supplementary materials for -...
TRANSCRIPT

www.sciencemag.org/cgi/content/full/science.1226743/DC1
Supplementary Materials for
Tricking the Guard: Exploiting Plant Defense for Disease Susceptibility
J. Lorang, T. Kidarsa, C. S. Bradford, B. Gilbert, M. Curtis, S.-C. Tzeng, C. S. Maier, T. J. Wolpert*
*To whom correspondence should be addressed. E-mail: [email protected]
Published 18 October 2012 on Science Express DOI: 10.1126/science.1226743
This PDF file includes: Materials and Methods
Figs. S1 to S6
References

Supporting Online Material Materials and Methods Agrobacterium-mediated transformation of Arabidopsis and transient expression in N. benthamiana
LOV1 was amplified by PCR from pXba1.5 (1) using the forward primer 5’-CACCATGGCTGAAGGAGTTGTGTTG-3’ and the reverse primer 5’-TTATTCGTCACAGTTGATAAATTGAAC-3’, and inserted into PENTR/D-Topo vector according to the manufacturer’s instructions (Invitrogen). The resulting construct, and TRX-h5 wildtype and mutant constructs, also in PENTR/D-Topo (8) were linearized and recombined into the following binary expression vectors using Gateway LR Clonase II enzyme mix (Invitrogen): for untagged proteins, pEarleyGate 100; for Myc-tagged proteins, pEarleyGate203 (18); and for bimolecular fluorescent complementation, pSITE-BIFC-C1nec, -1Ccec, -N1nen, and –N1cen (19). Recombination reactions were transformed into Escherichia coli strain DH5α, and the resulting clones were transformed into Agrobacterium tumefaciens strain GV3101 for Arabidopsis stable transformation and GV2260 for transient expression. Arabidopsis plants containing the attrx-h5-1 mutation in ecotype Cl-0 or wild-type Col-4 plants were transformed using the floral dip method (20) and putative transgenic seeds were planted in soil wet with 0.02% glufosinate-ammonium. For transient expression, overnight A. tumefaciens GV2260 cultures containing the appropriate constructs were harvested by centrifugation and resuspended in infiltration buffer (10 mM MES pH5.2, 10 mM MgCl, 150 µM Acetosyingone, Sigma) to a final density of 0.5 OD600, incubated at room temperature for 3 hours, and syringe-infiltrated into the youngest fully-expanded leaf of three-week-old N. benthamiana. For LOV1 and TRX-h5 co-expression, equal volumes of the cultures were mixed before infiltration. For victorin sensitivity assays, victorin or water was infiltrated 48 hours after A. tumefaciens infiltration. Pictures were taken 8 hours after Victorin treatment. Bimolecular fluorescence complementation and confocal microscopy
LOV1, RPS5 and TRX-h5 were cloned into pSITE-BIFC- C1nec, -C1cec, -N1nen, and –N1cen destination vectors (19) and transiently co-expressed in N. benthamiana, in all possible nYFP/cYFP combinations or with cYFP-GST and nYFP-GST control vectors, as described above. Images were taken 48 hours after A. tumefaciens infiltration using a Zeiss LSM 510 laser scanning confocal microscope (Carl Zeiss Jena GmbH, Jena, Germany) with an Argon laser and 488 nm excitation. YFP was collected using the beam splitter and filter configuration, HFT 488/543, NFT 490 and BP 505-530. Chloroplast autoflourescence was collected using the same configuration except a LP650 instead of BP 505-530 filter was used. The objective was a Plan-Neofluar 25x/0.8.
Insulin reduction assay of TRX-h5 activity
TRX-h5 activity was measured using insulin as a substrate according to Holmgren et al. (21). Reactions were performed in 63mM potassium phosphate buffer, pH 7.0, 2 mM EDTA, 0.1% insulin (w/v), and 7.5 µg / ml (approximately 0.05 µM) TRX-h5. Dithiothreitol was added to a final concentration of 1mM to initiate reactions, and activity

was measured by monitoring turbidity at 650 nm. Reactions were performed with and without 1-10 µM victorin.
TRX-h5 protein was isolated utilizing a His-tag. TRX-h5 was cloned into PENTR/SD/D-TOPO, recombined into pDEST 17 (both Invitrogen) and the resulting construct was transformed into E. coli BL21 (DE3). A 500 ml culture of transformed cells was grown to 0.5 OD600, induced with 2 µg/ml arabinose, and then allowed to grow for an additional 45 minutes at 37ºC. Cells were harvested by centrifugation, resuspended in 7 mLs BugBuster (Novagen), and incubated with gentle agitation for 20 minutes at room temperature. The cell extract was centrifuged at 10,000g for 15 minutes at 4ºC and the resultant supernatant was incubated with 2 mLs Ni-NTA resin (Novagen) in 2 mLs binding buffer (0.5 M NaCl, 5 mM imidazole, 40 mM Tris-HCL, pH 7.9) for one hour at 4ºC with gentle shaking. The resin was washed 2 times with 0.5 M NaCl, 120 mM imidazole, 20 mM Tris-HCL, pH 7.9, and eluted with 1 M imidazole, 0.5 M NaCl, 20 mM Tris-HCL, pH 7.9. Protein was dialyzed in 100mM potassium phosphate buffer, pH 7.0, 2 mM EDTA.
Protein purification, western blot analysis and immunoprecipitation
Biotinlylated victorin (BIC) was prepared as previously described (11). For analysis of protein extracts, leaves were infiltrated with 10 µg/mL victorin or 20 µg/mL BIC and incubated in the dark in a petri dish with moistened filter paper in the lid floating in a 25ºC water bath. At times indicated, 1 leaf per sample was ground in 200 µL TES (50 mM Tris pH 7.6, 5 mM EDTA, 2% SDS) with 20 mM DTT and 200 µL phenol. Samples were mixed and centrifuged at 13,000g for 5 minutes. The phenol phase was transferred to a clean microcentrifuge tube and 1 mL of 0.1 M ammonium acetate in methanol was added with 10 µL of 1 M DTT. Samples were precipitated overnight at 4ºC. Proteins were pelleted, washed with 100% methanol, dryed and resuspended in 30 µL SDS loading buffer with 20 mM DTT. 1.5 to 5 µL of each sample were run on a 16-18% SDS-PAGE gel. Soluble and plasma membrane protein partitioning was performed using an aqueous Dextran-polyethylene glycol two-phase system (22) Proteins were blotted onto a Protran nitrocellulose membrane with a 0.2-µm pore size (Whatman Schleicher and Schuell, Dassel, Germany). Membranes were blocked with 5% nonfat dry milk in PBST (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, 0.05% v/v Tween-20) or TBST (100 mM tris pH 7.5, 150 mM NaCl, 0.1% Tween) for 1hr to overnight. Anti-LOV1 antibody, anti-myc antibody (Invitrogen) and anti-H+ATPase antibody (Agrisera, Vannas, Sweden) were added at a 1:5000 dilution and Anti-Rubisco LSU antibody (Agrisera, Vannas, Sweden) at 1/10,000 and incubated for 1 h. Blots were washed three times for 10 min each with PBST or TBST. HRP-Rabbit Anti-Mouse IgG (Zymed Laboratories, South San Francisco, CA) and HRP-Goat Anti-Rabbit IgG (Promega, Madison, WI) were added at a 1:5000-10,000 dilution in PBST or TBST and incubated for 1 h. Blots were washed three times for 15 min each and developed with SuperSignal West Pico Chemiluminescent Substrate (Pierce, Rockford, IL)
For immunoprecipitation experiments, leaves from 3 1/2-week old Arabidopsis (5 leaves per sample) were infiltrated with 10 µg/mL victorin or 20 µg/mL BIC and incubated in the dark in a petri dish with moistened filter paper in the lid floating in a 25ºC water bath. At the indicated times, leaves were ground in 1 mL chilled lysis buffer (150 mM NaCl; 50 mM Tris pH 8.0; 0.1% NP-40) with protease inhibitors [1 mM PMSF;

1 mM EDTA; 1 Complete, Mini, EDTA-free Protease Inhibitor Cocktail Tablet per 10 mL buffer (Roche, Penzberg, Germany)]. All subsequent steps were performed at 4ºC. Samples were spun in a microfuge at 13,000g for 5 min and the supernatant removed to a clean microcentrifuge tube. 20 µL of a 50% resin slurry of Protein A Agarose (Sigma-Aldrich, St. Louis, MO) in lysis buffer was added and samples were placed on a rocker for 30 min. Samples were spun for 1 min at 13,000g and the supernatant removed to a clean tube. 2 µL of Anti-myc Antibody (Invitrogen) was added and samples were placed on a rocker for another 30 min. 50 µL of the Protein A slurry was added and samples were left on a rocker overnight. Samples were spun 30 sec at 13,000g and the supernatant discarded. The resin was washed four times with 500 µL of lysis buffer with protease inhibitors. The washed resin was boiled in 50 µL of SDS-PAGE loading buffer with 20 mM DTT. For Western blots, 40 µL of each sample was analyzed on a 16% SDS-PAGE gel. Gels were blotted and developed as described above except the 1º antibody was ImmunoPure Goat Anti-Biotin (Pierce) used at 1:25,000 and the 2º antibody was HRP-Rabbit Anti-Goat IgG (Zymed Laboratories, South San Francisco, CA) used at 1:12,500.
Pseudomonas syringae pv. maculicola inoculations
Arabidopsis seeds were incubated in 0.1% agarose for 5 days at 4 ºC and then applied to soil. Seedlings were grown in the growth chamber on a short day cycle (10h light/14h dark) at 22 ºC. Pseudomonas syringae pv. maculicola M6CΔE (Psm) (23) was grown overnight in Kings Medium B at 28 ºC and harvested by brief centrifugation. Psm cells were resuspended in 10 mM MgCl and 0.02%Silwett L-77 to 0.1 OD600 and sprayed until run-off on leaves of three to four-week-old Arabidopsis that were infiltrated, using a blunt-end 1 mL syringe, with water or 100 µg/mL victorin two hours prior to treatment with Psm. Two days after treatment with Psm, treated leaves were excised from plants, washed 3X in 40 mL water, and leaf disks were excised with a #3 cork borer. Three leaf disks/biological rep were combined in a 1.5 mL microcentrifuge tube and ground in 0.5 mL water with a plastic pestle. Serial dilutions of pulverized leaves were placed on Kings medium B, rifampicin (100 µg/mL) plates, incubated for two days at 28 ºC and colonies were counted.
For salicylic acid-induced resistance experiments, Arabidopsis seedlings were grown in hydroponic solution (24) in the growth chamber on a short day cycle (10h light/14h dark) at 22 ºC, and then transferred to water or 20 µg/ml victorin 24 hrs prior to spraying with water or 1mM salicylic acid (SA). Pseudomonas syringae pv. maculicola (Psm) was grown overnight in Kings Medium B at 28 ºC and harvested by brief centrifugation. Psm cells were resuspended in 10 mM MgCl and .02%Silwett L-77 to 0.1 OD600 and sprayed until run-off on twelve-day old seedlings. Two and one-half days after spraying Psm, 100 mg leaves/biological rep were combined in a 1.5 mL microcentrifuge tube, ground in 0.5 mL water and diluted and plated as described above. Quantitative RT-PCR analysis of PR1 expression
Leaves of three-week-old Arabidopsis seedlings grown in hydroponic solution (24) in the growth chamber on a short day cycle (10h light/14h dark) at 22 ºC, and then transferred to water or 20 µg/ml victorin 48 hrs prior to spraying with water or 1mM salicylic acid (SA), were harvested 24 hours after water/SA treatment. RNA from these

leaves was isolated using a plant RNAeasy kit (Promega) according to manufactures instructions. cDNA was synthesized from 1 µg RNA using Superscript III First-Strand Synthesis SuperMix (Invitrogen). Quantitative RT-PCR was performed using a C1000 thermal cycler (Bio-Rad), program 95ºC 3:00, (95ºC 0:10, 55ºC 0:10, 72ºC 0:30) X39, and Bio-Rad 2X SYBR Green Mix with primers F 5’-GATGTGATCATGCATACACACGTACA, R 5’-ATCCTGCATATGATGCTCCTTATTGA for PR1 (25) and F 5’-TCCAGCTAAGGGTGCC, R 5’-GGTGGGTACTCGGAGA for AthEf1a, which was used for transcript normalization. Data was analyzed with CFX Manager software (Bio-Rad) using the ΔΔC(t) method. Four biological x 2 technical replicates were used per data point and experiments were repeated at least two times.
Yeast-Two-Hybrid Assays
The clones for yeast-two-hybrid assays were PCR amplified and cloned in pENTR/D-Topo using the following primers: for the coiled-coil domain (LOV1 CC) LOVENTR-F (CACCATGGCTGAAGGAGTTGTGTTG) and LOVENTR-1R (TCAGTCTTCTTCTTTCCATACGTC) for the LRR domain (LOV1 LRR) LOVENTRLRR-F (CACCTCTGCCATCAATGCTCG) and LOVENTR-stop (TTATTCGTCACAGTTGATAAATTGAAC), and for RPS5CC, CACCATGGGAGGTTGTTTCTCTG and TCAATTCACTTTCTCCCATATGTC. Products cloned into pENTR/D-Topo were recombined, along with full-length LOV1 pENTR/D-Topo, into pJG4-5 (Clontech). TRX-h5 in pENTR/D-Topo (4) was recombined into pEG202 (26). Yeast strain EFY48 was transformed using an EasyComp Transformation Kit (Invitrogen) according to manufacturer’s recommendations. Yeast co-transformants harboring the appropriate constructs in pJG4-5 and pEG202 were selected on SD /-Trp/-His medium containing glucose (Yeast Protocols Handbook, Clontech). For evaluation of interaction between given TRX-h5 and LOV1 proteins, yeast co-transformants were plated on SD/-Leu/-Trp/-His medium containing galactose and raffinose (Yeast Protocols Handbook, Clontech). Mass Spectrometry
A C4 Ziptip (Millipore, MA) was utilized as a disposable desalting (or trapping) column to bind the proteins. To perform the Ziptip procedure, a Ziptip was prepared by rinsing with 10 µl acetonitrile (containing 0.1% of formic acid) three times, followed by three washes with 0.1% of formic acid. TRX sample solutions (10 µl each) was then bound to the Ziptip by pipetting the protein solution three times, followed by rinsing six times with 5 % acetonitrile containing 0.1% formic acid. The Ziptip was inserted between the LC and mass spectrometer via an in-house built interface as described previously (27). The sample was eluted into the mass spectrometry using 50 % aqueous acetonitrile containing 0.1% formic acid (27).
An Electrospray ionization LCT Premier time-of-flight mass spectrometer (Waters, Milford, MA, USA) was used. The instrument was operated in the positive-ion mode. Instrument settings were as follows: capillary voltage 2000V, cone voltage 60V, source temperature 90 °C. Nitrogen was used as a desolvation gas (400 L/h), with a desolvation temperature of 150°C. The scan range was m/z 400 to 2000. The instrument was calibrated using NaI (2 mg/mL) in isopropanol/water (1:1) over a mass range of 400

– 2000. Under the conditions used the mass accuracy is typically in the range of +/- 5-10 ppm. Data were collected using MassLynx v 4.0 software (Waters) and mass spectral deconvolution was performed using the Maximum Entropy (MaxEnt1) algorithm.
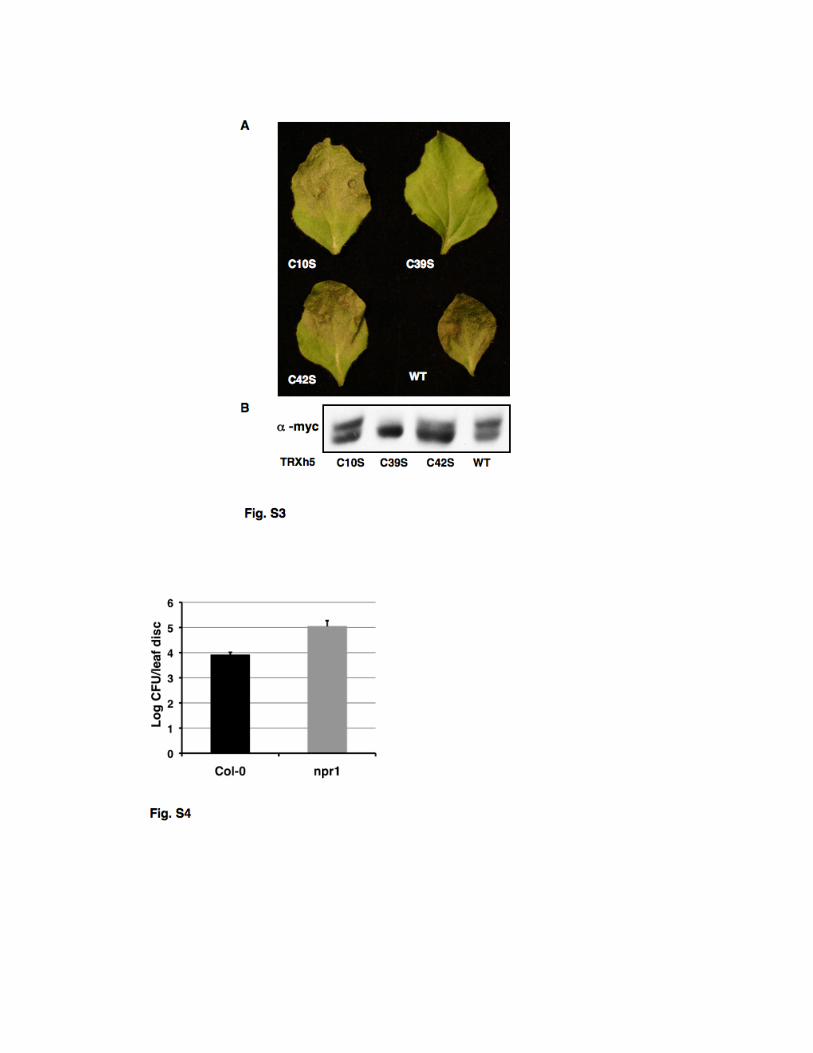
Supplemental figure legends Figure S1. Victorin binds to diverse thioredoxins. (A) SDS-polyacrylamide gel elctrophoresis of E. coli thioredoxin (TRX1) (Sigma) 20 minutes after incubation with 10 µM victorin or water. Victorin binding is seen as an ~ 1 kD increase in the apparent mass of TRX1 protein detected with Commassie blue. (B) Immunoblot of myc-tagged oat TRX-h1 and Arabidopsis TRX-h3 proteins from N. benthamiana leaves two hours after in vivo treatment with 10 µM victorin. Proteins were detected with α-myc antibody and victorin binding is seen as an ~ 1 kD increase in the apparent mass of TRX proteins on Western blots. Figure S2. Mass Spectrometry of TRX-h5, and the victorin-TRX-h5 adduct. (A) Mass of TRX was determined as 17,366 Da in presence of a reducing agent (DTT, MW 154). (B) After adding victorin (hydrated form, C31H45Cl3N6O13, MW 816; aldehydic form MW 798) to the TRX solution for binding, two major adducts were observed with deconvoluted masses of 18,127 and 18,282 Da, respectively. The adduct with a mass of 18,127 Da was tentatively assigned to the TRX-victorin adduct based on the mass shift of 761 Da which strongly suggested that the victorin bound to TRX and formed a stable adduct accompanied by loss of one “Cl”. The product with a mass of 18,282 Da was tentatively assigned as the DTT conjugate of the TRX-victorin adduct. Figure S3. The victorin-binding ability of transiently expressed TRX-h5 proteins correlates with the ability of TRX-h5s to support LOV-dependent cell death in Nicotiana benthamiana. (A) N. benthamiana transiently expressing LOV and wild-type (WT) or mutant TRX-h5s eight hours after treatment with 10 µM victorin. (B) Immunoblot of wildtype and mutant, myc-tagged TRX-h5 proteins from N. benthamiana leaves two hours after in vivo treatment with 10 µM victorin. Proteins were detected with α-myc antibody and victorin binding is seen as an ~ 1 kD increase in the apparent mass of TRX-h5 proteins in blotted, SDS-polyacrylamide gels.
Figure S4. Increased susceptibility of Arabidopsis Col-0 npr1to Pseudomonas syringae pv. maculicola M6CΔE (Psm). Colony forming units (CFUs) of Psm in leaves of four-week-old Arabidopsis seedlings two days after treatment with 0.1 OD600 Psm. Standard error bars represent N=6 biological replicates of 3 leaf disk each. Treatment means are significantly different by T-test (P < .006). Figure S5. Victorin inhibits salicylic acid-induced resistance. Colony forming units (CFUs) of Pseudomonas syringae pv. maculicola M6CΔE (Psm)/g of two-week-old Arabidopsis seedlings 2.5 days after treatment with 0.1 OD600 Psm. Arabidopsis were grown in hydroponic solution and transferred to water or 20 µg/ml victorin 24 hrs prior to spraying with water or 1mM salicylic acid (SA) to induce resistance. Plants were sprayed with Psm 24 hours after SA (or water) treatment. Standard error bars represent N=8 biological replicates of 100 mg leaf tissue each. Asterisk indicates a significant difference

from the other treatment by T-test (P < .006). Experiments were limited to a single time point because large amounts of victorin were required.
Figure S6. Victorin causes cell death in Common Bean. Leaf of 4-week old Common Bean (Pinto) 36 hours after infiltration with water (left) or 10 µM victorin (right).
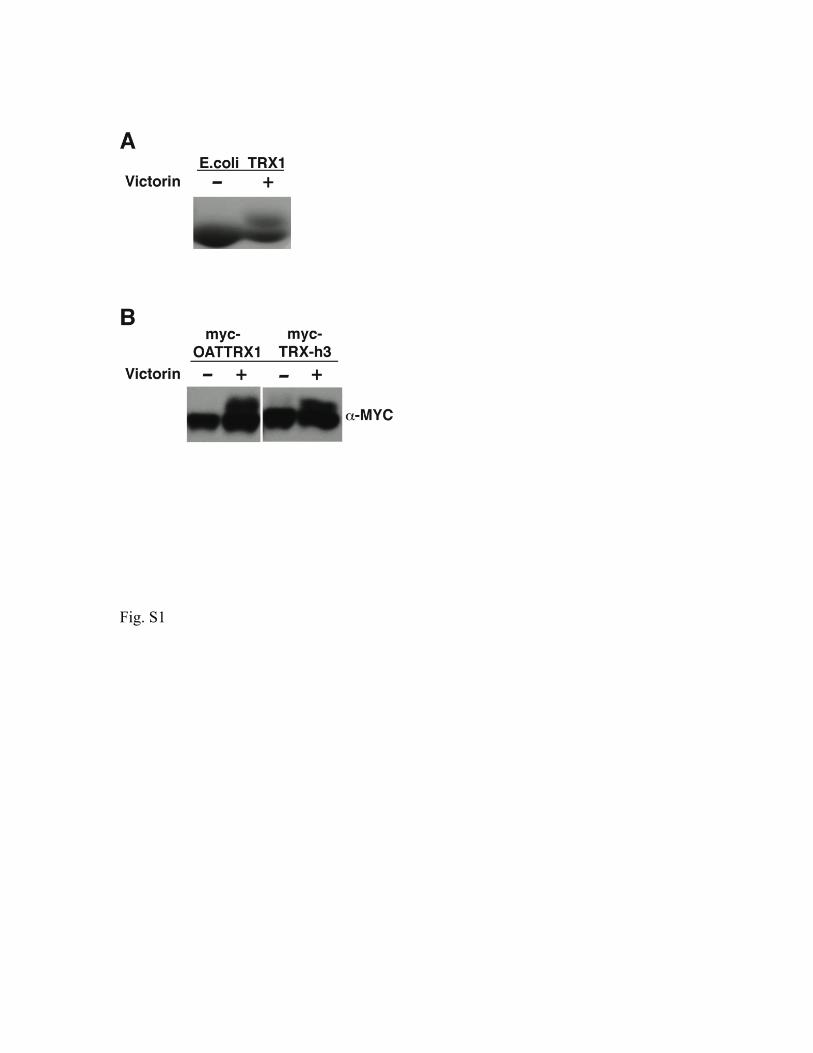
Fig. S1
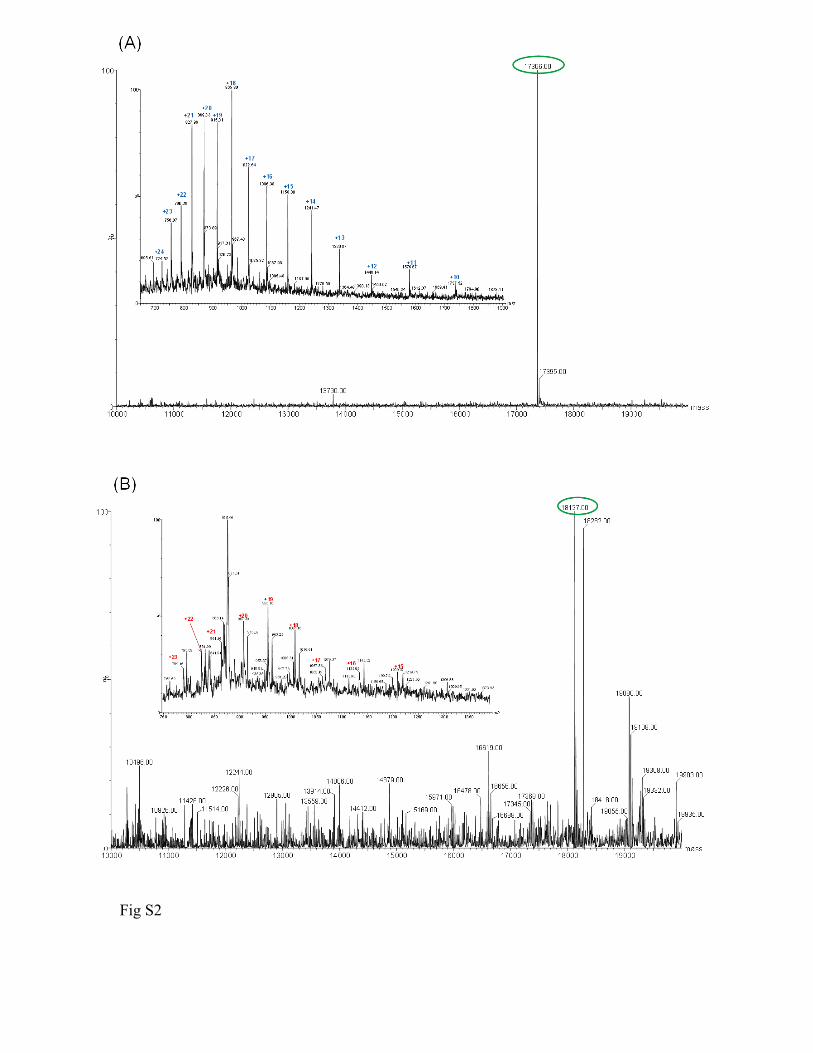
Fig S2


References
1. J. M. Lorang, T. A. Sweat, T. J. Wolpert, Plant disease susceptibility conferred by a
“resistance” gene. Proc. Natl. Acad. Sci. U.S.A. 104, 14861 (2007).
doi:10.1073/pnas.0702572104 Medline
2. J. D. G. Jones, J. L. Dangl, The plant immune system. Nature 444, 323 (2006).
doi:10.1038/nature05286 Medline
3. B. J. DeYoung, R. W. Innes, Plant NBS-LRR proteins in pathogen sensing and host defense.
Nat. Immunol. 7, 1243 (2006). doi:10.1038/ni1410 Medline
4. T. A. Sweat, J. M. Lorang, E. G. Bakker, T. J. Wolpert, Characterization of natural and
induced variation in the LOV1 gene, a CC-NB-LRR gene conferring victorin sensitivity
and disease susceptibility in Arabidopsis. Mol. Plant Microbe Interact. 21, 7 (2008).
doi:10.1094/MPMI-21-1-0007
5. T. J. Wolpert, L. D. Dunkle, L. M. Ciuffetti, Host-selective toxins and avirulence
determinants: What’s in a name? Annu. Rev. Phytopathol. 40, 251 (2002).
doi:10.1146/annurev.phyto.40.011402.114210 Medline
6. Y. Tada et al., Plant immunity requires conformational changes of NPR1 via S-nitrosylation
and thioredoxins. Science 321, 952 (2008). doi:10.1126/science.1156970 Medline
7. H. Cao, S. A. Bowling, A. S. Gordon, X. Dong, Characterization of an Arabidopsis mutant
that is nonresponsive to inducers of systemic acquired resistance. Plant Cell 6, 1583
(1994). Medline
8. T. A. Sweat, T. J. Wolpert, Thioredoxin h5 is required for victorin sensitivity mediated by a
CC-NBS-LRR gene in Arabidopsis. Plant Cell 19, 673 (2007).
doi:10.1105/tpc.106.047563 Medline
9. C. Laloi, D. Mestres-Ortega, Y. Marco, Y. Meyer, J. P. Reichheld, The Arabidopsis cystolic
thioredoxin h5 gene induction by oxidative stress and its W-box-mediated response to
pathogen elicitor. Plant Physiol. 134, 1006 (2004). doi:10.1104/pp.103.035782 Medline
10. See supplementary materials on Science Online.

11. T. J. Wolpert, V. Macko, W. Acklin, D. Arigoni, Molecular features affecting the biological
activity of host-selective toxins from Cochliobolus victoriae. Plant Physiol. 88, 37
(1988). doi:10.1104/pp.88.1.37 Medline
12. C. C. Mundt, Importance of autoinfection to the epidemiology of polycyclic foliar disease.
Phytopathology 99, 1116 (2009). doi:10.1094/PHYTO-99-10-1116 Medline
13. A. Marmagne et al., A high content in lipid-modified peripheral proteins and integral
receptor kinases features in the Arabidopsis plasma membrane proteome. Mol. Cell.
Proteomics 6, 1980 (2007). doi:10.1074/mcp.M700099-MCP200 Medline
14. T. Wroblewski et al., Comparative large-scale analysis of interactions between several crop
species and the effector repertoires from multiple pathovars of Pseudomonas and
Ralstonia. Plant Physiol. 150, 1733 (2009). doi:10.1104/pp.109.140251 Medline
15. J. Lorang, A. Cuesta-Marcos, P. M. Hayes, T. J. Wolpert, Identification and mapping of
adult-onset sensitivity to victorin in barley. Mol. Breed. 26, 545 (2010).
doi:10.1007/s11032-010-9458-5
16. E. D. Nagy, J. L. Bennetzen, Pathogen corruption and site-directed recombination at a plant
disease resistance gene cluster. Genome Res. 18, 1918 (2008). doi:10.1101/gr.078766.108
Medline
17. J. D. Faris et al., A unique wheat disease resistance-like gene governs effector-triggered
susceptibility to necrotrophic pathogens. Proc. Natl. Acad. Sci. U.S.A. 107, 13544 (2010).
doi:10.1073/pnas.1004090107 Medline
18. K. W. Earley et al., Gateway-compatible vectors for plant functional genomics and
proteomics. Plant J. 45, 616 (2006). doi:10.1111/j.1365-313X.2005.02617.x Medline
19. K. Martin et al., Transient expression in Nicotiana benthamiana fluorescent marker lines
provides enhanced definition of protein localization, movement and interactions in
planta. Plant J. 59, 150 (2009). doi:10.1111/j.1365-313X.2009.03850.x Medline
20. S. J. Clough, A. F. Bent, Floral dip: A simplified method for Agrobacterium-mediated
transformation of Arabidopsis thaliana. Plant J. 16, 735 (1998). doi:10.1046/j.1365-
313x.1998.00343.x Medline

21. A. Holgrem, Thioredoxin catalyzes the reduction of insulin disulfides by dithiothreitol and
dihydrolipoamide. J. Biochem. 254, 9627 (1979).
22. C. Larsson, S. Widell, P. Kjellbom, Preparation of high-purity plasma membranes. Methods
Enzymol. 148, 558 (1987). doi:10.1016/0076-6879(87)48054-3
23. L. Rohmer, S. Kjemtrup, P. Marchesini, J. L. Dangl, Nucleotide sequence, functional
characterization and evolution of pFKN, a virulence plasmid in Pseudomonas syringae
pathovar maculicola. Mol. Microbiol. 47, 1545 (2003). doi:10.1046/j.1365-
2958.2003.03402.x Medline
24. D. M. Gibeaut, J. Hulett, G. R. Cramer, J. R. Seemann, Maximal biomass of Arabidopsis
thaliana using a simple, low-maintenance hydroponic method and favorable
environmental conditions. Plant Physiol. 115, 317 (1997). doi:10.1104/pp.115.2.317
Medline
25. M. Tian et al., Arabidopsis actin-depolymerizing factor AtADF4 mediates defense signal
transduction triggered by the Pseudomonas syringae effector AvrPphB. Plant Physiol.
150, 815 (2009). doi:10.1104/pp.109.137604 Medline
26. pEG202 was a gift from Hironori Kaminaka and Jeff J. Dangl.
27. T. W. Rhoads et al., Measuring copper and zinc superoxide dismutase from spinal cord tissue
using electrospray mass spectrometry. Anal. Biochem. 415, 52 (2011).
doi:10.1016/j.ab.2011.03.029 Medline