studies on steroid fever -...
TRANSCRIPT

Studies on Steroid Fever
I. PRODUCTIONOF LEUKOCYTEPYROGENIN VITRO BY ETIOCHOLANOLONE
PHYLLIs BODEL and Momis Dn.LmiD
From the Department of Internal Medicine,Medicine, New Haven, Connecticut
A B S T RA C T When a serum-buffer solution ofetiocholanolone is incubated with human bloodleukocytes in vitro, a pyrogen is released. Likeendogenous pyrogen of leukocyte origin, thispyrogen produces prompt monophasic fevers inrabbits, does not induce fever tolerance when givendaily, and is inactivated by trypsin. In many re-spects, the characteristics of the in vitro reactionresemble experimental steroid-induced fever. Forexample, release of pyrogen varies directly withthe concentration of steroid. 4-8 hr of contact be-tween steroid and leukocyte is required for activa-tion of the cell. Rabbit leukocytes are not activatedby etiocholanolone. Finally, androsterone, the 5a-isomer of etiocholanolone, does not induce pyrogenrelease in vitro. These studies suggest that experi-mental steroid fever in man may be mediated byan endogenous pyrogen released from leukocytes.
INTRODUCTION
In 1956 it was reported that the intramuscularinjection of a naturally occurring steroid hor-mone metabolite, etiocholanolone (3a-hydroxy-etiocholane-17-one), produced fever in man (1).Many subsequent experiments have since con-firmed these findings and have provided additionalinformation characterizing this response (2, 3).
Dr. Bodel is a Research Associate in Medicine. Dr. Dil-lard is an Assistant Professor of Medicine. Address re-quests for reprints to Dr. Phyllis Bodel, Yale UniversitySchool of Medicine, 333 Cedar Street, New Haven, Conn.06520.
Received for publication 5 June 1967 and in revisedform 11 September 1967.
Yale University School of
Tests with etiocholanolone and with steroidsclosely related in structure indicate that theseagents produce fever only in man (4). Unlikefever induced by bacterial endotoxin, which de-velops within 1-11/ hr after intravenous injection,there is a delay of 416 hr before the onset ofsteroid-induced fever, which may then last for12 hr or more (2). Pyrogenic activity of varioussteroids is related to their particular structuralfeatures (5), and appears to depend on a 5,8-configuration (1).
Although local inflammation characteristicallyfollows intramuscular injection of the pyrogenicsteroids (6), and increased numbers of granulo-cytes appear in the blood at the time of the fever(2), the unusual features of steroid fever have ledto the suggestion that these agents elevate tem-perature by a mechanism that does not involve thewhite cell, and therefore, steroid fever differs fromfevers caused by various microbes and their prod-ucts (7). Since there is evidence that certain typesof periodic fever are associated with abnormalsteroid metabolism (8, 9), it seemed worthwhile tolook for the pathways for the pyrogenic action ofetiocholanolone.
Recent observations have demonstrated that hu-man leukocyte pyrogen, produced in vitro byincubation of blood leukocytes with endotoxin orheat-killed staphylococci, can be detected by in-injection into rabbits (10). Similar experiments,therefore, seemed appropriate in order to examinepossible effects of etiocholanolone on pyrogen pro-duction by human blood leukocytes.
The Journal of Clinical Investigation Volume 47 1968 107

METHODSAll glassware and standard reagents were renderedsterile and pyrogen-free by heating at 1600 C or auto-claving for 2 hr. Tris buffer and enzyme preparationswere filtered through an ultra-fine sintered glass filterand tested for pyrogenicity. Techniques for injection ofrabbits and temperature recording have been reportedpreviously (11). In early experiments, each rabbit re-ceived injections for only 2 or 3 consecutive days toavoid fevers arising from sensitization to human ma-terials. Later it was found that hypersensitivity fevers didnot occur in most rabbits until after 5 days of injections,as long as exposure to human materials was limited tosupernatants of incubated leukocytes as described below.
Preparation of leukocytes. 100 ml of blood containing10 U of heparin per ml was obtained from normal malevolunteers. To this was added 200 ml of 3% dextran1in saline. After 20-30 min at room temperature, the leuko-cyte-rich supernatant was removed, centrifuged at 300 gfor 15 min, and the cell buttons were washed once withKrebs-Ringer phosphate (KRP) buffer,2 containingheparin, 10 U/ml. After a second centrifugation, mostremaining red cells were lysed by brief exposure to dis-tilled water (12) and the leukocytes were washed oncemore with heparinized buffer and centrifuged. Leukocyteswere then counted on a Coulter Particle Counter and ad-justed to a concentration of 9-10 X 107 per ml.
Monocytes. The method of Bennett and Cohn (13)was used to prepare from normal blood a leukocyte frac-tion which contained predominantly mononuclear cells.100 ml of heparinized blood was mixed with 200 ml of 3%dextran in saline, and the red cells were allowed tosettle for 20-30 min. The supernatant was then removed,centrifuged at 300 g for 15 min, and the cell buttons wereresuspended in 5-8 ml of 25% human serum albumin.3This mixture was centrifuged for 36 min at 12'C at 2400g. The cells at the top of the tubes were carefully re-moved, along with a few milliliters of albumin, 30 ml ofheparinized KRP buffer was added, and the suspensionwas centrifuged again at 300 g for 15 min. The cells werethen suspended in a few milliliters of buffer or autologousserum, and a cell count and differential were performed.50-70% of the leukocytes were monocytes, the rest beingmostly small lymphocytes. Polymorphonuclear leukocytesvaried from < 1 to 15%.
Steroids. Etiocholanolone (Mann Research Labs., Inc.,New York) was autoclaved for 1 1/2-2 hr in an Erlen-meyer flask. KRP buffer was added and the flask wasshaken overnight in a Dubnoff incubator at 370C. Thesaturated solution was then passed through an ultra-fine sintered glass filter, and the filtrate was stored at40C. Solutions of androsterone were prepared in the sameway. Identification and characterization of the steroids
1 Dextran, M.W. 100,000-200,000 Nutritional Biochem-icals Corp., Cleveland, Ohio.
2 Modified to contain 14 the concentrations of Ca`+ andMg+', to prevent precipitation.
sNormal human serum albumin, salt-poor, Ameri-can National Red Cross.
were performed using various chromatographic pro-cedures. A 96% methanol:heptane system was usedfor paper chromatography. The colunui method employedthe gradient elution technique of Lakshmanan and Lieber-man (14). Gas-liquid chromatography was performed oila 1%o XE-60 coating of Celite (nmesh 100-120) in a 6 ftby 4 mmcoiled column with a flanme detector. Each pro-cedure was standardized en1l)loying kinown steroids whosepurity was demonstrated by melting point, mixed meltingpoint, and infrared spectroscopy. Steroid concentrationswere determined by using the Zimmerman colorimetrictechnique (15). Solutions of radioactive etiocholanoloneand androsterone of high specific activity, prepared bythe Wilzbach exchange method (16), were niade in anidentical manner, and aliquots were added to somie ex-perimcaits in order to follow any small changes in thesteroids during incubation.
Incubation. 50-mIl Erlenmleyer flasks contained 1 mlof the leukocyte preparation, 6 ml of heparinized KRPbuffer or a buffer solution of etiocholanolone, 1.2 ml ofautologous serum, 1.5 mg of glucose per nml, and 6000 Uof penicillin. The total incubation volume in each flaskwas 8-9 ml. The flasks were shaken onl a D)ubnoff incu-bator at 370C for 2-3 hr, and then placed in a stationaryincubator at 370C overnight. After about 18 hr of incu-lation, the flask contents were removed, centrifuged at 520g for 15 min, and the supernatants were either injectedat once or stored at 4°C for several (lays. Aln aliquotfrom each flask was cultured in thioglycollate broth toconfirm sterility. All experiments in which cultures werenot sterile were discarded. In each experiment a controlflask of white cells in serum-buffer was incubated, as wellas various experimental flasks. Supernatants from allflasks in each exl)eriment were usually injecte(l into thlesame two rabbits.
Rabbit leukocytes. Rabbit blood was obtained bycardiac puncture, and processed as described for humanblood. Techniques for preparing spleen and lymph nodecells have been described previously (11). Briefly, tissueswere minced, forced through a wire gauze with saline,and white cells were counted. Lung cells were obtained bya modification (17) of the technique of Myrvik, Leake,and Fariss (18). All mononuclear cells in these ex-periments were obtained from rabbits previously in-fected with BCG.
Leukocytes were suspended in KRP buffer and 15%7onormal rabbit serum for incubation, with added glucoseand penicillin as described previously. 1-2 X 108 cells wereusually added to each flask, in a total volume of 15 ml.Heat-killed staphylococcus albus, prepared as describedpreviously (10), was added to one incubation flask forsome experiments, in a ratio of 30-50 bacteria per whitecell.
Fever index. The area under fever curves 2 hr afterinjection was measured as previously described (19).
RESULTS
To determine whether etiocholanolone could ini-tiate release of pyrogen by leukocytes in vitro, we
108 P. Bodel and M. Dillard

FEVER(OC)
I.0-
0.5-
0-
+ Etio
I I lI0 1
TIME (Hours)
FIGURE 1 Febrile responses in one rabbit to injection of supernatantsfrom 5.0X107 white cells (WBC), incubated with or without etio-cholanolone (Etio) at a final concentration of 39 ,Ag/ml, or from etio-cholanolone incubated alone.
incubated human white cell preparations for 18 hrwith a buffered solution of etiocholaholone and15%o serum. Control leukocytes were incubated ina serum-buffer medium alone. The supernatantswere removed, and from each flask a volumederived from the same number of white cells(4-5 x 107) was injected into each rabbit. Theresults from a typical experiment are shown inFig. 1. Supernatants from the leukocytes incubatedwith etiocholanololne were p)yrogenic, whereas con-trols containing either leukocytes alone or etio-cholanolone incubated without leukocytes wereinactive. The shape of this fever curve, includingits early onset and rapid defervescence, resemblesthe curves l)rodtlce(l by injection of both rabbit(20) and human (10) leukocyte pyrogen. Theseresults were rep)eate(lly confirmed. In a series of25 consecutive exl)eriments, etiocholanolone failedto induce significant pyrogen only twice. In twoother experiments of this series, the supernatantderived from leukocytes incubated in serum-bufferalone contained pyrogen (> 0.3C average max-imum fever). This occasional occurrence of "spon-taneous" activation has been reported previously(10). All other experimeflts in the series gaveresults which were similar to those shown inFig. 1. The data from these experiments areaveraged in Table I. These findings, then, suggestthat a leukocyte pyrogen is produced when humanwhite, blood cells are incubated in the presence ofetiocholanolone.
The following controls were devised to exclude
the possible role of endotoxin contamination. Solu-tions of etiocholanolone were injected in volumesof 30 ml or more and were nonpyrogenic evenwhen preincubated with 15% rabbit serum at370 C for '/2 hr to potentiate the action of anyendotoxin that might be present. The possiblecontribution of endotoxin from other sourceswould appear to be eliminated by the negativecultures of all tested material and by the absenceof pyrogen in the supernatant from control leuko-cytes. Injections of etiocholanolone incubated aloneas controls in various experiments were uniformlynonpyrogenic. These curves have been omittedfrom subsequent figures for simplicity.
In order to confirm that the pyrogen being pro-duced during incubation of white cells with etio-cholanolone was a protein, like the endogenouspyrogens previously described of rabbit (21) andhuman (10) origin, we attempted to inactivate
TABLE IHuman Blood Leukocytes Incubated
with Etiocholanolone
No. of Maximum feverexperiments WBC* ('C)
WBC 21 47.01:1.7§ 0.061:0.017 (31)11WBCplus 21 47.0i1.7 0.75 40.047 (36)
etiocholanolone
* Average number of leukocytes from which supernatant was derivedfor each injection.
I Average temperature elevation above base line.§ SE of mean.
II Numbers of rabbits injected.
Production of Leukocyte Pyrogen In Vitro by Etiocholanolone 109

FEVER(IC)
I.0 -
I.-4
0.5 -
0-
.,x*"mePyrogen
/x
-x-, XPyrogen + Trypsin
.o ' 0,
0 l
TIME (Hours)
FIGURE 2 Average fevers in rabbits after injection ofpreformed etiocholanolone pyrogen, incubated in Trisbuffer, pH 8.4, with and without 3 mg of trypsin per in-jection. In this and the following figures, unless otherwisestated, each curve represents the average responses oftwo rabbits, and the same rabbits received all injectionsfor each experiment.
preformed "etiocholanolone pyrogen" by trypsinand pronase. Supernatant from an 18 hr incuba-tion of leukocytes and etiocholanolone was dividedinto two parts. To each was added 0.15 M Trisbuffer pH 8.4, CaCl2 to a final concentration of10-3 M, and to one flask 6 mg of trypsin .4 In otherexperiments, 0.5 mg of pronase5 was added in-
4 Trypsin, 2 X crystallized (salt-free), Nutritional Bio-chemicals Corp., Cleveland, Ohio.
5Pronase, B grade, Calbiochem, Los Angeles, Calif.The enzyme in buffer was preincubated for 2 hr beforeuse.
FEVER(oC)
I.0-
0.5-
0-
0F
stead of trypsin. Total volumes were 12-15 ml.After overnight incubation at 370C, both controland enzyme-treated samples were tested in tworabbits. The results of one experiment with trypsinare given in Fig. 2. Identical results were obtainedwith pronase. Since both enzymes removed thepyrogenic activity from such supernatants, "etio-cholanolone pyrogen" appears to have an essentialprotein nature like other endogenous pyrogens.
Since phagocytosis activates leukocytes to re-lease pyrogen (10), it seemed essential to showthat the activating properties of this preparationof etiocholanolone were not due to the presence ofvery small particles of crystalline steroid. Thefollowing experiment was designed to test thishypothesis. A solution of etiocholanolone was pre-pared in the usual way (see Methods). After fil-tration, a portion of this solution was spun at105,000 g for 2 hr in a Spinco Model L Ultracen-trifuge, in tubes which had been autoclaved for11/2 hr to inactivate any endotoxin. The super-natant was then carefully removed and incubatedwith white cells, along with an unspun aliquot ofthe original filtrate. As shown in Fig. 3, there wasequal pyrogen production from cells incubatedwith both samples. Since centrifugation at thisspeed should have removed all particles of greaterthan 400,000 molecular weight (22), it is ex-tremely unlikely that the presence of microcrystalsof steroid could be responsible for the activation ofleukocytes in our system.
WBC+ Etio (S-2)
--A WBC +Ei S
2
TIME (Hours)FIGURE 3 Average fevers in rabbits after injection of supernatantsfrom 5.0 X 107 white cells (WBC) incubated alone, with a solutionof etiocholanolone 39 ,ug/ml (S-i), or supernatant of the same solutionafter centrifugation at 105,000 g for 2 hr (S-2).
110 P. Bodel and M. Dillard

It seemed possible that changes had occurred insteroid structure during either its preparation orincubation with leukocytes. Chromatography, melt-iiig points, and infrared spectroscopy, as well aLslaleledl etiocholaiiolone, were used to examine thispossibility. No detectable change occurred in thesteroid either after autoclaving or after an 18-hrincubation with white cells. Thus, a buffer solu-tion of etiocholanolone, containing about 30 ,Fg/ml,appeared to be capable of activating white cells torelease pyrogen.
Since a closely related steroid, androsterone,which differs only in having a 5a-configuration,has been reported to be usually nonpyrogenic inhuman injection studies (3), attempts were madeto induce production of pyrogen in the in vitrosystem with solutions of this compound. Andro-sterone solutions were prepared in the same wayas were those of etiocholanolone. In seven experi-ments, in which several different preparations ofsteroid were used, varying in concentration from14 to 40 ug/ml, androsterone was completely non-pyrogenic five times. In two experiments, leuko-cytes incubated with androsterone released smallamounts of pyrogen. One experiment is shownin Fig. 4. In this experiment both etiocholanoloneand androsterone were present in nearly equalconcentrations. These results indicate that the twosteroids, etiocholanolone and androsterone, havedifferent capacities for inducing in vitro pyrogenrelease from leukocytes under these conditions.
In the next group of experiments, the relationbetween the release of pyrogen and the concentra-
FEVER(OC)
1.0-
0.5-
0-
FEVER(0c)
0.5-
01
WBC+Etiox
X--,
x-'
WBC
WBC+Andro0
TIME (Hours)
FIGURE 4 Average fevers in rabbits after injection ofsupernatants from 4.0 X 10' white cells (WBC) incu-bated alone, with 29 ,ug/ml androsterone, or 27 Ag/mletiocholanolone.
tion of etiocholanolone was examined. The usualconcentration of a buffer-saturated solution ofetiocholanolone was 20-40 tg/ml. Varying dilu-tions of steroid were used in four experiments, oneof which is shown in Fig. 5. When the concentra-tion was decreased 2- to 3-fold, production ofpyrogen was sharply reduced in all experiments.Production was usually maximal with levels of20-30 jug/ml (the highest concentrations tested),but when the concentration of steroid was reducedto 10 ug/ml or less, little or no pyrogen wasevolved.
Since one of the unusual characteristics of thepyrogenic steroids is their ability to evoke feveronly in man, it was of interest to determinewhether leukocytes from rabbits could releasepyrogen in vitro. For these experiments, rabbitand human cells were prepared in the same way,and the solutions of steroid used were known tobe effective when incubated with human cells.
WBC+ Etio
/'I x .720oLg/ml
i/ 30 jig /ml _ 0
/ ,- - <0 NN~~~~~~N
/ 'IO 1O/Ag/mI+ 6 sWBColone
I I
2I
TIME (Hours)FIGURE 5 Average fevers in rabbits after injection of supernatantsfrom 5.5 X 107 white cells (WBC) incubated alone or with varyingconcentrations of etiocholanolone (Etio).
Production of Leukocyte Pyrogen In Vitro by Etiocholanolone ill
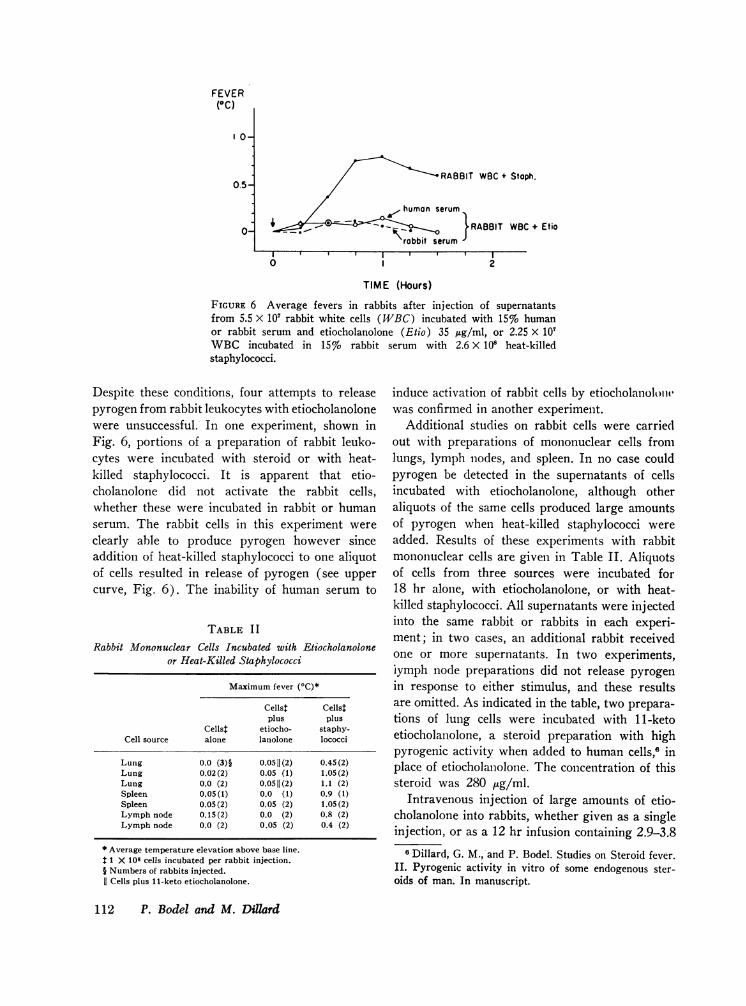
FEVER(OC)
0-
0.5-
0-
RABBIT WBC+ Staph.
ehuman serum
}RABBIT WBC+ Etiorabbit serum
0 2
TIME (Hours)
FIGURE 6 Average fevers in rabbits after injection of supernatantsfrom 5.5 X 107 rabbit white cells (WBC) incubated with 15% humanor rabbit serum and etiocholanolone (Etio) 35 tug/ml, or 2.25 X 107WBCincubated in 15% rabbitstaphylococci.
Despite these conditions, four attempts to releasepyrogen from rabbit leukocytes with etiocholanolonewere unsuccessful. In one experiment, shown inFig. 6, portions of a preparation of rabbit leuko-cytes were incubated with steroid or with heat-killed staphylococci. It is apparent that etio-cholanolone did not activate the rabbit cells,whether these were incubated in rabbit or humanserum. The rabbit cells in this experiment wereclearly able to produce pyrogen however sinceaddition of heat-killed staphylococci to one aliquotof cells resulted in release of pyrogen (see uppercurve, Fig. 6). The inability of human serum to
TABLE I IRabbit Mononuclear Cells Incubated with Etiocholanolone
or Heat-Killed Staphylococci
Maximum fever (oC)*
Cellst Cellstplus plus
Cellst etiocho- staphy-Cell source alone lanolone lococci
Lung 0.0 (3)§ 0.0511(2) 0.45 (2)Lung 0.02 (2) 0.05 (1) 1.05(2)Lung 0.0 (2) 0.0511 (2) 1.1 (2)Spleen 0.05(1) 0.0 (1) 0.9 (1)Spleen 0.05(2) 0.05 (2) 1.05(2)Lymph node 0.15(2) 0.0 (2) 0.8 (2)Lymph node 0.0 (2) 0.05 (2) 0.4 (2)
* Average temperature elevation above base line.t 1 X 108 cells incubated per rabbit injection.§ Numbers of rabbits injected.CICells plus 11-keto etiocholanolone.
serum with 2.6 X 10 heat-killed
induce activation of rabbit cells by etiocholaniolotiwas confirmed in another experiment.
Additional studies on rabbit cells were carriedout with preparations of mononuclear cells fromlungs, lymph nodes, and spleen. In no case couldpyrogen be detected in the supernatants of cellsincubated with etiocholanolone, although otheraliquots of the same cells produced large amountsof pyrogen when heat-killed staphylococci wereadded. Results of these experiments with rabbitmononuclear cells are given in Table II. Aliquotsof cells from three sources were incubated for18 hr alone, with etiocholanolone, or with heat-killed staphylococci. All supernatants were injectedinto the same rabbit or rabbits in each experi-ment; in two cases, an additional rabbit receivedone or more supernatants. In two experiments,iymph node preparations did not release pyrogenin response to either stimulus, and these resultsare omitted. As indicated in the table, two prepara-tions of lung cells were incubated with 11-ketoetiocholanolone, a steroid preparation with highpyrogenic activity when added to human cells,6 inplace of etiocholanolonie. The concentration of thissteroid was 280 jug/ml.
Intravenous injection of large amounts of etio-cholanolone into rabbits, whether given as a singleinjection, or as a 12 hr infusion containing 2.9-3.8
6 Dillard, G. M., and P. Bodel. Studies on Steroid fever.II. Pyrogenic activity in vitro of some endogenous ster-oids of man. In manuscript.
112 P. Bodel and M. Dillard

FEVERINDEX
HOURS OF EXPOSURETO ETIOCHOLANOLONE
FIGURE 7 Average fever indexes of rabbits after injection of super-natants derived from 18-hr incubations of 6.3 X 1O7 white cells. Eachaliquot of cells was exposed to etiocholanolone 32 /Ag/ml for varyingtimes before centrifugation and further incubation in serum buffer.
mg of steroid, did not produce fever. These lastdata confirm the work of others who have usedsomewhat different solvents and injection sched-ules (4).7
The long latent period (usually 4-6 hr) thatfollows the injection of steroid in man is an un-usual feature of this form of experimental fever.Further experiments were carried out with thein vitro system to determine how long the leuko-cyte requires exposure to etiocholanolone beforepyrogen production is initiated. The results of atypical experiment are shown in Fig. 7. Feverindexes rather than fever curves are plotted forconvenience. Control leukocytes in this experimentwere incubated for 18 hr in a serum-buffer me-dium. Experimental leukocytes were first incu-bated with etiocholanolone for varying intervals,then centrifuged, and the supernatant was re-moved. The cells were then suspended in freshserum-buffer for the remainder of 18 hr, afterwhich the second supernatant was removed. Thefirst and second supernatants were combined forinjections; thus, each fever index shown in Fig. 7represents the total pyrogen released during 18 hrfor each preparation. Cells exposed to etiocho-lanolone for only 2 or 4 hr. like control cells, didnot release a significant amount of pyrogen. How-ever, exposure of the cells to steroid for 6, 8, or18 hr all appeared to be equally effective in stim-
' Cohn, G. L., and E. Atkins. Unpublished observations.
ulating the cells to release pyrogen. In six othersimilar experiments the time of exposure requiredfor stimulation varied between 4 and 8 hr. Onlyone of five experiments in which the 4 hr incuba-tion time was tested showed pyrogen release after4 hr of exposure, whereas four of six experimentsin which cells were tested after 6 hr of incubationshowed pyrogen release. From these studies itseems clear that exposure to a solution of etio-cholanolone for a number of hours is required inorder for the leukocytes to become activated.
Since the blood leukocyte preparations in thesestudies contain several different cell types, andsince human mononuclear cells are now known torelease pyrogen under certain conditions (23), itwas of interest to determine which cell in theleukocyte preparations was responsible for produc-tion of pyrogen. In two experiments of differentdesign, blood mononuclear cells were clearly im-plicated as one source of pyrogen.
In the first experiment, blood leukocytes wereobtained on .two occasions from a patient withchronic agranulocytosis. Differential counts re-vealed only 3% eosinophils, with the remainderof the leukocytes either monocytes or lymphocytes.When the white cells from this patient were incu-bated with etiocholanolone, definite production ofpyrogen occurred. Sufficient pyrogen was releasedby 3-7 X 108 monocytes to cause an average feverof 0.7'C in five rabbits. The fever curves in these
Production of Leukocyte Pyrogen In Vitro by Etiocholanolone
.:i
...'.: ..:.
mI I 12 4 6 8
113

FEVER(OC)
AVERAGE M- 4.3 M- O.42 M- 2. 2NO.-CELLS
(XIO") G- 0.73 G- 5.0 G- 29.8
FIGURE 8 Average maximum fevers in rabbits after in-jection of supernatants from various white cell prepara-tions incubated with etiocholanolone. The average num-bers of monocytes (M) and granulocytes (G) contrib-uting to the supernatants is given below each bar. Thenumber of recipient rabbits is given in parentheses withineach bar. sE of mean is indicated. All blood cells wereobtained from normal donors except in two experiments(five recipients) included in the left-hand bar in whichleukocytes were from a patient with chronic agranu-locytosis.
rabbits exactly resembled those of normal bloodleukocyte pyrogen (see, for example, Fig. 1). Inthe second experiment, a preparation of monocytesand lymphocytes was obtained from normal bloodby differential centrifugation in human serum al-bumin (see Methods). Such preparations of mono-nuclear cells, containing less than 10%o granulo-cytes, were shown on three occasions to releasepyrogen following incubation with etiocholanolone.The total number of granulocytes in these prepara-tions was less than 1.5 x 106, a level well belowthe minimum of these cells required for significantproduction of pyrogen (see below). Results fromthese two types of experiments with mononuclearcells are summarized in the left-hand bar in Fig. 8.
It seems likely, then, that mononuclear cells canrelease pyrogen. following incubation with etio-cholanolone. Calculation of the per cent of mono-cytes and lymphocytes in these various prepara-tions indicates that release of pyrogen varieddirectly with the numbers of monocytes, and waseasily detectable when as few as 5 X 106 mono-cytes were present. The absolute number oflymphocytes, however, varied from less than 1-18X 106 in preparations with similar pyrogenicity.
Such evidence, although indirect, suggests that themonocyte is the important cell in these preparations.
Several experiments were designed to investigatethe action of etiocholanolone on human granulo-cytes. The results of some of these studies aresummarized in the right half of Fig. 8. Normalblood cells, containing 60-70%o granulocytes, re-leased little or no pyrogen when incubated withetiocholanolone in dosages of 3-6 x 106 granulo-cytes. When the numbers of granulocytes were in-creased to about 3 x 107, fevers of about 0.50Cwere produced in four experiments, in which num-bers of monocytes were between 1.5 and 3.5 x 100.Comparison with fevers induced by 4-6 X 100monocytes (see left half of Fig. 8) indicates thatetiocholanolone stimulates mononuclear cells to amuch greater extent than it does granulocytes.From results of several experiments in which num-bers of monocytes were very low, however, it ispossible that granulocytes can also release pyrogenafter incubation with steroid. Further studies willbe needed to confirm this inference.
DISCUSSION
Etiocholanolone and its isomer, androsterone, arenaturally occurring steroids which are formed asend products of androgen metabolism. Studies ofthe concentration of unconjugated etiocholanolonein normal plasma (24) have indicated that there isapproximately 1 ,ug/100 ml of plasma. When etio-cholanolone is injected (25, 26), it is rapidly con-jugated, cleared from plasma, and excreted in theurine in conjugated form, primarily as the glu-cosiduronate.
Etiocholanolone has no apparent androgenic ac-tivity, unlike its principal metabolic precursors,but it has antitumor activity in certain lymphomas(27), and recently has been shown to cause por-phyrin release from liver cells (28). The discoverythat etiocholanolone (1) and many other similarsteroids (5) are pyrogenic suggested that abnor-malities of steroid production or turnover mightgive rise to some clinical fevers. There is some evi-dence to support this hypothesis (8, 9). However,the exact role of these compounds in periodicfevers has not been clarified.
The studies reported here indicate that etio-cholanolone can activate human blood leukocytesto release an endogenous pyrogen in vitro. Al-
114 P. Bodel and M. Dillard

though it is not known whether this material isidentical to leukocyte pyrogen elicited by otherstimuli (10), the two agents resemble each otherclosely in all tested biologic activities. For example,both pyrogens elicit in rabbits a prompt mono-phasic fever which is usually maximal at 45-60min and rapidly defervesces. This fever curve dif-fers from those observed with many other pyrogens(7). Further, both agents are stable for weekswhen stored at 40C, and both are inactivated bytrypsin. 'Finally, rabbits do not develop toleranceto repeated injections of these leukocyte pyrogens,as they do to a number of microbial products.
In the following respects, release of endogenouspyrogen by human leukocytes exposed in vitro toetiocholanolone resembles the steroid fever thatfollows injection of etiocholanolone into humanvolunteers.
First, as reported by Kappas and Palmer (3),injection of androsterone, the 5a-analogue of etio-cholanolone, into volunteers did not elicit fever,except occasionally after repeated injections. Inour in vitro model, androsterone was tested underthe same conditions as etiocholanolone and wasnot pyrogenic in most experiments. Studies ofother steroids, both "pyrogenic" and "nonpyro-genic" by injection studies, are reported in anotherpaper,8 and confirm the specificity of the reaction.
Second, in our studies rabbit leukocytes failed torelease pyrogen when exposed to etiocholanolone.This was true whether or not human serum waspresent, or whether various mononuclear cell prep-arations were used. Similarly, this steroid hasrepeatedly failed to induce fever when injected ina variety of animals (4), suggesting that steroidfever occurs only in man. Our in vitro studies areat least consistent with this hypothesis, althoughexperiments with leukocytes from other specieswould provide more evidence. Since endogenouspyrogen from dog leukocytes does not producefever in rabbits (29), however, studies of pyrogenrelease from leukocytes of additional animals willonly be meaningful when it is established that thesepyrogens are active in rabbits.
A long period of incubation with steroid was re-quired in order to activate leukocytes in our experi-
8 Dillard, G. M., and P. Bodel. Studies on steroid fever.II. Pyrogenic activity in vitro of some endogenous steroidsof man. In manuscript.
ments. This unusual feature seems to correlate wellwith the long latent period described in vivo beforeonset of fever. By contrast, when endotoxin isadded to rabbit leukocytes in vitro, incubation ofthe cells for only 5 min or less is sufficient to in-duce activation (30). Similarly, endotoxin causesa prompt fever after injection in both rabbit andman (7). The activation of human cells by phago-cytosis also occurs within 1 hr after bacteria areadded.9 The reasons for the delay in activation ofleukocytes by etiocholanolone, which is presumablyrelated to the mode of action of the steroid on thewhite cell, are presently unknown. The steroid mayact on a different cell than the one stimulated byendotoxin or by phagocytosis, or it may activatethe cell by different pathways. It seems very un-likely that such a time delay in vitro is due to aproduct of steroid conversion, since no change incomposition of radioactive steroid could be demon-strated after incubation with leukocytes.
A relationship between steroid fever and leuko-cytes is suggested by previous observatioiis thatmost pyrogenic steroids produce marked localinflammation and leukocytosis (6), and that leuko-penic patients injected with etiocholanolone reactwith little inflammation or fever (31). However,it would be premature to propose a direct relation-ship between our in vitro studies and such injec-tion experiments in human volunteers. Studies inrabbits have shown that leukocytes from lung,lymph node, and spleen are all capable of pyrogenrelease (17), as are monocytes from peritonealexudates (32), and since the sites to which steroidis carried after injection are not known, it is pos-sible that blood leukocytes play no role in thefever induced by this agent. These studies do sug-gest, however, that steroid fever in man ismediated by an endogenous pyrogen released fromleukocytes of some kind. Since pyrogenic steroidsrelease lysosomal enzymes from suspensions ofrabbit leukocyte granules and liver lysosomes(33), their ability to activate human cells may alsodepend upon alterations in membrane permeability.Changes in membrane structure probably precederelease of endogenous pyrogen in other experi-mental models (7).
The contribution of steroids such as etiocho-lanolone to clinical fevers is still not clearly defined.
9 Bodel, P., and E. Atkins. Unpublished observations.
Production of Leukocyte Pyrogen In Vitro by Etiocholanolone 115

The capacity of this steroid to activate leukocytesto release pyrogen, however, indicates that thissteroid and related compounds may have importantbiological activities in man, including the capacityto induce fever. It is likely that further studies ofthe action of pyrogenic steroids on cells may leadto a better understanding of the role of these com-pounds in health and disease.
ACKNOWLEDGMENTS
We thank Dr. Elisha Atkins, in whose laboratory mostof this work was done, for his interest, encouragement,and invaluable advice, and Dr. Philip Bondy for his in-terest and support. Wealso thank David Kelly and Mrs.James Wechsler for excellent technical assistance.
This work was supported by grants (AI-01564 andAM-00254) from the U. S. Public Health Service.
REFERENCES
1. Kappas, A., L. Hellman, D. K. Fukushima, and T. F.Gallagher. 1957. The pyrogenic effect of etiocholanol-one. J. Clin. Endocrinol. Metab. 17: 451.
2. Kappas, A., P. B. Glickman, and R. H. Palmer. 1960.Steroid fever studies: Physiological differences be-tween bacterial pyrogens and endogenous steroid pyro-gens of man. Trans. Assoc. Am. Physicians. 73: 176.
3. Kappas, A., and R. H. Palmer. 1963. Selected aspectsof steroid pharmacology. Pharmacol. Rev. 15: 123.
4. Palmer, R. H., B. Ratkovits, and A. Kappas. 1961.Steroid pyrogen studies in laboratory and domesticanimals. J. Appl. Physiol. 16: 345.
5. Kappas, A., W. Soybel, P. Glickman, and D. K.Fukushima. 1960. Fever-producing steroids of endoge-nous origin in man. Arch. Internal. Med. 105: 701.
6. Glickman, P. B., R. H. Palmer, and A. Kappas. 1964.Steroid fever and inflammation. Arch. Internal Med.114: 46.
7. Atkins, E., and E. S. Snell. 1965. Fever. In TheInflammatory Process. B. W. Zweifach, L. Grant, andR. T. McCluskey, editors. Academic Press Inc.,N. Y. 495.
8. Bondy, P. K., G. L. Cohn, and C. Castiglione. 1960.Etiocholanolone fever; A clinical entity. Trains. Assoc.Am. Physicians. 73: 186.
9. Bondy, P. K., G. L. Cohn, and P. B. Gregory. 1965.Etiocholanolone fever. Medicine. 44: 249.
10. Bodel, P., and E. Atkins. 1966. Human leukocytepyrogen producing fever in rabbits. Proc. Soc. Exptl.Biol. Med. 121: 943.
11. Atkins, E., and C. Heijn, Jr. 1965. Studies on tuber-culin fever. III. Mechanisms involved in the release ofendogenous pyrogen in vitro. J. Exptl. Med. 122: 207.
12. Fallon, H. J., E. Frei, III, J. D. Davidson, J. S.Trier, and D. Burk. 1962. Leucocyte preparations
from human blood: Evaluation of their morphologicand metabolic state. J. Lab. Clin. Med. 59: 779.
13. Bennett, W. E., and Z. A. Cohn. 1966. The isolationand selected properties of blood monocytes. J. ExptlMed. 123: 145.
14. Lakshmanan, T. K., and S. Lieberman. 1954. An im-proved method of gradient elution chromatographyand its application to the separation of urinary keto-steroids. Arch. Biochelzm. Biophys. 53: 258.
15. Edwards, R. W. H. 1958. Steroids. In Chromato-graphic and Electrophoretic Techniques. I. Smith,editor. William Heineman, London. 1: 428.
16. Wilzbach, K. E. 1957. Tritium-labeling by exposureof organic compounds to tritium gas. J. Am. Chent.Soc. 79: 1013.
17. Atkins, E., P. Bodel, and L. Francis. 1967. Release ofan endogenous pyrogen iii vitro from rabbit mono-nuclear cells. J. Exptl. Med. 126: 357.
18. Myrvik, Q. N., E. S. Leake, and B. Fariss. 1961.Studies onl pulmonary alveolar macrophages from thenormal rabbit: A technique to procure them in a highstate of purity. J. Jumlmntzol. 86: 128.
19. Bo(lel, P. T., and E. Atkins. 1965. Studies in stap)lhy-lococcal fever. V. Staphylococcal filtrate pyrogen.Yale J. Hiol. AMcd. 38: 282.
20. Bennett, 1. L., Jr., and P. B. Beeson. 1953. Studies onpathogenesis of fever. II. Characterization of fever-producing substances from p)olymorplhonuclear leuko-cytes and from fluid of sterile exudates. J. Exptl.Mcd. 98: 493.
21. Rafter, G. W., R. D. Collins, and W. B. Wood. 1960.Studies on the pathogenesis of fever. VII. Preliminarychemical characterization of leucocytic pyrogen. J.Exptl. Med. 111: 831.
22. 1952. Spinco Technical Manual, Ultracentrifuge ModelL. Specialized Instruments Corp., Belmont, Calif.
23. Bodel, P., and E. Atkins. 1967. Release of endogenouspyrogen by human monocytes. Nezv Engl. J. Med. 276:1002.
24. Cohn, G. L., P. K. Bondy, and C. Castiglione. 1961.Studies on pyrogenic steroids. I. Separation, identifica-tion and measurement of unconj ugated dehydroepi-androsterone, etiocholanolone and androsterone in hu-man plasma. J. Clin. Invest. 40: 400.
25. Kappas, A., L. Hellman, D. K. Fukushima, and T. F.Gallagher. 1958. The thermogenic effect and metabolicfate of etiocholanolone in man. J. Clin. Endocrinol.Metab. 18: 1043.
26. Bondy, P. K., G. L. Cohn, and P. B. Gregory. 1965.Etiocholanolone fever. Medicine. 44: 249.
27. Hellman, L. Steroids. In Clinical Endocrinology.E. B. Astwood, editor. Grune and Stratton, N. Y. Inpress.
28. Levere, R., A. Kappas, and S. Granick. 1967. Theinduction of hemoglobin formation in erythroid cellculture by 5p-H steroid metabolites. J. Clin. Invest.46: 1084.
1166 P. Bodel and M. Dillard

29. Petersdorf, R. G., and I. L. Bennett, Jr. 1957. Studieson the pathogenesis of fever. VII. Comparative ob-servations on the production of fever by inflammatoryexudates ill ral)bits andl (logs. Bull. Johins HopkinslIosp. 100: 277.
30. Snell, E. S., and E. Alkins. l967. Interactions ofgram-negative bacterial endotoxin with rabbit bloodin vitro. Am. J. Physiol. 212: 1103.
31. Shulman, J. A., W. L. Herrmann, and R. G. Peters-
dorf. 1964. Experimental etiocholanolone fever. J.Clin. Endocrinol. Metab. 24: 1136.
32. Hahn, H. H., D. C. Char, W. B. Postel, and W. B.Wood(, Jr. 1967. Stu(dies on pathogenesis of fever.XV. Production (if endogeious pyrogen by peritonealniacrop)hages. J. lIxpll. Mcd. 126: 385.
33. Weissmann, G. 1965. Studies of lysosomes. VI. Theeffect of neutral steroids and bile acids on lysosomesin vitro. Biochem. Pharmacol. 14: 525.
Production of Leukocyte Pyrogen In Vitro by Etiocholanolone 117