horsfieldia ...." "In Knema, the flowers themselves are arranged in
terminal, umbel-like, contracted racemes" (de Wilde, 1979, p. 326), as is shown in Fig. 2511 for Knema pectinata Warb. In some members of this genus, e.g., Knema tridactyla Airy-Shaw, the brachyblasts can continue to produce lateral flowers for more than 15 years, developing about three flowers each season (de Wilde, pers. comm.). Thus older branches which long ago lost the subtending leaves of the still flowering short shoots appear to be cauliflorous, as in Knema furfuracea (Hook. f. & Thoms.) Warb.
The brief survey given here shows, that the inflorescences of the so called " primitive angiosperms" exhibit a high degree of variety, and in many taxa reached a high level of specialization, as in the long shoot-short shoot systems of some Winteraceae. Although monotelic inflorescences are prevailing and even solitary flowers can be found within the Magnoliales (as well as in Dilleniales), the question whether the solitary terminal flower may be regarded to be the primitive inflorescence cannot be answered clearly. Thus we may repeat a remark, which once was made on the phylogeny of angiosperms: we need primitive inflorescences, urgently!
Literature Cited Bailey, I. W. and A. C. Smith. 1942. Degeneriaceae, a new family of flowering plants from Fiji. J.
ArnoldArb. 23: 356-365, pls. I-V. , G. Nast and A. C. Smith. 1943. The family Himantandraceae. J. Arnold Arb. 24: 190-206, pls. I-VI.
Baillon, H. E. 1869. Histoire des plantes, vol. I: 289-295. Paris. Blake, T. S. 1972. Idiospermum (Idiospermaceae) a new genus and family for Calycanthus austra-
liensis. Contrib. Queensland Herb. No. 12. Briggs, B. and L. Johnson. 1979. Evolution in the Myrtaceae--Evidence from inflorescence structure.
Proc. Linn. Soc. N.S.W. 102: 157-272. Buchheim, G. 1964. Magnoliales. In: H. Melchior (ed.), A. Engler's Syllabus der Pflanzenfamilien,
12th ed., vol. 2. Berlin. De Candolle, A. P. 1827. Organographie vegetale, ou description raisonnee des organes des plantes.
Paris. Celakovsky, L. F. 1892. Gedanken iiber eine zeitgemasse Reform der Theorie der Bliitenstande. Bot.
Jahrb. Syst. 16: 33-51. Croft, J. R. 1978. Himantandraceae. In: J. S. Womersley (ed.), Handbook of theflora of Papua New
Guinea 1: 126-128. Univ. Press, Melbourne. Diels, L. 1919. Uber die Gattung Himantandra, ihre Verbreitung und ihre systematische Stellung.
Bot. Jahrb. Syst. 55: 126-134. Eckardt, Th. 1957. Zur systematischen Stellung von Eucommia ulmoides. Ber. Deutsch. Bot. Ges.
69: 487-498. 1964a. Some observations on the morphology and embryology of Eucommia ulmoides Oliv.
J. Indian Bot. Soc. 42A: 27-34. 1964b. Die natiirliche Verwandtschaft der Bliitenpflanzen. Umschau 64: 496-502.
Eichler, A. W. 1875/1878. Blithendiagramme, 2 vols. Leipzig. Endress, P. K. 1969. Gesichtspunkte zur systematischen Stellung der Eupteleaceen (Magnoliales).
Berichte Schweizer Bot. Ges. 79: 229-278, pls. I and II. 1977. Uber Bliitenbau und Verwandtschaft der Eupomatiaceae und Himantandraceae (Mag-
noliales). Ber. Deutsch. Bot. Ges. 90: 83-103. 1979. A second species of Steganthera (Monimiaceae) from Australia. Blumea 25: 315-318. 1986. Floral structure, systematics, and phylogeny in Trochodendrales. Ann. Missouri Bot.
Gard. 73: 279-324. and F. B. Sampson. 1983. Floral structure and relationships of the Trimeniaceae (Laurales).
J. Arnold Arb. 64: 447-473.
688
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

AUGUST 1988
Fries, R. E. 1919. Studien fiber die Bliitenverhaltnisse bei der Familie Annonaceae. Acta Horti Berg. 6: 3-48.
1941. Neue amerikanische Annonaceen. Acta Horti Berg. 13: 103-116. 1959. Annonaceae. In: A. Engler and K. Prantl, Die natiirlichen Pflanzenfamilien, 2nd ed.
(ed. H. Melchior), Vol. 17a II. Duncker and Humblot, Berlin. Gifford, E. M., Jr. 1950. The structure and development of the shoot apex in certain woody Ranales.
Amer. J. Bot. 37: 595-611. 1951. Ontogeny of the vegetative axillary bud in Drimys winteri var. chilensis. Amer. J. Bot.
38: 234-243. Gilg, E. and R. Schlechter. 1919. Uber zwei pflanzengeographisch interessante Monimiaceen aus
Deutsche-Neu-Guinea. Bot. Jahrb. Syst. 55: 195-201. Harms, H. 1897. Uber die Stellung der Gattung Tetracentron Oliv. und die Familie der Trocho-
dendraceen. Ber. Deutsch. Bot. Ges. 15: 350-360. 1916. Uber die Bliitenverhaltnisse und die systematische Stellung der Gattung Cercidiphyllum
Sieb. et Zucc. Ber. Deutsch. Bot. Ges. 34: 272-283, pl. V. 1930. Eucommiaceae. In: A. Engler and K. Prantl, (eds.), Die natiirlichen Pflanzenfamilien
2. Aufl., Vol. 18a: 348-351. Verlag Engelmann, Leipzig. King, G. 1891. The species of Myristica of British India. Ann. Roy. Bot. Gard. 3: 275-331, pls. 106-
174. Kohler, H. A. 1923. Kohler's Medizinalpflanzen. 2. Aufl. (ed. G. Pabst). H. Bernmuhler, Berlin. Kubitzki, K. 1969. Monographie der Hernandiaceen. Bot. Jahrb. Syst. 89: 78-148. Money, L. L., I. I. Bailey and B. G. L. Swamy. 1950. The morphology and relationships of the
Monimiaceae. J. Arnold Arb. 31: 372-404. Miiller-Doblies, D. and F. Weberling. 1984. Uber Prolepsis und verwandte Begriffe. Beitr. Biol.
Pflanzen 59: 121-144. Nast, Ch. G. and I. W. Bailey. 1945. Morphology and relationships of Trochodendron and Tetra-
centron. II. Inflorescence, flower and fruit. J. Arnold Arb. 26: 267-276. 1946. Morphology of Euptelea and comparison with Trochodendron. J. ArnoldArb. 27: 186-
192. Nicely, K. A. 1965. A monographic study of the Calycanthaceae. Castanea 30: 38-81. Nooteboom, H. P. 1985. Notes on Magnoliaceae with a revision of Pachylarnax and Elmerrillia
and the Malesia species of Manglietia and Michelia. Blumea 31: 65-121. Nordhagen, R. 1937. Studien fiber die monotypische Gattung Calluna Salisb. I. Bergens Museums
Arbok 1937, Naturvid. Rekke, Nr. 4. Parkin, J. 1914. The evolution of the inflorescence. J. Linn. Soc. Bot. 42: 511-563. Pax, F. 189 la. Monimiaceae. Pp. 94-105. In: A. Engler and K. Prantl (eds.), Die natirlichen Pflan-
zenfamilien, III. Teil, 2. Abt. Leipzig. 1891b. Hernandiaceae. Pp. 126-129. In: A. Engler and K. Prantl (eds.), Die natiirlichen
Pflanzenfamilien, III. Teil, 2. Abt. Leipzig. Perkins, J. and E. Gilg. 1901. Monimiaceae. In: A. Engler (ed.), Das Pflanzenreich IV. 101. Reprint
1960. Verlag Engelmann, Berlin. 1925. Ubersicht iber die Gattungen der Monimiaceae sowie Zusammenstellung der Abbil-
dungen und der Literatur fiber die Arten dieser Familie bis zum Jahre 1925. Verlag Engelmann, Leipzig.
Pilger, R. 1921. Bemerkungen zur phylogenetischen Entwicklung der Bliitenstande. Berichte d. Freien Vereinigungfur Pflanzengeographie und systematische Botanikfur das Jahr 1919: 69-77. Berlin.
1922. Ueber Verzweigung und Bliitenstandsbildung bei den Holzgewachsen. Bibliotheca Botanica 90. Stuttgart.
Prantl, K. 1891. Myristicaceae. Pp. 40-42. In: A. Engler and K. Prantl (eds.), Die natiirlichen Pflanzenfamilien, III. Teil, 2. Abt. Leipzig.
Rickett, H. W. 1944. The classification of inflorescences. Bot. Rev. 10: 187-231. Rodriguez, R., O. Matthei and M. Quezada. 1983. Flora arborea de Chile. Editorial de la Univ. de
Concepci6n, Chile. Royen, P. van. 1962. Sertulum Papuanum 6. Himantandraceae. Nova Guinea Botany 9: 127-135. Schodde, R. 1969. A monograph of the family Atherospermataceae R. Br. Ph.D. thesis, University
of Adelaide. 1970. Two new suprageneric taxa in the Monimiaceae alliance (Laurales). Taxon 19: 324-
328.
689
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions

TAXON VOLUME 37
1983. A new genus in the Monimiaceae alliance (Laurales). Parodiana 2: 298-305.
Sinclair, J. 1968. Florae Malesianae Precursores-XLII. The genus Myristica in Malesia and outside Malesia. Gardens' Bull. Singapore 23.
Smith, A. C. 1943. Taxonomic notes on the Old World species of Winteraceae. J. Arnold Arb. 24: 119-164.
1947. The families Illiciaceae and Schisandraceae. Sargentia 7. Arnold Arboretum, Jamaica
Plain, Massachusetts. Solereder, H. 1899. Zur Morphologie und Systematik der Gattung Cercidiphyllum Siebold et Zucc.,
mit Beriicksichtigung der Gattung Eucommia Oliv. Ber. Deutsch. Bot. Ges. 17: 387-406, pl. XXVIII.
St. Hilaire, A. F. C. P. de. 1840. Leqons de botanique comprenant principalement la morphologie vegetale. Paris.
Takhtajan, A. 1959. Die Evolution der Angiospermen (German translation by W. H6ppner). Jena.
Troll, W. 1964/1969. Die Infloreszenzen. Typologie und Stellung im Aujbau des Vegetationskorpers, Bd. I (1964), 11/1 (1969). Jena. and H. Weber. 1957. Morphologie einschlieBlich Anatomie. Fortschr. Bot. 19: 18-27.
Tucker, S. C. 1959. Ontogeny of the inflorescence and the flower in Drimys winteri var. chilensis. Univ. Calif Publ. Botany 30(4): 257-336.
Tulasne, L. R. 1855/1856. Monographia Monimiacearum. Archives du Museum d'Histoire Naturelle
(Paris) 8. Uphof, J. C. Th. 1959a. Eupomatiaceae. In: A. Engler and K. Prantl, Die natirlichen Pflanzenfa-
milien, 2. Aufl. (ed. H. Melchior), 17a II, 173-176. Duncker and Humblot, Berlin. 1959b. Myristicaceae. Pp. 177-220. In: A. Engler and K. Prantl, Die natirlichen Pflanzen-
familien, 2. Aufl. (ed. H. Melchior) 17a II. Duncker and Humblot, Berlin.
Vink, W. 1970. The Winteraceae of the Old World. I. Pseudowintera and Drimys-Morphology and
taxonomy. Blumea 18: 225-354. 1977. The Winteraceae of the Old World. II. Zygogynum-Morphology and taxonomy.
Blumea 23: 219-250. 1983. The Winteraceae of the Old World. IV. The Australian species of Bubbia. Blumea
28: 311-328. 1985. The Winteraceae of the Old World. V. Exospermum links Bubbia to Zygogynum.
Blumea 31: 39-55.
Wagner, R. 1922. Uber die Existenz anisophyller Monimiaceen. Akad. Wiss. Wien, Sitzung d. Math.- Naturw. Kl.-Akademischer Anzeiger Nr. 8-9: 1-2.
1942. Glossocalyx Zenkeri n. sp., eine anisophylle Monimiacee. Botan. Archiv 44: 1-7.
Warburg, O. 1897. Monographie der Myristicaceen. Nova Acta, Abh. Kais. Leop.-Carol. Deutsch. Akad. 68: 1-680.
Weberling, F. 1982. Current problems of modern inflorescence morphology. Austral. Syst. Bot. Soc. Newsletter No. 30: 5-21.
1983a. Evolutionstendenzen bei Bliitenstanden. Abh. Akad. Wiss. Lit. Mainz, Math.-Naturw. Kl. Jg. 1983, Nr. 1.
1983b. Fundamental features of modern inflorescence morphology. Bothalia 14: 917-922. 1985. Zur Infloreszenzmorphologie der Lauraceae. Bot. Jahrb. Syst. 107: 395-414.
Wilde, W. J. J. O. de. 1979. New account of the genus Knema (Myristicaceae). Blumea 25: 321- 478.
Williams, K. A. W. 1979/1980. Native plants of Queensland. Brisbane.
690
This content downloaded from 129.173.72.87 on Sun, 15 Sep 2013 18:32:51 PMAll use subject to JSTOR Terms and Conditions