studies of tissue permeability · across the membrane. ... permeability had been increased by...
TRANSCRIPT

THE JOURNAL OF HIOL.O~ICAL CHEMISTRY Vol. 240, No. 9, September 1965
Printed in U.S.A.
Studies of Tissue Permeability
X. CHANGES IN PERMEABILITY TO 3-METHYLGLUCOSE ASSOCIATED WITH CONTRACTION OF ISOLATED FROG MUSCLE*
JOHN 0. HoLLoszYt AND H. T. NARAHARA
Prom the Department of Biological Chemistry, Washington University School of Medicine, St. Louis, Missouri 63110
(Received for publication, March 26, 1965)
Exercise can lower the concentration of blood glucose in man (1) and laboratory animals (2). Goldstein et al. (3) showed that exercise lowered the concentration of injected n-galactose, n-xylose, and L-arabinose, but not of n-sorbitol, in the blood of eviscerated nephrectomized rats. Helmreich and Cori (4) studied the phenomenon more directly by measuring the con- centration of pentoses in plasma and muscles of nephrectomized rats and found that the accumulation of pentoses within muscle cells was increased by exercise. Their results have been con- firmed with pentoses in cats (5) and with galactoseJ4C in rats (6). Similar changes in the concentration of sugars have been seen in the blood and tissues of animals with experimental diabetes during exercise (2-4).
These findings indicate that exercise increases the uptake of sugars by muscle, but it is not clear that this effect is entirely the result of changes in cell permeability because alterations in blood flow during exercise might also influence the rate of ac- cumulation. In order to extend these studies with detailed measurements of permeability under more readily controlled conditions, it was of interest to examine the effect of muscle contraction in vitro.
Frog sartorius muscle has been shown to be well suited for use in the investigation of sugar transport across the cell mem- brane (7). Frog sartorii are also thin enough to permit adequate oxygenation at moderate frequencies of stimulation (8). Fur- thermore, if desired, stimulation can be carried out under strictly anaerobic conditions.
The substrate selected for the present studies was a-methyl- glucose, a nonmetabolizable sugar that closely resembles glucose in the characteristics of its transport across the cell membrane (7, 9, 10). In the presentation that follows, permeability will be expressed in terms of the initial rate of penetration of 3- methylglucose into cells. It was found that stimulation of isolated frog skeletal muscle causes a large increase in the permea- bility to 3-methylglucose. The effects of varying the tempera- ture of incubation or the frequency and duration of stimulation were investigated. The results suggest that electrical stimula- tion initiates a chain of events that continues to affect permea- bility even after the end of contraction. The penetration of
* This work was supported in part by Research Grant AM- 04082.05 from the National Institutes of Health, United States Public Health Service, and by a grant of the American Cancer Society to Washington University, St. Louis, Missouri.
t Special Research Fellow of the National Institute of Arth- ritis and Metabolic Diseases, United States Public Health Service.
sugar into stimulated muscle exhibits a saturation type of kinetics, and the increase in permeability is related more to a change in V,,, than of K,. Evidence is presented that insulin and contraction affect the same system for transport of sugar across the membrane. Electrical stimulation was able to pro- duce its effect on permeability in the absence of added insulin; the tissues probably contained little endogenous insulin because the permeability of resting muscles was very low.
EXPERIMENTAL PROCEDURE
Chemicals-3-O-Methyl-n-glucose-3H had a specific activity of approximately 20 & per pmole. The source and purity of the compound and the method of measuring radioactivity have been described in the preceding paper of this series (10). Man- nitol-1,6-W containing approximately 2 PC per pmole was obtained from the Volk Radiochemical Company.
Handling of Tisms-Female Rana pipiens purchased from a dealer in Minnesota were kept in water at approximately 4”. Sartorius muscles, weighing approximately 80 to 140 mg, were dissected out without severing the attachment to the pubic bone. Muscles from summer frogs are less permeable to sugar than those from winter frogs, but muscles were found to be responsive to stimulation in all seasons. Muscles for direct stimulation were kept overnight in oxygenated Ringer’s solution at 4” prior to use in order to permit them to recover from the effects of dissection. Nerve-muscle preparations for indirect stimulation were left in Ringer’s solution at room temperature for 90 min and then at 0” for 15 to 120 min before use. The Ringer’s solution was similar to that used previously (7) except that the concentration of potassium was lowered from 5.7 mM to 2.5 mM to make the composition more like that of the elec- trolytes in frog plasma (11).
Muscles that were to be stimulated directly were attached to a vertical Lucite rod by clamping the pubic bone against a platinum wire electrode that lay in a slightly raised area near the lower end of the rod. A second platinum wire loop 3.5 cm further up on the rod held the muscle away from the rod for optimal exposure to the medium and provided the second point of electrical con- tact. In some instances the muscle was stimulated indirectly through platinum electrodes applied to the motor nerve in a moist chamber that was located just above the surface of the fluid bathing the muscle. A stimulator from the L. N. Simpson Company of St. Louis was used to deliver rectangular, direct current impulses lasting 4 milliseconds at a voltage that was slightly greater than that required for maximal contraction
3493
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Studies of Tissue Permeability. X Vol. 240, No. 9
(approximately 20 volts for direct stimulation and 0.6 volt for stimulation through the nerve).
The tibia1 tendon of each muscle was tied to a jeweler’s chain that connected it with a transducer. In most of the experiments the resting tension of the muscle was adjusted to 1 g, and iso- metric contractions were elicited. The tension developed in isometric contractions was measured with a Sanborn force trans- ducer, model FTA 100-1, capable of detecting changes in the range of approximately 0.5 to 100 g. For experiments involving isotonic contraction the muscle was extended with a net load of 1 g, and an after loading stop device was adjusted to prevent further stretching of the muscle. The net load to be lifted was then increased to 10 g. Shortening was measured with a Sanborn Linearsyn transducer, model 585DT500-BM, capable of detecting displacements in the range of 0.1 to 25 mm. In order to decrease the inertia of the measuring system at lopr work loads, in some experiments the displacement transducer was not used, and the muscle lifted a 0.3-g weight by a silk thread that passed over a small pulley; movement of the weight was measured on a milli- meter scale.
The mounted muscle was immersed in 7 ml of Ringer’s solution in a test tube, and the tube was held in a constant temperature bath at 19” or 0”. In most cases stimulation was performed aerobically, and a gas mixture composed of 95% O2 and 5% COz was bubbled continuously through the medium. For stimulation under anaerobic conditions, a gas phase of 95% Nz and 5% COz was used; traces of oxygen were first removed from the gas by passing it through solutions of vanadyl sulfate (12), and the top of the test tube was closed with Parafihn except for a small opening around the chain leading to the transducer. Oxygen that might have been introduced into the medium with the muscle was flushed out for a period of 10 min before beginning stimulation.
In previous studies on the effects of insulin (10) it has been shown that initial rates of entry of 3-methylglucose into the cell can be measured at 19” (a) if the extracellular space of the iso- lated muscle is first equilibrated with the sugar at 0” and (5)
3 SIIOCKSIMIN. .
; I
OO I
60 120 180 DURATION OF STIMULATION,MINUTES
FIG. 1. Effect of frequency and durat,ion of stimulation on the permeability of muscle to 3-methylglucose. Muscles of winter frogs were stimulated at 19” for different periods of time. Values for U, the initial rate of penetration of 3-methylglucose measured immediately after stimulation, are expressed as micromoles of substrate penetrating per ml of intracellular water per hour. Each point represents the mean for two to four muscles.
provided that the intracellular accumulation does not exceed a level that is approximately 16% of the extracellular concentra- tion. This restriction of the extent of penetration permissible would limit the variety of experimental conditions that could be examined if 3-methylglucose transport were measured during the period of stimulation. However, the increase in permeability to 3-methylglucose that occurs as a result of stimulation at moderate frequencies persists essentially unchanged for long intervals after stopping stimulation. It was also found that 3-methylglucose entered cells very slowly at 0” even when the permeability had been increased by stimulation. Therefore, in the experiments to be reported here, initial rates of sugar penetration were measured following stimulation.
Each milliliter of the Ringer’s solution used for incubation contained 3.5 PC of 3-methylglucose-3H and 0.25 PC of man- nitol-i4C. The concentration of 3-methylglucose was adjusted to 8 mM with nonlabeled carrier in all experiments except for the study of the apparent K, of penetration, where various concentrations were used. Mannitol-i4C was used to determine the extracellular space (cj. Reference 10). Sufficient nonlabeled mannitol was added to bring the combined concentrations of 3-methylglucose and mannitol to 40 mM. In order to avoid changes in osmolarity during successive phases of the experi- ments, the Ringer’s solution used for stimulation or other types of prior incubation contained nonlabeled mannitol at a concen- tration of 40 mM; for convenience, these nonradioactive media have been referred to simply as Ringer’s solution.
After stimulation, the muscle was rapidly removed from the electrode, trimmed, and chilled in fresh ice-cold Ringer’s solution for 5 min. The muscle was then blotted on filter paper damp- ened with ice-cold Ringer’s solution and equilibrated for 15 min at 0” with 2 ml of medium containing 3-methylglucose-3H and mannitol-W in a stoppered 25.ml Erlenmeyer flask. The flask and muscle were finally incubated at 19”, generally for 10 to 20 min, in order to permit 3-methylglucose to accumulate within the cells. When the flasks were transferred from a 0” to a 19” bath, the temperature of the incubation medium rose to the higher value within 14 min. Preliminary studies indicated that mounting of the muscles on the electrode, and the presence or absence of the pubic bone, had no significant effect on permea- bility.
Chemical Analyses-Lactic acid was determined with lactate dehydrogenase and hydrazine (13) in perchloric acid extracts of muscles and media, and the creatine phosphate content of muscles was measured calorimetrically (14).
RESULTS
E$ect of Duration and Frequency of Stimulation on Permeability to S-Methylglucose-The permeability of isolated frog sartorius muscle to 3-methylglucose increased linearly with time in re- sponse to electrical stimulation (Fig. 1). No delay in the onset of this increase in permeability could be demonstrated. Even- tually, a plateau was formed that was higher and was reached earlier at the faster frequencies of stimulation. For frequencies in the range of 2.5 to 20 shocks per min the plateau, or greatest change in permeability attainable, was proportional to the rate of stimulation (Fig. 2).
E$ect of Stimulation at @‘-Contractions of frog muscle are readily elicited at 0”. Although permeability increased more slowly when stimulation was carried out at 0” than at 19”, even brief periods of stimulation at 0” had a significant effect (Table 1
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from

September 1965 J. 0. Holloszy and H. T. Narahara 3495
I -r/ I
A T
of I I I I 0 5
SHOCKS !EFl MINUTE 15 20
FIG. 2. Relationship of permeability to the frequency of stim- ulation. Muscles of winter frogs were stimulated at 19” at vari- ous frequencies, and the plateau of permeability attained after 2 hours was measured as in Fig. 1. Each point indicates the mean for four muscles, and vertical bars represent twice the standard error of the mean.
Experiments B, C, and D). When stimulation at 0” was con- tinued for sufficiently long periods of time, the effects on permea- bility were as great as those obtained with stimulation at 19’ (Table I, Experiments E and F). In these studies permeability was measured by incubating muscles with 3.methylglucose at 19” after st.imulation at either 0” or 19”.
It was conceivable that stimulation at 0” initiated changes in the muscle which then led to an alteration of permeability only after the muscle was warmed to 19” for the incubation with 3-methylglucose. Therefore, in other experiments incubation with 3-methylglucose was done at 0” in order to see whether or not permeability would increase when the muscles were not warmed either during or after stimulation. Two effects of temperature must be distinguished here. On the one hand, the rate of transport of Zkmethylglucose is faster at 19” than at 0” in resting muscles and in muscles that have been stimulated for 2 hours at 19” (Table I, Experiments F and G). On the other hand, the matter that is of principal concern here is the possibility that an increase of temperature may be necessary in order to allow a series of reactions initiated by electrical stimulation at 0” to progress to completion so that they can affect permeability. The results of Experiment G in Table I clearly indicate that permeability was increased above the corresponding basal value when the muscle was stimulat,ed at 0” and the measurement of permeability was also carried out at 0”. Thus, it was not neces- sary to warm the muscle above 0” at any time in order to obtain an effect of stimulation on permeability.
Effect Following Brief, Intense Stimulation-When short periods of intense stimulation were employed, permeability continued to increase after the end of stimulation. For example, when muscles were given 10 tetanic stimuli lasting 5 set each and spaced 1 min apart, not only did permeability increase during stimulation but a considerable rise also occurred in a period of 30 min following stimulation (Table II, Experiment B). When 10 tetanic stimuli were spaced 6 min apart so that the total elapsed time was 54 min, a further increase in permeability did not occur (Table II, Experiment C).
It was conceivable that a series of closely spaced tetanic shocks, in addition to augmenting permeability, might cause some other type of alteration in the cell that could interfere
transiently with sugar transport. I f this inhibitory effect of rapid stimulation were to subside in the period following stimula- tion, an apparent increase in permeability would result. In order to try to demonstrate an inhibitory effect of this nature, muscles were first subjected to a course of stimulation, 120 shocks per min for 30 min, that had been found in prior trials to produce a maximal effect on permeability. Muscles pre- treated in this manner should show no further increase in permea- bility dvring subsequent tetanization whereas any interference with sugar transport should still be demonstrable. After an initial course of maximal stimulation half of the muscles were allowed to rest for 20 min in Ringer’s solution in order to permit the permeability to become stabilized and were then subjected to 10 tetanic stimuli spaced 1 min apart. Paired control muscles were not stimulated tetanically but were allowed to rest for 30 min after the initial maximal stimulation before measuring permeability. The rate of penetration of 3-methylglucose in the tetanized muscles was 9.9 Imoles per ml of cell water per hour, which was a value not significantly different from the mean of 10.1 observed in the controls. Thus, no evidence of a tran- sient interference with sugar transport was found in rapidly tetanized muscles to account for the increase in permeability that was observed after the end of stimulation.
In muscles that were subjected to a series of closely spaced
TABLE I Effect of prior stimulation at 0” and 19’ on permeability
to S-methylglucose Muscles were stimulated to contract at either 0” or 19”; they
were then equilibrated with 3-methylglucose at, 0” as described under “Experimental Procedure,” and the initial rate of pene- tration, D, was measured at either 19” or 0”. Eight muscles were used in Experiment A, and three muscles for each group in Ex- periment E; four muscles were used for each group in the other experiments. The mean and standard error of the mean is given for each group.
%n”d” experiment
Summer A B
C
D
E
Winter F
G
- T a m
m
19” 19
19
19
19
19
0
Fre- mlcy 01 stimu- lation
ihocks/ n&in
None TW TS
60 60 20 20 16 16
None 20 20
None 20 20
Du- 63n 0 ,timu- lation
min
15 15 30 30
180 180
120 120
120 120
T< f ;
7
-
19” 0
19 0
19 0
19 0
19 0
19 0
0.72 f 0.14 4.3 f 0.7 1.8 f 0.3 4.2 f 0.4 2.4 f 0.5 3.1 i 0.4 2.2 f 0.1 3.9 f 1.1 4.6 f 1.2
0.78 f 0.09 6.6 f 0.6 6.1 f 0.2
0.051 f 0.004 1.01 f 0.15 0.85 f 0.17
<O.ool
<O.Ol
<O.Ol
NSDt
NSD
NSD
* TS, 10 tetanic stimuli lasting 5 set each were spaced 1 min apart.
v
t NSD, no significant difference between the means.
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3496 Studies of Tissue Permeability. X Vol. 240, No. 9
TABLE II Increase in permeability during rest period following brief, intense stimulation
Winter frogs were used. Paired muscles suspended in Ringer’s solution were given 10 tetanic stimuli lasting 5 set each.
Ringer’s solution for 30 min at 19” before its permeability was Perme- measured.
ability was measured immediately after stimulation in one muscle The extracellular space is expressed as milliliters per
g of tissue, wet weight. of each pair, and the other muscle was allowed to rest in fresh
Nine muscles per group were used in Ex- periments A and B, and six muscles per group in Experiment C.
I , 1 , Interval Period
Experiment yt;tu~ after II stimulation
Muscle weight P Extracellular space
~~ min min s vdes/nd/hr wwleslmusclejhr ml/g
A Not stimulated 0.094 f 0.607 0.42 f 0.08 0.025 zt 0.005 0.27 i 0.01 B 1 0 0.099 f 0.004 3.9 i 0.2 0.23 i 0.01 0.22 f 0.01
1 30 0.095 f 0.004 7.2 i 0.5 0.37 z!z 0.03 <O.OOl 0.26 f 0.01 C 6 0 0.112 f 0.003 7.7 f 1.0 0.46 i 0.05 0.27 f 0.01
6 30 0.108 f 0.605 7.9 f 1.1 0.45 i 0.06 NSD* 0.28 f 0.01
* NSD, no significant difference between the means.
TABLE III Changes in permeability and in distribution of tissue water during period following tetanic stimulation
P
<O.Ol
NSD
Muscles from winter frogs were given 10 tetanic stimuli lasting in hypotonic Ringer’s solution at 19”, and then the rate of pene- 5 set each and spaced 1 min apart. The permeability of one tration of 3-methylglucose was measured in a hypotonic medium. muscle of each pair was measured immediately after stimulation, Hypotonicity was produced by decreasing the concentration of in media of the usual tonicity, as described under “Experimental mannitol by 30 pmoles per ml, and the concentration of NaCl by Procedure.” The other muscles were allowed to rest for 30 min 10 pmoles per ml. There were five muscles in each group.
Period after stimulation Medium Extracellular space
I Muscle weight
I I I
min
0 30
Normal Hypotonic
ml/s s ,mwlcslml/hr ,mwlcs/musclc/hr
0.22 f 0.01 0.117 i 0.007 3.6 f 0.4 0.25 f 0.03 0.19 f 0.01 0.128 f 0.010 6.9 f 0.6 0.54 f 0.05
P
<O.Ol
Manic stimuli, a small but significant reduction in extracellular space occurred (Table II, Experiment B). A corresponding volume of water must have moved into the cell since the muscle weight did not change significantly. In the 30-min period that followed stimulation, the extracellular space increased again to approximately the resting value (Table II, Experiment B); presumably this change reflects an efflux of water from the cell. Although the extracellular space differs somewhat from one batch of frogs to another, this sequence of events has been seen consistently in other similar experiments not reported here. These shifts in tissue water probably occurred in response to alterations of osmolarity resulting from a transient accumulation of lactic acid and other glycolytic intermediates during stimula- tion. On account of these variations in cell water content, the penetration of sugar has been expressed as micromoles per muscle as well as micromoles per ml of cell water per hour (Table II). The interpretation of the results is not affected qualitatively by the manner of expression.
Since the increase in permeability that occurred in the period following a series of closely spaced tetanic shocks was accom- panied by an efflux of water from the cell, the possibility was considered that the increase in permeability might be related to changes in cell water content. However, when the efflux of water was prevented by transferring the muscle to a hypotonic medium immediately after stimulation, an increase in permea- bility still occurred (Table III). Thus, the change in permea- bility could be clearly dissociated from the usual decrease in cell water cont,ent.
Reversal of E$ect of Stimulation on Permeability-Isolated frog
sartorius muscles were stimulated in Ringer’s solution at 19” and then allowed to recover in fresh Ringer’s solution at the same temperature for various periods of time before measuring the permeability. When muscles were stimulated at a frequency of 60 shocks per min for 30 min, the permeability 5 hours after stimulation was just as high as it was 1 hour after stimulation (Fig. 3). Since this frequency of stimulation was close to the maximum one that could be used without causing marked fatigue, it was of interest to determine how long permeability would remain elevated after stimulation at a lower frequency. It was found that the increase in permeability resulting from stimulation at 26 shocks per min also persisted unchanged for 5 hours (Fig. 3). Then a moderately rapid decline took place, followed by a more gradual decrease; approximately 24 hours after stimulation, permeability was almost at the initial control value again. Other experiments of this type on different batches of frogs yielded similar results. An increased permeability of rat gastrocnemius muscle to pentoses was found by Helmreich and Cori (4) to persist for at least 30 min after stimulation of the muscles in situ.
The experiments on frog muscle described above were carried out in the absence of any utilizable substrate in the medium. However, when glucose was added to the incubation medium during the recovery period, the fall in permeability was only slightly accelerated (Fig. 3).
When stimulation through the motor nerve was substituted for direct electrical stimulation of muscle, a similar delay in the return of permeability to the resting level was observed. Thus, 1 hour after stimulation through the nerve at a frequency of 26
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from

September 1965 J. 0. Holloszy and H. T. Narahara 3497
shocks per min for 30 min, the average rate of penetration of 3-methylglucose for eight muscles was 4.2 f 0.3 pmoles per ml of cell water per hour; 5 hours after stimulation, the average rate in paired muscles was 4.0 f 0.6. These observations indi- cate that the prolonged delay in reversal of the effect of stimula- tion on permeability represents a normal physiological process rather than an effect of electrical injury of the muscle cell mem- brane.
The process that restores permeability to the basal state is markedly influenced by temperature, and very little reversal of the increase in permeability was seen at 4“ for approximately 24 hours after stimulation (Fig. 3).
Kinetics of Penetration of S-Methylglucose after Stimulation-A saturation type of kinetics was observed when the rate of penetra- tion of 3-methylglucose was measured in electrically stimulated muscles at different concentrations of substrate. The results obtained with unstimulated and stimulated muscles have been graphed in Fig. 4 according to the method of Lineweaver and Burk (15). In unstimulated muscles the I’,,, of penetration of 3-methylglucose was 1.5 Mumoles per ml of intracellular water per hour, and stimulation at 20 shocks per min increased the value to 9.0. The apparent K,,, of 4.2 mM in stimulated muscles was not significantly different from the value of 4.0 in resting muscles and is in the range of values that has been observed previously in resting frog muscles (10).
E&d of Insulin and Stimula.tion Together on Permeability- I f insulin and electrical stimulation could act independently on two separate sets of transport sites, one might expect their effects on permeability to be additive even when maximal amounts of the two agents were applied. It was of interest to test this possibility. First, it was established that stimulation at a frequency of 120 shocks per min for 30 min produced a nearly maximal increase in permeability, and that stimulation at this frequency for 60 min caused no further increase. In the same
v IO
9
8
1
6
5
4
3
2
I
a
26 SHOCKS/MM .
0 -
If, WlTtloUl GLUCOSE .
UNSTlMULATED IO: WITH GLUCOSE -_-- - -- - - - - ____________________-------------------- *
5 PERIOD AFT;: STIM"LAT&HOURS
20
FIG. 3. Reversal of the increase in permeability following stim- ulation. Muscles from winter frogs were stimulated at 60 or 26 shocks per min for 30 min at 19” and were then allowed to recover in fresh Ringer’s solution at 19” or 4” before measuring the initial rate of penetration of 3-methylglucose. In some experiments glucose at a concentration of 2 mM was included in the Ringer’s solution of one muscle of each pair during the recovery phase, but glucose was not present during the final incubation with 3- methylglucose. Each point is the mean for two to four muscles.
1.5
‘4
1.0
STIMULATEG . -.---*
0125 02% I/S
FIG. 4. Effect of stimulation on the initial rate of penetration of 3-methylglucose at different concentrations. Muscles from winter frogs were stimulated at a frequency of 20 shocks per min for 2 hours at 19”. The initial rate of penetration, V, was then measured at different concentrations (S) of 3-methylglucose. Values of 21 are expressed as micromoles per ml of intracellular water per hour, and S is given in micromoles per ml. Each point is the mean for four muscles. Values obtained in unstimulated muscles are shown for comparison.
TABLE IV Effect of insulin and stimulation together on permeability
to S-methylglucose Muscles from winter frogs were used. One group of muscles
was incubated for 3 hours at 19” in Ringer’s solution with or without insulin and then stimulated while immersed in the same solution for another 30 min. Other muscles were incubated with or without insulin for 33 hours but were not stimulated. Initial rates of penetration of 3-methylglucose were then measured. There were 4 muscles in each group except in Experiment A where 12 muscles were used.
Experiment Insulin
0 m t3Doo
0 2ooo
70 0
70
Stimulation
shocks/min ,moles/ml/kr
0 0.42 f 0.08 0 6.9 f 0.6 0 7.1 f 0.5
120 7.5 f 0.4 120 8.3 zk 0.6
0 2.4 f 0.4 20 3.0 f 0.4 20 4.9 f 0.4
-
v
6.5 6.7 7.1 7.9 2.0 2.6 4.5
batch of frogs a nearly maximal effect of insulin1 on permeability was obtained by exposing the muscles to the hormone for 3f hours at a concentration of 2000 microunits per ml (Table IV, Experiment B). Approximately 3 hours are required for insulin to exert its full effect on permeability in frog muscle at 19” (10). The combined effect of maximal amounts of insulin and electrical stimulation was not significantly different from the effect of stimulation alone in paired muscles (Table IV, Experiment B). As one might have anticipated, the effects of submaximal amounts of insulin and stimulation on permeability were ap-
1 The authors wish to thank Dr. Otto K. Behrens of Eli Lilly and Company for a generous gift of crystalline beef insulin.
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from
3498 Studies of Tissue Permeability. X Vol. 240, No. 9
proximately additive (Table IV, Experiment C). Sacks and Smith (5) observed an approximately additive effect of insulin and muscular activity on the accumulation of pentose in cat gastrocnemius muscle in wivo, but it is not stated whether or not the amounts of insulin and exercise used were maximal.
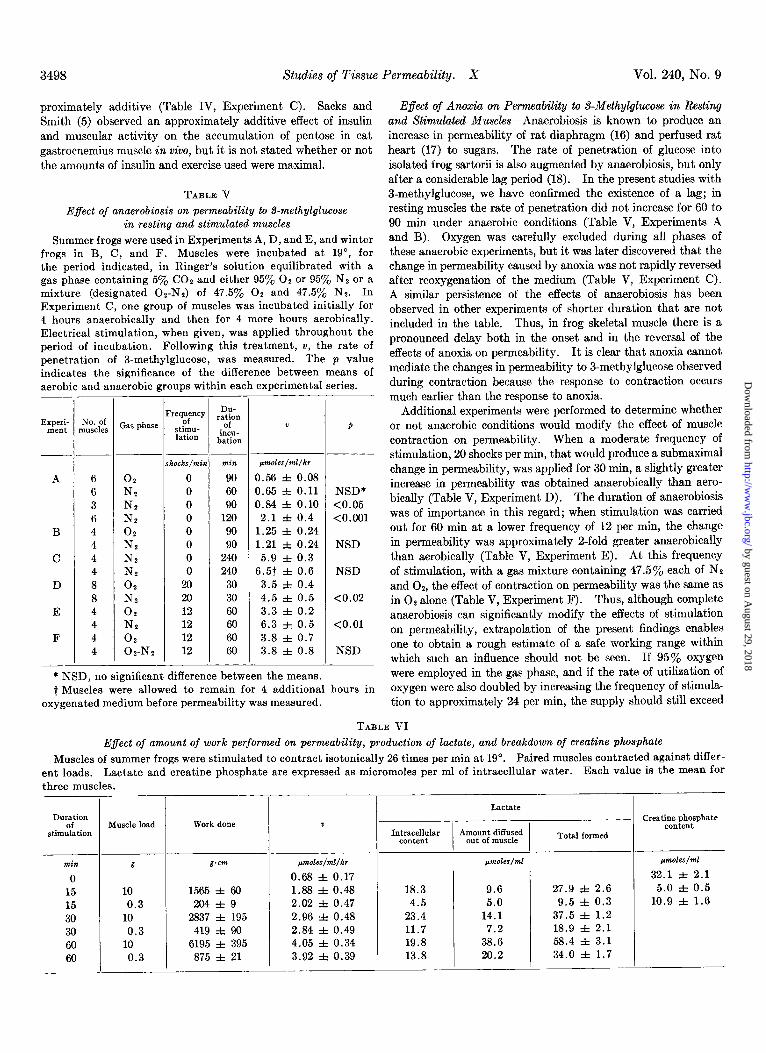
TABLE V
Effect of anaerobiosis on permeability to S-methylglucose in resting and stimulated muscles
Summer frogs were used in Experiments A, D, and E, and winter frogs in B, C, and F. Muscles were incubated at 19”, for the period indicated, in Ringer’s solution equilibrated with a gas phase containing 5% CO2 and either 95% 02 or 95% N, or a mixture (designated Oz-Ne) of 47.5y0 02 and 47.5% Nz. In Experiment C, one group of muscles was incubated initially for 4 hours anaerobically and then for 4 more hours aerobically. Electrical stimulation, when given, was applied throughout the period of incubation. Following this treatment, v, the rate of penetration of I-methylglucose, was measured. The p value indicates the significance of the difference between means of aerobic and anaerobic groups within each experimental series.
-
1
_-
-
Experi- ment Gas phase req:?y
stimu- lation
DU- ration
of incu-
bation
t P
rocks/& pmoles/ml/hr
0 0.56 f 0.08
0 0.65 f 0.11 NSD* 0 0.84 f 0.10 <0.05 0 2.1 f 0.4 <O.OOl 0 1.25 f 0.24 0 1.21 f 0.24 NSD 0 5.9 f 0.3 0 6.57 f 0.6 NSD
20 3.5 f 0.4 20 4.5 f 0.5 <0.02 12 3.3 f 0.2 12 6.3 f 0.5 <O.Ol 12 3.8 f 0.7 12 3.8 f 0.8 NSD
* NSD, no significant difference between the means. t Muscles were allowed to remain for 4 additional hours in
oxygenated medium before permeability was measured.
6 6 3 6 4 4 4 4 8 8 4 4 4 4
02 N? N2 N* 02
N2 N2 N-2 02 N2 02 N2 02 02-N2
min
90
60 90
120 90 90
240 240
30 30 60 60 60 60
-
-
E$ect of Anoxia on Permeability to S-Methylglucose in Resting and Stimulated Muscles-Anaerobiosis is known to produce an increase in permeability of rat diaphragm (16) and perfused rat heart (17) to sugars. The rate of penetration of glucose into isolated frog sartorii is also augmented by anaerobiosis, but only after a considerable lag period (18). In the present studies with 3-methylglucose, we have confirmed the existence of a lag; in resting muscles the rate of penetration did not increase for 60 to 90 min under anaerobic conditions (Table V, Experiments A and B). Oxygen was carefully excluded during all phases of these anaerobic experiments, but it was later discovered that the change in permeability caused by anoxia was not rapidly reversed after reoxygenation of the medium (Table V, Experiment C). A similar persistence of the effects of anaerobiosis has been observed in other experiments of shorter duration that are not included in the table. Thus, in frog skeletal muscle there is a pronounced delay both in the onset and in the reversal of the effects of anoxia on permeability. It is clear that anoxia cannot mediate the changes in permeability to 3-methylglucose observed during contraction because the response to contraction occurs much earlier than the response to anoxia.
Additional experiments were performed to determine whether or not anaerobic conditions would modify the effect of muscle contraction on permeability. When a moderate frequency of stimulation, 20 shocks per min, that would produce a submaximal change in permeability, was applied for 30 min, a slightly greater increase in permeability was obtained anaerobically than aero- bically (Table V, Experiment D). The duration of anaerobiosis was of importance in this regard; when stimulation was carried out for 60 min at a lower frequency of 12 per min, the change in permeability was approximately a-fold greater anaerobically than aerobically (Table V, Experiment E). At this frequency of stimulation, with a gas mixture containing 47.5% each of NP and 02, the effect of contraction on permeability was the same as in O2 alone (Table V, Experiment F). Thus, although complete anaerobiosis can significantly modify the effects of stimulation on permeability, extrapolation of the present findings enables one to obtain a rough estimate of a safe working range within which such an influence should not be seen. If 95% oxygen were employed in the gas phase, and if the rate of utilization of oxygen were also doubled by increasing the frequency of stimula- tion to approximately 24 per min, the supply should still exceed
TABLE VI
Effect of amount of work performed on permeability, production of lactate, and breakdown of creatine phosphate
Muscles of summer frogs were stimulated to contract isotonically 26 times per min at 19”. Paired muscles contracted against differ- ent loads. Lactate and creatine phosphate are expressed as micromoles per ml of intracellular water. Each value is the mean for three muscles.
D”6a:io stimulation
Muscle load Work done I I
min
0 15 15 30 30 60 60
g
10 0.3
10 0.3
10 0.3
g.cm
1565 f 60 204 f 9
2837 f 195 419 f 90
6195 f 395 875 f 21
pmoles/ml/hr
0.68 f 0.17 1.88 f 0.48 2.02 f 0.47 2.96 f 0.48 2.84 f 0.49 4.05 f 0.34 3.92 f 0.39
-
.-
-
- Lactate
Intracellular Amount diffused content out of muscle Total formed
18.3 9.6 4.5 5.0
23.4 14.1 11.7 7.2 19.8 38.6 13.8 20.2
27.9 f 2.6 9.5 f 0.3
37.5 f 1.2 18.9 f 2.1 58.4 f 3.1 34.0 f 1.7
- creatimp~;phate
pVdCS/?lil
32.1 f 2.1 5.0 f 0.5
10.9 f 1.6
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from

September 1965 J. 0. Holloszy and H. T. Narahara 3499
the requirement of the tissue, and changes in permeability caused
by the stimulation would not be modified because of a lack of oxygen. These results are in accord with the conclusions of Hill and Kupalov (8) that a thin sartorius muscle can contract for long periods in titro at frequencies up to 23 to 37 per min in a fully oxygenated medium at 20” without exhaust,ing the supply of oxygen within the tissue.
Effect of Amount of Work Performed on Permeability, Produc- tion of Lactate, and Brealcdown of Creatine Phosphate-Muscles were made to contract isotonically against either a 0.3-g or a 10-g load for various periods of time aerobically. Although the muscles contracting against the 10-g load performed approxi- mately 7 times more work than the more lightly loaded muscles, the same change in permeability occurred in both groups (Table VI). Under these conditions, muscles that lifted 10 g produced significantly larger amounts of lactic acid (p < 0.01) and broke down more creatine phosphate (p < 0.01) than the lightly loaded paired muscles (Table VI), in keeping with earlier reports of the effects of muscle loading on glycolytic reactions (19, 20) and creatine phosphate metabolism (21, 22).
Thus, it is clear that changes in the amount of work performed during isotonic contraction at a given frequency do not modify the effect of stimulation on permeability. Furthermore, the change in permeability is not correlated with the rate of forma- tion of lactate or with the intracellular levels of lactate and creatine phosphate.
nISCUSSION
Stimulation of isolated frog sartorius muscles has been shown to facilitate entry of 3-methylglucose into the cells. Indirect stimulation through the nerve is as effective as direct stimulation, and the changes in permeability are not mediated by inadequate oxygenation of the tissue. Since circulatory changes do not play a role in these experiments on isolated tissues, the results support the concept that the fall in concentration of blood sugar (2, 3) and the enhanced accumulation of sugars in muscle (4) that have been observed in z&o after exercise are largely attribut- able to an increase in permeability of the muscle cells.
The finding (Table II) that a period of brief, intense stimula- tion is followed by a further rise in permeability after the stimula- tion has been stopped is of interest because it suggest.s that the changes in permeability are caused by a process that is initiated but not completed during contraction. At slower frequencies of stimulation the changes in permeability appear to be com- pleted in the intervals between stimuli, and no further increase is seen after the end of stimulation. In all instances studied, the effect of contraction on permeability has persisted for a very long time after stimulation (Fig. 3). This persistence is difficult to reconcile with the observation that permeability rises to different plateau values after prolonged stimulation at different frequencies (Fig. 1). Formation of a plateau suggests the exist- ence of a steady state in which a process that tends to increase permeability is balanced by an opposing process. The delay in reversal of the effects of stimulation could be explained if sequential changes having different time courses were involved in mediating the effects of stimulation on permeability. Thus, an early, readily reversible alteration in the cell that is propor- tional to the frequency of stimulation could account for the different plateaus of permeability, while a later step that is
reversible only after a long lag period could explain the prolonged persistence of the increase in permeability. Although the exact nature of these intermediary changes that alter permeability is not known, the observations suggest that several different types of alterations occur in succession.
In the isolated muscle preparation it is possible to carry out detailed studies of the kinetics of sugar transport that help to characterize the changes in permeability that occur in response to stimulation. The penetration of 3-methylglucose exhibits a saturation type of kinetics in stimulated muscles, as is the case in muscles that have been exposed to insulin (9, 10). This finding suggests that there are discrete transport sites in the celi membrane and that these sites have a limited capacity to handle the substrate that is presented to the cells.
Various kinds of changes in the kinetic characteristics of cell membrane transport systems have been described in response to insulin under different experimental conditions (23-25) ; the changes seen after electrical stimulation in the present experi- ments closely resemble the effects of insulin on the permeability of frog muscle to 3-methylglucose. In frog sartorius, insulin and electrical stimulation both increase the vmax of sugar pene- tration without appreciably altering the apparent K, (cf. Fig. 4). These observations suggest that electrical stimulation, like insulin, may affect permeability by increasing the number of transport sites that are operative. The additional finding (Table IV) that electrical stimulation and insulin both produce approximately the same maximal increase in permeability sug- gests that the same number of transport sites are available for activation by both agents. These similarities in the transport systems affected by electrical stimulation and insulin suggest that the same system may be acted upon in each instance. This hypothesis is supported by the observation that the combined effect of maximal amounts of electrical stimulation and insulin on permeability is not significantly greater than the effect of either one alone (Table IV).
One might expect that when agents differ in type as much as insulin and electrical stimulation do, they might cause their effects through different mechanisms. This expectation is borne out by the finding that, at 19”, permeability is affected promptly by contraction, whereas there is an absolute lag of approximately 30 min before insulin begins to act. The differ- ence is even more pronounced at 0”; at this temperature electrical stimulation increases the rate of penetration of sugar promptly (Table I), whereas insulin has no detectable effect in 3+ hours (10). Thus, it appears that, although insulin and electrical stimulation may increase the number of operative sites for sugar transport in the same system of the cell membrane, they produce their effects by different mechanisms.
SUMMARY
The permeability of isolated frog sartorius muscles to sugar was studied at 19” with 3-methylglucose-3H as substrate. A marked increase in permeability was observed in electrically stimulated muscles. The permeability started to rise promptly and, in the range of 3 to 20 shocks per min, increased linearly with time until a plateau was reached that was approximately proportional to the frequency of stimulation. At 0”, permea- bility increased more slowly than at 19” in response to stimula- tion, but the effect obtained after long stimulation at a given
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3500 Studies of Tissue Permeability. X Vol. 240, No. 9
frequency was approximately the same at both temperatures. When stimulation at 19” was brief and sufficiently intense, an
additional increase in permeability occurred after the end of stimulation, showing that reactions affecting permeability had not gone to completion during contraction. Permeability remained elevated for approximately 5 hours after stimulation, and then began to fall to base-line values, which were reached in about 18 hours.
At a given frequency of stimulation, the amount of work performed in a certain time did not influence the magnitude of the change in permeability. As a corollary, no relationship could be established between the changes in permeabilit,y and changes in the concentration of lactate or creatine phosphate in the muscles.
Anoxia increased the permeability of resting muscles, but, in contrast to the prompt effect of stimulation, a lag period of 60 to 90 min was noted before permeability began to rise. It is concluded that stimulation produces changes in permeability that are not mediated by anoxia. On the other hand, complete anaerobiosis was able to augment the effects of contraction on permeability, especially after intervals longer than 30 min.
Penetration of 3-methylglucose into stimulated muscles followed a saturation type of kinetics. Muscle contraction, like insulin, increased the V,,, of sugar transport without appreciably altering the apparent K,. At a given concentration of substrate, electrical stimulation and insulin produced approxi- mately the same maximal change in permeability. Maximal
amounts of stimulation and insulin together had no greater effect than either one alone, but submaximal amounts produced effects that were approximately additive. It is concluded that contraction and insulin act on the same system for sugar trans- port in the muscle cell membrane.
Acknowledgment-The authors wish to express their sincere appreciation to Dr. Carl F. Cori for his encouragement of this work and for valuable suggestions concerning the manuscript.
1. 2.
3.
4.
5. 6. 7.
8.
9.
10.
11. 12. 13.
14.
15.
16. 17.
18.
19. 20.
21.
22.
23.
24.
25.
REFERENCES
STRANDELL, B., Acta Med. &and., 66 (suppl.), 1 (1934). INGLE, D. J., NEZAMIS, E., AND MORLEY, E. H., Am. J. Phys-
iol., 166, 469 (1951). GOLDSTEIN, M. S., MULLICK, V., HUDDLESTUN, B., AND LE-
VINE, R., Am. J. Physiol., 173, 212 (1953). HELMREICH, E., AND CORI, C. F., J. Biol. Chem., 224, 663
(1957). SACKS, J., AND SMITH, J. F., Am. J. Physiol., 192, 287 (1958). DULIN, W. E., AND CLARK, J. J., Diabetes, 10, 289 (1961). NARAHARA, H. T., ~ZAND, P., AND CORI, C. F., J. Biol. Chem.,
236, 3370 (1960). HILL, A. V., AND KUPALOV, P., Proc. Roy. Sot. London, Ser.
B, 106, 313 (1929). MORGAN, H. E., AND PARK, C. R., Federation PTOC., 17, 278
(1958). NARAHARA, H. T., AND OZAND, P., J. Biol. Chem., 236, 40
(1963). FENN, W. O., Physiol. Revs., 16, 450 (1936). MEITES, L., AND MEITES, T., Anal. Chem., 20, 984 (1948). HOHORST, H.-J., in H.-U. BERGMEYER (Editor), Methods of
enzymatic analysis, Academic Press, Inc., New York, 1963, p. 266.
FURCHGOTT, R. F., AND DEGUBAREFF, T., J. Biol. Chem., 223, 377 (1956).
LINEWEAVER, H., AND BURK, D., J. Am. Chem. Sot., 66, 658 (1934).
RANDLE, P. J., AND SMITH, H. G., Biochem. J., 70, 501 (1958). MORGAN, H. E., RANDLE, P. J., AND REGEN, D. M., Biochem.
J., 73, 573 (1959). ~)ZAND. P.. NARAHARA. H. T.. AND CORI. C. F., J. Biol. Chem.,
237, 3037 (1962). ’ FISHER, R. E., AND CORI, G. T., Am. J. Physiol., 112.5 (1935). ~ZAND. P.. AND NARAHARA, H. T., J. Biol. Chem., 239, 314G
(1964). ’ CAIN, D. F., INFANTE, A. A., AND DAVIES, R. E., Nature,
196, 214 (1962). CARLSON, F. D., HARDY, D. J., AND WILKIE, D. R., J. Gen.
Physiol., 46, 851 (1963). POST. R. L., MORGAN, H. E., AND PARK, C. R., J. Biol. Chem.,
236, 269 (1961). AKEDO, H., AND CHRISTENSEN, H. N., J. Biol. Chem., 237,
118 (1962). FISHER, R. B., AND ZACHARIAH, P., J. Physiol., 168, 73 (1961)
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from

John O. Holloszy and H. T. NaraharaISOLATED FROG MUSCLE
3-METHYLGLUCOSE ASSOCIATED WITH CONTRACTION OF Studies of Tissue Permeability: X. CHANGES IN PERMEABILITY TO
1965, 240:3493-3500.J. Biol. Chem.
http://www.jbc.org/content/240/9/3493.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/240/9/3493.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on August 29, 2018
http://ww
w.jbc.org/
Dow
nloaded from