survival and ecological effects of genetically engineered erwinia carotovora in soil and aquatic...
TRANSCRIPT

650
Environmental Toxicology and Chemistry, Vol. 16, No. 4, pp. 650–657, 1997q 1997 SETAC
Printed in the USA0730-7268/97 $6.00 1 .00
SURVIVAL AND ECOLOGICAL EFFECTS OF GENETICALLY ENGINEEREDERWINIA CAROTOVORA IN SOIL AND AQUATIC MICROCOSMS
SARAH PALMER,† VJERA S. SCANFERLATO,† DAVID R. ORVOS,† GEORGE H. LACY‡ and JOHN CAIRNS, JR.*††Department of Biology and ‡Department of Plant Pathology,
Physiology and Weed Science, Virginia Polytechnic Institute and State University,Blacksburg, Virginia 24061, USA
(Received 30 January 1996; Accepted 30 August 1996)
Abstract—The release of genetically engineered organisms into the environment has raised concerns about their potential ecologicalimpact. In this study, genetically engineered Erwinia carotovora strains expressing varying levels of reduced phytopathogenicityand wildtype E. carotovora strains were used in aquatic and soil microcosms to assess the survival, intraspecific competition, andeffects upon specific groups of indigenous bacteria. In aquatic microcosms, the densities of Erwinia genetically engineered organisms(GEMs) and wildtype strains declined and fell below the detectable limit of plate counts 15 d after the microcosms were inoculated.In aquatic microcosms, engineered E. carotovora did not exhibit a competitive advantage over the wildtype. The effect of engineeredand wildtype E. carotovora on densities of total and selected bacteria was not significantly different. Treatment with engineeredE. carotovora did not change biomass values of the receiving community but did cause a transitory increase in metabolic activity.In aquatic microcosms, the inability of genetically engineered E. carotovora to persist, displace resident species, or affect themetabolic activity of aquatic communities indicates the low risk of adverse ecological consequences in aquatic ecosystems. Unlikeprevious investigations involving soil microcosms, densities of both the genetically engineered and wildtype E. carotovora remainedat detectable levels over 60 d in both agricultural clay and forest loam soils. The type of soil significantly affected the survival ofthe GEM and the wildtype. The sorptive properties of clay particles, as well as low concentrations of soil nutrients and organicmatter, may have contributed to the unexpected patterns of GEM and wildtype survival.
Keywords—Genetically engineered microorganisms Risk assessment
INTRODUCTION
The widespread use of chemical pesticides and fertilizersand the accumulation of these and other nonbiodegradableproducts have resulted in extensive air, soil, and water pol-lution [1]. The long-term ecological implications of chemicaland consumptive resource wastes are only beginning to be-come apparent. Increased awareness of such ecological per-turbations has heightened the public’s acceptance and supportof biological control agents and bioremediation organisms de-veloped through advances in recombinant DNA technologyfor use in industrial and agricultural applications.
However, like chemicals, the introduction of engineeredorganisms into environments perturbed or stressed by pollutionor other anthropogenic activities may have unpredictable re-sults such as organism dissemination and successful coloni-zation beyond the release site. Environments that are partic-ularly vulnerable to exogenous organismal introductions arethose that are isolated, disturbed, simplified, and/or stressed[2]. Furthermore, genetically engineered microorganisms(GEMs) introduced into an ecosystem will interact with res-ident microbial populations [3]. Intraspecific competition be-tween the engineered and indigenous wildtype organisms mayfavor the establishment of the former, especially if it is eco-logically more successful in a specific environmental niche[4]. Additionally, interspecific competition may cause changesin natural microbial communities. The potential for displace-ment of bacterial populations, which play major roles in or-ganic matter mineralization, is considered particularly dele-
* To whom correspondence may be addressed.
terious in aquatic ecosystems where large amounts of biomassand organic carbon exist in the form of detritus [5]. Disruptionof mineralization processes may lead to changes in the pro-ductivity of aquatic ecosystems [6,7]. Microcosm studies havebeen effective in first-tier risk assessment fate-and-effect in-vestigations to evaluate potential ecological impacts [8]. Theuse of microcosms in preliminary studies of GEMs minimizesthe probability and severity of adverse ecological impacts thatmay occur during field trials.
In these investigations, a battery of genetically engineeredErwinia carotovora subspecies carotovora strains were usedas models in aquatic and soil microcosms. Erwinia carotovorasubspecies carotovora is a phytopathogenic enterobacteriumresponsible for soft rot in many agricultural plant tissues [9].Erwinia carotovora produces a battery of enzymes (pectatelyases, cellulases, phospholipases, and proteases) that are re-sponsible for plant tissue breakdown [10]. Although E. car-otovora is not indigenous to soil and aquatic environments, itis universally present in aquatic and terrestrial ecosystems [11–13]. Concentrations of erwiniae greater than 107 cells/ml havebeen found in surface waters receiving agricultural runoff (G.Lacy, unpublished data). Soil composition may play a key rolein the survival and ultimate distribution of Erwinia GEMs inaquatic systems. Because of the ubiquity and pathogenicity ofthe wildtype, genetically altered strains of erwiniae have po-tential ecological and economic importance as biological con-trol agents if they are able to survive and outcompete wildtypestrains.
Previous studies by Orvos and colleagues [14] and Scan-ferlato and colleagues [15] addressed the survival, competi-

Survival and effects of genetically engineered Erwinia Environ. Toxicol. Chem. 16, 1997 651
Table 1. Wildtype and genetically engineered Erwinia carotovora strains used to assess survival, competition, and effects on indigenousbacteria in aquatic and soil microcosms. References provide information on the genetic constructs of the genetically engineered (GEM) strains
StrainPheno-
type Genotype Assessment EnvironmentRefer-ences
L-543 w1 Wildtype with two endopectate lyases, pI 9.5, 10.4 Survival Soil [16]L-833 GEM Wildtype L-543 with disarmed extracellular pectat lyase and
kanamycin resistance (KanR)Competition Water and sediment [9,16]
L-863 w1 Spontaneous rifampin mutant of wildtype L-543 (RifR) Survival, competition, effecton receiving community
Water and sediment [9,16]
L-864 GEM Spontaneous rifampin mutant of GEM L-833 (RifR andKanR)
Survival, effect on receivingcommunity
Water and sediment [9,14]
L-872 GEM Deficient in a pI 10.4 endo-pectate lyase gene (KanR) Survival Soil [17]
tion, and effects on indigenous bacterial communities of ge-netically engineered E. carotovora in a single loam soil type.The present study is an extension of that work and expandsupon the findings of GEM survival, competition, and effectson indigenous organisms while taking into account the extentof genetic manipulation of the GEM strain. The present studyevaluates selected E. carotovora GEMs in different soil typesand profiles the response of selected E. carotovora GEMs inaquatic environments. The following hypotheses were testedto determine the microbial ecology and potential risk of ge-netically engineered E. carotovora in aquatic and terrestrialsystems: (1) engineered strains of E. carotovora are less fitthan the wildtype, (2) when introduced into an environmentalmatrix, both engineered and wildtype E. carotovora fail tocompete with the indigenous organisms present and do notnegatively impact the biomass or metabolic activity of thereceiving community, (3) when introduced into the same ma-trix, neither engineered nor wildtype E. carotovora is morecompetitive, and (4) the type of ecosystem and its conditionwill affect the length of survival time of the introduced en-gineered E. carotovora. To test these hypotheses, we deter-mined the ability of a genetically engineered strain to survivein the water and sediment of aquatic microcosms, the effectof GEMs on specific groups of indigenous bacteria, the resultof intraspecific competition between the GEM and the wild-type, the effect of GEMs on biomass and metabolic activityof the receiving community, and the survival in different soiltypes, one a clay soil of agricultural origins and the other aloam-based forest soil.
MATERIALS AND METHODS
Bacterial strains and plasmids
Table 1 summarizes the E. carotovora subsp. carotovorastrains, their phenotypes, genotypes, and the combinations ofGEM and wildtype Erwinia used in the aquatic and soil mi-crocosm studies. Wildtype E. carotovora subsp. carotovorastrain L-543 and a spontaneous rifampin (150 mg/ml)-resistantmutant (L-863) were used in the soil and aquatic microcosmstudies, respectively. The wildtype strains have two endo-pec-tate lyases focusing isoelectrically at pI 9.5 and 10.4. Genet-ically engineered E. carotovora L-833 was used in the aquaticmicrocosms and was constructed from the wildtype by partiallyremoving in vitro an extracellular pectate lyase gene on aplasmid (pDR40) by deletion mutation, inserting a 1.4-kb DNAfragment from Tn903 conferring kanamycin resistance, andchromosomally inserting, by gene replacement, part of theplasmid (pSV41) into the chromosome [9,16]. This strain,L-833, has a 30% reduction in its ability to rot potato tubertissue compared to the wildtype [9]. Strain L-864, used in the
aquatic microcosms, is a spontaneous rifampin (150 mg/ml)-resistant mutant of L-833 that is also kanamycin resistant[9,14].
Genetically engineered E. carotovora strain L-872 was se-lected for soil microcosm studies because it has the greatestreduction in pathogenicity of the engineered strains to date. Adetailed account of the construction of L-872 is given in Lacyand Stromberg [17]. Briefly, constructed plasmid pDR40 hasgenes mediating the production of two endo-pectate lyases thatfocus isoelectrically at pI 9.5 and pI 10.4. A 5.5-kb fragmentwas cleaved from pDR40 to form pDR41, which is missing558 bases of the pI 9.5 gene and all of the pI 10.4 gene. Anantibiotic resistance marker was added to pDR41 by ligationof a 1.4-kb fragment of pUC4K that carried kanamycin resis-tance from Tn903, thereby creating pVS20. Plasmid pVS20transformed into Escherichia coli was transferred to E. car-otovora by conjugation in triparental matings. Recombinantswere selected by phosphate starvation and selective platingfor kanamycin resistance. Mutants defective in pectate lyaseproduction were screened by isoelectric focusing, and an en-gineered E. carotovora strain L-872, deficient in a pI 10.4endo-pectate lyase gene, was identified.
Culture conditions
Cultures of genetically engineered and wildtype E. caro-tovora used in the aquatic microcosms were grown in nutrientbroth to a density of approximately 109 colony-forming units(CFU)/ml as estimated turbidimetrically at 550 nm. Engineeredand wildtype cultures used in the soil microcosms were grownin 2 L of nutrient broth (Difco Laboratories, Detroit, MI, USA)for 24 h at 308C to an absorbance of approximately 1.0 at 550nm. All cells were harvested by centrifugation at 10,000 g andwashed twice in sterile distilled water (SDW). Cells for thesoil microcosms were resuspended in 45 ml of SDW to deliver15 ml of inoculum to each microcosm (200 g soil), providingapproximately 108 cells/g soil.
Media and enumeration
All media and agar were purchased from Difco Laborato-ries. Samples from aquatic microcosms were serially dilutedand plated on appropriate media. Engineered strain L-833 wasenumerated using plate count agar (PCA) fortified with 40mg/ml kanamycin monosulfate. Densities of engineered strainL-864 were determined on PCA amended with 40 mg/ml kana-mycin monosulfate and 150 mg/ml rifampin. Wildtype strainL-863 was enumerated on PCA amended with 150 mg/ml rif-ampin. Total bacteria were enumerated on 1/10 PCA (2.35 g/LPCA with 13.5 g/L Bacto-agar). This concentration of mediawas determined to be optimal for recovery of environmentally
652 Environ. Toxicol. Chem. 16, 1997 S. Palmer et al.
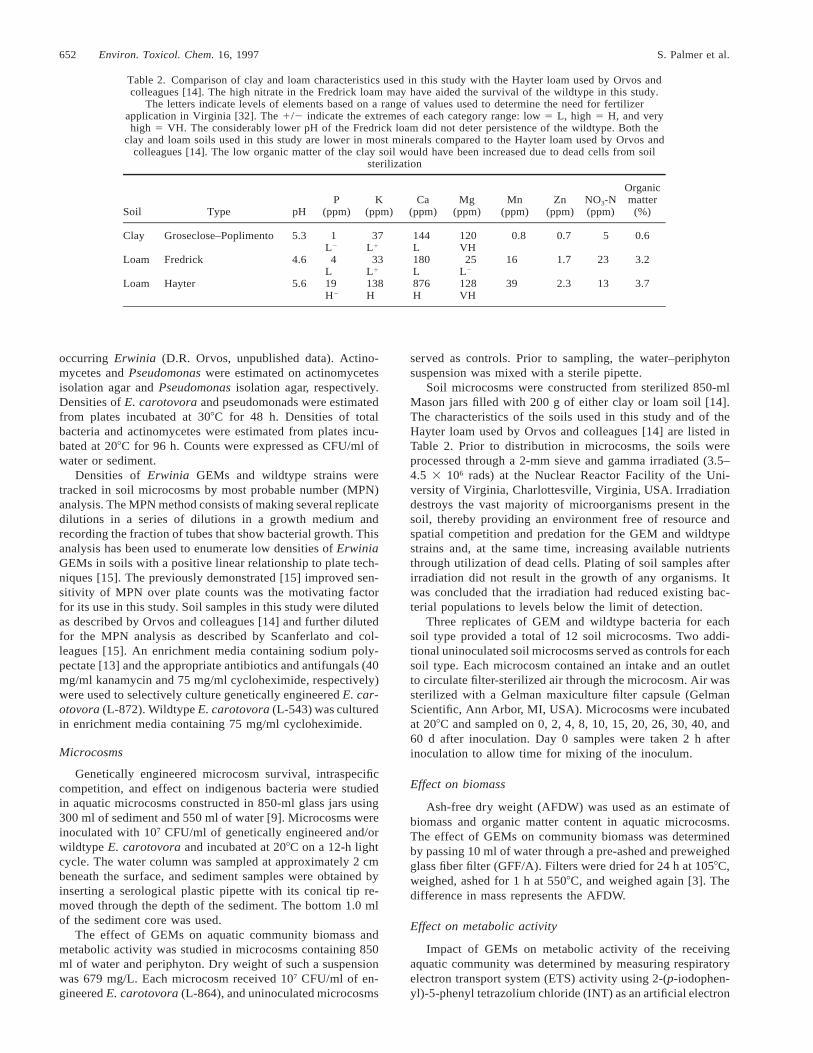
Table 2. Comparison of clay and loam characteristics used in this study with the Hayter loam used by Orvos andcolleagues [14]. The high nitrate in the Fredrick loam may have aided the survival of the wildtype in this study.
The letters indicate levels of elements based on a range of values used to determine the need for fertilizerapplication in Virginia [32]. The 1/2 indicate the extremes of each category range: low 5 L, high 5 H, and veryhigh 5 VH. The considerably lower pH of the Fredrick loam did not deter persistence of the wildtype. Both the
clay and loam soils used in this study are lower in most minerals compared to the Hayter loam used by Orvos andcolleagues [14]. The low organic matter of the clay soil would have been increased due to dead cells from soil
sterilization
Soil Type pHP
(ppm)K
(ppm)Ca
(ppm)Mg
(ppm)Mn
(ppm)Zn
(ppm)NO3-N(ppm)
Organicmatter
(%)
Clay
Loam
Loam
Groseclose–Poplimento
Fredrick
Hayter
5.3
4.6
5.6
1L2
4L19H2
37L1
33L1
138H
144L180L876H
120VH
25L2
128VH
0.8
16
39
0.7
1.7
2.3
5
23
13
0.6
3.2
3.7
occurring Erwinia (D.R. Orvos, unpublished data). Actino-mycetes and Pseudomonas were estimated on actinomycetesisolation agar and Pseudomonas isolation agar, respectively.Densities of E. carotovora and pseudomonads were estimatedfrom plates incubated at 308C for 48 h. Densities of totalbacteria and actinomycetes were estimated from plates incu-bated at 208C for 96 h. Counts were expressed as CFU/ml ofwater or sediment.
Densities of Erwinia GEMs and wildtype strains weretracked in soil microcosms by most probable number (MPN)analysis. The MPN method consists of making several replicatedilutions in a series of dilutions in a growth medium andrecording the fraction of tubes that show bacterial growth. Thisanalysis has been used to enumerate low densities of ErwiniaGEMs in soils with a positive linear relationship to plate tech-niques [15]. The previously demonstrated [15] improved sen-sitivity of MPN over plate counts was the motivating factorfor its use in this study. Soil samples in this study were dilutedas described by Orvos and colleagues [14] and further dilutedfor the MPN analysis as described by Scanferlato and col-leagues [15]. An enrichment media containing sodium poly-pectate [13] and the appropriate antibiotics and antifungals (40mg/ml kanamycin and 75 mg/ml cycloheximide, respectively)were used to selectively culture genetically engineered E. car-otovora (L-872). Wildtype E. carotovora (L-543) was culturedin enrichment media containing 75 mg/ml cycloheximide.
Microcosms
Genetically engineered microcosm survival, intraspecificcompetition, and effect on indigenous bacteria were studiedin aquatic microcosms constructed in 850-ml glass jars using300 ml of sediment and 550 ml of water [9]. Microcosms wereinoculated with 107 CFU/ml of genetically engineered and/orwildtype E. carotovora and incubated at 208C on a 12-h lightcycle. The water column was sampled at approximately 2 cmbeneath the surface, and sediment samples were obtained byinserting a serological plastic pipette with its conical tip re-moved through the depth of the sediment. The bottom 1.0 mlof the sediment core was used.
The effect of GEMs on aquatic community biomass andmetabolic activity was studied in microcosms containing 850ml of water and periphyton. Dry weight of such a suspensionwas 679 mg/L. Each microcosm received 107 CFU/ml of en-gineered E. carotovora (L-864), and uninoculated microcosms
served as controls. Prior to sampling, the water–periphytonsuspension was mixed with a sterile pipette.
Soil microcosms were constructed from sterilized 850-mlMason jars filled with 200 g of either clay or loam soil [14].The characteristics of the soils used in this study and of theHayter loam used by Orvos and colleagues [14] are listed inTable 2. Prior to distribution in microcosms, the soils wereprocessed through a 2-mm sieve and gamma irradiated (3.5–4.5 3 106 rads) at the Nuclear Reactor Facility of the Uni-versity of Virginia, Charlottesville, Virginia, USA. Irradiationdestroys the vast majority of microorganisms present in thesoil, thereby providing an environment free of resource andspatial competition and predation for the GEM and wildtypestrains and, at the same time, increasing available nutrientsthrough utilization of dead cells. Plating of soil samples afterirradiation did not result in the growth of any organisms. Itwas concluded that the irradiation had reduced existing bac-terial populations to levels below the limit of detection.
Three replicates of GEM and wildtype bacteria for eachsoil type provided a total of 12 soil microcosms. Two addi-tional uninoculated soil microcosms served as controls for eachsoil type. Each microcosm contained an intake and an outletto circulate filter-sterilized air through the microcosm. Air wassterilized with a Gelman maxiculture filter capsule (GelmanScientific, Ann Arbor, MI, USA). Microcosms were incubatedat 208C and sampled on 0, 2, 4, 8, 10, 15, 20, 26, 30, 40, and60 d after inoculation. Day 0 samples were taken 2 h afterinoculation to allow time for mixing of the inoculum.
Effect on biomass
Ash-free dry weight (AFDW) was used as an estimate ofbiomass and organic matter content in aquatic microcosms.The effect of GEMs on community biomass was determinedby passing 10 ml of water through a pre-ashed and preweighedglass fiber filter (GFF/A). Filters were dried for 24 h at 1058C,weighed, ashed for 1 h at 5508C, and weighed again [3]. Thedifference in mass represents the AFDW.
Effect on metabolic activity
Impact of GEMs on metabolic activity of the receivingaquatic community was determined by measuring respiratoryelectron transport system (ETS) activity using 2-(p-iodophen-yl)-5-phenyl tetrazolium chloride (INT) as an artificial electron
Survival and effects of genetically engineered Erwinia Environ. Toxicol. Chem. 16, 1997 653
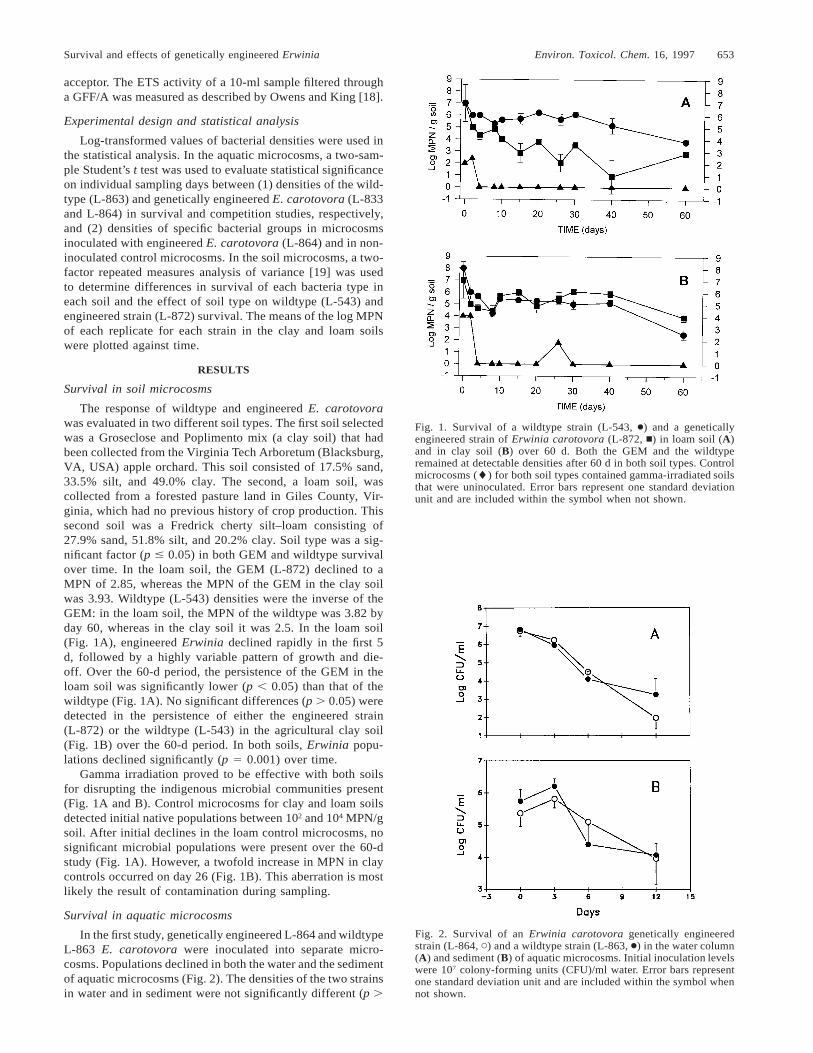
Fig. 1. Survival of a wildtype strain (L-543, ●) and a geneticallyengineered strain of Erwinia carotovora (L-872, n) in loam soil (A)and in clay soil (B) over 60 d. Both the GEM and the wildtyperemained at detectable densities after 60 d in both soil types. Controlmicrocosms (l) for both soil types contained gamma-irradiated soilsthat were uninoculated. Error bars represent one standard deviationunit and are included within the symbol when not shown.
Fig. 2. Survival of an Erwinia carotovora genetically engineeredstrain (L-864, C) and a wildtype strain (L-863, ●) in the water column(A) and sediment (B) of aquatic microcosms. Initial inoculation levelswere 107 colony-forming units (CFU)/ml water. Error bars representone standard deviation unit and are included within the symbol whennot shown.
acceptor. The ETS activity of a 10-ml sample filtered througha GFF/A was measured as described by Owens and King [18].
Experimental design and statistical analysis
Log-transformed values of bacterial densities were used inthe statistical analysis. In the aquatic microcosms, a two-sam-ple Student’s t test was used to evaluate statistical significanceon individual sampling days between (1) densities of the wild-type (L-863) and genetically engineered E. carotovora (L-833and L-864) in survival and competition studies, respectively,and (2) densities of specific bacterial groups in microcosmsinoculated with engineered E. carotovora (L-864) and in non-inoculated control microcosms. In the soil microcosms, a two-factor repeated measures analysis of variance [19] was usedto determine differences in survival of each bacteria type ineach soil and the effect of soil type on wildtype (L-543) andengineered strain (L-872) survival. The means of the log MPNof each replicate for each strain in the clay and loam soilswere plotted against time.
RESULTS
Survival in soil microcosms
The response of wildtype and engineered E. carotovorawas evaluated in two different soil types. The first soil selectedwas a Groseclose and Poplimento mix (a clay soil) that hadbeen collected from the Virginia Tech Arboretum (Blacksburg,VA, USA) apple orchard. This soil consisted of 17.5% sand,33.5% silt, and 49.0% clay. The second, a loam soil, wascollected from a forested pasture land in Giles County, Vir-ginia, which had no previous history of crop production. Thissecond soil was a Fredrick cherty silt–loam consisting of27.9% sand, 51.8% silt, and 20.2% clay. Soil type was a sig-nificant factor (p # 0.05) in both GEM and wildtype survivalover time. In the loam soil, the GEM (L-872) declined to aMPN of 2.85, whereas the MPN of the GEM in the clay soilwas 3.93. Wildtype (L-543) densities were the inverse of theGEM: in the loam soil, the MPN of the wildtype was 3.82 byday 60, whereas in the clay soil it was 2.5. In the loam soil(Fig. 1A), engineered Erwinia declined rapidly in the first 5d, followed by a highly variable pattern of growth and die-off. Over the 60-d period, the persistence of the GEM in theloam soil was significantly lower (p , 0.05) than that of thewildtype (Fig. 1A). No significant differences (p . 0.05) weredetected in the persistence of either the engineered strain(L-872) or the wildtype (L-543) in the agricultural clay soil(Fig. 1B) over the 60-d period. In both soils, Erwinia popu-lations declined significantly (p 5 0.001) over time.
Gamma irradiation proved to be effective with both soilsfor disrupting the indigenous microbial communities present(Fig. 1A and B). Control microcosms for clay and loam soilsdetected initial native populations between 102 and 104 MPN/gsoil. After initial declines in the loam control microcosms, nosignificant microbial populations were present over the 60-dstudy (Fig. 1A). However, a twofold increase in MPN in claycontrols occurred on day 26 (Fig. 1B). This aberration is mostlikely the result of contamination during sampling.
Survival in aquatic microcosms
In the first study, genetically engineered L-864 and wildtypeL-863 E. carotovora were inoculated into separate micro-cosms. Populations declined in both the water and the sedimentof aquatic microcosms (Fig. 2). The densities of the two strainsin water and in sediment were not significantly different (p .
654 Environ. Toxicol. Chem. 16, 1997 S. Palmer et al.
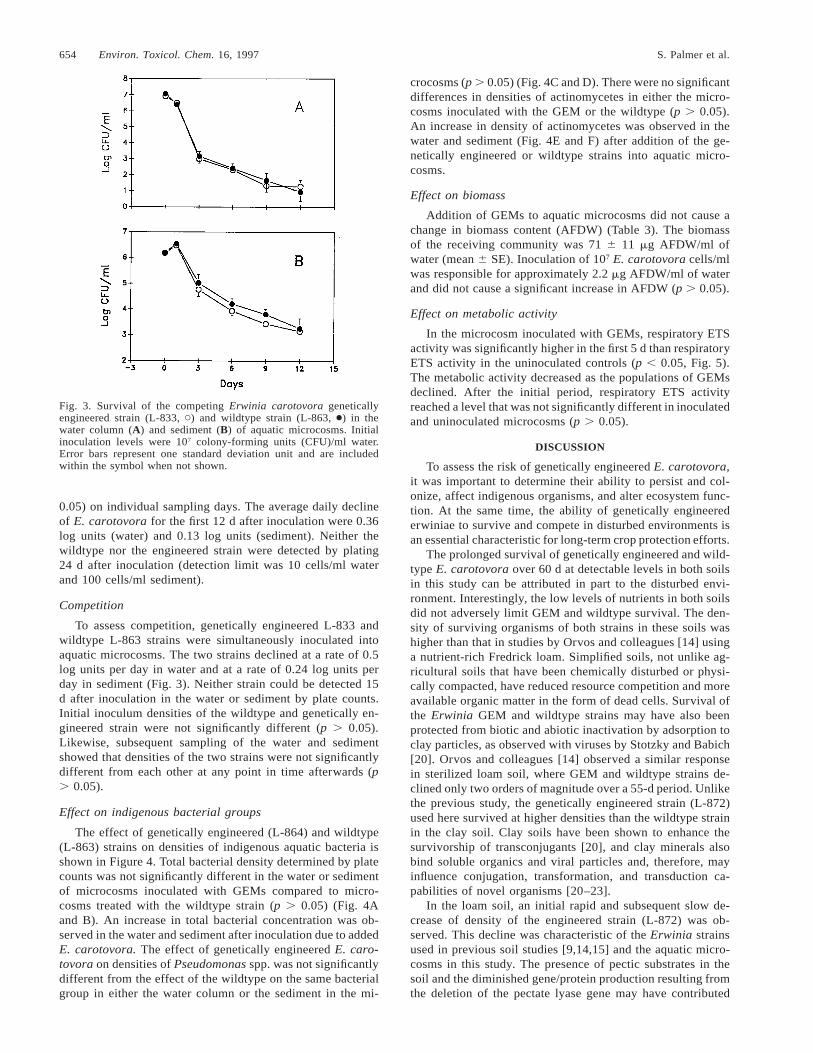
Fig. 3. Survival of the competing Erwinia carotovora geneticallyengineered strain (L-833, C) and wildtype strain (L-863, ●) in thewater column (A) and sediment (B) of aquatic microcosms. Initialinoculation levels were 107 colony-forming units (CFU)/ml water.Error bars represent one standard deviation unit and are includedwithin the symbol when not shown.
0.05) on individual sampling days. The average daily declineof E. carotovora for the first 12 d after inoculation were 0.36log units (water) and 0.13 log units (sediment). Neither thewildtype nor the engineered strain were detected by plating24 d after inoculation (detection limit was 10 cells/ml waterand 100 cells/ml sediment).
Competition
To assess competition, genetically engineered L-833 andwildtype L-863 strains were simultaneously inoculated intoaquatic microcosms. The two strains declined at a rate of 0.5log units per day in water and at a rate of 0.24 log units perday in sediment (Fig. 3). Neither strain could be detected 15d after inoculation in the water or sediment by plate counts.Initial inoculum densities of the wildtype and genetically en-gineered strain were not significantly different (p . 0.05).Likewise, subsequent sampling of the water and sedimentshowed that densities of the two strains were not significantlydifferent from each other at any point in time afterwards (p. 0.05).
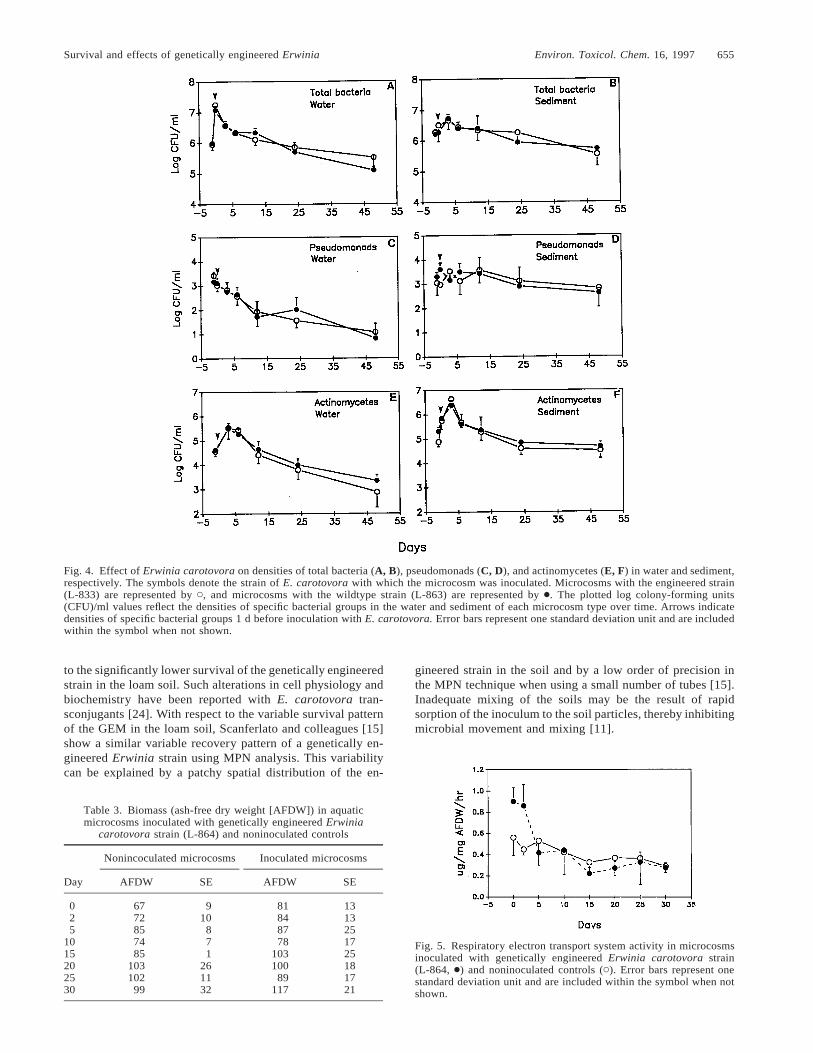
Effect on indigenous bacterial groups
The effect of genetically engineered (L-864) and wildtype(L-863) strains on densities of indigenous aquatic bacteria isshown in Figure 4. Total bacterial density determined by platecounts was not significantly different in the water or sedimentof microcosms inoculated with GEMs compared to micro-cosms treated with the wildtype strain (p . 0.05) (Fig. 4Aand B). An increase in total bacterial concentration was ob-served in the water and sediment after inoculation due to addedE. carotovora. The effect of genetically engineered E. caro-tovora on densities of Pseudomonas spp. was not significantlydifferent from the effect of the wildtype on the same bacterialgroup in either the water column or the sediment in the mi-
crocosms (p . 0.05) (Fig. 4C and D). There were no significantdifferences in densities of actinomycetes in either the micro-cosms inoculated with the GEM or the wildtype (p . 0.05).An increase in density of actinomycetes was observed in thewater and sediment (Fig. 4E and F) after addition of the ge-netically engineered or wildtype strains into aquatic micro-cosms.
Effect on biomass
Addition of GEMs to aquatic microcosms did not cause achange in biomass content (AFDW) (Table 3). The biomassof the receiving community was 71 6 11 mg AFDW/ml ofwater (mean 6 SE). Inoculation of 107 E. carotovora cells/mlwas responsible for approximately 2.2 mg AFDW/ml of waterand did not cause a significant increase in AFDW (p . 0.05).
Effect on metabolic activity
In the microcosm inoculated with GEMs, respiratory ETSactivity was significantly higher in the first 5 d than respiratoryETS activity in the uninoculated controls (p , 0.05, Fig. 5).The metabolic activity decreased as the populations of GEMsdeclined. After the initial period, respiratory ETS activityreached a level that was not significantly different in inoculatedand uninoculated microcosms (p . 0.05).
DISCUSSION
To assess the risk of genetically engineered E. carotovora,it was important to determine their ability to persist and col-onize, affect indigenous organisms, and alter ecosystem func-tion. At the same time, the ability of genetically engineerederwiniae to survive and compete in disturbed environments isan essential characteristic for long-term crop protection efforts.
The prolonged survival of genetically engineered and wild-type E. carotovora over 60 d at detectable levels in both soilsin this study can be attributed in part to the disturbed envi-ronment. Interestingly, the low levels of nutrients in both soilsdid not adversely limit GEM and wildtype survival. The den-sity of surviving organisms of both strains in these soils washigher than that in studies by Orvos and colleagues [14] usinga nutrient-rich Fredrick loam. Simplified soils, not unlike ag-ricultural soils that have been chemically disturbed or physi-cally compacted, have reduced resource competition and moreavailable organic matter in the form of dead cells. Survival ofthe Erwinia GEM and wildtype strains may have also beenprotected from biotic and abiotic inactivation by adsorption toclay particles, as observed with viruses by Stotzky and Babich[20]. Orvos and colleagues [14] observed a similar responsein sterilized loam soil, where GEM and wildtype strains de-clined only two orders of magnitude over a 55-d period. Unlikethe previous study, the genetically engineered strain (L-872)used here survived at higher densities than the wildtype strainin the clay soil. Clay soils have been shown to enhance thesurvivorship of transconjugants [20], and clay minerals alsobind soluble organics and viral particles and, therefore, mayinfluence conjugation, transformation, and transduction ca-pabilities of novel organisms [20–23].
In the loam soil, an initial rapid and subsequent slow de-crease of density of the engineered strain (L-872) was ob-served. This decline was characteristic of the Erwinia strainsused in previous soil studies [9,14,15] and the aquatic micro-cosms in this study. The presence of pectic substrates in thesoil and the diminished gene/protein production resulting fromthe deletion of the pectate lyase gene may have contributed
Survival and effects of genetically engineered Erwinia Environ. Toxicol. Chem. 16, 1997 655
Fig. 4. Effect of Erwinia carotovora on densities of total bacteria (A, B), pseudomonads (C, D), and actinomycetes (E, F) in water and sediment,respectively. The symbols denote the strain of E. carotovora with which the microcosm was inoculated. Microcosms with the engineered strain(L-833) are represented by C, and microcosms with the wildtype strain (L-863) are represented by ●. The plotted log colony-forming units(CFU)/ml values reflect the densities of specific bacterial groups in the water and sediment of each microcosm type over time. Arrows indicatedensities of specific bacterial groups 1 d before inoculation with E. carotovora. Error bars represent one standard deviation unit and are includedwithin the symbol when not shown.
Fig. 5. Respiratory electron transport system activity in microcosmsinoculated with genetically engineered Erwinia carotovora strain(L-864, ●) and noninoculated controls (C). Error bars represent onestandard deviation unit and are included within the symbol when notshown.
Table 3. Biomass (ash-free dry weight [AFDW]) in aquaticmicrocosms inoculated with genetically engineered Erwinia
carotovora strain (L-864) and noninoculated controls
Day
Nonincoculated microcosms
AFDW SE
Inoculated microcosms
AFDW SE
025
1015202530
6772857485
103102
99
910
871
261132
81848778
103100
89117
1313251725181721
to the significantly lower survival of the genetically engineeredstrain in the loam soil. Such alterations in cell physiology andbiochemistry have been reported with E. carotovora tran-sconjugants [24]. With respect to the variable survival patternof the GEM in the loam soil, Scanferlato and colleagues [15]show a similar variable recovery pattern of a genetically en-gineered Erwinia strain using MPN analysis. This variabilitycan be explained by a patchy spatial distribution of the en-
gineered strain in the soil and by a low order of precision inthe MPN technique when using a small number of tubes [15].Inadequate mixing of the soils may be the result of rapidsorption of the inoculum to the soil particles, thereby inhibitingmicrobial movement and mixing [11].

656 Environ. Toxicol. Chem. 16, 1997 S. Palmer et al.
The decline and disappearance of genetically engineeredand wildtype E. carotovora from the water column and sed-iment after inoculation into aquatic microcosms confirms theresults of previous investigations [15]. Similar decline curveswere observed in the competition experiment, in which thedensities of simultaneously introduced wildtype and GEMstrains declined at the same rate. A low level of nutrients andpredation were probably the factors most responsible for elim-ination of both the engineered and wildtype E. carotovorafrom the aquatic system [25,26]. The genetic alteration ap-parently did not affect the fitness and competitiveness of theengineered strain L-864 compared to the wildtype strain L-863in water and in sediment. The presence of plant tissue may beexpected to accentuate any difference between the two pop-ulations. However, a separate study demonstrated that genet-ically engineered E. carotovora increase in density at the samerate as in the wildtype on potato tuber tissue [27].
Although large densities of wildtype erwiniae that occa-sionally appear in natural surface waters originate from ter-restrial rotting plant tissue [28], it is doubtful if the externalsources can account for the continuous presence of E. caro-tovora [12]. Niches may exist in natural aquatic systems wherethe wildtype can survive for an extended period of time, suchas in stream bed sediments [29], or perhaps as pathogens orepiphytes on aquatic plants. Because of the similar physio-logical and ecological characteristics of the genetically engi-neered and wildtype E. carotovora, such habitats may be suit-able for the persistence of both strains.
After release of genetically engineered organisms, residentmicrobial communities will first respond to potential pertur-bation [30]. We monitored changes in densities of total bacteriaand densities of two bacterial groups involved in mineraliza-tion of organic matter—actinomycetes and Pseudomonas spp.In our aquatic microcosm experiments, the GEM did not dis-place these bacterial groups and did not produce microbialchanges in the water or sediment in a manner different fromthat of the wildtype.
Biomass and metabolic activity were monitored after in-oculation of GEMs into aquatic microcosms. Changes in bio-mass (AFDW) were not detected, but a significant increase inmetabolic activity of the system followed the inoculation ofGEMs. The absence of a noticeable increase in biomass canbe explained by the fact that the added 107 cells/ml representedthe addition of only 2.2 mg AFDW/ml. This amount is neg-ligible when compared to AFDW content in noninoculatedcontrol microcosms (71 mg AFDW/ml). On the other hand,the actively respiring E. carotovora cells caused a significantincrease in respiratory ETS activity when added to micro-cosms. This enzymatic activity decreased as the populationdensity decreased. These results support the findings of Mar-tinez and colleagues [31] that inactivity of metabolic enzymesinvolved in ETS reactions is associated with culturabilitylosses and cell lysis.
In the aquatic microcosms in this study, the GEMs did notoutcompete the wildtype strain, displace the indigenous bac-teria, or have a long-lasting effect on the biomass or metabolicactivity. In soil microcosms, both GEM and wildtype strainsdisplayed soil-dependent persistence patterns in two simplifiedagricultural and nonagricultural soil types. The unique re-sponse of the engineered Erwinia strains and wildtypes reflectsthe importance of subtle genetic and environmental variations.The behavior of novel organisms predicted from generalizedmicrocosm studies is not always valid in a risk assessment
model. Further examination of the fate and effects of GEMsin a wide range of environments (soils, sediments, and aerialdispersion; perturbed and nonperturbed systems) is warrantedin order to develop a comprehensive risk assessment. The vari-ation of GEM survival resulting from physio-biochemical al-terations warrants further attention, particularly in studies ofGEMs of commercial value.
Acknowledgement—These studies were supported by the E.I. DuPontde Nemours and Company Educational Foundation, the Mobil Foun-dation, National Science Foundation grant BSR-8705445, and Envi-ronmental Protection Agency grant R-813805-01-0. D.R. Orvos wassupported, in part, by the Society of Environmental Toxicology andChemistry Pre-Doctoral Fellowship, sponsored by the Procter & Gam-ble Company, and the Cunningham Dissertation Fellowship from Vir-ginia Tech. We are grateful to Julie Yim for excellent technical as-sistance, the Nuclear Reactor Facility at the University of Virginia,Charlottesville, Virginia, for sterilization of the soils, and Darla Don-ald for editorial preparation of the manuscript for publication.
REFERENCES
1. Hoy, M.A. 1991. Biotechnology and the environment, review.Science 253:89–90.
2. Sharples, F.E. 1983. Spread of organisms with novel genotypes:Thoughts from an ecological perspective. Recomb. DNA Tech.Bull. 6:43–56.
3. Alexander, M. 1971. Microbial Ecology. John Wiley & Sons,New York, NY, USA.
4. Stacey, G. 1985. The rhizobium experience. In H.O. Halvorson,D. Pramer and M. Rogul, eds., Engineered Organisms in theEnvironment: Scientific Issues. American Society for Microbi-ology, Washington, DC, pp. 109–121.
5. Wetzel, R.G. 1983. Limnology, 2nd ed. W.B. Saunders, Phila-delphia, PA, USA.
6. Tiedje, J.M., R.K. Colwell, Y.L. Grossman, R.E. Hodson, R.E.Lenski, R.N. Mack and P.J. Regal. 1989. The planned intro-duction of genetically engineered organisms: Ecological consid-erations and recommendations. Ecology 70:298–315.
7. Cairns, J., Jr. and J.R. Pratt. 1986. Factors affecting the ac-ceptance and rejection of genetically altered microorganisms byestablished natural aquatic communities. In T.M. Poston and R.Purdy, eds., Aquatic Toxicology and Environmental Fate: NinthSymposium, STP 921. American Society for Testing and Mate-rials, Philadelphia, PA, pp. 207–221.
8. Orvos, D.R. 1992. Assessing environmental risk from geneticallyengineered microorganisms and products containing recombinantDNA. In J. Cairns, Jr., B.R. Niederlehner and D.R. Orvos, eds.,Predicting Ecosystem Risk. Princeton Scientific, Princeton, NJ,USA, pp. 215–235.
9. Scanferlato, V.S., D.R. Orvos, G.H. Lacy and J. Cairns, Jr.1989. Genetically engineered Erwinia carotovora in aquatic mi-crocosms: Survival and effects on functional groups of indigenousbacteria. Appl. Environ. Microbiol. 55:1477–1482.
10. Kotoujansky, A. 1987. Molecular genetics of pathogenesis bysoft-rot erwiniaes. Annu. Rev. Phytopathol. 25:405–430.
11. Gannon, J.T., V.B. Manilal and M. Alexander. 1991. Rela-tionship between cell surface properties and transport of bacteriathrough soil. Appl. Environ. Microbiol. 57:190–193.
12. Maddox, D.A. and M.D. Harrison. 1988. Presence and popu-lation dynamics of Erwinia carotovora in irrigation water in southcentral Colorado. J. Appl. Bacteriol. 64:169–182.
13. Meneley, J.C. and M.E. Stanghellini. 1976. Isolation of soft-rotErwinia spp. from agricultural soils using an enrichment tech-nique. Phytopathology 66:367–370.
14. Orvos, D.R., G.H. Lacy and J. Cairns, Jr. 1990. Geneticallyengineered Erwinia carotovora: Survival, intraspecific compe-tition and effects upon selected bacterial genera. Appl. Environ.Microbiol. 56:1689–1694.
15. Scanferlato, V.S., G.H. Lacy and J. Cairns, Jr. 1990. Enu-merating low densities of genetically engineered Erwinia caro-tovora in soil. Lett. Appl. Microbiol. 10:55–59.
16. Allen, C., V.K. Stromberg, F.D. Smith, G.H. Lacy and M.S.Mount. 1986. Complementation of an Erwinia carotovora subsp.carotovora protease mutant with a protease-encoding cosmid.Mol. Gen. Genet. 202:276–279.

Survival and effects of genetically engineered Erwinia Environ. Toxicol. Chem. 16, 1997 657
17. Lacy, G.H. and V.K. Stromberg. 1990. Pre-release microcosmtests with genetically engineered plant pathogen. In D.R.MacKenzie and S.C. Henry, eds., International Symposium onthe Biosafety Results of Field Tests of Genetically ModifiedPlants and Microorganisms. Kiawah Island, SC, USA, November27–30, pp. 81–89.
18. Owens, T.G. and F.D. King. 1975. The activity measurement ofrespiratory electron-transport-system activity in marine zooplank-ton. Mar. Biol. 30:27–36.
19. Sokal, R.R. and F.J. Rohlf. 1981. Biometry. W.H. Freeman, NewYork, NY, USA.
20. Stotzky, G. and H. Babich. 1984. Fate of genetically engineeredmicrobes in natural environments. Recomb. DNA Tech. Bull. 7:163–188.
21. Stotzky, G. 1980. Surface interactions between clay minerals andmicrobes, viruses and soluble organics and the probable impor-tance of these interactions to the ecology of microbes in soil. InR.C.W. Berkeley, J.M. Lynch, J. Melling, P.R. Rutter and B. Vin-cent, eds., Microbial Adhesion to Surfaces. Ellis Harwood,Chichester, UK, pp. 231–247.
22. Stotzky, G. and V.N. Krasovsky. 1981. Ecological factors thataffect the survival, establishment, growth and genetic recombi-nation of microbes in natural habitats. In S.B. Levy, R.C. Clowesand E.L. Koenig, eds., Molecular Biology, Pathogenicity, andEcology of Bacterial Plasmids. Plenum, New York, NY, USA,pp. 31–42.
23. Suter, G.W., II. 1985. Application of environmental risk analysisto engineered organisms. In H.O. Halvorson, D. Pramer and M.Rogul, eds., Engineered Organisms in the Environment: Scien-tific Issues. American Society for Microbiology, Washington, DC,pp. 211–219.
24. Kozyrovskaya, N.A., R.I. Gvozdyak, V.A. Muras and V.A.Kordyum. 1984. Changes in properties of phytopathogenic bac-teria affected by plasmid pDR1. Arch. Microbiol. 137:338–343.
25. DeBoer, S.H. 1982. Survival of phytopathogenic bacteria in soil.In M.S. Mount and G.H. Lacy, eds., Phytopathogenic Prokary-otes, Vol. 1 Academic, New York, NY, USA, pp. 88–108.
26. Liang, L.N., J.L. Sinclair, L.M. Mallory and M. Alexander.1982. Fate in model ecosystems of microbial species of potentialuse in genetic engineering. Appl. Environ. Microbiol. 44:708–714.
27. Stromberg, V.K., D.R. Orvos, V.S. Scanferlato, G.H. Lacy andJ. Cairns, Jr. 1990. In planta competition among cell-degradingenzyme mutants and wildtype strains of Erwinia carotovora. InZ. Klement, ed., Plant Pathogenic Bacteria. Proceedings, 7thInternational Conference on Plant Pathogenic Bacteria, HungaryAcademy of Sciences, Budapest, Hungary, June 11–16, 1989, pp.721–725.
28. Lacy, G.H., R.C. Lamb and C.M. Berg. 1981. Iris soft rot causedby Erwinia chrysanthemi, associated with overhead irrigation andits control by chlorination. Proceedings, Southern Regional In-ternational Plant Propagator’s Society Annual Meeting, Houston,TX, USA, December 6–9, pp. 624–634.
29. Jorge, P.E. and M.D. Harrison. 1986. The association of Er-winia carotovora with surface water in northeastern Colorado. I.The presence and population of the bacterium in relation to lo-cation, season and water temperature. Am. Potato J. 63:517–531.
30. Brock, T. 1985. Prokaryotic population ecology. In H.O. Hal-verson, D. Pramer and M. Rogul, eds., Engineered Organismsin the Environment: Scientific Issues. American Society for Mi-crobiology, Washington, DC, pp. 177–179.
31. Martinez, J., J. Garcia-Lara and J. Vives-Rego. 1989. Esti-mation of Escherichia coli mortality in seawater by the decreasein 3H-label and electron transport system activity. Microb. Ecol.17:219–225.
32. Donohue, S.J. and G.W. Hawkins. 1979. Guide to computerprogrammed soil test recommendations in Virginia. CooperativeExtension Division Publication 834. Virginia Polytechnic Insti-tute and State University, Blacksburg, VA, USA.