studies on the control ofsodium excretiondm5migu4zj3pb.cloudfront.net/manuscripts/106000/... ·...
TRANSCRIPT

Studies on the Control of Sodium Excretion
in Experimental Uremia
RAYMONDG. SCHULTZE, HowARDS. SHAPIRo, and NEAL S. BRICKER
From the Renal Division, Department of Internal Medicine, WashingtonUniversity School of Medicine, St. Louis, Missouri 63110
A B S T R A C T A study of the mechanisms governingthe high rate of sodium excretion per nephron charac-teristic of patients with chronic renal disease has beenmade in dogs. A "remnant kidney" was produced by85% infarction of the left kidney while the right kidneywas left intact. A bladder-splitting procedure allowedsimultaneous measurement of glomerular filtration rateand the rate of sodium excretion by each kidney. Theanimals were fed a constant known amount of sodiumchloride and 0.1 mg of 9 a-fluorohydrocortisone twicedaily throughout the study.
In a group of dogs fed 3 or 5 g of salt per day, sodiumexcretion by the remnant kidney averaged 6.5 /AEq/minwhile the intact kidney was present and 53.7 /AEq/minwhen the animals became uremic after the intact kidneywas removed. The increased sodium excretion per neph-ron by the remnant organ often occurred within 18 hrafter contralateral nephrectomy and persisted despiteexperimentally induced acute reductions in the glomerularfiltration rate to below prenephrectomy levels. A secondgroup of animals studied in the same manner but receiv-ing 1 g of salt per day or less failed to develop a natri-uresis after contralateral nephrectomy despite high gradeuremia. Thus an increased impermeable solute load pernephron was not a regulatory factor in the productionof the natriuresis. The increased rate of sodium excre-tion per nephron in uremia resembles that after salineloading in that it may occur without an increase in glo-merular filtration rate or a reduction in mineralocorticoidstimulation. It follows that an additional factor or fac-tors must be involved in the genesis of the natriuresis.
A portion of this work was presented in preliminary format the 79th meeting of the Association of American Physi-cians, Atlantic City, N. J., 3 May 1966. Trans. Ass. Amer.Physicians Philadelphia. 79: 322.
Dr. Schultze is a U. S. Public Health Service SpecialResearch Fellow. Dr. Shapiro is a U. S. Public Health Serv-ice Research Fellow. Dr. Bricker is a recipient of a U. S.Public Health Service Research Career Award.
Received for publication 10 October 1968 and in revisedform 3 January 1969.
In contrast to the natriuresis that is seen in normalanimals subjected to saline loading, these uremic ani-mals were found not to have a detectable increase inextracellular fluid volume or blood volume in the pres-ence of high fractional sodium excretion rates.
Sodium excretion in response to a small salt load bythe remnant organ in uremia was 30% greater than theresponse of both kidneys in the preuremic state despitea markedly reduced total GFR. These data are consistentwith the view that the volume control mechanism be-comes more responsive in uremia.
INTRODUCTIONPatients with chronic renal disease generally are capableof maintaining sodium balance on an average salt intakedespite a marked reduction in their functioning nephronpopulation. It follows that if the range of salt intakeremains uniform, sodium excretion per nephron must in-crease by approximately twofold for every 50% re-duction in glomerular filtration rate (GFR). At very lowlevels of GFR, therefore, the fraction of filtered sodiumexcreted must be quite large. For example on a 7 g saltintake with a GFRof 4 ml/min, 16% of the filtered so-dium must be excreted to maintain balance. Similar con-siderations indicate that the range over which both ab-solute sodium excretion per nephron and the fractionof filtered sodium excreted must change in response torelatively small changes in salt intake must becomeprogressively larger as GFR becomes progressivelylower.
The mechanisms responsible for the renal regulation ofsodium excretion in uremia are only partially understood.The present studies are addressed to a further definitionof these mechanisms.
METHODSStudies were performed on female mongrel dogs weighingbetween 10-20 kg. A "remnant" kidney was produced in onekidney in each dog by ligating sufficient second and third
The Journal of Clinical Investigation Volume 48 1969 869

order branches of the left renal artery to reduce the renalmass by approximately 85%. The extent of the infarctioncould be established at the time of surgery by observing thearea of ischemia. The residual nephrons, when examined atnecropsy, were free of any pathologic stigmata of chronicdisease except for limited round cell infiltration at the junc-tion of the infarcted and noninfarcted zones. The dogs wereallowed to recover from the surgical procedure for at leasta week, after which the urinary bladder was divided intotwo permanent hemi-bladders in the manner described previ-ously (1). An additional week was allowed for recoveryfrom this procedure.
86 studies were performed on 27 dogs. Each of the ani-mals was maintained on a constant caloric intake containinga known amount of salt throughout the duration of thestudies (from 7 days to 6 wk). The basic diet contained lessthan 1 mEq of sodium per day. In all but one group of ani-mals a weighed amount of sodium chloride was added to thediet. 0.2 mg of 9-a-fluorohydrocortisone also was givendaily. The diet, the added sodium chloride (1, 3, or 5 g/day),and the mineralocorticoid hormone were administered bystomach tube twice daily in two equal parts. The animalswere maintained on this regimen for a minimum of 5 daysbefore experiments were begun. Several different types ofexperiments were performed.
Serial studies. (a) 3-5 g salt intake. Serial experimentsdesigned to measure the changes in the patterns of sodiumexcretion by the remnant kidneys were performed in eightdogs maintained on either a 3 or a 5 g salt diet. From oneto three control experiments were performed with the nor-mal kidney in situ. Simultaneous measurements were madeon both the remnant and the contralateral kidneys. There-after the normal kidney was removed surgically, and re-peated experiments were performed on the remnant kidneyover an 8 day period. In six of the eight dogs, the initialpostnephrectomy study on the remnant organ was carriedout within 18 hr of contralateral nephrectomy. (b) 0-1 gsalt intake. In six animals serial studies were performedaccording to the above schedule; however, these dogs re-ceived either no supplementary salt or 1 g of supplementarysalt.
Renal artery constriction. In six animals, the effects ofrenal arterial constriction on sodium excretion were studiedapproximately a week after the control kidney had been re-moved and uremia had developed. A constricting band wasplaced loosely about the renal artery of the remnant kidney24-48 hr before the study. The experiments then were per-formed with the dogs awake and in the standing position.Several control clearance periods were obtained, after whichthe band was constricted so as to reduce GFR and filteredload of sodium below the level observed in the prenephrec-tomy control studies in the same kidney.
Timed response to a 2.5 g salt load. A series of studieswas performed to determine the quantitative response of theremnant kidneys. A total of 31 studies was performed on 13dogs. Each animal was maintained chronically on a 5 g saltdiet in the manner described above. On the morning of study,one-half of the daily salt load (2.5 g) was administered inone of four different ways: (a) intravenously as isotonicsaline at 5 ml/min; (b) intravenously as isotonic saline at50 ml/min; (c) by stomach tube as isotonic saline (thelow-salt basic diet was omitted in these experiments) ; (d)by stomach tube as sodium chloride dissolved in 250 ml ofwater together with half of the basic daily diet. In all dogsthe effects of at least two different routes of administrationwere examined both before and after removing the normal
kidneys. In the typical study, the infusion for the measure-ment of creatinine clearance was initiated approximately 2 hrbefore administering the salt load. Three to four preliminaryclearance periods were obtained to determine the base linepatterns of sodium excretion. Thereafter the salt load wasdelivered and the excretion of sodium was determined dur-ing five successive 1-hr periods. GFR was measured duringeach of the urine collection periods. The patterns of sodiumexcretion during the 5 hr after administration of the saltload in the postnephrectomy studies were compared withthose in the prenephrectomy control studies. Comparisonsalso were made between the response to the various routesof administration of sodium chloride. In all of these stud-ies, the animals were awake and standing quietly in a sup-porting sling. The sodium load was administered at the sametime of day in all studies in individual dogs.
Studies of inulin space. In nine animals maintained on a5 g salt diet, extracellular fluid volume was estimated beforeand after removing the normal kidney. Inulin space was mea-sured using the method of Deane (2). GFRand sodium ex-cretion were determined simultaneously with the measure-ments of inulin space.
Red cell mass and blood volume. In five of the animalsin which inulin space was determined, simultaneous mea-surements were made of blood volume with 51Cr-labeled redblood cells from the same animal (3).
In all studies, GFR (exogenous creatinine clearance), fil-tered load of sodium, and sodium excretion rates were de-termined. 7-10 ml of blood were collected at the midpointof each period through a catheter placed in the externaljugular vein. Infusions were delivered via a peripheral veingenerally in a hind limb. Creatinine dissolved in 5% dextrosein water was delivered at a rate of 3 ml/min in the pre-nephrectomy studies and 1 ml/min in postnephrectomy stud-ies. Creatinine was determined according to the method ofBonsnes and Taussky (4). Sodium was measured on aflame photometer. When serial studies were performed, thered blood cells obtained during each clearance period werereconstituted to the original hematocrit with isotonic salineand returned at the end of each experiment.
RESULTS
In Table I are shown the results of prenephrectomy con-trol values for GFR and sodium excretion on normaland remnant kidneys in 18 dogs used in the serial stud-ies. Six dogs were maintained on a 5 g salt diet, six ona 3 g salt diet, and six on 1 g of salt per day or less.GFR for the normal kidneys ranged from 30.5 to 81ml/min. Values for the remnant kidneys ranged from3.6 to 11.1 ml/min. The fraction of filtered sodium ex-creted was slightly greater for the remnant than for thecontrol kidneys in all but one of the dogs. Nevertheless,all of the remnant kidneys reabsorbed at least 98.5% ofthe filtered sodium, and in 14 of the 18 dogs, sodium re-absorption exceeded 99% of the filtered load.
Serial studies. The results of five successive studieson a representative dog maintained on a 5 g salt diet areshown in Table II. Values are recorded for the rem-nant kidney only. During the prenephrectomy study, thesodium excretion rate averaged 6.0 uEq/min. The nor-mal kidney excreted an average of 56.1 IAEq/min during
870 R. G. Schultze, H. S. Shapiro, and N. S. Bricker

TABLE ISummary of Control Measurements of Dogs in Serial Studies
GFR UN&V UN&V/FLN&No. ofdogs Diet I R I R I R
ml/min uEq/min %12 3 or 5 g NaCl 58.0(sE) 4-3.6 6.4(SE) ±0.5 24.5(sE) ±4.2 7.2(SE) :+:0.8 0.34(SE) :1:0.07 0.85(sE) 40.10
6 0.25 or 1 g NaCl 55.3 ±6.1 8.4 40.9 6.5 41.6 4.1 ±0.9 0.10 ±0.02 0.39 40.58
Values represent means i standard error.GFR, glomerular filtration rate; I, intact kidney; R, remnant kidney; UNaV, sodium excretion rate; UN.V/FLN., fractionalsodium excretion.
this study. 18 hr after removing the normal kidney, theGFR of the remnant organ had increased from an av-erage prenephrectomy value of 6.1 to 8.0 ml/min. At thesame time the sodium excretion rate had risen to 50,uEqcmin. The fraction of filtered sodium excreted in-creased from an average prenephrectomy value of 0.7 to4.7%. In the three subsequent studies performed on days2, 3, and 6, GFR remained between 7 and 7.8 ml/min,sodium excretion rate varied from 61 to 87 ,uEq/min,and fractional sodium excretion increased further to 8%.
The results of the serial studies on all eight dogsmaintained on the 3 or 5 g NaCl diets are summarizedin Table III. In four of the six dogs which were studiedwithin 18 hr of removing the normal kidney, sodiumexcretion rates by the remnant kidney increased by atleast threefold. In studies performed from the 2ndthrough the 8th postnephrectomy day substantial in-creases in sodium excretion by the remnant kidneys oc-curred in all eight dogs. In most animals, GFRand thefiltered load of sodium increased. However in two ani-mals (dogs 6 and 7) GFRand filtered sodium decreasedmodestly. The natriuresis occurred nevertheless. All ani-
TABLE I IRepresentative Experiment in Which the Sodium Excretion
of the Remant Kidney Was Followed for 6 Daysafter Contralateral Nephrectomy*
UNaV/Day Weight BUN GFR FLNa UNaV FLNa
kg mg/100 ml ml/min pEq/min uEq/min %
Control 12.4 10.0 6.1 819 6 0.7Intact kidney removed
18 hr 12.5 31.0 8.0 1069 50 4.72 12.7 52.5 7.0 984 51 6.23 12.5 54.0 7.8 1101 81 7.96 11.8 63.0 7.1 995 76 7.7
* Experiment performed on dog No. 8.BUN, blood urea nitrogen. All other abbreviations are ex-plained in Table I.
mals exhibited an appreciable increase in the fractionof filtered sodium excreted. The average blood urea ni-trogen (BUN) on the final day of study in this groupwas 77 mg/100 ml.
In order to examine the role of the increase in GFRon the increased rates of sodium excretion by the rem-nant kidneys, the effects of reduction of GFR by con-striction of the renal artery were measured. The datafrom the six dogs subjected to this procedure are re-corded in Table IV. Values are shown for the pre-nephrectomy control study, as well as for the post-nephrectomy experiment before and after constrictingthe renal artery. In three of the six dogs no evidence ofan increase in GFR was observed in the postnephrec-tomy state. Conceivably this may have related to somedegree of arterial constriction created by the band evenbefore experimental constriction. Yet each of these dogsexhibited a typical increase in sodium excretion in thepostnephrectomy study. In one animal (dog No. 9),GFRand urine flow were sufficiently reduced that fur-ther constriction was not carried out. In all six animals,sodium excretion rate in postnephrectomy studies hadincreased markedly over prenephrectomy values; andwith constriction, excretion rates remained well abovethe corresponding values in the prenephrectomy studiesdespite the fact that the filtered load of sodium was from15 to 48% lower.
One factor that could contribute to the acquired natri-uresis by the remnant kidneys is an increase in the filtra-tion of impermeable or poorly permeant solutes whichcan accumulate in the extracellular fluid in uremia. Thispossibility was examined in two ways. The first involvesthe study of dogs maintained on a low sodium diet (1.0g of NaCl or less). Data from this group of experimentsare summarized in Table V. Sodium excretion rate byremnant kidneys averaged 4.1 ,uEq/min and by the rem-nant plus control kidneys averaged 10.7 ,uEq/min in theprenephrectomy studies. In the postnephrectomy studies,the excretion rates by the remnant kidneys averaged only13.1 ,uEq/min despite the existence of chemical ab-normalities of uremia. Two postnephrectomy studies
Studies on the Control of Sodium Excretion 871
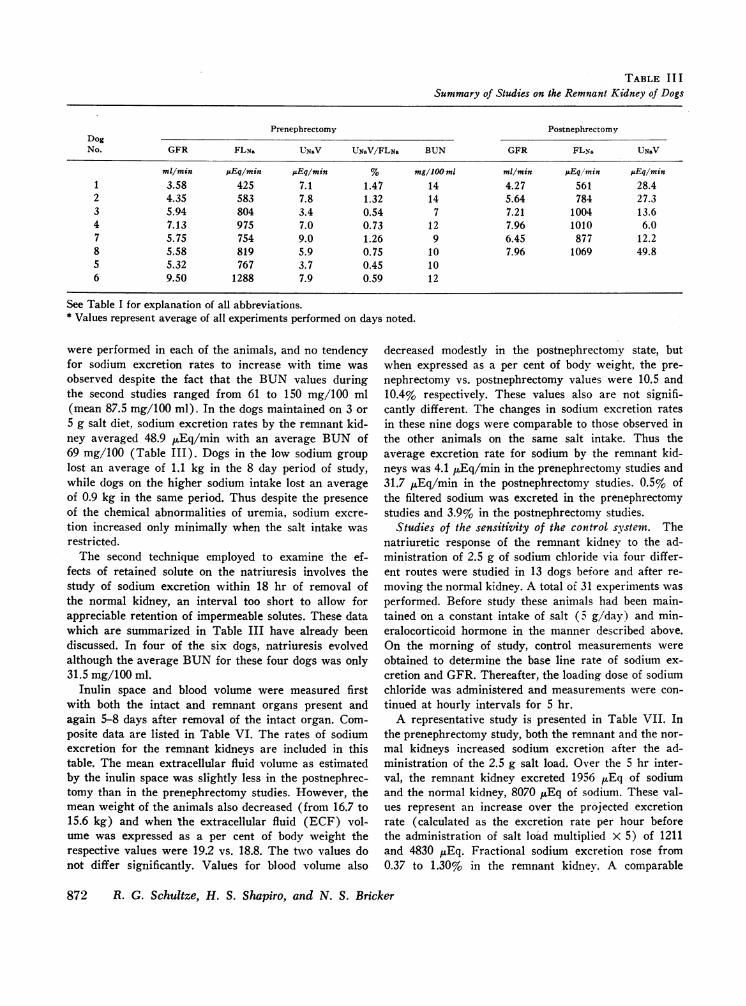
TABLE II ISummary of Studies on the Remnant Kidney of Dogs
Prenephrectomy PostnephrectomyDogNo. GFR FLN, UNaV UNaV/FLNa BUN GFR FLNs UNaV
ml/min pEq/min pEq/min % mg/100 ml ml/min pEqlmin pEq/min
1 3.58 425 7.1 1.47 14 4.27 561 28.42 4.35 583 7.8 1.32 14 5.64 784 27.33 5.94 804 3.4 0.54 7 7.21 1004 13.64 7.13 975 7.0 0.73 12 7.96 1010 6.07 5.75 754 9.0 1.26 9 6.45 877 12.28 5.58 819 5.9 0.75 10 7.96 1069 49.85 5.32 767 3.7 0.45 106 9.50 1288 7.9 0.59 12
See Table I for explanation of all abbreviations.* Values represent average of all experiments performed on days noted.
were performed in each of the animals, and no tendencyfor sodium excretion rates to increase with time wasobserved despite the fact that the BUN values duringthe second studies ranged from 61 to 150 mg/100 ml(mean 87.5 mg/100 ml). In the dogs maintained on 3 or5 g salt diet, sodium excretion rates by the remnant kid-ney averaged 48.9 uEq/min with an average BUN of69 mg/100 (Table III). Dogs in the low sodium grouplost an average of 1.1 kg in the 8 day period of study,while dogs on the higher sodium intake lost an averageof 0.9 kg in the same period. Thus despite the presenceof the chemical abnormalities of uremia, sodium excre-tion increased only minimally when the salt intake wasrestricted.
The second technique employed to examine the ef-fects of retained solute on the natriuresis involves thestudy of sodium excretion within 18 hr of removal ofthe normal kidney, an interval too short to allow forappreciable retention of impermeable solutes. These datawhich are summarized in Table III have already beendiscussed. In four of the six dogs, natriuresis evolvedalthough the average BUNfor these four dogs was only31.5 mg/100 ml.
Inulin space and blood volume were measured firstwith both the intact and remnant organs present andagain 5-8 days after removal of the intact organ. Com-posite data are listed in Table VI. The rates of sodiumexcretion for the remnant kidneys are included in thistable. The mean extracellular fluid volume as estimatedby the inulin space was slightly less in the postnephrec-tomy than in the prenephrectomy studies. However, themean weight of the animals also decreased (from 16.7 to15.6 kg) and when the extracellular fluid (ECF) vol-ume was expressed as a per cent of body weight therespective values were 19.2 vs. 18.8. The two values donot differ significantly. Values for blood volume also
decreased modestly in the postnephrectomy state, butwhen expressed as a per cent of body weight, the pre-nephrectomy vs. postnephrectomy values were 10.5 and10.4% respectively. These values also are not signifi-cantly different. The changes in sodium excretion ratesin these nine dogs were comparable to those observed inthe other animals on the same salt intake. Thus theaverage excretion rate for sodium by the remnant kid-neys was 4.1 ,uEq/min in the prenephrectomy studies and31.7 /AEq/min in the postnephrectomy studies. 0.5% ofthe filtered sodium was excreted in the prenephrectomystudies and 3.9% in the postnephrectomy studies.
Studies of the sensitivity of the control system. Thenatriuretic response of the remnant kidney to the ad-ministration of 2.5 g of sodium chloride via four differ-ent routes were studied in 13 dogs before and after re-moving the normal kidney. A total of 31 experiments wasperformed. Before study these animals had been main-tained on a constant intake of salt (5 g/day) and min-eralocorticoid hormone in the manner described above.On the morning of study, control measurements wereobtained to determine the base line rate of sodium ex-cretion and GFR. Thereafter, the loading dose of sodiumchloride was administered and measurements were con-tinued at hourly intervals for 5 hr.
A representative study is presented in Table VII. Inthe prenephrectomy study, both the remnant and the nor-mal kidneys increased sodium excretion after the ad-ministration of the 2.5 g salt load. Over the 5 hr inter-val, the remnant kidney excreted 1956 uEq of sodiumand the normal kidney, 8070 uEq of sodium. These val-ues represent an increase over the projected excretionrate (calculated as the excretion rate per hour beforethe administration of salt load multiplied X 5) of 1211and 4830 uEq. Fractional sodium excretion rose from0.37 to 1.30% in the remnant kidney. A comparable
872 R. G. Schultze, H. S. Shapiro, and N. S. Bricker

Fed a Normal Salt Die} and Followed Longitudinally
(18 hr) Postnephrectomy* (2nd-8th day)
DaysUN&V/FLN& BUN GFR FLN& UNNV UNaV/FLN, BUN observed
% mg/1JO ml ml/min pEq/min ;LEq/min % mg/100 ml
5.05 40 4.39 597 79.1 12.60 81 2-43.50 36 5.77 813 53.2 6.55 66 21.33 19 7.05 988 56.2 5.60 81 2-5,80.41 36 7.55 1067 38.6 4.49 86 2-81.37 23 4.34 608 56.5 9.32 73 2,3,6,84.43 31 6.78 1040 47.2 6.31 73 2,3,6
7.76 1053 65.6 5.95 68 3-5,88.66 1171 32.0 3.20 47 2,3,5
change occurred in the normal organ. Maximal excre- Table VIII presents the data from the 31 observationstion occurred in the 3rd hr bilaterally. in the 13 dogs. The combined 5 hr sodium excretion rates
The same salt load was administered via the same for remnant plus control kidneys in the prenephrectomyroute after the control kidney had been removed. So- studies was 8914 uEq. The 5-hr value for the remnantdium excretion rate in the 5 hr interval after adminis- kidney alone in the postnephrectomy studies was 11,705tering the load was greater than the combined excretion uEq. For the group the difference between the 5-hr ex-rate of remnant plus the normal kidney in the pre- cretion rate and the projected 5-hr excretion rate wasnephrectomy studies. The increase over the projected 3900 uEq for the two kidneys in the prenephrectomyexcretion rate was slightly less than the combined two studies and 2100 ,uEq for the remnant alone in the post-kidney values in the prenephrectomy studies. Fractional nephrectomy studies. This occurred despite the factsodium excretion increased from an initial level of 2.43 that the GFRin the remnant kidney in the postnephrec-to a maximum of 6.41%. Maximum sodium excretion tomy studies averaged only 15% of the total (i.e. com-occurred in the 3rd hr. Thus in response to the same bined two kidney) GFR in prenephrectomy studies. Thesalt load, the remnant kidney excreted more sodium route of sodium administration did not affect the re-both in absolute terms and in terms of the increase over sponse, and there was no difference for either individualprojected base line excretion rates in the uremic state animals or group data between slow intravenous infu-than when the contralateral control kidney contributed sion (11 experiments), rapid intravenous infusion (sixto function. experiments), intragastric saline instillation (seven ex-
TABLE IVSummary of Data from the Constriction Experiments: Glomerular Filtration Rate, Filtered Sodium,
and Sodium Excretion by the Remnant Kidney
PostnephrectomyPrenephrectomy Postnephrectomy with constriction
DogNo. GFR FLNa, UN&V GFR FLNa UNaV GFR FLNa UN.V
ml/min AEq/min pEq/min ml/min uEq/min pEq/min ml/min jsEq/min sEq/min
4 7.13 975 6.99 6.96 1029 86.7 5.72 832 37.55 5.32 767 3.69 5.38 716 114.0 4.01 524 56.5
10 8.68 1180 11.2 11.0 1528 55.9 6.76 903 17.611 5.55 760 3.74 7.38 1022 77.0 5.28 708 29.4
4.80 628 18.612 7.05 967 7.43 8.64 1150 88.3 4.41 575 39.4
3.90 498 29.49 8.53 1117 11.8 5.59 729 25.3
See Table I for explanation of all abbreviations.
Studies on the Control of Sodium Excretion 873
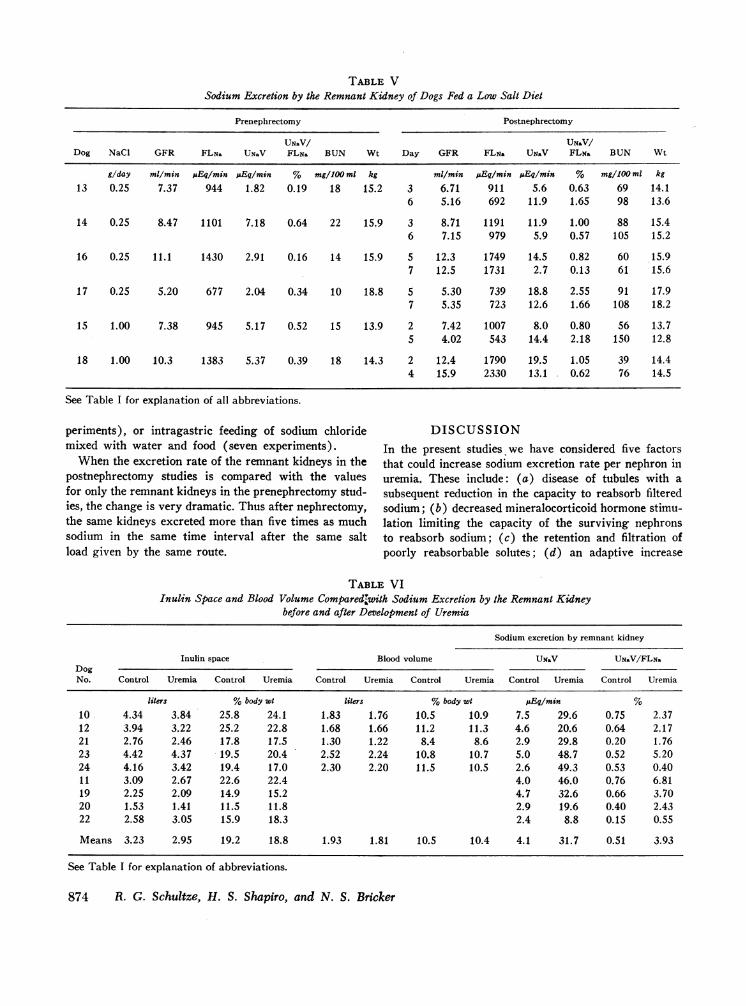
TABLE VSodium Excretion by the Remnant Kidney of Dogs Fed a Low Salt Diet
Prenephrectomy Postnephrectomy
UNaV/ UNaV/Dog NaCl GFR FLNa UN&V FLNa BUN Wt Day GFR FLN, Ug.V FLN& BUN Wt
g/day ml/min uEq/min pEqlmin % mg/100 ml kg ml/min j&Eq/min .Eq/min % mg/100 ml kg13 0.25 7.37 944 1.82 0.19 18 15.2 3 6.71 911 5.6 0.63 69 14.1
6 5.16 692 11.9 1.65 98 13.6
14 0.25 8.47 1101 7.18 0.64 22 15.9 3 8.71 1191 11.9 1.00 88 15.46 7.15 979 5.9 0.57 105 15.2
16 0.25 11.1 1430 2.91 0.16 14 15.9 5 12.3 1749 14.5 0.82 60 15.97 12.5 1731 2.7 0.13 61 15.6
17 0.25 5.20 677 2.04 0.34 10 18.8 5 5.30 739 18.8 2.55 91 17.97 5.35 723 12.6 1.66 108 18.2
15 1.00 7.38 945 5.17 0.52 15 13.9 2 7.42 1007 8.0 0.80 56 13.75 4.02 543 14.4 2.18 150 12.8
18 1.00 10.3 1383 5.37 0.39 18 14.3 2 12.4 1790 19.5 1.05 39 14.44 15.9 2330 13.1 0.62 76 14.5
See Table I for explanation of all abbreviations.
periments), or intragastric feeding of sodium chloride DISCUSSIONmixed with water and food (seven experiments). In the present studies we have considered five factors
When the excretion rate of the remnant kidneys in the that could increase sodium excretion rate per nephron inpostnephrectomy studies is compared with the values uremia. These include: (a) disease of tubules with afor only the remnant kidneys in the prenephrectomy stud- subsequent reduction in the capacity to reabsorb filteredies, the change is very dramatic. Thus after nephrectomy, sodium; (b) decreased mineralocorticoid hormone stimu-the same kidneys excreted more than five times as much lation limiting the capacity of the surviving nephronssodium in the same time interval after the same salt to reabsorb sodium; (c) the retention and filtration ofload given by the same route. poorly reabsorbable solutes; (d) an adaptive increase
TABLE VIInulin Space and Blood Volume Compareduwith Sodium Excretion by the Remnant Kidney
before and after Development of Uremia
Sodium excretion by remnant kidney
Inulin space Blood volume UN&V UNaV/FLN&DogNo. Control Uremia Control Uremia Control Uremia Control Uremia Control Uremia Control Uremia
liters %body wt liters %body wt AEq/min %10 4.34 3.84 25.8 24.1 1.83 1.76 10.5 10.9 7.5 29.6 0.75 2.3712 3.94 3.22 25.2 22.8 1.68 1.66 11.2 11.3 4.6 20.6 0.64 2.1721 2.76 2.46 17.8 17.5 1.30 1.22 8.4 8.6 2.9 29.8 0.20 1.7623 4.42 4.37 19.5 20.4 2.52 2.24 10.8 10.7 5.0 48.7 0.52 5.2024 4.16 3.42 19.4 17.0 2.30 2.20 11.5 10.5 2.6 49.3 0.53 0.4011 3.09 2.67 22.6 22.4 4.0 46.0 0.76 6.8119 2.25 2.09 14.9 15.2 4.7 32.6 0.66 3.7020 1.53 1.41 11.5 11.8 2.9 19.6 0.40 2.4322 2.58 3.05 15.9 18.3 2.4 8.8 0.15 0.55
Means 3.23 2.95 19.2 18.8 1.93 1.81 10.5 10.4 4.1 31.7 0.51 3.93
See Table I for explanation of abbreviations.
874 R. G. Schultze, H. S. Shapiro, and N. S. Bricker

TABLE VI IRepresentative Experiment of Response of a Dog to a Small Salt Load before and after the Development of Uremsa
Control Uremia
Intact kidney Remnant kidney Remnant kidney
UN&V/ UNaV/ UN&V/Hour GFR UNNV FLNa GFR UNaV FLNa GFR UNaV FLNa
ml/min uEq/hr % ml/min ;&Eq/hr % ml/min AEq/hr %
Control 44.5 648 0.19 5.3 149 0.37 6.2 1176 2.43
2.5 g of NaCI given by gastric tube with foodFirst 45.3 490 0.17 5.6 115 0.32 6.4 1134 2.22Second 44.0 1110 0.32 5.5 300 0.69 7.0 1680 2.92Third 44.3 2670 0.64 5.2 493 0.99 6.7 3348 6.41Fourth 40.4 1994 0.64 4.6 463 1.30 6.7 2562 4.97Fifth 40.7 1806 0.57 4.6 436 1.22 6.4 1670 3.235 hr total 8070 1956 11394Expected total* 3240 745 5880Difference 4830 1211 5514
See Table I for explanation of all abbreviations.* Value obtained by multiplying excretion rate in control hour by 5.
in GFRper nephron; and (e) a regulated reduction in number. The patterns of sodium excretion changed in afractional reabsorption of filtered sodium by the renal fashion that was both striking and appropriate for thetubules, largely unrelated to any of the foregoing maintenance of sodium balance. The data provide somefactors. insight into which the possible contributory factors may
The experiments were designed to differentiate be- have served in a primary role in the functional changes.tween these events. Each of the dogs studied had one The experimental model precludes the contribution ofkidney with a markedly reduced nephron population and damaged residual nephrons to the genesis of the natri-a contralateral control organ, with a normal comple- uresis per nephron. The experimental kidneys were "rem-ment of nephrons. Both sodium intake and caloric in- nant" organs, and there was no apparent intrinsic renaltake were maintained constant throughout the periods of disease in the surviving nephron population. The ex-study. The patterns of sodium excretion by the remnant perimental design also excludes the contribution ofkidneys were studied first with the contralateral normal mineralocorticoid insufficiency in that each of the dogskidney in situ. Then the normal kidney was removed was maintained on a large dose of a potent salt-retainingand the transformation that took place in the residual steroid throughout the period of study. The contributionnephrons of the remnant kidney was observed often in of a sustained natriuresis mediated by poorly reabsorb-serial fashion as uremia evolved. The opportunity thus able solutes is more difficult to examine directly, butexisted to compare sodium excretion by the same neph- two observations suggest that this mechanism probablyrons in the same animals first under conditions wherein was not the key factor in the genesis of the natriuresis.the total neprhon population exceeded 60% of the The first observation is that the natriuresis began withinoriginal number, then under conditions where the total 18 hr of removal of the normal kidney, an interval toonephron population approximated 10-20% of the original short to allow for substantial retention of poorly reab-
TABLE VIIISummary of 31 Experiments of the Type Depicted in Table VII
Control Uremia
Intact kidney Remnant kidney Total Remnant kidney
UNaV, MEq/Shr 6954(SE) ±i1193 1960(SE) --189 8914(SE) -±1323 11705(SE) --759Expected* 3524 --384 1468 --118 4092 -±510 9575 -765Difference 3430 --894 492 --146 3922 -±1016 2130 --533
* Value obtained bv multiplying excretion rate in control hour by 5.
Studies on the Control of Sodium Excretion 875

sorbable solutes. The second observation is that the re-tention of such solutes per se was not associated with anatriuretic effect in animals maintained on a low salt diet.Thus an increase in sodium excretion per nephron occur-red before the overt retention of impermeable soluteswhen salt intake wsas maintained at 3-5 g/day and markednatriuresis per nephron did not occur despite overt soluteretention when the salt intake was restricted. Wewouldfavor the view, therefore, that while poorly reabsorbablesolutes may contribute to an increase in sodium excre-tion rate per nephron in uremia, a regulated natriuresiswas probably not dependent upon such a mechanism;nor would such a mechanism appear to allow for the typeof regulatory ability that the chronic uremic patientpossesses in altering sodium excretion rates in accord-ance with sodium intake (5).
An adaptive increase in GFRper nephron could wellincrease the excretory capacity for sodium in survivingnephrons, and it is of interest that GFRdid increase inthe remnant kidneys in most animals after contralateralnephrectomy. Yet, the increase in GFR was not uni-versal, and the natriuresis appeared in those animals inwhich GFRdid not increase as well as in those in whichhyperfiltration did develop. Moreover, when GFR pernephron -was reduced acutely in the remnant kidneys ofuremic animals, sodium excretion rates remained ap-preciably higher than the values observed in the samekidneys in prenephrectomy studies. A third observationmay be relevant: in several animals maintained on a lowsalt diet, GFR increased but the natriuresis was sub-stantiallv smaller than in the dogs maintained on a highersalt intake. Thus, hyperfiltration could well contributeto the capacity of the residual nephrons to increase therate of sodium excretion; but natriuresis can occurwithout it and need not occur with it. Therefore it seemsunlikely that hyperfiltration is the key modulating eventin the control system for sodium excretion in uremia.Two recent studies have provided evidence that is con-sistent with this interpretation. In both, GFR was in-creased without a concomitant alteration in ECF volume.Lindheimer, Lalone, and Levinsky raised GFR in nor-mal dogs by up to 30% either by feeding of high pro-tein meal, administering glucocorticoids, or infusingdopamine (6). Sodium excretion rates increased onlyslightly, and fractional sodium excretion rates changedlittle. Slatopolsky, Elkan, Weerts, and Bricker raisedGFR acutely in uremic patients (5) and in normalindividuals (7) by infusing parathyroid hormone. Al-though filtered load of sodium increased by an averageof 20.4 and 18.9% in uremic patients and normal indi-viduals respectively, sodium excretion rates increasedmodestly, and fractional sodium excretion remained es-sentially unchanged. In the present studies, fractionalsodium excretion increased from an average of 0.89 to
6.75% in the remnant kidney of dogs receiving 3-5 g ofNaCl per day.
The evidence, therefore, points to the modulation oftubular reabsorption of sodium by a mechanism unrelatedto any of the foregoing factors. Phenomenologically, thenatriuresis has characteristics in common with thenatriuretic response to saline loading. As was first shownby De Wardner, Mills, Clapham, and Hayter (8) andsubsequently confirmed in many laboratories, the natri-uresis after acute saline loading is not dependent oneither an increase in GFR or a decrease in mineralo-corticoid stimulation. Similarly in the present dogs, theincrease in sodium excretion rate by the remnant kid-neys produced by the removal of the contralateral nor-mal organs did not require either an adaptive increasein GFRor a decrease in mineralocorticoid hormone ac-tivity. The natriuresis attendant upon both saline load-ing in normal animals and nephron reduction in ani-mals on a modest salt intake is an appropriate responsefor the preservation of sodium balance and ECF volume.The question therefore must be asked as to whether themarked natriuresis in the remnant kidneys of the uremicmodels relates to chronic expansion of either ECF vol-ume or of blood volume. The present studies do not sup-port either of these possibilities. Within the limits ofthe methods used, neither ECF volume (inulin space)nor blood volume was greater in the uremic state thanin the prenephrectomy control studies in the same ani-mals, although sodium excretion by the remnant kidneyswas increased by over 700%. Theoretically a homeo-static mechanism requires some displacement from the"normal" state in order to effect a change in end organresponse; in the case of the volume control system in theuremic animals, this displacement appears to be veryslight if the afferent limb detects an alteration in ECFor blood volume. The failure to detect an increase inblood volume or inulin space in the uremic dogs mightalso suggest that the control system continues to operatewith striking efficiency.
In comparing the response of the remnant kidneys toa fixed load of sodium chloride given by four differentroutes of administration, the natriuresis was increasedgreatly in the uremic vs. the prenephrectomy state.Thus the nephrons of the remnant kidneys excretedfour times as much sodium in the same time intervaland after the same amount of salt when the normal kid-ney was absent than when it was present and con-tributing to urine formation. Indeed, the remnant kidneyexcreted more sodium than the remnant and the normalkidneys combined had excreted in the same animals be-fore nephrectomy. When the timed response over the 5hr interval was compared, no lag period was observedin the excretion response of the remnant kidney in theuremic state. Thus it cannot be assumed that the ex-
876( R. G. Schultze, H. S. Shapiro, and N. S. Bricker

cretion lagged behind such that the stimuli to increasedexcretion multiplied over the 5 hr interval.' Ratherhowever, from the present data, as well as those fromuremic man (5), it would seem that as steady-stateGFR decreases due to a permanent reduction in neph-ron population, the total system becomes more sensitiveand more responsive to a given displacement from thesteady state. The expression of this change is an en-hanced response of the individual nephrons to a givenload of sodium. It is unclear whether this enhancementinvolves an increased sensitivity of the detector mecha-nisms of the volume control system in uremia, a greaterresponsivity of the efferent mechanisms to a given dis-placement from the steady state, or to an exaggeratedend organ response to a given efferent stimulus.
A number of different events have been shown to in-fluence sodium excretion under specific experimentalcircumstances. During saline loading fractional reab-sorption of sodium in the proximal tubule diminishesstrikingly. Martino and Earley have presented evidencethat physical factors may play a role in this suppression(9). Among these physical factors are decreased renalvascular resistance, a decreased plasma protein concen-tration, and increased renal perfusion pressure. Lewyand Windhager have presented evidence that peritubularoncotic pressure will influence proximal sodium reab-sorption and have suggested that changes in filtrationfraction by virtue of the associated changes in peri-tubular capillary protein concentration may exercise aninfluence over the rate of sodium reabsorption (10).Biochemical factors also may contribute to the natriure-sis of saline loading. Recently Waugh and Kubo haveshown that when substrates normally utilized by thekidney are added in small amounts to the infused saline,natriuresis is blunted (11). Finally, there is evidencefrom several sources that a humoral material withnatriuretic capacity appears in the plasma of saline-loaded animals (8, 12, 13). The natriuresis of salineloading also appears to involve inhibition of sodium re-absorption beyond the proximal tubule. However, to date,the mechanisms involved in this inhibition remain to beelucidated.
Which of the foregoing mechanisms is responsible forthe natriuresis of nephron reduction and for the enhancedsensitivity of the control system is not established by the
'The base line sodium excretion rates were higher in thepostnephrectomy than in the prenephrectomy studies on thesame dogs. The apparent explanation for this is that thediurnal rhythm for sodium excretion was altered in the ure-mic state. Thus in serial measurements of sodium excretionmade over 24-hr intervals, an increase in excretion rate oc-curred between 1 and 4 a.m. in the prenephrectomy studies,but not in the postnephrectomy studies.
present observations. It seems unlikely, however, thatdilution of plasma proteins or of substrate plays a rolesince the amounts of saline administered wvere small andthe same load of salt was administered before and afterthe development of uremia. Preliminary observationssuggest that blood pressure does not change consistentlyafter removing the normal organ, and data also of apreliminary nature suggest that no changes in cardiacoutput occur. The relative contribution of a change inthe activity of a natriuretic hormone vs. changes inphysical factor(s) within the renal interstitium re-main for future investigation.
REFERENCES1. Desautels, R. E. 1957. Hemisection of the bladder for the
collection of separate urine samples. Surg. Gynecol. Ob-stet. Int. Abstr. Surg. 105: 767.
2. Deane, N. 1952. Methods in Medical Research. A. C.Corcoran, editor. Yearbook Medical Publishers Inc., Chi-cago. 5.
3. Albert, S. N. 1966. Blood volume. J. Amter. Med. Ass.198: 770.
4. Bonsnes, R. W., and H. H. Taussky. 1945. On the colori-metric determination of creatinine by the Jaffe reaction.J. Biol. Chem. 158: 581.
5. Slatopolsky, E., I. 0. Elkan, C. Weerts, and N. S.Bricker. 1968. Studies on the characteristics of the con-trol system governing sodium excretion in uremic man.J. Clin. Invest. 47: 521.
6. Lindheimer, M. D., R. C. Lalone, and N. G. Levinsky.1967. Evidence that an acute increase in glomerular fil-tration has little effect on sodium excretion in the dogunless extracellular volume is expanded. J. Clin. Invest.46: 256.
7. Slatopolsky, E., P. Hoffsten, J. Mc Culley, and N. S.Bricker. 1968. On the demonstration of a third factor(s)in the natriuresis induced by ECF volume expansion inman. J. Lab. Clin. Med. 72: 1018.
8. De Wardener, H. W., I. H. Mills, XV. F. Clapham, andC. J. Hayter. 1961. Studies on the efferent mechanism ofthe sodium diuresis which follows the administration ofintravenous saline in the dog. Clin. Sci. 21: 249.
9. Martino, J. A., and L. E. Earley. 1967. Demonstrationof a role of physical factors as determinants of the na-triuretic response to volume expansion. J. Clin. Invest.46: 1963.
10. Lewy, J. E., and E. E. Windhager. 1968. Peritubularcontrol of proximal fluid reabsorption in the rat kidney.Amer. J. Physiol. 214: 943.
11. Waugh, W. H., T. Kubo. 1968. Antinatriuretic effectof exogenous metabolic substrate on sodium diuresis ofsaline infusion. Life Sci. 7: 35.
12. Lichardus, B., and J. W. Pearce. 1966. Evidence for ahumoral natriuretic factor released by blood volume ex-pansion. Nature (London). 209: 407.
13. Johnston, C. I., J. 0. Davis, S. S. Howards, and F. S.Wright. 1967. Cross-circulation experiments on themechanism of the natriuresis during saline loading inthe dog. Circ. Res. 20: 1.
Studies on the Control of Sodium Excretion 877