survival of the quarantine root-knot nematodes m....
TRANSCRIPT

University: Gent
Faculty: Science
Department: Biology
ACADEMIC YEAR 2014-2015
Survival of the quarantine root-knot nematodes M.
chitwoodi and M. fallax in waste products of the processing
industry
Martin Baiye Baiye
Promoter: Prof. dr. ir. Wim Wesemael Thesis submitted to obtain the degree of
Master of Science in Nematology

Survival of the quarantine root-knot nematodes M.
chitwoodi and M. fallax in waste products of the processing
industry
Martin B. BAIYE1, Wim M. L. WESEMAEL1,2,
1University of Ghent, Faculty of Sciences, Department of Biology, K.L Ledeganckstraat 35,
9000, Ghent, Belgium
2 Institute for Agricultural and Fisheries Research (ILVO), Burgemeester Van Gansberghelaan
96, 9820 Merelbeke, Belgium

1
SUMMARY
Root-knot nematodes (RKN), Meloidogyne spp., are ubiquitous plant pests that affect a wide
range of crops thereby posing enormous threat to global food security. The quarantine species
M. chitwoodi and M. fallax have become prevalent in some potato and vegetable production
areas causing significant crop losses. Effective treatment of waste soil from the potato and
vegetable processing industry is important to avoid the dissemination of these nematodes. This
study was designed to examine the effect of different temperature treatments on the survival of
M. chitwoodi, M. fallax and M. incognita. Both incubator and warm water bath treatments were
used in this study. For incubator treatment of second-stage juveniles (J2), survival after 40°C
during 1 hour was high with 88.12 ± 2.38%, 85.26 ± 2.46% and 94.11 ± 1.69% survivals for M.
chitwoodi, M. fallax, and M. incognita respectively. Lowest survival was recorded at 50°C for
M. chitwoodi and M. fallax and at 60°C for the tropical M. incognita (0% survival). The lowest
hatching percentage (hp < 5%) for egg masses for both quarantine nematodes and M. incognita
was reported at 50°C and 60°C respectively. For warm water bath experiments, highest survival
of J2 was observed at 40°C for quarantine species and at 50°C for M. incognita. Nonetheless,
lowest hatching percentage (hp < 5%) was observed at 50°C for all species. Generally,
temperature influences the survival and hatching of juveniles with survival decreasing with
increase in temperature and time. Warm water treatment seems more effective than incubator
treatment and is recommended as an efficient treatment option. Further studies are required to
examine the difference in hatching percentage between warming up egg masses from an initial
room temperature to that of a stable temperature regime.
Key words: Quarantine nematodes, hatching, survival, heat treatments, waste soil

2
INTRODUCTION
A major global challenge facing the agricultural sector is the need to enhance food security for
a constantly increasing human population (De Waele & Elsen, 2007). Food or agricultural
product is plaque with several plant pathogens and pests which include several viruses, bacteria,
fungi, insects and nematodes. Plant-parasitic nematodes are among the most economically
damaging pest on horticultural and field crops, causing an estimated US$100 billion loss
globally on an annual basis (Oka et al., 2000). These nematodes have an exceedingly wide host
range and survive varying environmental conditions (Perry, 1999).
Among plant-parasitic nematodes, root-knot nematodes (Meloidogyne chitwoodi and M. fallax)
have become prevalent in potato and vegetable production causing significant crop losses
(Wesemael et al., 2011). These nematodes have been reported from Argentina, Belgium,
Germany, Netherlands, USA, Mexico, Portugal, France, Switzerland and South Africa
(OEPP/EPPO, 2009;Wesemael et al., 2011). Nonetheless, M. chitwoodi was also reported in
Turkey and M. fallax has so far been detected in New Zealand, Australia and UK (Van Der
Gaag et al., 2012). An evaluation of pest risk analysis showed that these pests could establish
wherever potatoes can be grown and coarse soil texture with high sand content is considered to
favor establishment (Suffert & Giltrap, 2012).
However, in addition to direct root damage, the gall-forming sedentary endoparasites elicit
infection by secondary pathogens such as fungi and bacteria and therefore, act as a major cause
of preventable crop disease and yield loss (Nicol et al., 2011). Above-ground symptoms are not
readily apparent, but may consist of various degrees of stunting, lack of vigor and wilting under
moisture stress (OEPP/EPPO, 2004). Nevertheless, substantial crop losses from RKN could be
much greater if species currently causing localized damage (e.g. quarantine spp.) became more
widespread (Singh et al., 2013). For example, cumulative losses from potato cyst nematodes
introduction to Australia were estimated at 370 million AUD over 20 years (Hodda & Cook,
2009), and Pine wood nematode introduction to Europe estimated at 22 billion EUR over 22
years (Soliman et al., 2012).
Dispersal or dissemination of the plant-parasitic nematodes is largely restricted to the
movement of soil (McNeill et al., 2006), infected plants and planting materials. In the
processing industry, plant-parasitic nematodes can be found on heaps of muddy waste,
including soil washed off vegetables and vegetable waste, waste water and mud from lagoons
where the waste water collects (Gamon & Lenne, 2012). Nevertheless, infested waste soil can

3
also be found in potato and vegetable farms (Akhtar, 1993), crop stock warehouses and adhering
to harvesting machines and transport vehicles. Thus, effective treatment of waste soil before
disposal is essential to avoid further spreading of the quarantine root-knot nematode M.
chitwoodi and M. fallax. Moreover, in Belgium it’s prohibited to grow crops from which the
below ground parts are harvested in M. chitwoodi or M. fallax infested soil to avoid further
spread. Only when a sound system to treat waste products is in place, this measure might be
altered.
In this study, different temperature treatments of sterilized soil inoculated with M. chitwoodi,
M. fallax and M. incognita at varying temperatures were investigated. The effect of temperature
on survival was then examined.
MATERIALS AND METHODS
Nematode cultures
Three nematodes species were used in this study (Meloidogyne chitwoodi, M. fallax and M.
incognita). Cultures of these nematode species were obtained from the ILVO stock cultures and
maintained on tomato plants in plastic pots (1.5l) filled with sandy soil under temperature
controlled glass house conditions (16 hours light, 17-26°C). Roots of the tomato plants were
harvested between 6 to 7 weeks after inoculation. The roots were washed, cut into small pieces
of 1-2 cm and placed on a modified Baermann’s funnel (Hooper, 1986) in order to obtain freshly
hatched J2s. The Baermann’s funnel technique (Baermann, 1917) is based on the motility of
nematodes and enables nematodes to be separated from organic material. To obtain egg masses,
fresh roots were kept in water and under a binocular microscope (50× objective) small root
pieces containing a female with egg mass were cut from the roots with a scalpel. The root pieces
with the egg mass were transferred to a water filled plate and kept for further use.
Preparation of juveniles and egg masses
Freshly hatched J2 (<24 hour) were tapped from the Baermann’s funnel. The nematodes were
allowed to settle for about 20 minutes and a vacuum pump (Vacuubrand, BVC 21) was used to
suck out the excess water. The remaining volume in the beaker was noted using a measuring
cylinder. The content in the beaker was homogenized by blowing thin air through a 10ml pipette
and subsamples (1ml) taken and transferred into a counting dish. Successive counts of juveniles
per subsamples were noted aided by a counter. The average number of juveniles per subsamples

4
was then obtained and multiplied by the total volume to obtain the total number of juveniles in
the total sample. Subsamples of 100 J2 to be inoculated were obtained by multiplying the
number (100 J2) by the total volume divided by the total juveniles (Total volume × 100 J2/Total
juveniles).
For egg masses, root pieces containing 1 egg mass were picked and placed on a filter (mesh
size of 48 µm) in a small sieve. A second filter was then placed on top of the one beneath to
hold the egg mass in place and the sieve was closed for firmed grip. The sieves with the egg
masses were then placed unto a glass plate filled with water ready for use.
Sterile soil preparation
Sandy soil composed of 74% sand, 17% sandy loam, 3% loam and 6% clay and sterilized at
100°C for 16 hours, was filtered through a metal sieve of 3.055 mm into a bath. The dry soil
was measured and water was added to obtain 20% humidity. The soil was mixed thoroughly
for homogeneity to keep the soil moist for further use.
Temperature experiments for juveniles and egg masses
To examine the effect of temperature on survival, both incubator (Binder 53) and warm water
bath (Grant Y14) treatments were used. In all cases, two temperature regimes were noted with
10 replicates each, the warmed up regime (wu) and a desired temperature regime. Each test
sample was labelled appropriately and ran for 1 or 2 hours. In the warm water bath experiment,
the test samples were placed in a rack and immersed in the warm water with the water level
slightly above the soil in the tube to ensure uniform temperature. For the incubator experiment,
the samples were placed in a plastic plate of about 26cm in diameter and into the incubator with
the set temperature.
For juveniles’ incubator experiments, 30g of prepared sterile soil was filled into tubes (3.2cm
in diameter and 5.5cm height) and 50ml test tubes (3.1cm in diameter by 11.6cm height) were
used in warm water bath experiments. A micropipette was used to inoculate subsamples of 100
J2 into the tubes. The test sample tubes were then placed in an incubator or warm water bath
depending on the desired experiment. For M. chitwoodi and M. fallax, the experiments were
run in an incubator and warm water bath at 40°C and 50°C. For M. incognita, the experiment
was run at 40°C, 50°C and 60°C for incubator treatment, 40°C and 50°C for warm water bath
treatment.

5
For the experiment with egg mass, 15g of prepared sterile soil was filled into the test tubes.
Sieves containing egg masses were transferred unto the test tubes with a gentle push. Additional
15g of soil was then added on top of the sieve in the tube to ensure it was properly covered.
Control treatments for the experiments were established for juveniles and egg masses as
described above but sample tubes were kept at room temperature and the number of live J2 and
hatched juveniles from egg masses were counted with time.
Nematode extraction
Automatic zonal centrifuge was used for nematode extraction (Hendrickx, 1995). The principle
is based on the density of the nematode.
Extraction was done one day after treatment. For juveniles, tubes with soil (30g) were properly
rinsed with water in a 1000ml beaker. A second beaker of 100ml (receiver) was labelled
correspondingly with the first and both were placed on the automatic zonal centrifuge for the
extraction process to begin. After the extraction, the beaker (100ml) with the nematodes was
allowed to stand for at least 3 hours for the nematodes to settle. A vacuum pump was then used
to suck out the excess water to facilitate counting.
For egg masses, both the soil and the sieves were rinsed thoroughly with water in a beaker.
Sieves with egg masses were then transferred to small glass bottles with their corresponded
labelings. Water was then added to ensure the sieves were immersed completely in the water.
The glass bottles were then closed with a perforated lid or parafilm for ventilation. The bottles
were kept at room temperature and hatched juveniles counted on a weekly basis for 4 weeks.
On the fourth week, the egg masses were crushed with a few drops of Sodium hypochlorite
aided with a small glass rod of about 15cm. Both juveniles and unhatched eggs were counted.
Counting of nematodes
A binocular microscope (50× objective), handling needle and a hand tally counter were used to
count nematodes. J2 were counted and separated based on live or dead. Live juveniles were
detected by observing movement. Seemingly, dead J2 were stimulated with the handling needle
and monitored for possible movement.
6
Data analysis
Statistical analyses were performed using GraphPad Prism Version 5 for Windows (GraphPad
Software, San Diego California USA) and Excel 2013 for Windows (Microsoft Corporation).
The hatching data obtained were fitted to the logistic model y = c/(1+exp (-b×(time-m))), where
y is the cumulative % hatch; the model is described by three parameters: the time at which 50%
of the hatch is reached (m), the hatching rate (b) and the final hatching percentage (c)
(Wesemael et al., 2006). Those parameters were calculated for all replicates and all treatments.
Mortality rates as well as m, b and c values were subjected to the Shapiro-Wilk test of normality
and when data was found to be non-normally distributed, the nonparametric Mann-Whitney
and Kruskal-Wallis tests were used. For more Gaussian data, the Student’s t test for grouped
data and one-way analysis of variance followed by Dunnett ׳s multiple comparisons test or
Bonferroni correction were used as appropriate to compare mortalities between and within
treatments. P values <0.05 were considered statistically significant. Unless otherwise stated,
error bars represent standard error of the mean (SEM) values.
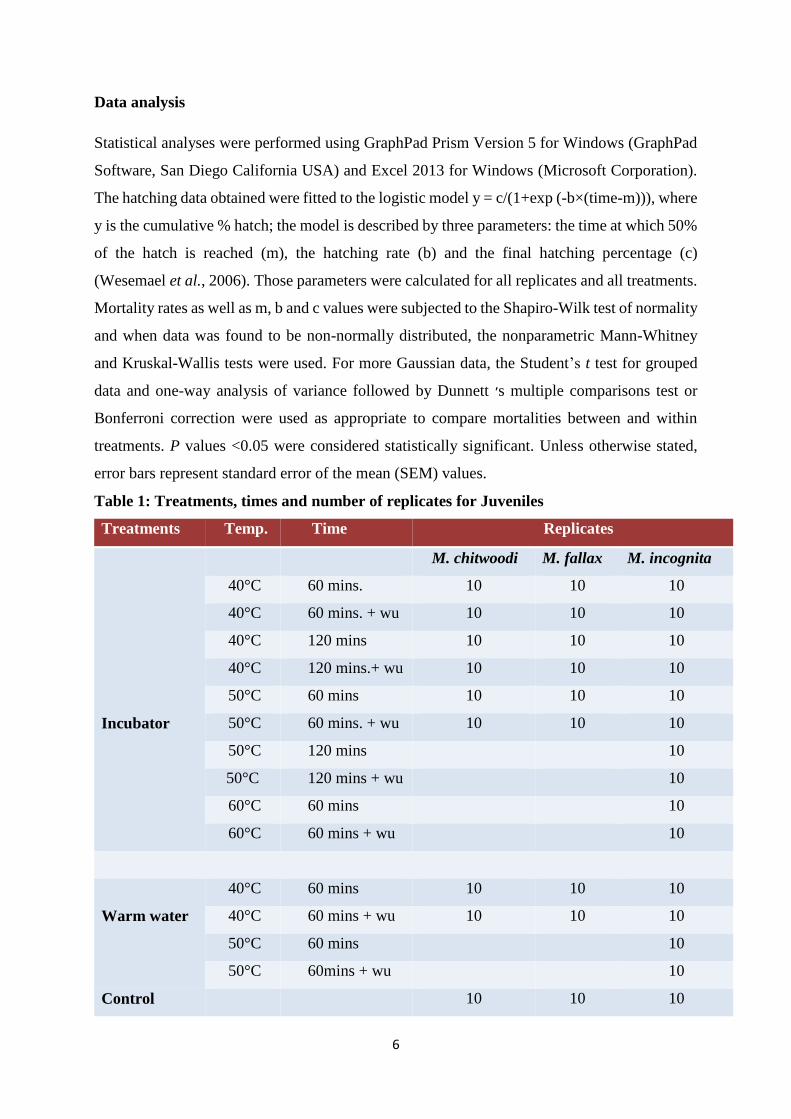
Table 1: Treatments, times and number of replicates for Juveniles
Treatments Temp. Time Replicates
Incubator
M. chitwoodi M. fallax M. incognita
40°C 60 mins. 10 10 10
40°C 60 mins. + wu 10 10 10
40°C 120 mins 10 10 10
40°C 120 mins.+ wu 10 10 10
50°C 60 mins 10 10 10
50°C 60 mins. + wu 10 10 10
50°C 120 mins 10
50°C 120 mins + wu 10
60°C 60 mins 10
60°C 60 mins + wu 10
Warm water
40°C 60 mins 10 10 10
40°C 60 mins + wu 10 10 10
50°C 60 mins 10
50°C 60mins + wu 10
Control 10 10 10
7
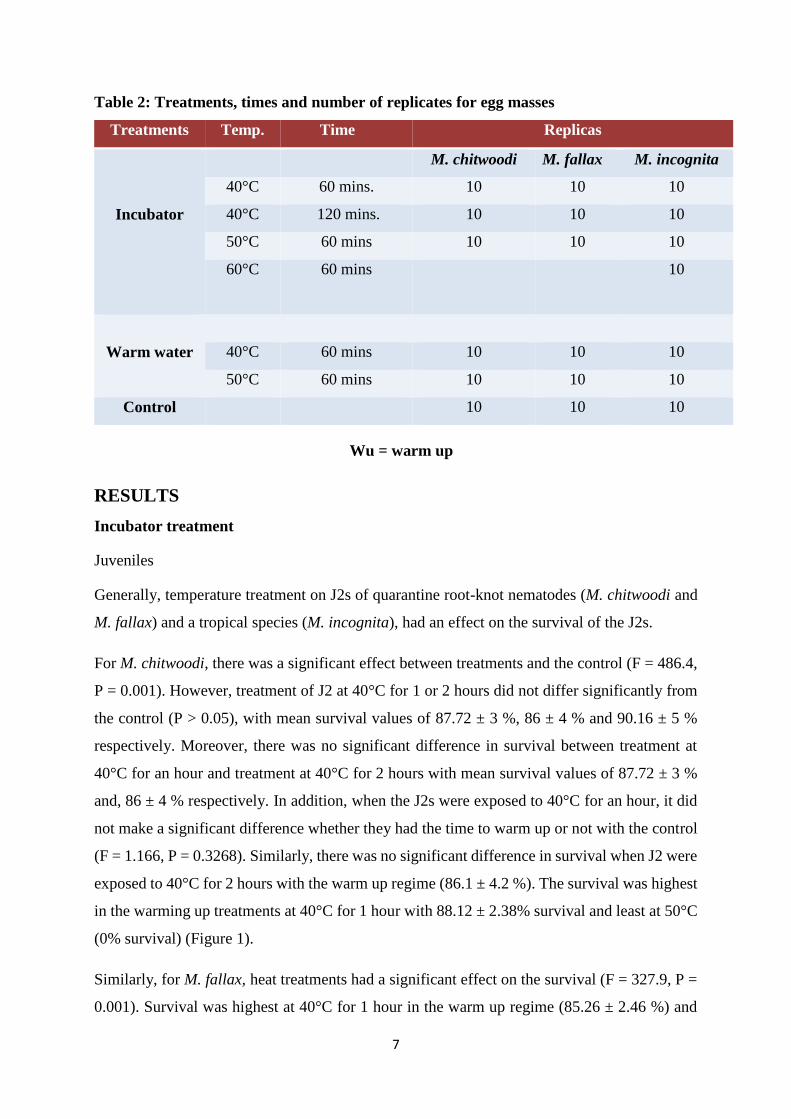
Table 2: Treatments, times and number of replicates for egg masses
Treatments Temp. Time Replicas
Incubator
M. chitwoodi M. fallax M. incognita
40°C 60 mins. 10 10 10
40°C 120 mins. 10 10 10
50°C 60 mins 10 10 10
60°C 60 mins 10
Warm water
40°C 60 mins 10 10 10
50°C 60 mins 10 10 10
Control 10 10 10
Wu = warm up
RESULTS
Incubator treatment
Juveniles
Generally, temperature treatment on J2s of quarantine root-knot nematodes (M. chitwoodi and
M. fallax) and a tropical species (M. incognita), had an effect on the survival of the J2s.
For M. chitwoodi, there was a significant effect between treatments and the control (F = 486.4,
P = 0.001). However, treatment of J2 at 40°C for 1 or 2 hours did not differ significantly from
the control (P > 0.05), with mean survival values of 87.72 ± 3 %, 86 ± 4 % and 90.16 ± 5 %
respectively. Moreover, there was no significant difference in survival between treatment at
40°C for an hour and treatment at 40°C for 2 hours with mean survival values of 87.72 ± 3 %
and, 86 ± 4 % respectively. In addition, when the J2s were exposed to 40°C for an hour, it did
not make a significant difference whether they had the time to warm up or not with the control
(F = 1.166, P = 0.3268). Similarly, there was no significant difference in survival when J2 were
exposed to 40°C for 2 hours with the warm up regime (86.1 ± 4.2 %). The survival was highest
in the warming up treatments at 40°C for 1 hour with 88.12 ± 2.38% survival and least at 50°C
(0% survival) (Figure 1).
Similarly, for M. fallax, heat treatments had a significant effect on the survival (F = 327.9, P =
0.001). Survival was highest at 40°C for 1 hour in the warm up regime (85.26 ± 2.46 %) and

8
least at 50oC for 1 hour (100% mortality). However, neither the survival for an hour nor 2 hours
at 40°C differed significantly from the control (P > 0.05), with mean survival of 83.75 ± 3.82
%, 83.73 ± 4.3 % and 86.5 ± 4.13 % respectively (Figure 1). Again, treatment for an hour at
40°C did not differ significantly from treatment at 40°C for 2 hours. Moreover, there was no
significant effect in treatments between warming up the juveniles to a desired temperature and
a stable temperature regime with the control (F = 0.7769, P = 0.4698). Nevertheless,
temperature has little or no influence on the survival between warming up and not warming up
the juveniles at 40°C for 1 or 2 hours (P > 0.05), with mean values of 85.26 ± 2.5 %, 83.75 ±
3.82 % for 1 hour and 83.75 ± 3.82 % and 83.73 ± 4.3 % for 2 hours accordingly.
Contrary to M. chitwoodi and M. fallax, M. incognita showed slightly different results. The
lowest survival was observed at 60°C (100% mortality) as opposed to 50°C for quarantine
species. Moreover, M. incognita showed highest survival (94.11 ± 2.66 %) at 40°C (+ wu)
among the different species examined. Although, heat treatment had a significant effect on the
survival (F = 369.7, P = 0.001), treatment at 40°C for 1 or 2 hours did not differ significantly
from each other and the control (P > 0.05) with mean survival values of 93.47 ± 2.52 %, 92.94
± 1.93 % and 94.37 ± 2.66 % respectively. However, treatment at 50°C for 1 and 2 hours
differed significantly from each other and the control (P < 0.05) with mean values of 30.08 ±
7.42 %, 21.3 ± 9.95 % and 94.37 ± 2.66 % correspondingly. Nonetheless, there was no
difference in survival between the warm up regime and a fixed temperature regime (Figure 1).
Figure 1: Mean survival of second-stage juveniles from Meloidogyne chitwoodi, M. fallax
and M. incognita after heat treatments in an incubator during different time periods. (wu
= juveniles exposure in the incubator at room temperature and warm up to the given
temperature)
Egg masses
0
20
40
60
80
100
M. chitwoodi M. fallax M. incognita
Surv
ival
(%
)
Control
wu + 40°C for 60 mins.
40°C for 60 mins.
wu + 40°C 120 mins.
40°C for 120 mins.
wu + 50°C for 60 mins.
50°C for 60 mins.
wu + 50°C for 120 mins.
50°C for 120 mins
wu + 60°C for 60 mins.

9
Similar to the juveniles, heat treatment had an influence on the hatching percentage of J2s from
RKN.
For M. chitwoodi, temperature incubator treatment had an effect on the hp (F = 34.88, P =
0.001), hatching rate and the time at which 50% of the hatching is reached (F = 6.761, P =
0.0042). The lowest hatching percentage (hp < 5%) was obtained with the treatments at 50°C
for 1 hour (Fig. 2). Moreover, at 40°C, the hatching percentage for 1 (88.3 ± 4.64 %) or 2 hours
(62.68 ± 12.1%) differed significantly from each other (P < 0.05). However, the hp for 1 hour
(86.09 ± 2.29 %) did not differ from the control (Fig. 2, Table 3).
Also, for M. fallax, temperature had an influence on the hatching percentage (P = 0.001). The
lowest hp was observed at 50°C (hp < 5%). Moreover, the final hp for 1 (91.04 ± 6.1245 %) or
2 hours (65.6 ± 9.69 %), control (93.48 ± 2.55 %) and 2 hours at 40°C differed significantly
from each other (P < 0.05). However, the hp at 40°C for 1 hour did not differ from the untreated
control (P > 0.05). In addition, the time at which 50 % of the hatching was reached was
significantly different across the different treatment and the control (P = 0.001). No effect of
temperature on the hatching rate was observed (P = 0.1352) (Fig. 3, Table 3).
For M. incognita, the hp differed across the different treatments (P = 0.001). Although the hp
was higher for the treatment at 40°C for 1 hour (94.45 ± 0.83 %) than the untreated control
(93.32 %), the effect was not significant. Neither did temperature had an effect on the untreated
control and treatment at 40°C for 2 hours. However, temperature did influence the hp at 50°C
for 1 hour. The lowest hatching percentage was observed at 60°C for 1 hour (hp < 5%).
In figure 4, the hatching patterns can be seen for M. incognita. Heat treatment did not influence
the moment at which 50% of the hatching was reached (P = 0.2532). However, the hatching
rate was affected (P = 0.001) (Table 3).

10
Table 3: Parameters of the logistic curve y = c/(1 + exp(-b×(time-m))) describing hatching
of second-stage juveniles of Meloidogyne chitwoodi, M. fallax and M. incognita at heat
treatment in an incubator at different time period. The results are the means of 10
replicates of the time at which 50% of the total hatching is reached (m), the hatching rate
(b) and the maximum hatching percentage (c).
Warm water treatment
Juveniles
Like incubator treatment, warm water treatment had an influence on the survival of quarantine
and tropical nematodes with very high variability between both treatments.
M. chitwoodi recorded highest survival at 40°C for 1 hour (1.21 ± 1.81 %). The influence of
temperature on survival did not differ significantly between 1 and 2 hours (100% mortality) at
40°C (P > 0.05). The treatments differed significantly with the control (F = 357, P = 0.001). In
addition, when the J2s were exposed to 40°C for an hour, there was no significant difference
whether they had the time to warm up or not (P > 0.05) (Fig. 5).
Similar results were obtained for M. fallax with 100% mortality at 40°C for 1 hour and the
treatment was significantly different from the control (P < 0.05). However, there was no
difference in survival between warming up the juveniles and not warming up.
Contrary to the above, M. incognita showed least survival at 50°C (100% mortality). The
highest survival was observed at 40°C for 1 hour (80.2 ± 7.59 %). Nevertheless, treatment
between 1 or 2 hours did not differ from each other but differed from the control (F = 509.4, P
= 0.001). Equally, there was no difference in survival between warming up the juveniles to a
desired temperature regime and a fixed temperature regime (P > 0.05) (Fig. 5).
Species Temp. Time m b c
M. chitwoodi 40°C 60 mins. 2.5 1.27 88.3
Control
40°C 120 mins. 3 1.5 62.7
2.6 1.23 86.1
M. fallax 40°C 60 mins. 2.3 1.36 91
Control
40°C 120 mins. 2.8 1.42 66
2.2 1.33 93.5
M. incognita 40°C 60 mins. 2.3 1.4 94.45
Control
40°C 120 mins. 2.5 1.32 88
50°C 60 mins. 2.6 1.67 42.7
2.4 1.28 93.3

11
Figure 5: Mean survival of second-stage juveniles from Meloidogyne chitwoodi, M. fallax
and M. incognita after heat treatments in warm water bath during different time periods.
(wu = juveniles exposure in the warm water at room temperature and warm up to the
given temperature)
Egg masses
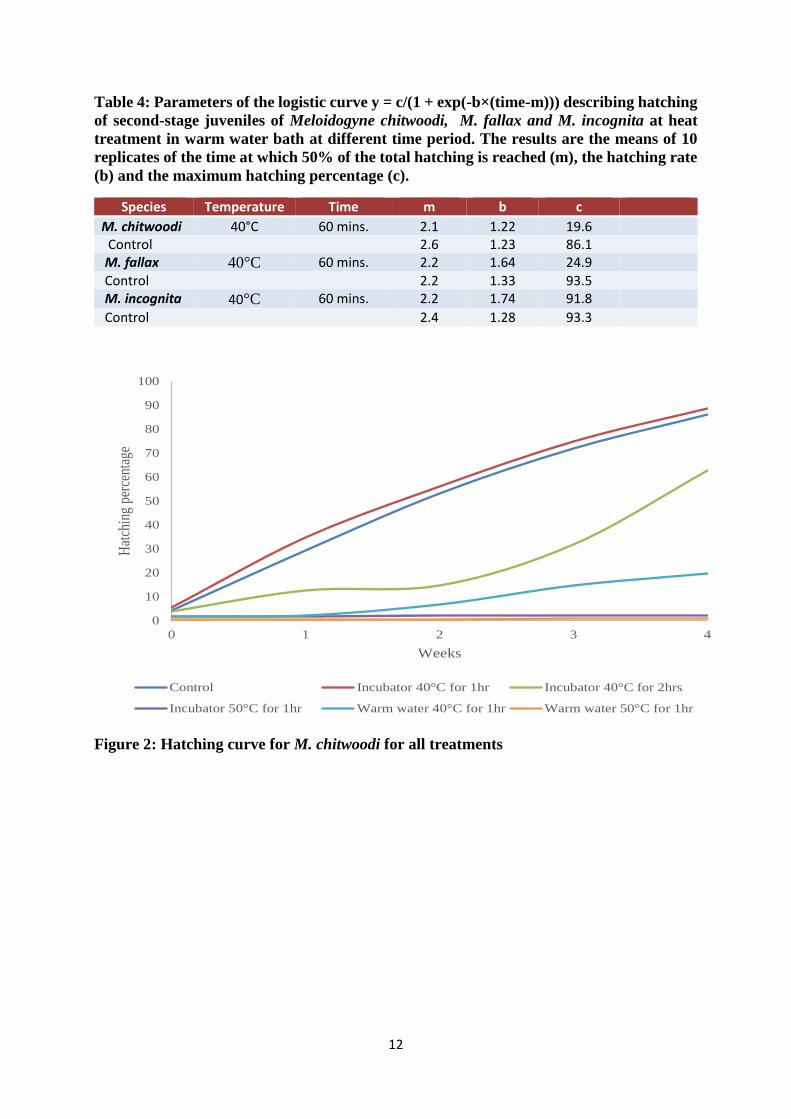
From figure 2, it can be deduced that warm water treatment has an effect on the hatching
percentage (hp), the hatching rate and the time at which 50% of the hatching was reached (P <
0.05). For M. chitwoodi, the lowest hatching percentage was obtained at 50°C for 1 hour (hp <
5%). At 40°C for 1 hour, the hatching percentage was 19.6 ± 4.79 % (Table 4).
Likewise for M. fallax, temperature had an effect on the hp, the hatching rate and the time at
which 50% of the hatching was reached (Fig. 3). The hatching percentage was 24.9 ± 12.51 %
at 40°C for 1 hour. The lowest hatching percentage (hp <5%) was obtained at 50°C for 1 hour
(Fig. 3, Table 4).
For M. incognita, the hatching percentage at 40°C for 1 hour did not differ significantly from
the control (P = 0.2735), with mean hp of 91.77 ± 3.65 % and 93.3 ± 2.31 respectively (Table
4). The lowest hatching percentage (hp < 5%) was observed at 50°C for 1 hour. In addition, the
time at which 50% of the hatching was reached did not differ from the control. However,
temperature had a significant effect on the hatching rate (P = 0.001) (Fig. 4).
0
20
40
60
80
100
M. chitwoodi M. fallax M. incognita
Surv
ival
(%
)
Control
wu + 40°C for 60 mins.
40°C for 60 mins
wu + 40°C for 120 mins.
40°C for 120 mins
wu + 50°C for 60 mins.
50°C for 60 mins.
12
Table 4: Parameters of the logistic curve y = c/(1 + exp(-b×(time-m))) describing hatching
of second-stage juveniles of Meloidogyne chitwoodi, M. fallax and M. incognita at heat
treatment in warm water bath at different time period. The results are the means of 10
replicates of the time at which 50% of the total hatching is reached (m), the hatching rate
(b) and the maximum hatching percentage (c).
Species Temperature Time m b c
M. chitwoodi 40°C 60 mins. 2.1 1.22 19.6 Control 2.6 1.23 86.1 M. fallax 40°C 60 mins. 2.2 1.64 24.9 Control 2.2 1.33 93.5 M. incognita 40°C 60 mins. 2.2 1.74 91.8
Control 2.4 1.28 93.3
Figure 2: Hatching curve for M. chitwoodi for all treatments
0
10
20
30
40
50
60
70
80
90
100
0 1 2 3 4
Hat
chin
g pe
rcen
tage
Weeks
Control Incubator 40°C for 1hr Incubator 40°C for 2hrs
Incubator 50°C for 1hr Warm water 40°C for 1hr Warm water 50°C for 1hr

13
Figure 3: Hatching curve for M. fallax for all treatments
Figure 4: Hatching curve for M. incognita for all treatments
0
10
20
30
40
50
60
70
80
90
100
0 1 2 3 4
Hat
chin
g pe
rcen
tage
Weeks
control Incubator 40°C for 1hr Incubator 40°C for 2hrs
Incubator 50°C for 1hr Warm water 40°C for 1hr Warm water 50°C for 1hr
0
10
20
30
40
50
60
70
80
90
100
0 1 2 3 4
Hat
chin
g p
erce
nta
ge
Weeks
control Incubator 40°C for 1hr Incubator 40°C for 2hrs
Incubator 50°C for 1hr Incubator 60°C for 1hr Warm water 40°C for 1hr
Warm water 50°C for 1hr

14
DISCUSSION
Temperature is an important environmental factor for organisms. Different organisms have
different optimal temperatures for normal activities and high and low temperature limits they
can tolerate. This is also true for nematodes (Tsai, 2008). Temperature has been shown to
influence all aspects of nematode life cycle and behavior including hatching, motility, invasion,
development and survival (Wallace, 1963; Davide & Triantaphyllou, 1967; Bird, 1972; Evans
& Perry, 2009). Knowledge on optimum or detrimental temperature regimes for survival of the
quarantine root-knot nematodes M. chitwoodi and M. fallax is essential for control while
preserving soil structure and nutrients.
Generally, in this study, it was observed that the survival of nematodes decreased with increase
in temperature, exposure time, type of treatment and vice versa. After heat treatment in an
incubator, survival of M. chitwoodi and M. fallax was highest at 40°C (≥ 88%) and least at 50°C
(0% survival) for J2s. This is not surprising because higher temperature has been proven to
lower the survival of nematodes in many studies (Sutherland & Sluggett, 1974; Geden & Axtell,
1988; Kung et al., 1991). Bridge (1996) observed that most treatments calls for temperatures
from 50 to 55°C to cause complete mortality of nematodes. For M. incognita, 60°C was required
to obtain 100% mortality. This might be due to the fact that as tropical species, M. incognita
may possessed a more tolerant heat resistant protein which protects the organism against
slightly elevated temperature. However, Tsai (2008) noted exposure time as being vital to cause
mortality of M. incognita at lower temperatures. He observed that 6 days was required to kill
J2 of M. incognita at 40°C (100% mortality). This also explains why ≥88% of quarantine
nematodes survived temperature at 40°C for 1 or 2 hours, noting time as an important factor.
Moreover, with the onset of global warming, it is possible that the temperate quarantine species
might be adjusting with a more heat tolerant protein as an adaptation for survival.
In this study, warm water bath treatment appears to be more effective than incubator treatment.
Least survival was attained at 40°C (0%) for M. fallax (50°C for incubator treatment) and 1.21%
survival was observed for M. chitwoodi for 1 hour. However, with increased in time (2 hours),
survival of M. chitwoodi decreased to 0% (50°C for incubator treatment). The same is true with
M. incognita where 0% survival was attained at 50°C as opposed to 60°C incubator treatment.
Earlier studies have reported decreased survival of J2s with time as well as decreased survival
of J2s with higher temperature but shorter time (Qiu et al., 1993; Tsai, 2008). Wang and
McSorley (2008) compared the exposure time to lethal temperatures for M.

15
incognita suppression with both warm water and soil solarization. They found that the hours of
exposure above a specific temperature to kill nematodes in soil solarization exceeded the
minimum number of hours required to kill 100% of M. incognita eggs or J2s as determined in
the warm water bath experiment. It is likely that warm water bath is more efficient because
water as the solvent, has higher heat capacity and thermal conductivity than air, and remains in
continuous contact with the test tube inoculated with nematodes, as such heat absorb by the
water are not easily dissipated.
For egg masses, temperature has a direct impact on the hatching of juveniles with decreasing
hatching percentages when temperatures are above the optimum for hatching (Bird, 1972;
Inserra et al., 1983; Charchar & Santo, 2001). In this study, the maximum hatching percentage
was observed at 40°C for all nematodes in both incubator and warm water bath experiments
while the minimum was noted at 60°C for M. incognita in incubator treatment and 50°C for M.
chitwoodi and M. fallax. For both experiments, M. fallax had higher hp than M. chitwoodi while
M. incognita had highest hp. However, Inserra et al. (1983) and Khan et al. (2014) observed
higher hp of M. chitwoodi than M. fallax at 20°C for 23 days and attributed the observed
differences to the protective role of the egg. Moreover, eggs used in this study were collected
from egg masses of approximately the same age, they may have differed in their embryonic
development (Khan et al., 2014).
The time at which 50% hatched was reach was similar for all species incubated for 1 hour
(approx. 2.5 weeks) at 40°C. However, with the exception of M. incognita at 40°C warm water
treatment where m was similar to incubator treatment at 40°C for 1 hour, delays in hatching
were noted for warm water bath treatment and species incubated for 2 hours. Charchar and
Santo (2001), Morris et al. (2011) and Khan et al. (2014) observed delayed hatching at lower
temperatures. However, this delay might be interpreted as egg being in diapause in response to
adverse condition. On the other hand, some eggs might not be viable as a result of extreme
temperature above their normal threshold which they can tolerate.
CONCLUSION
From the study, it was concluded that temperature treatment has an effect on the survival and
hatching percentage of M. chitwoodi, M. fallax and M. incognita. Survival decreased (hp
decreased) with increased temperature and time duration. The tropical species M. incognita was
more resistant to heat treatment than its temperate counterparts M. chitwoodi and M. fallax.
Moreover, warm water treatment was more efficient than incubator treatment and required

16
temperature at 40°C to cause complete mortality for M. chitwoodi and M. fallax whereas 50°C
was required for incubator treatment to attain 0% survival. However, M. incognita required
50°C for warm water treatment and 60°C for incubator treatment to achieve 0% survival.
RECOMMENDATIONS
Since warm water treatment is more efficient than incubator treatment, this treatment should
often be used in a processing industry to save time and energy and 50°C should be the optimum
temperature limit. In addition, this study did not take into account whether there is a difference
in hatching percentage between the warming up regime and a fixed temperature regime. Further
studies are necessary to examine these differences.
ACKNOWLEDGEMENTS
I heartily thank Almighty God for the grace, wisdom and good health granted to me throughout
this study.
I am immensely grateful to my promoter Prof. dr. Wim Wesemael for his prompt response,
constructive comments, critical review and closely following up of my research despite his busy
schedule. I will like to say working with him has been a privilege for me. My very earnest and
profound gratitude goes to Mr & Mrs Ojong Arrrey and Mr & Mrs Enoh Jean-Pierre who made
it possible through their spiritual, moral and financial support for my entire study.
Special thanks and gratitude to the Institute for Agricultural and Fisheries Research (ILVO) for
allowing me carryout my thesis and use their facilities throughout my research work. Also,
special thanks to the staff of ILVO for the pleasant atmosphere at the Institute. I want to say it
was a great opportunity working with them.
A word of gratitude goes to Nancy De Sutter for her guidance and helpful demonstrations in
the laboratory, Stephanie Beelaert who has always been there for helpful assistance and
directions throughout my work.
Finally, many thanks go to my entire family particularly my parents for their love and care.
Special thanks also go to Mr. & Mrs. Teboh for their material and financial assistance and not
given up on me.

17
REFERENCES
Akhtar, M. (1993). Utilisation of plant-origin waste materials for the control of plant-parasitic
nematodes. Bioresource technology 46, 255-257.
Baermann, G. (1917). Eine einfache methode zur auffindung von ankylostomum (Nematoden)
larven in erdproben. Geneeskunding Tijdschrift voor Nederlandsch-Indië 57, 131-137.
Bird, A. F. (1972). Influence of temperature on embryogenesis in Meloidogyne javanica.
Journal of nematology 4, 206.
Bridge, J. (1996). Nematode management in sustainable and subsistence agriculture. Annual
Review of Phytopathology 34, 201-225.
Charchar, J. M. & Santo, G. S. (2001). Effect of temperature on the embryogenic
development and hatching of Meloidogyne chitwoodi races 1 and 2 and M. hapla.
Nematologia brasileira 25, 71-77.
Davide, R. & Triantaphyllou, A. (1967). Influence of the environment on development and
sex differentiation of root-knot nematodes. Nematologica 13, 102-110.
De Waele, D. & Elsen, A. (2007). Challenges in tropical plant nematology. Annu. Rev.
Phytopathol. 45, 457-485.
Evans, A. A. & Perry, R. N. (2009). 9 Survival Mechanisms. Root-knot nematodes, 201.
Gamon, A. & Lenne, N. (2012). Meloidogyne chitwoodi and Meloidogyne fallax in France:
initial management experiences. EPPO Bulletin 42, 122-126.
Geden, C. & Axtell, R. (1988). Effect of temperature on nematode (Steinernema feltiae
[Nematoda: Steinernematidae]) treatment of soil for control of lesser mealworm
(Coleoptera: Tenebrionidae) in turkey houses. Journal of economic entomology 81,
800-803.
Hendrickx, G. (1995). An automatic apparatus for extracting free-living nematode stages from
soil. Nematologica 41, 308.
Hodda, M. & Cook, D. (2009). Economic impact from unrestricted spread of potato cyst
nematodes in Australia. Phytopathology 99, 1387-1393.
Hooper, D. (1986). Extraction of nematodes from plant material. Laboratory methods for
work with plant and soil nematodes. London: Her Majesty’s Stationery Office, 51-58.
Inserra, R., Griffin, G. & Sisson, D. (1983). Effects of temperature and root leachates on
embryogenic development and hatching of Meloidogyne chitwoodi and M. hapla.
Journal of nematology 15, 123.

18
Khan, A., Wesemael, W. M., Moes, M. (2014). Influence of temperature on the development
of the temperate root-knot nematodes Meloidogyne chitwoodi and M. fallax. Russian
journal of nematology 22 (1), 1-9.
Kung, S.-P., Gaugler, R. & Kaya, H. K. (1991). Effects of soil temperature, moisture, and
relative humidity on entomopathogenic nematode persistence. Journal of Invertebrate
Pathology 57, 242-249.
Mcneill, M., Phillips, C., Bell, N. & Proffitt, J. (2006). Potential spread of pests in New
Zealand through commercial transport of nursery plants. New Zealand Plant
Protection 59, 75.
Morris, K. S., Horgan, F. G., Downes, M. J. & Griffin, C. T. (2011). {null= The effect of
temperature on hatch and activity of second-stage juveniles of the root-knot nematode,
Meloidogyne minor, an emerging pest in north-west Europe, en= The effect of
temperature on hatch and activity of second-stage juveniles of the root-knot
nematode,< i> Meloidogyne minor</i>, an emerging pest in north-west Europe}.
Nematology 13, 985-993.
Nicol, J. M., Turner, S. J., Coyne, D., Den Nijs, L., Hockland, S. & Maafi, Z. T. (2011).
Current nematode threats to world agriculture. Genomics and molecular genetics of
plant-nematode interactions. Springer, pp. 21-43.
OEPP/EPPO, (2004). Post-entry quarantine for potato. OEPP/EPPO Bulletin 34, 443-454.
OEPP/EPPO, (2009). Meloidogyne chitwoodi and Meloidogyne fallax. OEPP/EPPO Bulletin
39, 5-17.
Oka, Y., Koltai, H., Bar‐Eyal, M., Mor, M., Sharon, E., Chet, I. & Spiegel, Y. (2000). New
strategies for the control of plant‐parasitic nematodes. Pest Management Science 56,
983-988.
Perry, R. (1999). Desiccation survival of parasitic nematodes. Parasitology 119, S19-S30.
Qiu, J., Westerdahl, B., Giraud, D. & Anderson, C. (1993). Evaluation of hot water treatments
for management of Ditylenchus dipsaci and fungi in daffodil bulbs. Journal of
nematology 25, 686.
Singh, S., Hodda, M. & Ash, G. (2013). Plant‐parasitic nematodes of potential phytosanitary
importance, their main hosts and reported yield losses. EPPO Bulletin 43, 334-374.
Soliman, T., Mourits, M. C., Van Der Werf, W., Hengeveld, G. M., Robinet, C. & Lansink,
A. G. O. (2012). Framework for modelling economic impacts of invasive species,
applied to pine wood nematode in Europe.

19
Suffert, M. & Giltrap, N. (2012). EPPO Workshop on Meloidogyne chitwoodi and
Meloidogyne fallax (2011‐03‐08): importance for potato production and experience of
management in EPPO countries. EPPO Bulletin 42, 117-121.
Sutherland, J. & Sluggett, L. (1974). Time, temperature and soil moisture effects on
Xiphinema bakeri nematode survival in fallow soil.
Tsai, B. Y. (2008). Effect of temperature on the survival of Meloidogyne incognita. Plant
Pathology Bulletin 17, 203-208.
Van der Gaag DJ, Viaene N, Anthoine A, Ilieva Z, Karssen G, Niere B (2012) Pest Risk
Assessment of Meloidogyne fallax: Revised Test method 2b. In: Pest Risk Assessment
for the European Community Plant Health: A Comparative Approach with Case
Studies (Eds MacLeod A, et al.), pp. 1053. http://www.efsa.europa.eu/en/
supporting/doc/319e.pdf [accessed on 13 November 2013].
Wallace, H. R. (1963). The biology of plant parasitic nematodes. The biology of plant
parasitic nematodes.
Wang, K.-H. & Mcsorley, R. (2008). Exposure time to lethal temperatures for Meloidogyne
incognita suppression and its implication for soil solarization. Journal of nematology
40, 7.
Wesemael, W. M., Perry, R. N. & Moens, M. (2006). The influence of root diffusate and host
age on hatching of the root-knot nematodes, Meloidogyne chitwoodi and M. fallax.
Nematology 8, 895-902.
Wesemael, W. M., Viaene, N. & Moens, M. (2011). Root-knot nematodes (Meloidogyne
spp.) in Europe. Nematology 13, 3-16.