studies of the biology and control of the alfalfa blotch
TRANSCRIPT

University of Massachusetts Amherst University of Massachusetts Amherst
ScholarWorks@UMass Amherst ScholarWorks@UMass Amherst
Doctoral Dissertations 1896 - February 2014
1-1-1981
Studies of the biology and control of the alfalfa blotch leafminer, Studies of the biology and control of the alfalfa blotch leafminer,
Agromyza frontella (Rondani) (Diptera: Agromyzidae): a pest of Agromyza frontella (Rondani) (Diptera: Agromyzidae): a pest of
Massachusetts alfalfa. Massachusetts alfalfa.
John Thomas Andaloro University of Massachusetts Amherst
Follow this and additional works at: https://scholarworks.umass.edu/dissertations_1
Recommended Citation Recommended Citation Andaloro, John Thomas, "Studies of the biology and control of the alfalfa blotch leafminer, Agromyza frontella (Rondani) (Diptera: Agromyzidae): a pest of Massachusetts alfalfa." (1981). Doctoral Dissertations 1896 - February 2014. 5619. https://scholarworks.umass.edu/dissertations_1/5619
This Open Access Dissertation is brought to you for free and open access by ScholarWorks@UMass Amherst. It has been accepted for inclusion in Doctoral Dissertations 1896 - February 2014 by an authorized administrator of ScholarWorks@UMass Amherst. For more information, please contact [email protected].


STUDIES ON THE BIOLOGY AND CONTROL OF THE ALFALFA BLOTCH-
LEAFMINER, AGROMYZA FRONTELLA (RONDANI) (DIPTERA: AGROMY-
ZIDAE): A PEST OF MASSACHUSETTS ALFALFA.
A Dissertation Presented
By
John Thomas Andaloro
Submitted to the Graduate School of the
University of Massachusetts in partial fulfillment
Of the requirements for the degree of
DOCTOR OF PHILOSOPHY
September I981
Department of Entomology

STUDIES ON THE BIOLOGY AND CONTROL OF THE ALFALFA BLOTCH-
LEAFMINER, AGROMYZA FRONTELLA (RONDANI) (DIPTERA: AGRO-
MYZIDAE): A PEST OF MASSACHUSETTS ALFALFA
A Dissertation presented
By
John Thomas Andaloro
Approved as to style and content by:
Dr. David N. Ferro Dept. Head Department of Entomology
ii

John T . Andaloro 1981
All Rights Reserved

DEDICATION
This thesis is dedicated to Bridget VJalsh-Andaloro, my wife of two
and a half years, who unselfishly contributed her time in the
preparation of this manuscript. Her continued assurance, enduring
understanding, and friendship throughout my prolonged graduate ordeal
are proof of exemplary marital and Christian love for which I am
grateful.
This thesis is also dedicated to my parents, Mr. and Mrs. Samuel
and Josephine Andaloro, without whose support, encouragement, and pride
my graduate career would never have been undertaken.
iv

ACKNOWLEDGEMENTS
I wish to express my sincere gratitude to Dr. T. Michael Peters
for his guidance, encouragement, and friendship throughout the research
and preparation of this manuscript. I would like to give special
thanks to Dr.*s James B. Kring, Ronald J. Prokopy, Martin E. Weeks and
Roy Van Driesche for their time, assistance, and suggestions in all
aspects of this research.
In addition, the author extends his gratitude to the following
people:
Ms. Robyn Rufner, Electron Microscopy Lab., Univ. of Mass., for
her assistance in the taking of scanning electron microphotographs, the
Feed, Seed, and Fertilizer Lab., West Exp. Sta., Univ. of Mass., for
supplying protein analyses of alfalfa plant material, Alfred Alicandro
and David Cushing for their help in the collection of field data, and
Misters L. Chromak and E. Tidlund for performing ail the necessary
agronomic practices in the maintenance of the alfalfa test plots.
V

ABSTRACT
STUDIES ON THE BIOLOGY AND CONTROL OF THE
ALFALFA BLOTCH LEAFMINER, AGROMYZA FRONTELLA
(RONDANI): A PEST OF MASSACHUSETTS ALFALFA
(September, 1981)
John T. Andaloro, B.S., Rutgers University
M.S., Rutgers University, Ph.D., University of f^ssachusetts
Directed by: Professor T. Michael Peters
Studies were conducted from 1976-1978 to determine the population
dynamics of Agromyza frontella (Rondani) (Diptera: Agromyzidae) and
evaluate a modified harvest manipulation as a cultural control
technj.qua. Three complete generations and a partial fourth were
observed in v/estern Massachusetts with peak larval densities occurring
in early June, mid-July, mid-August, and late September. An average
U76 heat units (base 6.8^C) v/ere necessary for the development of one
generation. The first two ABL larval generations were synchronized
with the first two alfalfa harvest periods when cutting occurred at 105^
bloom. The later generations were less synchronized with time of
harvest, future larvae were commonly found mining 30 to ^10^ of the
alfalfa leaflets during the first two cuttings. Oven-dried leaflets
with at least one blotch mine weighed 8.2^ less than oven-dried unmined
leaflets. There was also a significant reduction in crude
protein/rained leaflet of 0.18 mg, 13.65^ less than unmined leaflets.
Evidence was obtained that indicated postponement of the first two
vi

harvests until after adult ABL egg deposition can reduce densities of
later ABL life stages. However, this practice resulted in unacceptable
crop quality. Older ABL larvae were observed prematurely evacuating
drying leaves. This indicates that if early harvest is to be utilized
as an effective cultural control then it must precede the development
of the third larval instar (most damaging instar), otherwise the
alfalfa must be immediately removed and not be allowed to dry in the
field.
Another cultural control investigated involved the retention of
uncut borders (perimeters) at harvest to serve as insectaries for
biological control agents of ABL. Fields with uncut "perimeters"
(approximately of crop), achieved 13.6^ fewer ABL late larval mines
during the two year study compared to the totally harvested alfalfa in
the control plots. However, increases in populations of Empoasca fabae
(Harris), Acyrthosiphon pisum (Harris), and mirids resulted in fields
where perimeters were retained compared to total harvest fields.
Reductions in alfalfa stem height, weight, and leaf surface area
occurred in these perimeter fields resulting in a yield loss of 631
kg/h, 18.5^ less than the yield of the total cut fields.
Two treatments of a malathion 5EC (0.84 L/h) plus methoxychlor 2EC
(2.2 L/h) mixture/harvest period reduced these herbivore populations
and increased yield by a mean 536 and 510 kg/h/harvest over the
untreated alfalfa in the perimeter and total cut fields respectively.
Fields with unharvested perimeters required insecticidal treatment to
achieve yields comparable to those of unsprayed totally cut alfalfa
vii

fields. Poor plant growth due to herbivore pressure the initial year
of the study was observed to result in reduced harvest yields on first
cutting the following year when cultural or chemical treatments were
not applied.
Results indicate that chemically treated and totally harvested
alfalfa maintained greater stem densities over the two year period than
did no chemical treatments and retaining uncut field portions.
Vlll

TABLE OF CONTENTS
DEDICATION. iv
ACKNOWLEDGEMENT.v
ABSTRACT.vi
LIST OF TABLES.x
LIST OF FIGURES.xi
LIST OF APPENDICES...xii
Chapter I. .1
Chapter II. POPULATION DYNAMICS OF THE ALFALFA BLOTCH LEAFMINER, AGROMYZA FRONTELLA, (RONDANI) AND ITS INJURY TO ALFALFA.
Introduction. Materials and Methods. 5
Results.. Discussion. 19
Chapter III. RETENTION OF ALFALFA CROP PERIMETERS AS REFUGES: IMPACT ON THE ALFALFA BLOTCH LEAFMINER, AGROMYZA FRONTELLA (RONDANI), OTHER SELECTED INSECTS AND CROP GROWTH.
Introduction.25
Materials and Methods. 27
Results... Discussion..
BIBLIOGRAPHY..
APPENDIX..
ix

LIST OF TABLES
Chapter II.
1. Comparison of protein content and dry weight between paired basal alfalfa leaflets...20
2. Protein loss in alfalfa leaflets due to ABL larval feeding.21
Chapter III.
3. Total number of selected insect species collected from culturally and chemically treated alfalfa. ^1
4. Alfalfa stand densities of experimental research plots.55
Appendix
5. A comparison of oven dried leaf weights and total leaf area among perimeter and total cut plots.94
6. Comparison of leaf weights as a percent of total stem weights am.ong culturally and chemically treated experimental plots. ..95
7. Cost/Benefits of alfalfa perimeter retention and chemical control of insect pests based on optimal stand density. 95
I
X

LIST OF FIGURES
Chapter II,
1, Seasonal occurrence of ^ frontella and crop develop¬ ment under a 4 harvest regime in 1977.
2, Seasonal occurrence of ^ frontella and crop develop¬ ment under a 3 harvest regime in 1977.
3, Seasonal occurrence of ^ frontella and crop develop¬ ment under a ^ harvest regime in Franklin Cty, 1978...15
4, Seasonal occurrence of frontella and crop develop¬ ment under a 4 harvest regime in Hampshire Cty, 1978.17
Chapter III.
5, Experimental plot design for studies on the effect of uncut crop borders upon alfalfa insects.29
6, Comparison of the total number of ABL mines and blotch mines/stem within unsprayed plots between perimeter and total cut fields...35
7, Effect of chemical and cultural control treatments on Empoasca fabae adults, Acyrthosiphon pisum adults and immatures, Lygus spp, adults, and other mirid adults and immatures.38
8, Effect of 1977 chemical and cultural control treatments on total alfalfa dry weight/25 stems,....44
9, Effect of 1978 chemical and cultural treatments of total alfalfa dry weight/25 stems at first harvest.46
10. Effect of 1978 chemical and cultural control treatments on total alfalfa dry weight/25 stems at second, third and fourth harvests. i\Q
11. Differences in net monetary return/h/season due to cultural and chemical control treatments.53
xi

Appendix
12, Seasonal life history of the alfalfa blotch leafminer, Agromyza frontella on harvested alfalfa.....bS
13. Spatial distribution of ^ frontella eggs at specific alfalfa stem heights.71
14. Seasonal occurrence of ^ frontella and crop development under a 4 harvest regime in Deerfield, Ma. 1977.73
15. Dorsolateral view of partially everted female A. frontella postabdominal structure.76
16. Dorsolateral view of ^ frontella ovipositor at early stage of eversion. 78
17. Dorsolateral view of ^ frontella ovipositor at intermediate stage of eversion.80
18. Nearly complete eversion of ^ frontella ovipositor.82
19. Lateral view of completely everted A. frontella ovipositor showing elongated egg laying structure on right.84
20. Oviposit ion puncture on underside of alfalfa leaflet.86
21. Comparison of 1978 peak mean ABL third instars/stem within unsprayed plots between per^imeter and total cut fields just prior to each harvest...88
22. Comparison of mean alfalfa heights from sprayed and unsprayed plots between perimeter and total cut fields at second, third, and fourth harvest periods in 1977.90
23. Comparison of alfalfa heights from sprayed and unsprayed plots between perimeter and total fields at first, second, third, and fourth harvest periods in 1978.
• •
Xll

CHAPTER I
STUDIES ON THE BIOLOGY AND CONTROL OF THE ALFALFA BLOTCH LEAFMINER,
AGROMYZA FRONTELLA (RONDANI) (DIPTERA:AGROMYZIDAE): A
PEST OF MASSACHUSETTS ALFALFA.
The alfalfa blotch leafminer (ABL), Agromyza frontella, was first
detected in Massachusetts in 1968 (Miller and Jensen 1970). It has
since spread westward to Ohio, southward to Virginia, and northward to
the Canadian provinces of Ontario, Quebec, and the Maritimes
(Hendrickson and Barth 1978a; Bereza 1977).
The taxonomy of the insect has been reviewed by both Spencer
(1973) and Steyskal (1972). Biological aspects of A. frontella in
Europe (Durseau and Jeandel 1977) and the United States (Mellors and
Helgesen 1978; Hendrickson and Barth 1978b) have also been studied.
A. frontella is widespread in Europe (Spencer 1973), but at low
population levels with rare localized outbreaks (Bollow 1955). This
has been attributed to the effectiveness of control by natural enemies.
In northeastern U.S. the ABL is considered to be an occasional pest
(Hendrickson and Barth 1978a). Reported economic losses due to larval
feeding have been estimated at $0.2 million in Quebec, in 1975 (Richard
and Gagnon 1976) and at $9/acre in Connecticut 1969-1970 (anonymous
1972).
Plants are damaged by the feeding punctures of adults (pinholing)
(Helgesen and Baxendale 1978; Durseau and Jeandel 1977) and leaf mining
of larvae (Hendrickson and Barth 1978a; Bereza 1977; Bollow 1955).
Byers and Valley (1978) have developed a tentative economic threshold
1

2
for the ABL which indicates that when approximately 22? of all alfalfa
leaflets are infested with at least one blotch mine protein loss
occurs.
Parasitism of ABL in the northeast by native species is minimal,
although season long parasitism rates of up to 36? have been reported
(Hendrickson and Barth 1979). It is believed that these native
parasites have switched over to ABL from native agromyzids, Liriomyza
trifoliearum spencer and L. trifolii. also found on alfalfa. A
possible reason for the occurrence of low levels of parasitization may
be due to the typical alfalfa harvesting practices. Harvesting removes
appreciable numbers of ABL infested leaflets in which immature
parasitoids are developing on the host larvae. Unparasitized third
instar larvae normally escape these harvesting dangers because they
have already abandoned their mines at maturity to pupate in the soil
prior to harvest. Similar occurrences were observed on Liriomyza spp.
leafminers and their parasites in California alfalfa (Jensen and
Koehler 1970),
The alfalfa blotch leafminer has not been studied in Massachusetts
since its presence was originally reported. Information on the extent
of larval mining damage, seasonal occurrence, and control strategies on
Massachusetts alfalfa needed to be collected and analyzed. This thesis
study was, thus, initiated to determine the population dynamics of A.
fronts and the relationship of ABL larval feeding to leaf weight and
protein loss. In addition, a form of modified alfalfa harvesting that
left uncut crop perimeters as a refuge to harbor beneficial-insects and
their hosts was also evaluated. Insect and plant responses to this

cultural control method were compared to conventional insecticidal
control methods.

CHAPTER II
POPULATION DYNAMICS OF THE ALFALFA BLOTCH LEAFMINER,
AGROMYZA FRONTELLA (RONDANI), AND ITS INJURY TO ALFALFA
Introduction
Agromyza frontella, (Rondani), alfalfa blotch leafminer (ABL), was
first recorded in western Massachusetts in 1968 (Miller and Jensen
1970). By 1979, A. frontella adults constituted 63/^ of the insects
swept all season from alfalfa in central and western Massachusetts
(Miller and Shaw 1969). Rapid spread throughout the northeast and
Canada quickly followed (Hendrickson and Barth 1978, Harcourt 1973) and
reached epizootic proportions in many locales.
A. frontella is widespread in Europe (Spencer 1973), but maintains
low population levels and outbreaks are localized and rare (Bollow I
1955). This has been attributed to effective natural control. In
northeastern United States, 14 species of native parasites have been
recovered from A. frontella. They have been reported parasitizing an
average 36^ of the leafminer population throughout the season
(Hendrickson and Barth 1979). It is believed that the native parasites
of the agromyzids Liriomyza trifoliearum Spencer and L. trifolii, also
found on alfalfa, have adapted to the ABL.
A. frontella is considered an occasional economic pest of
northeast alfalfa (Mellors 1979; Hendrickson and Barth 1978). Economic
losses due to larval feeding have been estimated at 0.2 million dollars
in Quebec, Canada in 1975 (Richard and Gagnon 1976) and at 9
dollars/acre in Connecticut 1969-1970 (Anonymous 1972).
A

5
Adults damage plants in the form of punctures, termed pinholing
(Helgesen and Baxendale 1978; Durseau and Jeandel 1977), for the
purpose of feeding on exuded sap. Such adult pinholing and larval
feeding have been described (Hendrickson and Barth 1978; Bereza 1977).
Byers and Valley (1978) have indicated that when approximately 22% of
the alfalfa leaflets are infested with at least one blotch mine,
significant protein loss occurs.
The extent of damage by ABL larval mining and the insect’s
population dynamics in Massachusetts have not been previously examined.
The present study was initiated to determine the seasonal occurrences
and lix’e history of A. frontella’s life stages in western Massachusetts
and to determine the relationship of ABL larval feeding to leaf weight
and protein loss.
Materials And Methods
Experimental plots. Seasonal ABL activity was studied on commercial
and university alfalfa fields in Franklin and Hampshire Counties, MA.
In 1976, 5 commercial fields were sampled weekly from July to
September. In 1977, if commercial fields in Conway (Franklin County),
elevation 500 feet, and 4 university research fields in Deerfield
(Franklin County), elevation 60 feet, were sampled weekly for A.
fVont^ life stages. Commercial fields were in close proximity to
one another and harvested 3-4 times depending on grower preference.
University fields were harvested 4 times. The first 3 harvests of the

6
latter were made when the alfalfa was in approximately 10^ bloom. By
this time many mature ABL larvae had already pupated in the soil,
thereby escaping the hazards of harvest. The fourth harvest occurred
in early to mid-October, after the first killing frost, regardless of
alfalfa height.
In 1978, ABL life stages were monitored approximately weekly in
the same ^ Deerfield plots and every 2-3 days in an Amherst (Hampshire
County) field. The harvesting scheme in both Deerfield and Amherst in
1978 was the same as for the 1977 Deerfield plots. Alfalfa stands at
all sites were 1-3 year old Saranac AR or Saranac varieties.
Sampling adults. Adults preferentially alight on the leaf tips of
alfalfa stems to feed and oviposit (Bremer 1976). When disturbed, they
make short flights to adjacent stems. This makes sweep net sampling
appropriate for estimating relative densities of A. frontella adults
despite limitations reported by other authors for other pests
(Southwood 1966; Cothran and Summers 1972). Samples consisted of 10
sweeps per site with a standard 30.1 cm dia sweep net with a 0.9m long
handle swung in a I80 degree arc through the top 20-25 cm. of alfalfa
growth. Five sites per field were sampled.
Sampling larvae. A fixed sample of 20 stems/field was randomly
selected along a diagonal transect with stems cut at ground level. The
fixed sample size was a compromise between acquiring more accurate
estimates of ABL immature stages in the higher densities and less
accurate estimates for low densities.

7
In the larval census, stems were sampled weekly at all sites,
except Amherst, where stems were taken every other day. For each stem,
the height, number of leaves, and number of mines were recorded. Mines
were divided into specific categories and used to determine specific
age classes of ABL immatures. Mining by the first and second instars
account for linear or early mines. These mines rarely exceed 4.2 mm
and 9 mm respectively in length (Hendrickson and Barth 1978). The
third instar widens the mine into the characteristic comma-shaped
blotch. The category of mature mines was further divided into
abandoned mines and mines still occupied by third instar larvae.
Host Injury Measurement. The "opposite leaflet" technique (Byers and
Valley 1978) was utilized to assess leaf weight and percent protein of
infested and non-infested leaflets. This assay treats the two basal
leaflets of an alfalfa leaf as exact duplicates. Comparative
measurements can then be made when one of the basal leaflets contained
a blotch mine and the other did not. Weight and percent protein
content of uninfested basal leaflets of alfalfa, oultivar Saranac AR,
was acquired by selecting uninfested basal leaflets from field grown
alfalfa. Two samples of 250 paired leaflets each were dried at 22
degrees C for 36h, weighed, and then analyzed for percent crude protein
by the Kjeldahl method (Horwitz 1970). Comparisons between samples
showed little differences in dry weight and only an average 0.17%
difference in crude protein. Therefore, this analysis was used on H
replicates of 250 paired trifoliolates (leaflets) each to determine
protein content and leaf weight. Each leaf had one basal leaflet with

8
at least 1 blotch mine and an opposite leaflet not infested. During
the drying process, those few larvae which still occupied the blotch
mine abandoned it and thus, did not interfere with the protein
analysis.
Results
■Seasonal phenology of A. frontella. During the 3 year period of this
study, ABL completed 3 generations per season in western Massachusetts
(Figs. 1, 3, ^). Adults from overwintering puparia appeared in
mid-May. The first larval generation extended into mid-June and ended
when mature third instar larvae abandoned their mines at the first
harvest (10^5 bloom). First generation adults were present in June and
their progeny began pupation in early to mid-July. Second generation
adults occurred through late-July and August and the ensuing third
generation larvae continued development into September. A partial
fourth generation also occurred, but larval development was protracted
and larvae eventually died due to the cooler fall vreather.
Harvesting regimes and population dynamics. The comparative effects of
different alfalfa harvest regimes on ABL larval population dynamics are
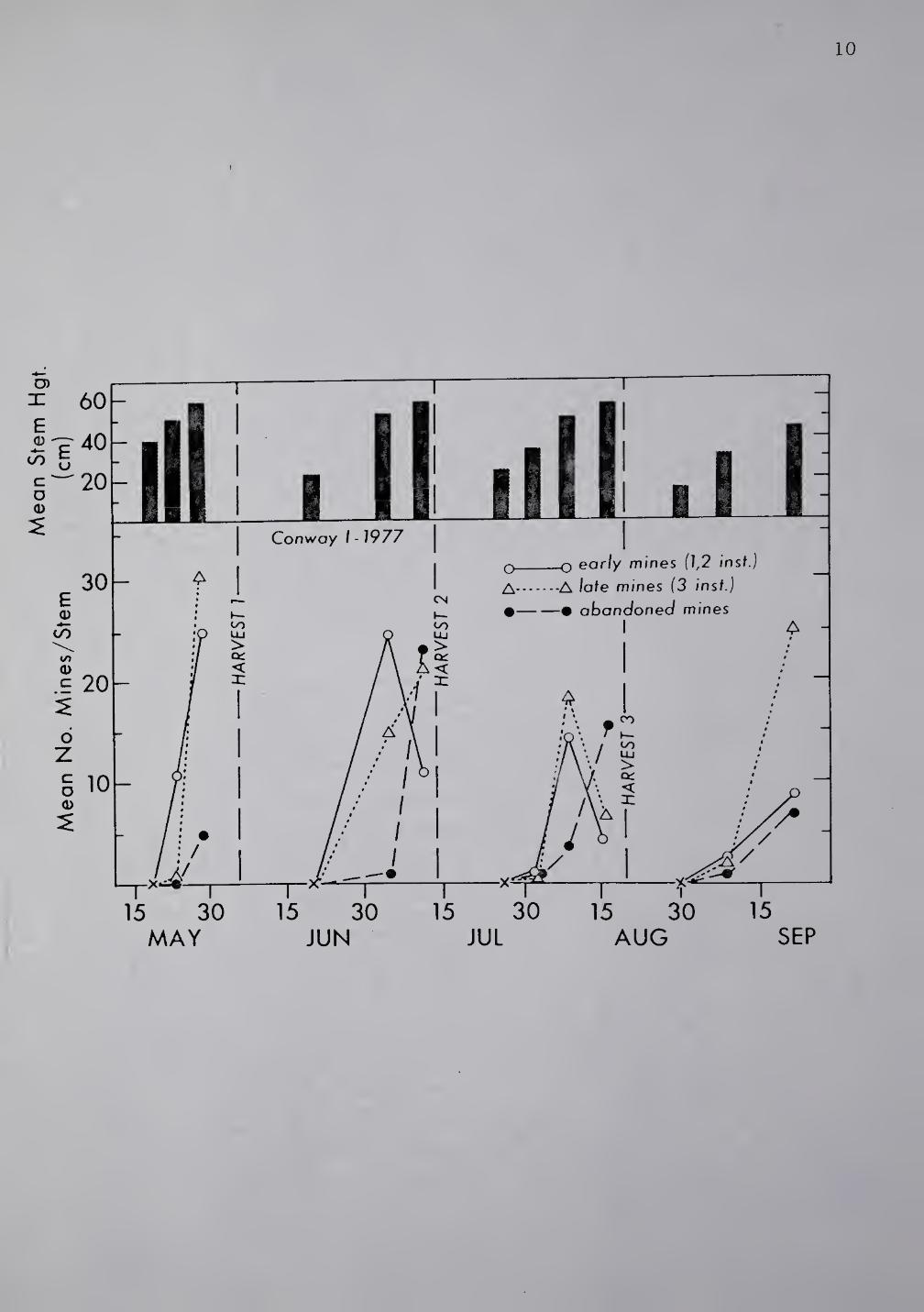
depicted in Figs. 1 and 2. First harvest of Conway I alfalfa (Fig. 1)
in early June, interfered with the majority of third instar larvae
completing development. At first harvest, an average of 5 abandoned
mines/stem was recorded. At Conway II (Fig. 2), a delay in harvest of

9
Fig. 1. Seasonal occurrence of A. frontella and crop development
under a 4 harvest regime. Conway, Franklin Co., MA, 1977. For sample
means, n=20.
Mea
n N
o. M
ines
/Ste
m
10
15 30 MAY

11
Fig. 2. Seasonal occurrence of A. frontella and crop
under a 3 harvest regime. Conway, Franklin Co., MA, 1977.
means, n=20.
development
For sample

80-
60
O) X
E a>
*^'g40l-
1-20
40-
E (U
Co «/) <D
C
O Z c o 0) 5
30-
20
10
Conway 11-1977 j p pearly mines (1,2 inst.) j
r A:—-Alate mines (3 inst.)
0— abandoned mines
I i
15 30 MAY
HA
RV
EST
2

13
two weeks resulted in a seven fold increase in abandoned mines over
that of Conway I. Initiation of first generation adult emergence
occurred in both fields the third week of June. At this time, regrowth
of alfalfa in Conway I had already achieved an average height of 20 cm,
due to the early cutting, whereas the later cut Conway II field was
still in stubble. The number of abandoned raines/stem that occurred in
Conway I during the second, third, and fourth harvest periods, despite
an early first harvest, was 22, 15, and 7 respectively. The timing of
the second and third harvests were favorable for larval development and
mine abandonment. Post harvest alfalfa in Conv^ay I was in the stubble
stage as the majority of leafminers pupated in the soil. Subsequent
fly emergence and oviposition then occurred in synchrony with new
growth of alfalfa. Conversely, in Conway II, second generation flies
oviposited on mature alfalfa which was harvested late in mid-August.
This late harvest terminated a large number of developing third
generation larvae resulting in insignificant levels of fourth
generation larvae on the third cutting. When fields are harvested in a
timely manner, third generation larvae are found in third cutting
alfalfa.
^^glglilI£,.^^equenGy and population dynamics. In 1978, the population
tr ends of both ABL adults and immatures were observed in Deerfield
(Fig. 3) and in Amherst (Fig. 4). Sampling of ABL life stages occurred
weekly in Deerfield and every 2-3 days in Amherst. Both fields were
harvested four times on approximately the same dates.
Sampling ABL lifestages every 2-3 days resulted in highly detailed

14
Fig. 3. Seasonal occurrence of A frontella and crop development
undsr a. 4 harvast ragiina. Deerfield, Franklin Co., MA, 197B. For
sample means, n=20.

Mea
n N
o.
Min
es/S
tem
Mea
n S
tem
Hgt
. (c
m)
15
/
100
80
60
40
20
40
30
Deerfield -1978
O early mines (1,2 inst.)
.‘A late mines (3 inst.) •-• abandoned mines A-A adults
B i 300
200
100 1
\ 1
CN CO
to Uj
\ •
K- »—
>
< a:
\ % \ QC
< X
1 A Uj > Q£ < X
1
Mean
N
o.

16
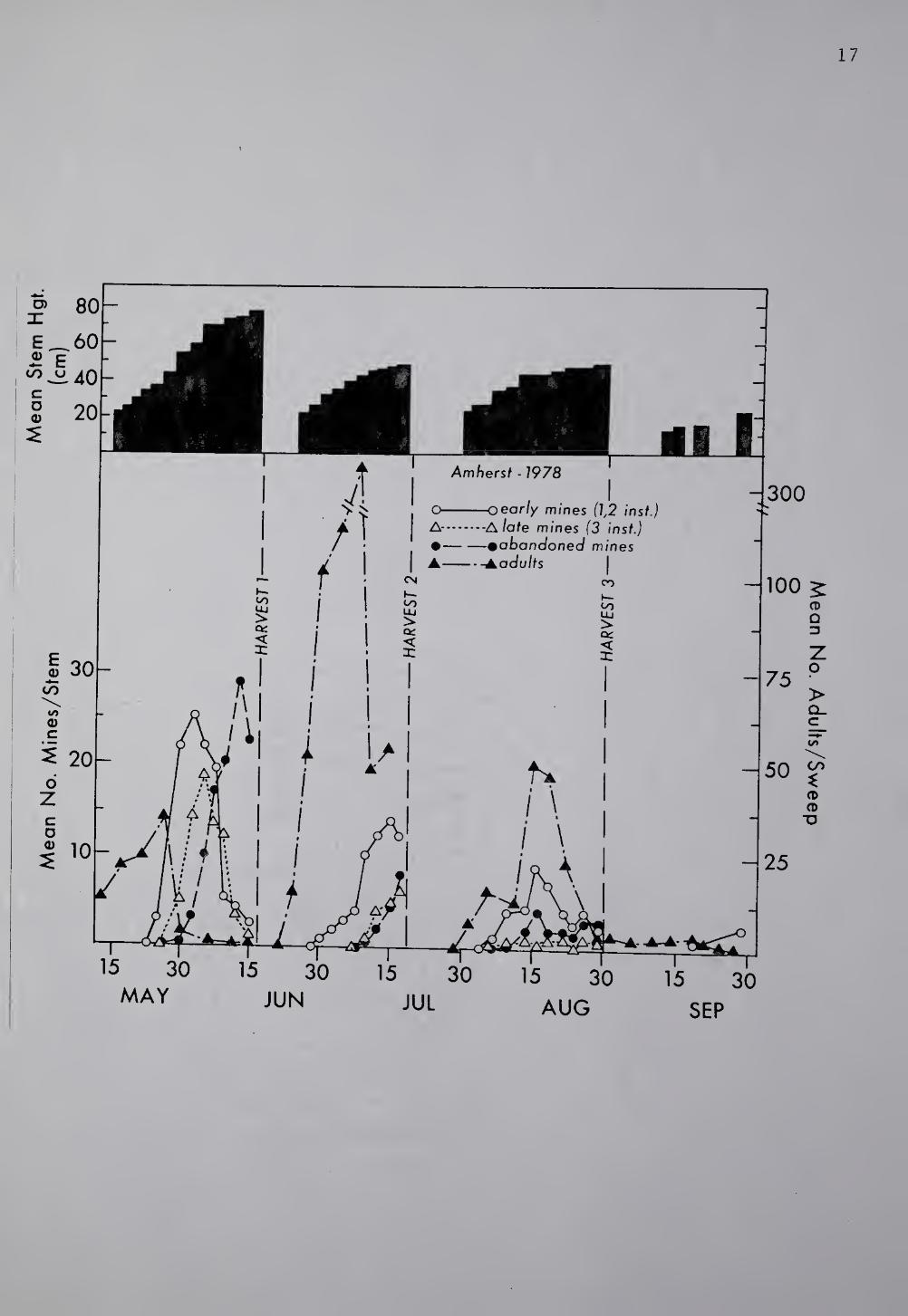
Fig. 4. Seasonal occurrence of A. fronted and crop developmen
under a 4 harvest regime. Amherst, Hampshire Co., MA, 1978. For
sample means n=20.
SEP
Mean N
o.
Adu

18
data (Fig. 4), but involved four times as many samples during the first
harvest period and approximately three times as many in the second and
third periods compared to a weekly sampling scheme. Small changes in
ABL density were more noticeable in the Amherst field where more
frequent samples were taken. At Deerfield (Fig. 3), initial emergence
of adults may have been missed by weekly sampling during the first
three harvest periods. Consequently, peak adult occurrences in both
harvest periods 1 and 2 are questionable since the level prior to the
first sampling date is unknown. The same situation arose while
sampling for early mines in Deerfield, during the first harvest.
Regardless, weekly sampling of adults and immatures in Deerfield gave
accurate indications of population trends within each harvest period
compared to similar trends achieved by more frequent sampling at
Amherst. Sampling frequency has little effect on the accuracy of the
number of abandoned mines detected since they essentially remain
unchanged, or may slightly decrease due to dehiscence of leaves, and
thus, are accumulative once larval evacuation occurs.
Developmental heat units. Heat units (base 6.8^ C) (Hendrickson and
Barth 1978) were accumulated throughout 1977 and 1978 according to
Arnold (I960) and correlated to the developmental time of the first 3
ABL generations. In 1977, a generation was determined as time between
first appearance of first larval instars of 1 generation to first
appearance of first larval instars of the next generation.
Approximately ^59 heat units accumulated for completion of the first
generation, and 525 and 500 for the second and third respectively. In

19
1978, heat units were totaled for each generation starting at first
appearance of eggs until first appearance of adults. The thermal units
accumulated during development of first, second, and third generations
in 1978 were 468, 450, and 455 respectively.
Effects of larval feeding. Mean comparisons of weight and percent
protein content of paired basal leaflets, one containing a blotch mine
and one unmined are presented in Table 1. Unmined leaflets weighed
8.2^ more than mined leaflets. A significantly lower percentage of
protein was found in mined leaflets (27.2) than in unmined leaves
(30.7), giving a 3.55^ loss. To find the actual crude protein
CP/leaflet in rag, the mean dry weight/leaflet was multipled by the
percent protein content. Table 2 lists the average CP/leaflet and the
difference in CP between unmined and rained leaflets per replicate. The
average CP loss for four replicates was O.I8 mg/leaflet or 13.8^.
Discussion
The data from this three year study indicate that the generations
of j^fomyza frontella appear synchronous with the growth periods of
alfalfa under a four harvest regime. This may be attributed to the
normal developmental time of an ABL generation of 36, 31, and 27 days
at 20, 23, and 26 C respectively (Hendrickson and Barth 1978) which
closely relates to the time temperature requirements for an alfalfa
harvest period. The lack of overlap between generations may also be
due to the harmful effects of harvesting on ABL life stages. Removal

TABLE 1
Comparison of protein content and dry weight between paired basal alfalfa leaflets.^
Mean^ Dry Wt/leaflet (mg) Protein Content (%)
Unmined A.40± 0.
Mined^ 3a 4.04± 0.4a
Uninined 30.7± 3.4a
Mined^ 27.2± 0.9b
Means followed by different letters indicate a significant difference at the P = 0.01 level.
Average of four replicates; one replicate = 250 pairs of basal leaflet.
Blotch Mine either complete (abandoned) or near complete (containing mature larvae) .

TABLE 2
Protein loss in alfalfa leaflets due to ABL feeding.
Crude Protein/Leaflet
Unmined Mined Difference Crude Protein Loss/Leaflet (%)
1.3±0.05a 1.12+0.08b 0.18±0.04 13.8%
Crude Protein/Leaflet = dry wt/leaflet x protein content (%).
Means followed by different letters indicate a significant difference at the P = 0.01 level.

22
of alfalfa canopy exposes the soil to direct ■mlight and results in
increased soil temperatures with decreases in soil moisture (Pinter et
al 1975). These conditions speed pupal development beyond the rate
which occurs under a canopy cover (Bremer 1976) and may also reduce
pupal survival due to harsh conditions.
Reduction of A. frontella populations appears to be an attainable
goal for alfalfa producers. Insecticidal control of ABL has proven to
be effective, but unacceptable for economic and environmental reasons
particularly because of the interference with the natural enemies which
exists between the alfalfa weevil Hypera postica (Gyllenhal) and its
natural enemies. However, the use of cultural controls would be less
costly, more advantageous to the stability of other alfalfa insect
complexes, and more acceptable to the grower. An effective cultural
control of ABL involves early harvest of alfalfa (Bremer 1976) which
removes large numbers of immatures from the field along witn the plant
matter. In this study, the first harvest of Conway I (Fig. 1) was
assumed to be too la.te to reduce the subsequent second larval
generation. Observations in the laboratory on the ability of older
larvae to evacuate their mines and form puparia either because of loss
of turgor or temperature change indicate that many nearly mature larvae
may escape the adverse effects of harvest. This phenomenon probably
also occurs in the field after alfalfa is cut and left to dry. Thus,
early harvest of alfalfa for leafminer control is not only to achieve
maximum yield of crop, but may be ineffective unless it is implemented
prior to the appearance of third instars (blotch mines), particularly
since it is this stage which causes the majority of damage.

23
Conversely, delaying first and second harvests as occurred at Conway II
(Fig. 2) results in minimal shelter for emerging first generation flies
and in the removal of newly laid eggs and young larvae of the third
generation. Late harvest is a less acceptable method of cultural
control due to the reduction of alfalfa quality and the loss of
dehisced lower leaves during post bloom.
Predicting the general time of ABL occurrence through v;eather
measurements could aid in the optimal sampling of ABL adults. The
average number of ambient thermal units, base 6.8^ C, accumulated from
the field in 1977-78 was found to be similar to the 484 heat units
reported by Hendrickson and Barth (1978) for laboratory reared flies.
Such consistency indicates the use of thermal unit accumulations as an
adequate tool for use in forecasting ABL generations. Since the
immature ABL spends approximately two-thirds of its time pupating in
the soil, acquiring thermal units based on soil temperatures may be
more accurate in predicting adult emergence.
Although A. frontella is considered an occasional economic pest of
alfalfa, studies comparing yields of alfalfa treated with insecticides
have shown no significant yield increases (MacCollom et al 1980, Byers
and Valley 1978), The results of our experiment indicated that oven
dried leaflets infested with one or more blotch mines weighed 8.2?^ less
than uninfested leaflets. We also found an average 13.8^ reduction in
protein content/infested leaflet which concurs with the results of
Byers and Valley (1978). This may represent potential economic loss,
if many leaves have mines, assuming supplemental protein is.required to
replace the protein lost to the leafminer.

24
Reduction in plant weight and digestable dry matter may be of
greater significance than indicted in this study dependent upon the
extent of larval infestation. Chronic leafmining damage may also
adversely affect the plant's susceptibility to crown rot, its ability
to overwinter, and stand longevity.

CHAPTER III
RETENTION OF ALFALFA CROP PERIMETERS AS REFUGES:
IMPACT ON THE ALFALFA BLOTCH LEAFMINER, AGROMYZA
FRONTELLA (RONDANI), OTHER SELECTED INSECTS AND CROP PRODUCTION
Introduotion
The alfalfa blotch leafminer (ABL) Agromyza frontella (Rondani) is
a European agromyzid species recently introduced into the United States
(Miller and Jensen 1970). It has since spread to Ohio, V/est Virginia,
Ontario, Quebec, and the Maritimes (Hendrickson and Barth 1978, Bereza
1977). Large adult (Kim 1975) and larval (Andaloro et al 1978)
populations occur in the northeast, with occasional reportings of
economic losses in alfalfa yield and quality (Byers and Valley 1978,
Richard and Gagnon 1976). In Europe A, frontella is of little economic
importance (Bollow 1955); a condition attributed to the high percentage
of parasitization (70-90^) by a complex of parasitoids (Durseau and
Jeandel 1977). In the northeastern U. S,, a complex of parasitoids
exists and is believed to have adapted to ABL from native agromyzids
found on alfalfa (Hendrickson and Barth 1979). This parasitoid complex
was responsible for a season long average parasitism rate of 36% in
Delaware, where 5 host generations occur. The average parasitization
rate for the first two ABL larval generations was ^3%, whereas, during
the remaining 3 generations average parasitization was 55^. Low levels
of parasitism early in the season were attributed to poor parasite-host
synchrony and too few parasitoids. Conventional alfalfa harvesting
practices which remove the leafminer host and drastically alter the
25

26
physical environment may prevent the buildup or rebound of the ABL
parasitoid complex early in the season. Such harvesting also removes
appreciable numbers of ABL-infested leaflets in which immature
parasitoids are developing on the host larvae. Many mature ABL larvae
normally escape these harvesting dangers because they have already
abandoned their mines to pupate in the soil. Such harvesting appears
to be less harmful to the leafminers than to their parasites. Similar
phenomena were observed on Liriomyza leafminers and their
parasites in California alfalfa (Jensen and Koehler 1970).
A variety of harvest modifications have been employed in alfalfa
insect pest rnamagement. Certain of these innovative harvest practices
were designed to reduce environmental instability (VanDen Bosch and
Stern 1969) and allow for maximum natural enemy conservation while
others either suppressed pest numbers by early removal of hay (Bremer
1976) or reduced pest emmigration into adjacent high cash crops (Stern
et al 1964). The objective of this study was to evaluate a form of
modified alfalfa harvesting that left uncut crop perimeters as a refuge
to harbor beneficial insects and their hosts. The effects of retained
crop perimeters on other alfalfa pests and the subsequent impact that
their population levels had on alfalfa crop production were also
monitored. Insect and plant responses to this cultural control method
were compared with responses to conventional insecticidal control
methods.

27
Materials And Methods
Experimental plot design. A two year old stand of Saranac AR variety
alfalfa in Deerfield, Franklin County, MA was divided into H 30m by 15ra
fields (Fig. 5). At each harvest, 2 of these designated "total cut"
fields (C), were completely harvested. Starting with the first cutting
the other 2, specified as "perimeter" fields (P), were left with an
uncut crop border or perimeter of 0.6m width (10-12^ of total area
represented by dotted lines in (Fig. 5). At the second harvest, an
uncut crop border of similar size was established inside the original
border which was then cut and removed with the rest of the crop. At
the third harvest, the outermost portion of alfalfa was again retained
as the uncut refuge. There was no fourth harvest either year,
although, data collected post 3rd harvest are referred to as occurring
in the fourth harvest period.
Each of the 4 fields was further subdivided into 4, 8m by 15m
plots. Two plots (labeled S) in each field were sprayed 1-3 times per
harvest with a malathion (5EC) plus methoxychlor (2EC) mixture (0.84L +
2.2L/ha) after the initial harvest. The remaining two plots (labeled
U) in each field were left unsprayed. The uncut crop borders in the
perimeter fields were not sprayed.
In an attempt to minimize movement of insects between experimental
areas, 4m wide strips of field corn (1977) and Balboa rye (1978) were
planted between and around the fields.

28
Fig. 5. Experimental plot design for 1977-1978 studies on the
effect of uncut crop borders upon alfalfa insects. Unsprayed (U) and
sprayed (S) plots are located within the perimeter (P) and total cut
(C) fields. (The dashed line within the P fields indicates position of
the originally retained border. Dashed line between fields indicates
sites of 2m high tobacco net fence). Due North points directly to the
upper right hand corner, Franklin Co., MA.

30
m

30
Sweep samples. A sweep net was chosen as the sampling tool to
determine the relative densities of the alfalfa pests and predators
included in the study. It*s use for sampling alfalfa insects has been
well documented (Pruess et al 1977, Cothran et al 1975, and Callahan et
al 1966).
A sweep sample consisted of three 180° sweeps with a standard 30.1
cm diameter insect net through the top 20-25 cm of alfalfa. Two random
samples/week were taken from each plot except v/hen alfalfa was in the
stubble stage. No samples were taken directly from the uncut crop
borders in the perimeter fields.
Insects swept in 1977 were identified and separated into 7
categories: adults of 1) A. frontella, 2) Empoasca fabae (Harris)
(potato leafhopper), and 3) Lygus spp.; all life stages of
Acyrthosiphon pisum (Harris) (pea aphid), 5) nabids,
6) coccinellids, and 7) plant bugs other than Lygus. In 1978, the
only changes in tallying insect taxa included the separation of plant
bug adults from their nymphs and the addition of potato leafhopper
nymphs and Therioaphis maculata (Buckton) (spotted alfalfa aphid)
adults and nymphs.
Stem sampling. Stem samples were collected for stem height, weight,
and leaf area and to document the ABL stages. Each sample consisted of
35 random main alfalfa stems collected at 2-3 step intervals along a
diagonal transect and cut at ground level. Samples did not-include
stems from the uncut border nor from the area where borders had been

31
during the previous harvest.
Plant height was determined for 25 stems from each sample. Stems
were then defoliated and both leaves and stems were oven dried at
75° C for approximately 30 h. Immediately after drying, weights of the
25 stems and also of the leaflets were recorded. Population densities
of the 3 ABL larval instars were determined on the remaining 5 stems.
Mines were used to determine specific age classes of immature ABL*s.
Mining by the first and second ABL larval instars in linear or early
mines. These mines rarely exceed 4.2mm and 9mm respectively in length
(Hendrickson and Barth 1978). Third (final) instar larvae widen the
^ine into the characteristic comma—shaped blotch. This category was
further divided into mines already "abandoned” by mature larvae which
had.dropped to the soil to pupate and mines still occupied by either
live or dead third instar larvae. Any leaflet which exhibited any of
the ABL type mines was categorized as a mined leaflet.
Results
Response of A. frontella to control treatments. Sampling A. frontella
adults during first harvest period, prior to initiation of the
experiment, indicated homogeneity among the plots. During the
remaining three harvest periods in 1977, there was no overall
significant difference (P=0.45) betv/een total number of flies swept
from the perimeter (P) and total out (C) fields. Analysis was based on
the variance of the fly populations observed over the 11 sampling

32
dates, A significant difference (P=0.06) occurred in 1978, as a
greater number of ABL adults were collected from the P fields over all
sampling dates. Statistical comparison tests performed on
individual sampling dates in 1978 revealed no significant differences
in overwintering fly numbers between the P and C fields during the
first alfalfa growth before the initial retention of borders.
Significant differences in number of first generation adults did occur
between the P and C fields in 1978 after the establishment of borders.
Uncut alfalfa borders accumulated adults which had emerged when the cut
alfalfa was at minimal growth. Over a prolonged period the uncut
alfalfa vjithin the retained perimeter became rank and harbored fewer
flies.
‘ Methoxychlor plus malathion treatments applied 1~2 days prior to
sampling in 1977 significantly (P=0.01) reduced A. frontella
populations in the S (sprayed) plots. Insecticidal treatments applied
5 to 7 days prior to sampling in 1978 resulted in no significant
differences in fly numbers between the U (unsprayed) and S plots. This
may have been due to the intense fly populaton pressure in 1978, short
residual of malathion and methoxychlor, and taller more lush alfalfa in
the S plots causing fly dispersal from the unsprayed plots. During one
particular sampling date during first generation in 1978, significantly
more flies were collected from the S plots of both the P and C fields.
The numbers of ABL mines/stem, particularly blotch mines, v^ere
used to assess jft, frontella larval populations and the extent of plant
damage. Differences in numbers of blotch and abandoned mine numbers
within the plots were also used as a comparative measure of successful

33
larval development and, thus, an indication of the cumulative effect of
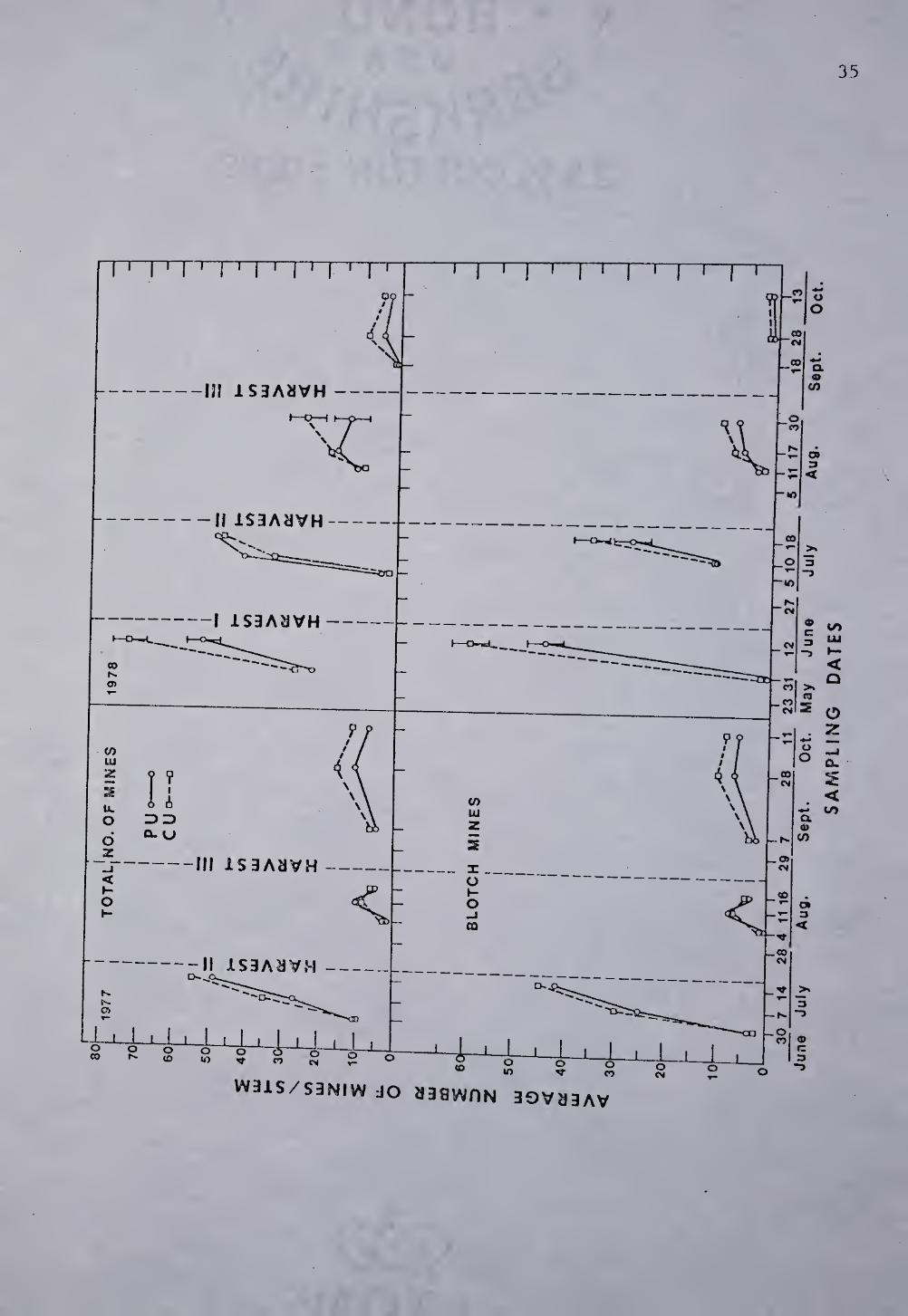
natural enemies. Fig. 6 shows a comparison of the number of blotch
mines (either abandoned or still occupied by live third instars)/stem
and the total number of mines (formed by first, second, third or
abandoned third instars)/stem found within the PU and CU plots in 1977
and 1978. Since insecticidal treatments reduced the larval populations
to such low levels in all the S plots, the data from the PS and CS
plots are not presented.
In 1977, mine populations in all unsprayed plots were similar. In
1978 significantly fewer mines were found in the perimeter plots.
There were more total mines at first and third harvest and a greater
number of blotch mines at first and second harvest in the CU plots than
in the PU plots. This is the reverse of A. frontella adult response
where greater fly numbers were swept from the PU plots in 1978.
Third instar ABL mortality V7as measured in 1978. The number of
mines containing dead third instars was counted separately from the
number of blotch mines containing live third instars and abandoned
mines in Fig. 6. There was an overall significant (P=0.01) difference
in the total number of dead third instars between the PU and CU plots
in 1978. The most pronounced difference occurred during second harvest
period where more dead third instar larvae/stem were found in the PU
plots (8) than in CU plots (4.8).
Msponse of selected insects to control treatments. The effect of the
2 control regimes upon the more important alfalfa insect pest
populations present during the experiment are sho\m in Fig. 7.

34
Fig. 6. Comparison of the total number of ABL mines (formed by
first, second, and third abandoned third instars) and blotch nines
(formed by third and abandoned third instars)/stem within upsprayed (U)
plots between perimeter (P) and total out (C) fields in Franklin Co.,
MA. If means differ significantly at a particular sampling date they
are indicated by 95? confidence limits according to Scheffe’s multiple
comparison test. For each sample mean, n=20.
35
SA
MP
LIN
G
DA
TE
S

36
Response of some insect populations to the treatments during post
treatment may be misleading. Close proximity of sprayed plots to
unsprayed plots created a situation where insects dispersed from
unsprayed (U) plots of high densities to the sprayed (S) plots of low
densities. This insect movement masked the total effectiveness of the
chemical treatment due to rapid recolonization of the S plots, but
concurrently reduced insect numbers in the U plots.
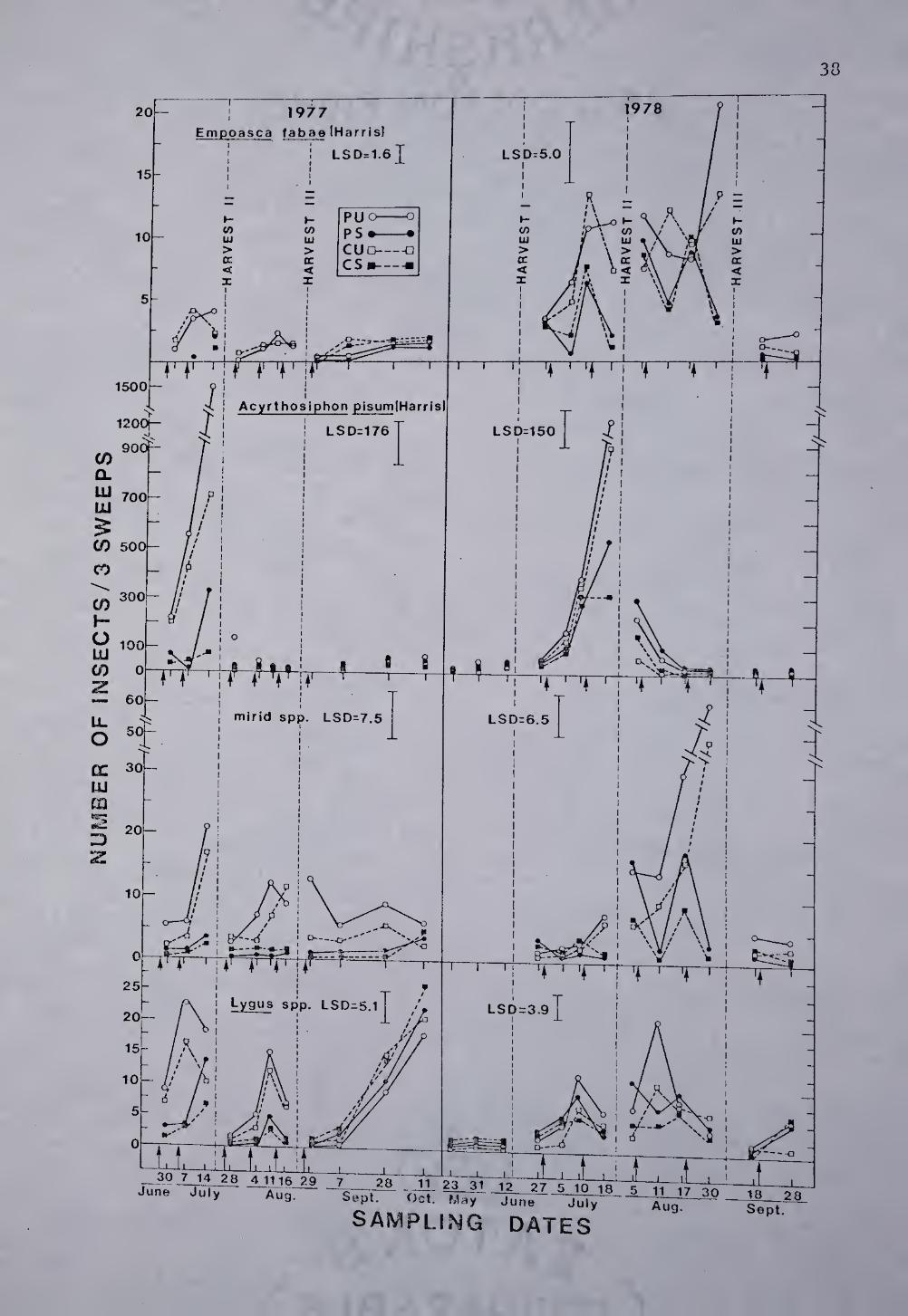
E, fabae was first collected in Massachusetts within the first 2
weeks of June, just after first harvest in both 1977 and 1978.
Economically injurious populations (Edwards et al 1978) occurred during
second harvest in 1977 and during second and third harvest in 1978
(Fig. 7). Insecticidal treatments significantly reduced PLH numbers in
the'S plots during second and third alfalfa growth periods both years.
However, in 1978 chemical control measures did not adequately suppress
—• ^^^ae populations belov; the economic threshold level of 1 to 2
adults and or nymphs/sweep. A significantly greater number of
leafhoppers occurred in the PU plots than in the CU plots only on
August 30, 1978. Sampling within the 0.6m wide uncut perimeter in 1978
revealed a 10-20 fold increase in PLH*s compared to numbers found in
the cut portion of the P plots. Large numbers of PLH persisted within
the border even as the alfalfa aged.
Pea aphid populations were greatest during second harvest in July
1977 and 1978 (Fig. 7). Applications of methoxychlor plus malathion
significantly reduced aphid numbers in the S plots compared to those in
the U plots. In 1978, despite application of 2 sprays during second
harvest period, pea aphids were still numerous in the S plots.

37
Fig. 7. Effect of chemical (sprayed, S; unsprayed, U) and
cultural (perimeter, P; total cut, C) control treatments on Empoasca
fabae adults, Acyrthosiphon pisum adults plus imraatures, Lygus spp.
adults, and other mirid adults plus immatures in Franklin Co., MA. * ,
Least significant differences (LSD) represent significance to the P=.05
level for each sample mean, n=8.
i
NU
MB
ER
OF IN
SE
CT
S/3 S
WE
EP
S
38

39
Significantly greater aphid numbers were found in the perimeter U and S
plots compared to the total cut U and S plots at time of peak
population. Pea aphid numbers were observed to build up faster in the
PS plots than in the CS plots following pesticide applications in the
second harvest period.
Fluctuations in the densities of specific mirids under the two
treatment regimes are presented in Fig. 7- Response of adult Lygus
spp. populations, predominately Lygus lineolaris Palisot de Beauvois,
varied considerably from that of other mirid spp. and are, thus,
depicted separately. The prevalent mirid species other than Lygus spp.
were Adelphocorus lineatus (Goeze), A. rapidus (Say), Plagiognathus
politus Uhler, P. cuneatus Knight, Poecilocapsus lineatus (F), and
Reuterscopus ornatus (Reuter).
Lygus bugs and other mirids vjere present in high numbers
throughout 1977 and 1978. Lygus bug adult numbers were greater than
all other mirid adults and nymphs together at all harvest periods
except third harvest in 1978. Insecticide sprays had a lesser effect
on adult Lygus spp. than on remaining plant bugs. Lygus adults were
one of the first insects to recolonize sprayed plots and when left
uncontrolled (Sept. - Oct. 1977), rapidly reach high density levels.
Significantly greater numbers of adult Lygus bugs were present in
the PU plots than in the CU plots at 5 sampling dates throughout the
study. The reverse occurred only once in 1977. Populations of mirid
spp. other than I^gus responded to perimeter retention, in the same
manner as adult Lygus. Differences in mirid numbers between the PU and
CU plots were most pronounced at third harvest in 1978 when highest

40
populations were present. Significantly more bugs were found at this
time in the PU and PS plots compared to the CU and CS plots.
Nabis alternatus (Parshley) and N. americoferus (Carayon) were the
only representatives of the nabid taxa found in the research plots,
Nabids were the most abundant insect predators sampled, with adult plus
nymphal populations reaching up to 10/sweep in the unsprayed plots.
Nabid population levels within unsprayed plots varied little from that
of the sprayed plots both years. The retention of uncut borders had
little impact on nabid numbers. There was little variation in
population levels between perimeter and total cut unsprayed fields.
Peak populations of coccinellid adults plus immatures never
exceeded 1 and 1,25/sweep in 1977 and 1978 respectively. The
predominant coccinellid species collected were Coleomegilla maculata
Timberlake, Coccinella trifasciata Mulsant, and Coccinella
transversoguttata Brown. These predator species were detrimentally
affected by insecticidal treatment, with populations suppressed to near
zero in 1977 and at second and fourth harvests in 1978. There were no
significant differences in coccinellid numbers between the PU and CU
plots either year, nor was there a significant correlation between
coccinellid and aphid numbers.
Summary of treatment effects on insect population densities. The total
number of individuals of the selected taxa that were collected from
each field at the 25 sampling dates is listed in Table 3. Of the
72,621 herbivorous individuals (excluding ABL life stages) swept from
the U plots, 42,921 were captured in the P fields, 1.5 times the number
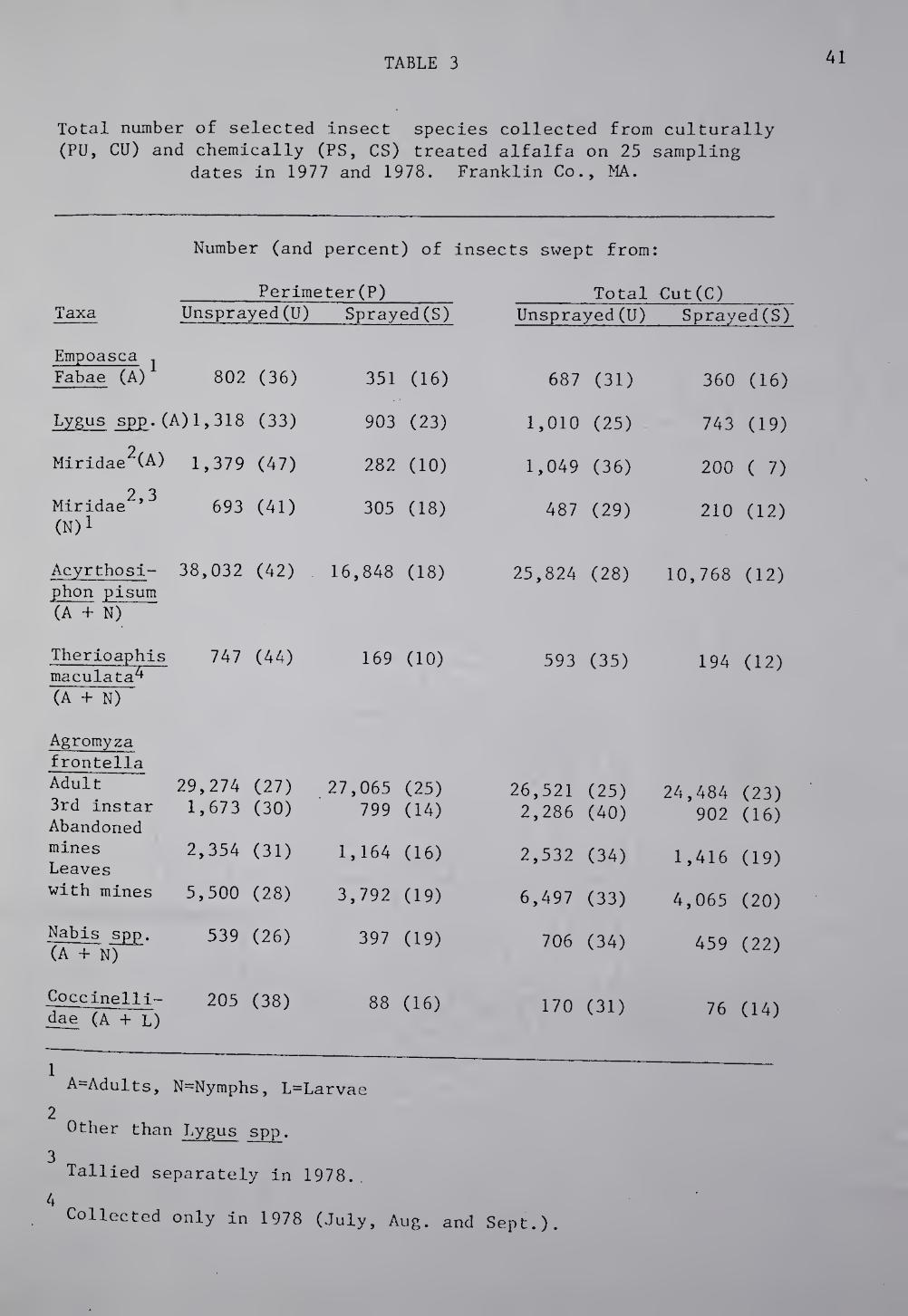
TABLE 3 41
Total number of selected insect species collected from culturally (PU, CU) and chemically (PS, CS) treated alfalfa on 25 sampling
dates in 1977 and 1978. Franklin Co., MA.
Number (and percent) of insects swept from:
_Perimeter(P)_ Total Cut(C) Unsprayed(U) Sprayed(S) Taxa
Therioaphis maculata4 (A + N)~
Agromyza frontella Adult 3rd instar Abandoned mines Leaves with mines
Nabis spp. (A + N)
Unsprayed(U)
747 (44)
29,274 (27) 1,673 (30)
2,354 (31)
5,500 (28)
539 (26)
Sprayed(S)
351 (16)
903 (23)
282 (10)
305 (18)
16,848 (18)
169 (10)
27,065 (25) 799 (14)
1,164 (16)
3,792 (19)
397 (19)
88 (16)
687 (31)
1,010 (25)
1,049 (36)
487 (29)
25,824 (28)
593 (35)
26,521 (25) 2,286 (40)
2,532 (34)
6,497 (33)
706 (34)
170 (31)
360 (16)
743 (19)
200 ( 7)
210 (12)
10,768 (12)
194 (12)
24,484 (23) 902 (16)
1,416 (19)
4,065 (20)
459 (22)
76 (14) Coccinelli- 205 (38) dae (A + L)
Empoasca Fabae (A)^ 802 (36)
Lygus spp.(A)1,318 (33)
Miridae^(A) 1,379 (47)
Miridae^’^ 693 (41) (N)l
Acyrthosi- 38,032 (42) phon pisum (A + N)
A Adults, N—Nymphs, L=Larvae
2 Other than Lygus spp.
3 Tallied separately in 1978.
4 Collected only in 1978 (July, Aug. and Sept.).

from the CU plots. Perimeter retention in the PS plots resulted in a
34it increase of herbivorous insects over the number collected in the CS
plots.
Seventy percent of all herbivores (including ABL) collected from
the sprayed and unsprayed plots were from the unsprayed plots. The use
of rnalathion plus methoxychlor reduced pest numbers by a factor of 2.3.
Insecticidal use reduced the 2 predator complex populations by a factor
of 1.6.
Mature ABL larvae (third instar), abandoned mines, and leaves with
mines were distributed unevenly with a greater percentage in the total
cut fields. This is the reverse of the trend in overall response of
other herbivorous insects to the cultural treatment.
Effect of control treatments on stem height, weight, and total lea_f_
area. Plant growth differed greatly in the experimental plots in 1977
and 1978. Alfalfa treated with insecticides was significantly taller
than unsprayed alfalfa within the perimeter and total cut fields in
1977 and 1978. Alfalfa height was significantly reduced within both U
and S plots of the P fields. In 1978, at first and third harvests, the
CS plots produced significantly taller alfalfa than the PS plots. In 4
of the harvest periods during the 2 years, the CU plots yielded alfalfa
of a mean height statistically the same or greater than that of the PS
plots.
Differences in oven dried stem weights from the various treatments
were consistent with their respective height differences and are shown
in figs. 8, 9, and 10. However, statistical differences in stem height

43
Fig, 8, Effect of 1977 chemical (sprayed,S; unsprayed, U) and
cultural (perimeter, P; total cut, C) control treatments on total (stem
+ leaf) alfalfa dry weight/25 stems at second, third and fourth
harvests in Franklin Co., MA. At each harvest date means accompanied
by different letters indicate a significant difference at P=.05
according to Duncans New Multiple Range test. For each sample mean,
n=4.

44
r 1 IS
(0
o JD u
D o
“D (D >.“D
o D ®
5 £ ^ >.
a o lim in !>-
O o C Q. a. D in
It ii II II u O. 3 LO
h-
O)
(D
bD
f“n I I r I i I I 1."I'” r I I I lO O lO o csj CM
SUJ9JS SZ/(S)mSi3M X
CU
LT
UR
AL
TR
T.

45
Fig. 9. Effect of 1978 chemical (sprayed, S; unsprayed, U) and
cultural (perimeter, P; total cut, C) control treatments on total (stem
+ leaf) alfalfa dry weight/25 stems at first harvest in Franklin Co.,
MA. Means accompanied by different letters indicate a significant
difference at P=.05 according to Duncans New Multiple Range test. For
each sample mean, n=4.

Iyii
ei2
,i9
78
46
Q> "a
© s
O Ss^ ■3 rs e>
^_ E a» >» fS CL ra
Lm
© Q c o. 3 (f)
II II II II
CD
.T’T I I ' i If) o U) If)
If)
Cu
ltu
ral
Trt
.

47
Fig, 10. Effect of 1978 chemical (sprayed, S; unsprayed, U) and
cultural (perimeter, P; total cut, C) control treatments on total (stem
+ leaf) alfalfa dry weight/25 stems at second, third, and fourth
harvests in Franklin Co., MA, At each harvest date means accompanied
by different letters indicate a significant difference at P=.05
according to Duncans New Multiple Range test. For each sample mean,
n=4.

T3 o 0>
>»t3 3 o m 0) O E La
cx ra ra La (/) b o O s a.
& II J» li II
O d. =>
CO CU
LT
UR
AL
TR
T.

49
did not always indicate a significant difference in stem weight. In
1977, stem weights were statistically greater in the C fields than in
the P fields at third (August 16) and fourth (October 11) harvest
periods (Fig. 8). Stem weights also were greater in the S fields than
in the U fields at second (July 14) and fourth harvest periods. The
first alfalfa crop in 1978 achieved the greatest stem weights (Fig. 9).
Although the alfalfa had not yet been subjected to either cultural or
chemical treatments for the 1978 season, the weight differential/25
stems between alfalfa in P and C labeled fields was the greatest of any
harvest period. There was a total dry stem weight increase of 11
grams/25 stems from alfalfa in CU plots compared to that in PU plots.
This weight difference was mainly due to stem rather than leaf weight.
No significant differences in total stem weights existed between S and
U plots within either P fields or C fields at this time. During the
third and fourth harvest periods in 1978, (Fig. 10) PU alfalfa weight
was significantly less than CU alfalfa weight. During the final three
harvests in 1978 the sprayed alfalfa weighed more than the unsprayed
alfalfa within each cultural treatment.
Leaf area is proportional to leaf weight and the factor 331.5cm^/g
oven dried leaf has been calculated by Robinson and Massengale (1967)
for Moapa variety alfalfa. The total leaf area (TLA) is twice the
measured leaf area value since entomophagous insets search both the
upper and lower sides of leaves. In this study, the two year overall
average of TLA at harvest for PU, PS, CU, and CS plots were 64.4, 75.5,
77.1, and 87.6 sq. m/sq. m of ground surface. Leaf area was greater in
the insecticide treated fields compared to the unsprayed fields and in

50
the total cut fields compared to the perimeter fields.
The effect of treatments on alfalfa yield and crop value. Optimal
yield was derived by correcting mean oven dry alfalfa weight/25 stem
sample to optimal stand density of 350 stems/sq. m and converting to
kg/ha. The overall mean for all harvests in 1977 and 1978 for PU, PS,
CU, and CS plots are 2782, 3318, 3^13, and 3923 kg/ha respectively.
Malathion plus methoxychlor treatments significantly increased the mean
yield of treated plots over untreated plots at harvest by approximately
536 kg/ha or 16^ in P fields and 509 kg/ha or 13^ in C fields.
Retention of uncut perimeters at each harvest had a statistically
significant negative effect on forage yield: unsprayed P plots yielded
a mean deficit of 631 kg/ha/harvest or 18.5^ less than that of
unsprayed C plots. Sprayed P plots yielded a mean reduction of 604
kg/ha/harvest or ^5% less than sprayed C plots. Percent differences in
yield between P and C fields do not reflect unharvested crop perimeters
which were retained. Yield differences are only those of the harvested
areas.
Differences in yield do not accurately indicate what the monetary
return per treatment is since the final yield does not reflect
accumulated treatment costs. Thus, a cost/benefit analysis on the
chemical and cultural control treatments was obtained to acquire a more
realistic view of the results of the treatment. Chemical
cost/treatment was valued at $7.95/ha. There were 6 insecticidal
treatments in 1977 and 5 in 1978. Costs for labor, machine, use, and
depreciation relative only to chemical application were evaluated and

51
combined with chemical cost to equal a total of $16.05/ha/treatment.
Retention of a border left approximately of the crop unharvested
aiid, thus, comprised a ^0% loss in yield/season. Alfalfa was valued
both years at $88,14/I000kg of dried alfalfa (Lyford 1979). All other
expenses such as harvesting, fertilizer use, and labor costs which were
similarly performed on all the plots were not included in the
cost/benefit analysis. For the purposes of this study no additional
cost was subtracted to compensate for possible reduced quality of
alfalfa which made up the perimeter.
The annual monetary net return/ha for alfalfa harvested from the
research plots after chemical and cultural treatment costs were
subtracted, are depicted graphically in Fig. 11. The monetary mean
return/ha of the unsprayed total cut plots (CU) was chosen as a base
line or check from which all comparisons were made.
The PU and PS plots in 1977 similarly netted an approximate $165
loss/ha/yr compared to the CU control plots. The disparity in monetary
net return between the PS and CU plots is due vjholly to chemical and
cultural treatment costs because the alfalfa yields of the 2 plots were
similar. Although the PS plots out-yielded the PU plots by
approximately 1300kg/ha for the 1977 season, the monetary return did
not significantly offset the chemical costs necessary to achieve that
difference.
The 1978 CS and PS plots netted approximately $ 100/ha more than
their respective unsprayed counterpart plots. Retention of a perimeter
in the 1978 PU plots resulted in a loss of about $390/ha/yr .due to the
combination of reduced yields and the 10? hay loss for border use.

52
Fig. 11. Difference in net monetary return/h/season due to
cultural and chemical control treatments using the total cut unsprayed
(CU) plots as a baseline. Franklin Co., MA. P=perimeter; C=total cut
S=spraysd; U-unsprayed.

125

54
Chemical treatments'on perimeter plots (PS) did not increase yields to
the extent where the net return/h i-ias comparable to that of PU.
.<;tand density. The differential decline in alfalfa stand during this
study is depicted in Table 4. Stand densities taken in October 1978 at
termination of the project were greatly reduced from those taken at the
beginning of the study in July 1977. Stem densities followed the
pattern previously defined for alfalfa stem height, weight, and yield
namely CS>PS>CU>PU.
Discussion
The results of this study indicate that the retention of uncut
alfalfa perimeters as a refuge for natural enemy conservation and
buildup suppressed ABL larval populations, but promoted a significant
increase in the numbers of other alfalfa insect pests and an overall
deleterious effect on yield and stand quality.
Uncut alfalfa perimeters seemed to attract insect migrants and ABL
adults which had begun to emerge V7hen the cut alfalfa was at minimal
growth. This may partly explain why there were significantly more ABL
flies in the perimeter unsprayed plots than in the total cut unsprayed
plots in 1978. Conversely, a significantly greater number of blotch
mines were recorded in CU plots than in the PU plots in 1978. This may
be attributed to the occurrence of greater rates of ABL larval
mortality caused by parasitism and predation as was found with 3rd
instar larvae in the P fields in 1978. The existence of a large ABL

55
TABLE 4
Alfalfa stand dansitias of axparlinental research plots. Franklin
Co., MA. P=perimeter; C=total cut; S=sprayed; U=unsprayed.
Treatment Number of stems/sq. m
July 1977
Oct. 1973
PU 318 96
PS 315 184
CU 349 152
CS 334 215

56
parasitoid complex (Hendrickson and Barth 1979) may be a factor in the
reduction of ABL populations, particularly since the most abundant
parasite, Diglyphus intermedius (Girault), kills additional larvae by
probing without oviposition.
Studies on Liriomyza spp. leafminers in California showed that
infestations were less severe in strip cut fields than in completely
cut ones (Jensen and Koehler 1979). Predation was indicated as a key
factor in reducing leafminer infestations. The high correlation of
Lygus spp. with third instar larvae in this experiment may lend support
to the possible role mature plant bugs play as falcultative predators
of inactive leafminers (Wheeler 1976; 1977). Other factors may be
involved in explaining the difference in ABL larval numbers.
The proliferation of plant growth and significant (P=.001)
increases in TLA in the total cut fields may have hindered the
searching effectiveness of predators. This could result in the greater
survival of ABL larvae, thus, more blotch mines in the CU plots.
Unless the retention of perimeters was specific to the buildup of
natural enemies of A. frontella it would seem that the other herbivores
would also have been subjected to more intensive predation and
parasitization. However, this did not appear to have occurred in this
study and it can only be speculated that crop border retention
maintained and promoted pest populations to levels such that natural
enemy action could not inhibit them. This agrees with earlier reports
by authors on the effects of modified harvesting practices. Summers
(1976) reported 62^ more Lygus spp. collected from fields where 3.1ra
wide uncut strips every 50m were left compared to a solid cut field.

57
In addition, Schlinger and Dietrick (1960) found 79.5^ of the captured
insects from alternate strip cut and solid cut fields v/ere from the
strip cut fields. Simonet and Pienkowski (1979) found that when leafy
material or uncut stems remained in poorly harvested fields, field
survival of potato leafhopper nymphs was higher and contributed to the
insect*s increase during alfalfa re-growth. These authors did not
define the detrimental effects increased pest populations had on
alfalfa hay production.
The PU plots which maintained significantly higher populations of
herbivores in this study produced significantly shorter alfalfa which
weighed less than the alfalfa in the CU plots. The same difference in
crop quality was also observed between the PS and CS plots despite
insecticidal applications. Crop growth parameters measured at first
harvest, 1978, indicate that insect feeding on alfalfa, one year may
affect alfalfa re-growth the next year. The response of alfalfa growth
at first harvest, 1978, prior to any treatments for that year, was
similar to the response of alfalfa growth in 1977 which was subjected
to different insect damage levels due to the different treatements.
Leath and Byers (1977) reported potato leafhopper and aphid feeding one
year affected alfalfa production the next.
Position effect was partly precluded when the findings of soil
analyses was similar for all test plots. However, other unmeasured
soil and environmental factors specific to location, i.e. soil
moisture, may have had a favorable impact on alfalfa production in the
C fields.
The application of malathion plus methoxychlor proved to be

58
effective against non-mobile pests. Indirectly, through herbivore
control, spraying was believed to have improved alfalfa crop growth in
the S plots compared to the U plots within the P and C treated fields.
Myers et al (1974) similarly reported increased alfalfa growth through
the use of specific insecticides in the field. Effect of chemicals
directly or indirectly on the physiological growth of alfalfa (Leigh
1963, Chapman and Allen 1948) is a lesser, but possible factor for
increased stem growth. Insecticidal applications on alfalfa in the
perimeter field (PS) was required to produce alfalfa of comparable
height and yield to that of unsprayed alfalfa in the total cut field.
Spraying was economical only in 1978 (fig. 10), but the results also
indicate that insectical applications in the P plots were not
sufficient to overcome the deleterious effects and cost of the retained
perimeter
The differential decline in alfalfa stand during the study
suggests insects play a major role both directly and indirectly in the
longevity of alfalfa in addition to diseases and climate. Reports of
the effects of insect feeding on decreasing cold hardiness of alfalfa
(Harper and Freyman 1979; Hower and Muka 1976), and predisposing plants
to root rots (Leath and Byers 1977) comply with our results. Such
typical stand reductions which growers normally attribute to winterkill
may actually be the result of cumulative insect and pathogen stress
factors. Our stand density data indicate that insecticidal treatments
and complete cutting of fields may not only produce superior yielding
alfalfa, but may also maintain a denser alfalfa stand compared to no
chemical treatment and retaining unharvested field portions.

Bibliography
Andaloro, J. T., T. M. Peters, and A. J. Alicandro. 1978. Correlation of common plant taxa phenologies to development of Agromyza frontella (Rondani) (Diptera: Agromyzidae). J. N. Y. Entomol. Soc. 86:276.
Anon. 1972. Alfalfa blotch leafminer situation in eastern United States. Coop. Econ. Insect Report. 22:132.
Arnold, C. Y. I960. Maximum-minimum temperatures as a basis for computing heat units. Am. Soc. Hort. Science. 76:682-92.
Bereza, K. 1977. Alfalfa blotch leafminer. Ontario (Canada) Factsheet No. 77-067. 2pp.
Bollow, H. 1955. Ein Massenauftreten der minierfliege Agromyza frontella (Rondani) an Luzerne. Pflanzenschutz. 7:141-43.
Byers, R. A. and K. Valley. slf3-lfa blotch leafminer. Insects Conf., Purdue Univ p40-43.
1978. An economic injury level for the Proc. 15th Ann. N. E. Inv. Alfalfa
., W. Lafayette, IN, Oct 17-18, 1978.
59

Callahan, R. A., F. R. Holbrook, and F. R. Shaw. 1966. A comparison of sweeping and vacuum collecting certain insects affecting forage crops. J. Econ. Entomol. 59:478-81.
Chapman, R. K. and T. C. Allen. 1948. Stimulation and suppression of some vegetable plant by DDT. J. Econ. Entomol. 4l;6l6-23.
Cothran, W. R. and C. G. Summers. 1972. Sampling for the Egyptian alfalfa weevil: a comment on the sweep net method. J. Econ Entomol. 65:689-91.
Cothran, W. R., C. G. Summers, and C. E. Franti. 1975. the Egyptian alfalfa weevil: Comparison of 2 standard techniques. J. Econ. Entomol. 68:563-4.
Sampling for sweep net
Durseau, L-. Jr. and D. Jeandel. 1977. Alfalfa blotch leafminer (Diptera: Agromyzidae) laboratory studies of biology in Europe Proc. Entomol. Soc. Wash. 79:259-65. urope.
Edwards, C. R., R. i. Abrams, and P. Sutton. 1978, Extension
1 T prog.„ 1,
Lafayette, IN. Oct! 17-^8,^197r p?9-22.^“'’‘^"®
Harcourt, D. G. 1973. Agromyza frontella (Rnndanil (Diptera: Entomol. Soc.

61
Harper, A. M. and S. Freyman. 1979. Effect of the pea aphid Acyrthosiphon pisum (Homoptera: Aphididae) on cold-hardiness of alfalfa. Can. Entomol. 111:635-36.
Helgesen, R. G. and F. Baxendale. 1978. Feeding activity of the alfalfa blotch leafminer, Agromyza frontella (Rondani) (Diptera: Agromyzidae). J. N. Y. Entomol. Soc. 86:274. Abstract.
Hendrickson, R. M. and S. E. Barth. 1978a. Notes on the biology of ^^Slyphus intermedius (Hymenoptera: Eulophidae), a parasite of the alfalfa blotch leafminer, Agromyza frontella. Proc. Entomol. Soc. Wash. 80:210-15.
Hendrickson, R. M., Jr., and S. E. Barth. 1978b. Biology of the Blotch Leafminer. Ann. Entomology. Soc. Am. 71 :295-8.
Hendrickson, R. M., Jr., and S. E. Barth. 1979. Effectiveness of the native parasites against Agromyza frontella (Rondani) (Diptera-
87^85-S!^^^’ P®st of alfalfa. J. N. Y. Entomol.’ Soc.
Horwitz, W. (ed.). 1970. Official Methods of Analysis of the AssociatioOpOf Agricultural Chemists. 11th ed. p. 16. Washington,
“°a?f;cung’at?fif; as factors affecting alfalfa stand persistence. Rep. 25th Alfalfa Improvement Conference. ARS-NC-52. p88-95. J-mprovernent

62
Jensen, G. L. and C. S. Koehler. 1970. Seasonal and distribution abundance and parasites of leafminers of alfalfa in California. J. Econ. Entomol. 63:1623-28.
Kim, K. C. 1975. Annual summary. Alfalfa blotch leafminer. Penn. Coop. Econ. Insect Rep. 22:132.
Lyford, S. J. 1979. Corn silage versus alfalfa. Dairy Digest, Publication No. USPS 915-480, Coop. Ext. Serv., University of Massachusetts.
Death, K. T. and R. A. Byers. 1977. Interaction of Fusarium root rot with pea aphid and potato leafhopper feeding on forage legumes. Phytopath. 67:226-29.
Leigh, T. F. 1963. The influence of two systemic organophosphates on growth, fruiting and yield of cotton in Calif. J. Econ. Entomol. 56:517-22.
MacCollom, G. B., G. Baumann, and J. G. Welch. I98O. Effect of Agromyza frontella (Rondani) (Diptera: Agromyzidae) on alfalfa Q^^^lity and yield. J. N. Y. Entomol. Soc. 88:56—7. Abstract.
Mellors, W. K. 1979. User’s Manual For: MINERSIM, A computer Simulation Model For The Alfalfa Blotch Leafminer. Cornell Univ Ithaca, N. Y. 28pp.

63
Mellors, W. K. and R. G. Helgesen. 1978. Developmental rates for the alfalfa blotch leafminer Agromyza frontella, at constant temperatures. Ann. Entomol. Soc. Am. 711886-88.
Miller, D. E. and G. L. Jensen. 1970. Agromyzid alfalfa leafminer and their parasites in Massachusetts. J. Econ. Entomol. 63:1337-38.
Miller, D. E. and F. R. Shaw. 1969. Forage crop insects in Massachsuetts. Coop. Econ. Insect Report. 19:849-50.
Pinter, P. J., Jr., N. F. Hadley, and J. H. Lindsay. 1975. Alfalfa crop micrometeorology and it*s relation to insect pest biology and control. Env. Entomol. 4:153-62.
Pruess, K. P., K. M. Lai Saxena, and S. Koinzan. 1977. Quantitative estimation of alfalfa insect populations by removal sweeping. Environ. Entomol. 6:705-8.
Richard, C. and C. Gagnon. 1976. Pertes dues aux maladies chez la luzerne au Quebec en 1975. Canadian plant disease survey. 56:82-4.
Robinson, C. D. and M. A. Massengale. 1967. Use of area weight relationship to estimate leaf area in alfalfa. Crop Sci. 7:394-5.

64
Schlinger, E. I. and E. J. Dietrick. I960. Biological control of insect pest aided by strip-farming alfalfa in experimental program. Calif. Agr. 14:8-9.
Simonet, D. E. and R. L. Pienkowski. 1979. Impact of alfalfa harvest on potato leafhopper populations with emphasis on nymphal survival. J. Econ. Entomol. 72:428-31.
Southwood, T. R. E. 1966. Ecological Methods With Particular Reference To The Study Of Insect Populations. Methuen and Co. London. 39 Ip.
Spencer, K. A. 1973. Agromyzidae (Diptera) of Economic Importance In Series Entomologica, vol. 9, E. Schimitschek (ed.), W. Junk, The Hague, 418pp.
Stern, V. M., R. Vanden Bosch, and T. F. Leigh. 1964. Strip cutting alfalfa for Lygus bug control. Calif. Ag. 18:4-6.
Steyskal, G. C. 1972. The taxonomy of the alfalfa blotch leafminer, Agromyza frontella (Rondani). USDA Coop. Econ. Insect Rep. 222:134-7.
Summers, C. G. 1976. Population fluctuations of selected arthropods in alfalfa: Influence of two harvesting practices. Environ. Entomol. 5:103-110.

65
Vanden Bosch, R., and V. M. Stern. 1969. The effect of harvesting practices on insect populations in alfalfa. Proc. Tall Timbers Conf. Ecol. Anim. Control by Habitat Manage. 1:47-51.
Wheeler, A. G., Jr. 1976. Lygus bugs as falcultative predators. Proc. Lygus Bug: Host plant interaction workshop. Aug. 21, 1976. Univ. Press of Idaho, Moscow.
Wheeler, A. G., Jr. 1977. Studies on the arthropod fauna of alfalfa. VII Predaceous insects. Can. Entomol. 109:423-27.

• 4 ^
1H< i,*
:vr
APPENDIX
r^'*'

67.
Fig. 12. Seasonal life history of the alfalfa blotch leafminer,
Agromyza frontella on harvested alfalfa in Hampshire and Franklin
counties, Massachusetts 1976-1978. (P=pupae, A=Adults, E=eggs,
L=larvae)

68
overwintering P j
GEN I
GEN II
GEN III
GEN IV
A E L
P partial diapause? A L HZL.
P porfiol diapause?
1 L
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
I

69
distribution of A» frontella. Gggs. In 1976 observations were
made on ^ frontella*s propensity to lay eggs vertically along the
alfalfa stem. In Fig. 13 are egg counts/third of stem taken during the
third harvest period in Hampshire and Franklin county alfalfa fields.
They indicate that a larger percentage of the eggs were consistently
deposited within the top two thirds of the plant with a special
preference to the top third. A large number of eggs were laid into new
unfolded leaflets located near the apical meristem. This behavior
supports the appearance of large numbers of flies in the upper canopy.
In Northfield and Amherst fields egg populations were increasing
in the middle third plant section while decreasing or remaining the
same in the top third section toward the end of the growing period.
This may be due to the fact that after formation of terminal flower bud
later in the harvest period more growth is from lateral buds, and thus
these may be more attractive to gravid females.
Oviposition in new leaves appears to be an adaptive survival trait
evolved by the leafminer to insure optimal nutrition for the larva and
liklihood of the leaf staying intact long enough for larval
ae,.lop.,„t. Old loodted od th. botlo. ihlrd of the pi.„t
senesce and fall from the plant prior to larval completion. In
addition, suboptimal nutrient levels in the old leaflets may prolong
time to larval maturity thus increasing risk of larva
leaflet at time of its abcission from the leaf.
s presence in the

70
Fig. 13. Spatial distribution of frontella eggs at specific
alfalfa stem heights in Hampshire (Amherst) and Franklin (Northfield,
Deerfield) counties, Massachusetts commercial fields in 1976 during
third harvest periods. For each sample mean n=10.

Mea
n N
o.
Eggs/
3rd o
f S
tem
71
I
I

72
Fig, 14. Seasonal occurrence of A. frontella and crop ^"development
under a 4 harvest regime. Conway, Franklin Co., Mass., 1977. For
sample means n=20.

Mea
n N
o. M
ines
/Ste
m
73
-190
> Q. C
(/>
CO
(D <0
TD
I
Mean N
o.

74
Figs. 15-19- Scanning electron microscopy. The Agromyza
frontella female utilizes her ovipositor for the dual purposes of
oviposition and production of feeding punctures, (pinholing). The
latter are special holes bored into the underside of the leaflet that
produces an exuded sap which the female laps up for nourishment.
Rarely does the hole break the epidermis of the upper side of the
leaflet. Fig. 15 depicts a dorsolateral view of the female’s
post-abdominal structure in the stage believed to be utilized in the
production of feeding punctures. The ovipositor has numerous small
chitinous rasping teeth x^hich aid in lacerating the side of the hole
produced in the leaflet tissue thus allowing for maximal juice flow.
The sculptured sheath also gives the ovipositor rigidity.
The ovipositor is normally held telescoped inside the abdomen
(Fig. 16). During the preceding oviposition and pinholing the tube is
everted (Figs. 17> 18), until at full expansion, the egg laying
apparatus comes into view. Bollow (1955) observed that during
oviposition the ovipositor is placed vertically on the leaf and bores
with turning movements into the epidermis and underlying parenchyma.
As the ovipositor reaches the parenchyma, it is bent over at a right
angle, and the hols is lengthened horizontally so that the egg can be
laid into the parenchymal layer of the leaf as depicted in Fig. I9.
The female is able to remove her ovipositor keeping the membraneous
leaf epidermis intact thus preventing dessication. This is probably
accomplished by the female turning her ovipositor sideways. The
lateral surfaces of the ovipositor have few chitinous teeth* (Figure 16)
which enable it to brush against the top of the epidermis without
breaking it.

75
Fig. 15. Dorsolateral view of (240x) partially everted female
Agromyza frontella (Rondani) post abdominal structure. This stage of
ovipositor expansion is utilized by the female for producing feeding
punctures (pinholing) in alfalfa leaflets.

- niW
;

77
Fig. 16. Dorsolateral view (590x) of A. frontella overpositor at
early stage of eversion.


79'
Fig. 17. Dorsolateral view (570x) of A. frontella ovipositor at
intermediate stage of eversion.


81
Fig
(470x).
18. Nearly complete eversion of A. frontella ovipositor
Egg laying structure with receptors are becoming visible.


83
Fig. 19. Lateral view (480x) of completely everted A.
ovipositor showing elongated egg laying structure on right.
)
frontella


Fig. 20. Oviposition puncture on underside of alfalfa leaflet.
Separation of the leaf cuticle from the lower epidermis occurs during
egg placement in the spongy mesophyll. Egg (not visible) is in upper
left hand corner of picture obscured by the lower epidermal cell layer.


87
Fig. 21. Comparison of 1978 peak mean (+/- standard deviation)
dead ABL third instars/stem within unsprayed (U) plots between
perimeter (P) and total cut (C) fields just prior to each harvest in
Franklin Co., Massachusetts. For each sample mean n=20.

88
si3©IS / sjeisuf
o
VI-
12
VII
-18
VII
I-30
X-1
3
1978
Sam
pli
ng
Dat
e

89
Fig. 22. Comparison of mean alfalfa heights from sprayed (S) and
unsprayed (U) plots between perimeter (P) and total cut (C) fields at
second, third, and fourth harvest periods in 1977 in Franklin Co.,
Massachusetts.
If means differ significantly from each other at a particularly
date they are indicated by 95^ confidence intervals (according to
Scheffe’s multiple comparison test). S above arrow indicates
insecticide application. For each sample mean, N=100.

90
T-^-1-1-1-^-1 I I r
»W=hioi
o • □ Ea II II 11 II
O) 3 U) Ou CL o O ED
t/v
HX 1S3A2IVH
to-
to¬
rn i/i-
-O)
O) CM
-lS3AliVH-
to-►
to-►
„L o (O
J_I_L_i J_L
CO CM
(O
CO CM
-to
o CO
U) CO CM
(IU011H013H NV31^
Jun
e
July
Au
g.
Sept.
Oct.
SA
MP
LIN
G
DA
TE
S-1
977

91
Fig. 23. Comparison of alfalfa heights from sprayed (S) and
unsprayed (U) plots between perimeter (P) and total cut (C) fields at
first, second, third, and fourth harvest periods in 1978 in Franklin
Co., Massachusetts.
If means differ significantly from each other at a particular date
they are indicated by 95^ confidence intervals (according to Scheffe’s
multiple comparison test). S above arrow indicated insecticide
application. For each sample mean, N=100.
/
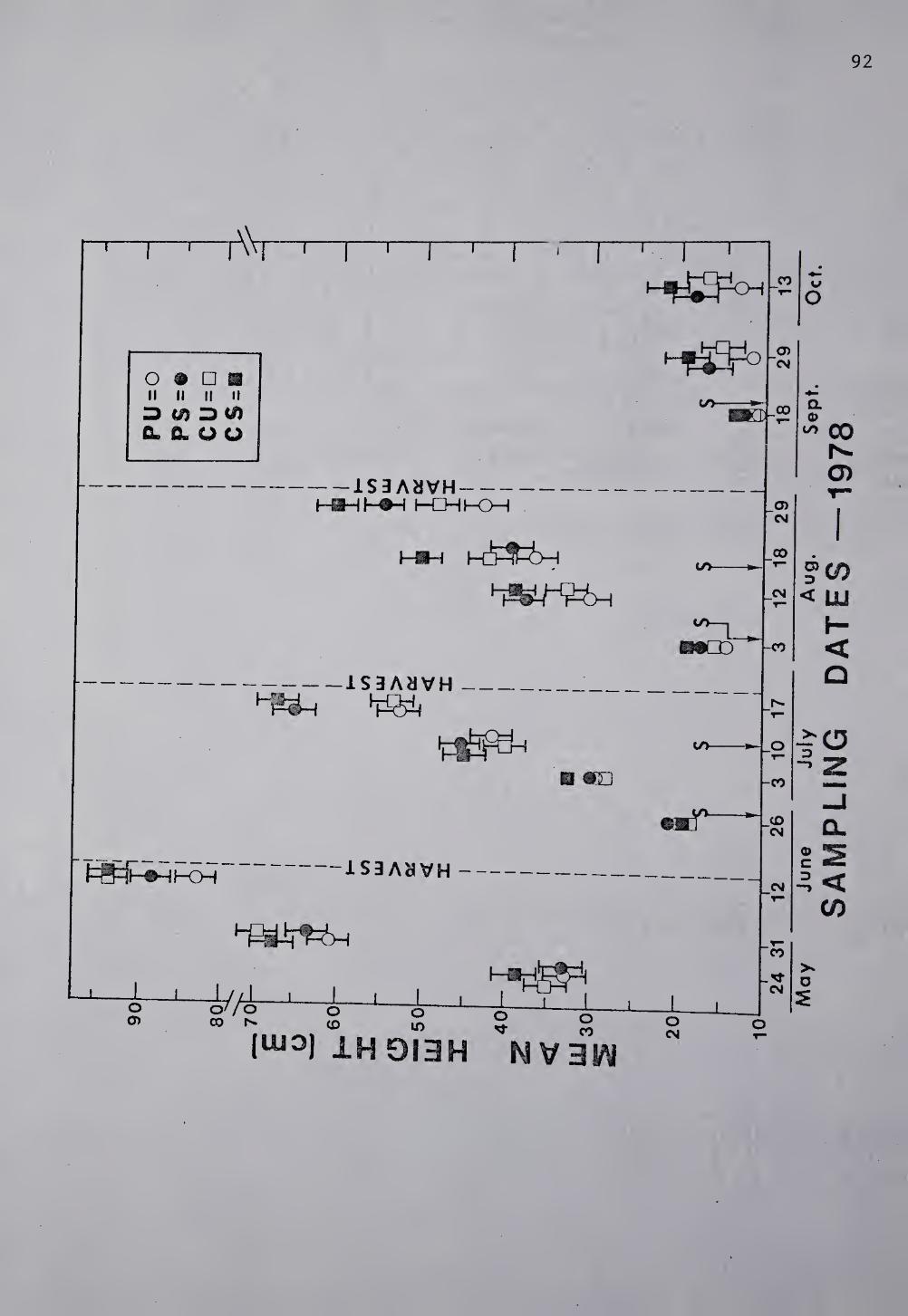
92

93
Alfalfa leaf weight. Oven dried leaf weights/25 stems were obtained
from stems in Figs, 8, 9, and 10, (Chapter 3), at all sampling dates.
Leaf samples were taken to measure canopy growth parameters and
approximate leaf area. Leaf weights averaged over all 7 harvests
indicate relationships similar to those that existed among the
previously mentioned stem heights and weights. The 2 year overall
average leaf weights/25 stems were 6.95, 8.13, 8.30 and 9.^1 grams from
the PU, PS, CU, and CS plots respectively.
Alfalfa leaves constituted an overall average of 41 percent of the
total stem dry weight at harvest Table 6. Variation in percent leaf
weight at different harvests ranged from 26 percent (June 1978) when
alfalfa v/as approximately 90cm tall to 52 percent (Oct 1978) when
alfalfa was approximately 17cm tall. Leaf weight/stem weight ratio was
found to be inversely proportionate to stem height which supports
similar findings by Bula and Hintz (1978). Therefore, taller alfalfa
will yield a greater percentage of stem weight rather than leaf weight.
This is of importance for nutritional purposes since 75^ of the
^^^^l^ble protein in the alfalfa plant is present in the leaves. Late
cutting of first harvest alfalfa, when it achieves great heights, would
result in high yields of plant matter but a proportionally lower yield
in protein content than that which would have been attained at an
earlier cutting.

94
m
w
pq < H
< (-J H
cd 0)
cd
4-1 td 0)
ro #>
CN'
< CJ 4J
O
cn
0
Cd
CXl Od
<
H
4-1 :2
44 Cd 0
h-3
F—4 00 CO o 00 VO VO CO CO
m 4d- crv vO CO
cd a o o o
VO f-H • • • • • CM o o •
f-4 .-4 + 1 +1 +1 o + 1 + 1 VO o o +1
1-H • • • • • <r o •
00 1^ m
vO CM
•
vD
i~H
cr>
o f~l
Cd u f—1
cd 4J c 44 1-^ 'd' r—t O CO O •H O < lO cr» CTv CO VO 4-1 H 'O •
0 H 'cr 'd- ov vO CM 'O C >. c cd cd Cd 4i >4 ,o cd rO
cx 0 rO r-4 Cd rO rO CO • m VO • 00 CO
CO c c:: 44 • • CM • • • B •H IS r-4 o + 1 o o o 0 44 + 1 +1 O +1 41 41 4-» CO Cd 00 CM • CO O CO 44 0 • • lO • • •
o VO t-4 crv 'd- m r-^ CM a
'd" r-H o o crv 00 4J CM CM CO CO CO
Nw^ 3 • • • • • • O CO H m CO CJV CO VO CM
CO 4-> T)
Cd 0 o cd 00 44 cd lO rO rO rO
•H O cd • m o • O CO 0 44 >4 44 • • .-4 • • • IS cx IS o + 1 rH o
'3 Ph CO 44 +1 +1 OV 41 41 fl 44 C cd (TV VO • CO CO VD cd Cd u 0 • • CO • • • 0 0 lo 00 o\ CO
iH 4i 44 0 0
Td 4J B r-H r—1 00 00 VO 0 0 •H <C CO 00 .-4 00
•H B >4 hXl • V4 •H 0 CO H CO 00 'd' >d- 1-4
4i p4 0 xi cd
c Cl. 0 0 cd 00 0 cd cd 0 • r-4 CTV • o CO > 00 cd 44 • • .-4 • • • o c >4 IS 1-4 o 41 o t—1 o
o P- 44 + 1 +1 kT 41 41 41 44 B CO 0 lO VO • CO t-4 00 O cd C 0 • • CO • • •
VO CO vO CM
cn
m
o CO
00
a\ CO
m
CO f—I
• 00
o vD
'd-
m CTv
VO
o w w e
•H 0 Cd CO a e o u
<
<1- 44 cn 0 CO > 0 >4 44
Cd Q
M >
00
CM
M >
00
I—t CO M M r-l
M M \ > > X
c Cd 0
•
c o
•H 4-1 Cd
•H > 0
nd 'O 4i cd
■o C 0 4-» CO
+1
CO 0 4-» Cd CJ
•H I—I
(X 0 u
44 O
CO c cd 0 B
0 >4 cd
CO >4 0
JOt B o c
•
4J 4->
C CO cd 0 O 4-)
•H 44 0 •H 00 C C 00 cd
•H J4 CO
0 cd >~i
a 0 ♦H ♦ 44 44 W Cd r—l 44 CJ P W
*H B • 0 *3 . >
►> ►— r-H Pi •H 0 3
C crv rC CO t-H 3 CO 3 B — >> 3 P 3 3 CO
I—1 3 o O 3 o 3 •3 3
3 3 3 0 Q 3
rC 3 44 O U 3
44 0 CM CO rH CO 00 44 44 o 3 P 3 54 •H PQ u 3 V—^ cd U
O C44 CTt CO O 3 rH u O 0 0 3 rH 3 44 •H 44 iH 35 0 0 0 0
rH > •H U 0 3 •H
44 iH 3 IS C 44 0 CO 3 54 • 0 to 0 O > rH
C44 O 3 C44 v O •fH 00 MO PH 3
CM 0 0 B rl*J
rO r3 u 3 44 CO 44
•3 VD 0 44 vO 0
3 54 O II 0
1—1 0 «—1 O 3 o 3 0 W
C44 0 yi 44 54 3 IS
CO 0 3 C44 C44 C44 3 144 3 3 0 •H 0 0
1^ 3 kJ
CM CO >;r

TABLE 6
Comparison of leaf weights as a percent of total stem weights among culturally and chemically treated experimental plots in
Franklin Co.,
Harvest Dates
Perimeter(P) Unsprayed(U) Sprayed(S)
Total Cut(C) Unsprayed(U) Sprayed(S)
VII-14-77 40 39 40 39
VIII-16-77 46 43 45 44
VI-12-78 27 28 25 26
VII-17-78 37 36 39 37
VIII-29-78 47 43 47 40
X-13-78 56 51 50 51
Mean 42 40 41 40
Each number is the mean % leaf weight of 4 replicates.
% leaf weight = leaf weight(g) ^ total stem weight (g) .

TABLE 7
Cost/Benefits of alfalfa perimeter retention and chemical control of insect pests based on optimal stand density.
Perimeter(P) Total Cut(C) Unsprayed(U) Sprayed(S) Unsprayed(U) Sprayed(S)
Seasonal Yield kg/ha I977I 6633 7952 7885 9217
$ Value of Hay (3 $88.14/1000kg 584.69 700.98 695.06 812.43
$ Cost/ha for Chem Trt/yr^ — -96.33 — -96.33
Yield Loss/ha/yr Due to Perimeter Retention^ -58.46 -70.09
Actual $ Return/ ha/yr 526.23 534.56 695.06 716.10
Seasonal Yield (kg/ha) 1978'^ 12845 15275 16007 18243
$ Value of Hay @ $88.14/1000kg 1132.24 1346.44 1410.96 1608.07
$ Cost/ha for Chem Trt/yr^ — -80.27 -80.27
Yield Loss/ha/yr Due to Perimeter Retention^ -113.22 -134.64
Actual $ Return/ ha/yr 1019.02 1131.53 1410.96 . 1527.80
Total of 2nd, 3rd, and 4th harvest.
Cost includes labor, machine use and chemicals for 6 insecticide applications.
Equals 10% of total yield/harvest.
Total of all 4 harvests.
Cost includes labor, machine use and chemicals for 5 insecticide applications.
