studies in animal locomotion - home | journal of...
TRANSCRIPT

STUDIES IN ANIMAL LOCOMOTION
I. THE MOVEMENT OF FISH WITH SPECIALREFERENCE TO THE EEL
BY J. GRAY.
King's College, Cambridge.
(From the Laboratory of Experimental Zoology, Cambridge.)
(Received 20th October, 1932.)
(With Four Plates and Eleven Text-figures)
WHEN a body is moving in water it encounters a resistance in the direction of itsmotion, and consequently the body must be supplied with energy if motion is tooccur at a uniform speed. A study of the mechanism of propulsion of a fish fallstherefore into two parts, (1) a study of the forces resisting motion through the water,and (2) a study of the mechanism whereby the fish utilises the energy liberated byits muscles for overcoming the forces of resistance. To some extent these twoaspects of the problem are interdependent and involve considerable hydrodynamicaldifficulties, but in the present paper an attempt will be made to show that the move-ments of a fish's body1 are such as to generate forces capable of opposing the forcesof resistance whatever be the nature or magnitude of the latter. The problem wasattacked two centuries ago by Borelli and by Pettigrew in 1873; since then com-paratively little attention has been devoted to the subject except by Breder (1926),whose results will be considered later.
Since all propellers operate by driving astern a volume of water, the reactionfrom which compensates the surface resistance of the moving object, the initialproblem of the fish's movements consists in demonstrating that the fish moves itsbody in such a way as to drive water away from its surface in a backward direction.All inanimate propellers belong to one of three types: (1) The jet propellers—asexemplified by all reaction turbines which project a current of water from a nozzle.The reaction caused by the water moves the nozzle in a direction opposite to that ofthe movements of the water. (2) Paddles—whereby a backward thrust is exerted onthe water, parallel to the direction of motion of the paddle and at right angles to thesurface of the paddle. The paddle can only be submerged during one-half of itscomplete movement, or it must be capable of being rotated about an axis at rightangles to its line of motion, in order that no appreciable thrust is exerted during theperiod which follows the effective phase of the movement. (3) Screws—the theory
1 The present paper deals only with the propulsive properties of the bodies of a selected number offish whose appendages play little or no part in the propulsion when the fish are moving at reasonablespeeds. The propulsive properties of the caudal fin will be considered in a subsequent paper.

Studies in Animal Locomotion 89
of screw propulsion is essentially that of an inclined plate—which, by motionthrough the water, generates a force at right angles to its surface (see Fig. 1). Thisforce (P) has a component (T) at right angles to the direction of motion of the plate,which tends to move the plate along a line at right angles to its original direction ofmotion.
The mechanism of propulsion of a typical fish does not conform to the design ofa jet or a paddle, and since all screws operate by means of a true rotary movement,the possibility of a screw is, at first sight, excluded. The object of this paper is toconsider the motion of a fish's body and to compare the underlying mechanism withthat of a typical screw propeller. During the whole of the work, an attempt has beenmade to record the form and position of the fish at known intervals of time by photo-
Fig. 1. AB is a cross-section of the blade of a screw moving along cd in the direction of the largearrow. A force P is generated at right angles to AB. This has a component T which tends to move thescrew in the direction of a.
graphic means. An experimental tank was set up in the field of a timed cinemato-graph camera, so that the position of the fish could be determined by means of agraduated field placed immediately underneath the fish. The method of recording theinterval between successive photographs has been described elsewhere (Gray, 1930).
I am greatly indebted to Mr J. E. Harris for his valuable help in the preparationof these photographic records.
I. OBSERVATIONS ON THE MOVEMENTS OF FISH.
As observed by the human eye, the motions of various types of fish appear tovary considerably from one species to another. The most conspicuous features of amoving eel—as are seen in the photographs taken by Marey (1894)—are the wavesof curvature which pass along the length of the body from head to tail. In thedogfish (and, still more, the mackerel and whiting), the presence of such waves isless obvious, and the visible movements appear to be due to transverse strokes

90 J. GRAY
executed by the posterior end of the body across the axis of motion. It can be seenfrom the photographs reproduced in Figs. 2-10 (Pis. I—IV), however, that in all thesecases waves of curvature pass along the body alternately on the two sides, but that theydiffer in the various fish in certain important characteristics. Firstly, their speed ofpropagation along the body varies greatly. In the examples illustrated the approxi-mate speeds of the waves and the rates of movement of the fish are as shown inTable I. Secondly, the form of the waves differs. In the reversing eel (Fig. 10) theamplitude of the waves is very large, and is of approximately the same value astheir wave-length. In Ammodytes (Fig. 7) and the mackerel (Fig. 5), the relativeamplitude is very much smaller, while the dogfish, glass-eel, butterfish, androckling occupy intermediate positions. Thirdly, when the fish are swimming at asteady rate, the frequency of the waves per second varies in the different species. Inthe examples illustrated, the approximate number of waves passing down each sideof the body are shown in Table II. Fourthly, the amplitude of the waves is alwaysgreatest at the posterior end of the body, but the variation between the amplitude ofthe head and tail varies very greatly in different types. In the small eel the amplitudeof the movements of the head is relatively very much greater than those of themackerel or whiting.
Table I.
Gla9s eel (Anguilla vulgaris)Butterfish (Centronotus gunnellus)Whiting (Gadus merlangtit)Dogfish (Acantlrias vulgaris)Mackerel (Scot/tber tcombrus)Ammodytes (A. lanceolatut)
Velocity of wavecm. per sec.
6-2I7-S25-055771 6 0
Velocity ocm. per
4-01 1 71 6 82942-5
8 0
Table II.
Glass-eelButterfishWhitingDogfishMackerelAmmodytes
Waves per min.
931 2 01 2 0
541701 2 0
The movement of the muscular waves along an eel's body was recordedphotographically by Marey (1894), who made no attempt to define the mechanicalprinciples which are responsible for the forward movement of the whole fish. Theseprinciples have been considered by Breder (1926), whose description of "anguilli-form" movement is as follows: "The forward motion is certainly attained by thepressure of the fish's body against the water in the following manner. The mechani-cal forces brought to bear on the water are diagonally backwards (from the posteriorsurfaces of each of the curves of the body). As these are distributed symetricallyabout the line of progression, a forward resultant of reaction follows, for pressure

Studies in Animal Locomotion 91
from a moving plane is always at right angles to its surface." That the fish's bodyexerts a pressure on the water at right angles to its own surface is in accordance withthe analysis given later in this paper, but Breder goes on to state that " It might beobjected that as the eel is moving ahead there is likewise adverse pressure diagonallyforward from the anterior sides of these backwardly moving waves. The truth ofthis is evident and it simply makes it necessary for the fish to pass these wavesposteriorly at a rate considerably faster than it expects to move forward Thespeed of the waves moving backward must exceed that of the forward motion of theanimal as a whole. If the two speeds just equalled each other it would mean that anypoint on a wave such as its crest would be stationary with reference to the sea-bottom; but as one is dependent on the other this is obviously impossible." Themechanical principles involved by this explanation are by no means clear, for itis certain that the propulsive thrust of the moving body is due to the fact thateach part of the body is executing a series of transverse movements. Althoughthese movements can be expressed in terms of longitudinally moving waves ofcontraction, the principles of propulsion of a fish are much more readily derivedfrom a study of the transverse movements of each section of the body than froma direct investigation of the propagated waves of contraction. In the present paperan attempt will be made to investigate the propulsive effect of those transversemovements which are induced in the various parts of the body by a series ofmuscular contractions which are of such a nature as to produce the phenomenon ofa propagated wave.
The movements of the body can be considered in two ways. Firstly, it is possible,from a series of instantaneous photographs taken at equal and known intervals oftime, to plot the position in space of any particular point on the surface of the body.Secondly, it is possible to consider the movements executed by one part of thebody relative to any other part, and not to fixed points in the environment of thefish. By combining these two sets of observations it is possible to form an idea ofthe way in which the contractions of the muscles induce changes in the relativeposition of the parts of the body, one to another, which are such as enable the fish,as a whole, to transmit to the water a backward momentum equal and opposite tothat of the frictional forces which oppose the motion of the fish through the water.Owing to the well-defined nature of its muscular waves, attention may be con-centrated on the small glass-eel (Anguilla vulgaris), about 7 cm. in length, shown inFig. 2; the same type of analysis can be applied to other forms, but for variousreasons it is convenient to defer this until later.
Fig. 12 A-C shows the track of the head, the middle point of the body, and thetip of the tail of an Anguilla whose form during motion is shown in Fig. 12 D. Itwill be noticed that the successive positions of each point lie along a sinusoidal curvewhose " pitch " or wave-length is the same in all cases, namely 3-2 cm., and that thisis less than the "pitch" or wave-length of the waves which characterise the bodyitself, viz. 4-7 cm. It can also be seen that the amplitude (to) of the waves is least atthe head and greatest at the tip of the tail. The axis of motion (ab) of the fish isshown in the figure, and it can be seen that if a line (cd) is drawn at right angles to

92 J. GRAY
this axis at points where the track of the point on the body cuts the axis of motion,then the angle (8°) between the track of the fish and this line cd (the transverse axisof movement) becomes progressively less as the tip of the tail of the fish is ap-proached. If we now examine (Fig. 13) the angle (6m) made by any part of the bodyof the fish and the line cd as this particular part crosses the line ab (i.e. crosses theaxis of forward movement), it can be seen from Fig. 13 that this angle also decreases
Head " Mid Point "Tip of TailA B C D
Fig. 12. A-C. The paths followed by (i) the head, (ii) the middle of the body, (iii) the tip of the tailof a young Anguilla (glass-eel). Note that the amplitude is greatest at the posterior end of the body,and that the wave-length of each track (A) is less than that of the curve of the animal's body (D).
from the head to the tail; since the pitch of the body is greater than the pitch of thecurve of movement, it follows that the angle (8m) made by the body with thetransverse axis (cd) must be greater than the angle (80) between the path of motionand the transverse axis of movement. The difference between these two angles(8m — 8p) is of fundamental importance and will be called the angle of attack; it isdesignated by the symbol a. Similar curves to those shown in Figs. 12 and 13 canbe constructed for other types of fish with similar results except that in most fish theamplitude of the transverse movements of the head are very small compared with

Studies in Animal Locomotion 93those of the tail. It can be seen in Figs. 2-10 that not only are the wave creststravelling along the body of the fish but they are also travelling backwards withreference to the environment.
To define the movements of a point on the body relative to other parts of thebody it is necessary to adopt two fixed axes of reference. One of these is providedby the axis of forward movement (ab), for this is also the axis about which each pointof the body is moving in a transverse direction relative to any other point. It is notso easy to obtain a fixed transverse axis. The ideal procedure would be to plot theposition of each point on the body with reference to a transverse axis which is
< a
Fig. 13. Tracings from enlarged photographs of Anguilla showing that the angle between the bodyand the axis of forward movement (ab) decreases from the anterior to the posterior end of the body.
moving forward with the fish at a velocity equal to the average forward velocity ofthe fish. This can be done within small limits of error if successive photographs areenlarged and then superimposed on each other in such a way that the tip of the headlies along the same transverse axis and if the longitudinal axis of motion (ab) of eachphotograph is superimposed on that of the others. This has been done in Figs. 14and 15, which represent a fish whose waves are moving down the body in the normalway, but whose body is unable to progress forwards. It can be seen that during eachphase of its motion, any given point forms part of a segment1 of the body (Fig. 14)which is inclined with its leading surface (i.e. the surface towards the direction of
1 The term "segment" is not used in its strict morphological sense.

f l J. GRAYtransverse movement) turned towards the hinder end of the body. Thus in Fig. 14the segment XY is travelling from the right side of the axis of movement towards theleft side and its leading surface is facing backwards and towards the left. Con-
\a
Fig- IS-
Fig. 14. Enlarged drawings of a young Anguilla arranged to show the movements of short segmentsof the body during the passage of the complete wave past the segments. Note that the segment XYis travelling from right to left and is directed backwards and to the left. The segment X1Y, is travellingfrom left to right and is directed backward and towards the right. Note also that the tip of the tail ismoving in a figure of 8 curve.Fig. 15 A. Tracings of left side of a butterfish showing the passage of a wave, and the correspondingpositions of the tail (1—7). The dotted line shows the longitudinal axis of motion.Fig .15 B. Shows the relative transverse velocity of the tail at different phases of its motion. Notethat the velocity is greatest when the tail is crossing the axis of forward movement.
versely, the segment X1 Yy shows the corresponding movement of a segment fromleft to right, and the leading surface is facing backwards and towards the right. Itwill be noted that as the segment XY is passing from right to left, the segment formspart of a wave whose crest is travelling down the right side of the body, and that as

Studies in Animal Locomotion 95
X1 yx moves from left to right it is part of a wave travelling down the left side of thebody.
Although the movements executed by each segment of the fish closely resemblethe movements of the blade of an oar when sculled from the back of a boat, thebody of the fish exhibits certain peculiar features of considerable theoreticalimportance.
(1) The speed at which a segment moves along its transverse path is notuniform. When displaced to its maximum extent from the longitudinal axis ofmovement (ab), the segment is moving very slowly; as it crosses the axis of longi-tudinal movement it is travelling at its maximum speed. Fig. 15 B shows the positionof a segment at equal periods of time, and it can be seen that the speed of itsmovement varies inversely with its displacement from the longitudinal axis ofmovement; during the phase of movement towards this axis the segment isaccelerating, and during the phase of movement away from this axis the segmentis decelerating.
(2) During each phase of movement the angle (9) between the body segment andthe longitudinal axis of movement is changing. It is greatest at the extreme positionsand is least as the segment is crossing the line (ab). Towards the end of each phase,the segment is parallel to the axis of longitudinal movement, but as it begins to movetowards this axis, the segment becomes more and more inclined backwards—aftercrossing the axis the process is reversed until the segment again points directlyforward; finally it becomes inclined in the opposite direction as it begins to movebackwards towards the axis (ab). The important point to notice is that the angle 6is least when the segment is crossing the line ab—i.e. when it is travelling at itsmaximum transverse speed.
(3) It is only when the segment is near to the axis ab that its leading surface isbounded by a plane—in all other positions the surface is curved. As the segmentapproaches the axis ab the leading surface is bounded by a curve which is concavetowards the direction of movement; after passing the axis (ab) the leading surface isconvex towards the direction of movement.
(4) If a point is marked on the surface (e.g. the base of the tail-fin in Fig. 15 B), itis found to travel in a figure of 8 curve relative to the head. The transverse axis ofthe figure of 8 is at right angles to the axis ab. The greater the amplitudes of trans-verse movement relative to the pitch or wave-length of the curve of the body, themore marked are the figures of 8 (see footnote, p. 97). The figure of 8 movementcan be expressed in another way, namely, when the whole fish is travelling forwardat a constant average velocity, the forward velocity of segments at the extremepositions of transverse displacement is rather greater than the average velocity ofthe whole fish and the forward velocity of segments which are crossing the axis ofmotion is rather less than this average value.
When the fish is in motion, the path traced out in space by any given point onthe body represents, of course, the locus of a point travelling along a figure of8 curve which is endowed with a forward velocity equal to the average forward speedof the fish. The angle between the body of the fish and its path of motion is of

96 J. GRAY
vital significance when we consider the propulsive properties of the body; the effectof the figure of 8 motion is, as is seen in Fig. 16, to increase this angle when asegment of the body is crossing the axis of longitudinal motion, and to decrease it asit approaches the extreme positions of its transverse displacement.
The figure of 8 motion of the tail of a slowly moving sturgeon was observed byPettigrew (1873), who implied that the movement is a physiological adaptation toefficient propulsion. It can, however, be shown that it is the inevitable result of thepropagation of a wave of curvature along an inextensible body.
12
Fig. 16. If AB is a segment travelling transversely along the dotted line a,^ and endowed with avelocity of 1 cm. per unit of time towards the right, then the track of the mid-point of the segmentis shown by the dotted curve to the right of the figure. If AB travels along the figure of 8 (0,0,0,ft, 6,6,) and is endowed with the same transverse velocity as before, then the track of the mid-point isshown by the full curve on the right of the figure. Note that the angle ot, is greater than the angleat,. The larger the amplitude of the figure of 8 the greater is the increase effected in ot at the mid-point of the transverse movement. [The reverse is the case at the extreme positions.]
Fig. 17 shows an inextensible string along which is passing a wave of curvatureof constant form. If one end of the string be allowed to move along the axis (cd),and the position of any given point on the string be marked in respect to this axisand the longitudinal axis (ab), it can be seen that except at stated intervals (of half-wave-lengths from the constrained end of the string) the path traced out by a fixedpoint along the string is a figure of 8.
The greater be the relative amplitude to the wave-length of the waves the moremarked is the figure of 8 movement. Since the relative amplitude at the tail end of

Studies in Animal Locomotion 97a fish is greater than at any other point, the tail exhibits a more marked figure of8 than does any other point1.
The observed movements of the fish's body can be summarised as follows:(1) All parts of the fish's body which are in transverse motion have their leading
surfaces directed backwards and towards the direction of transverse movement, butthe angle of inclination is most pronounced when the segment is crossing the axis oflongitudinal motion, and at this point the segment of the body is travelling at itsmaximum speed.
(2) Each point on the body is travelling along a figure of 8 curve relative to atransverse line which is moving forward at the average forward velocity of the wholefish. In other words, a segment when moving across the axis of longitudinal motionis travelling backwards relative to a segment which has reached the extremeposition of lateral displacement. The track of any point on the body (relative to theearth) is a sinusoidal curve whose pitch or wave-length is less than that of a curve
1 2 .3 4. 5 6 7 8 9Cr
Fig. 17. The dotted lines show the loci of points situated at i, 5, 10, 175 cm. from the end of aninextensible string along which a sine wave is moving. Note that each point travels on a figure of 8curve unless it is situated at or about one-half of a wave-length from the front end of the string.The larger numerals show successive positions of the crest of wave; the smaller numerals show thecorresponding positions of the selected points.
which defines the body of the fish. There is therefore a definite angle between thesurface of the fish and its path of motion.
The movements executed by the body of a fish are closely analogous to thoseexhibited by a flexible but elastic body, one end of which is made to vibrate along atransverse axis. The validity of this analogy will be discussed elsewhere.
1 It might be objected (see Fig. 17) that if the tail were to lie at a point equal to a multiple of halfa wave-length from the head, it should travel approximately in a straight line. It must be remembered,however, that this is only the case under two purely artificial conditions, (a) when the amplitude ofthe movements is the same along the whole length of the body, (b) when the axis of reference is suchthat the head is moving along a straight transverse line. If it were possible to refer the movements toan axis which is moving forward at the same average forward velocity of the whole fish (i.e. if thepropagation of the wave involved no displacement of the centre of gravity of the system), every pointof the body would appear to travel in a figure of 8, the horizontal amplitude of which would varydirectly with a power of the amplitude of the metachronal wave. If the wave have the form of a sinecurve and the amplitude be small, it can be shown that the longitudinal amplitude of the figure of
8 curve is —r-, where OJ is the amplitude of the sine wave and A is the wave-length.

98 J. GRAY
It now remains to be shown that (i) the movements of the body are such as todrive the fish forwards through the water, and (2) the movements of the body are thedirect effect of a series of waves of muscular contraction which start at the anteriorend of the body and pass backwards towards the tail. The latter consideration willbe discussed in a later paper.
II. THE BODY AS A PROPELLER.
Towards the extreme positions of each transverse cycle the velocity, form andinclination of the body are rapidly changing and although it is possible to see in anempirical way their general propulsive significance, these changes render impossiblean analysis of the effect of a complete cycle on the distribution of the surroundingwater. This distribution may be affected by the acceleration as well as by thevelocity of the body, and for this reason it is convenient to consider in the first instancethe forces acting on a segment as the latter passes the axis of forward movement,i.e. when it is inclined to this axis to its maximum extent, and when the velocity ofits transverse movement is greatest and reasonably constant.
H
Fig. 18 a. Fig. 186.Fig. 18 a. The dotted line shows the locus of the point O on the segment AB when the latter istravelling transversely from left to right. The position and inclination of the segment at variousphases of its movement are shown at 1-7. ab, longitudinal axis of movement, cd transverse axis ofmovement. The fish is depicted as stationary.Fig. 186. Diagram showing the position of the segment AB of a stationary fish in respect to its owndirection of motion and to the longitudinal axis ab. The segment is travelling along OE with avelocity equal to OE. The angle BOE = a, and is the angle of attack. The angle BOd is the angle(6m) at which the segment is inclined to the axis cd. The movement of water relative to the segmentis at first EO; after meeting the body, a finite volume of water flows along OA with a velocity OH.The effect of the water on the body is to endow the latter with a velocity GO relative to the water.

Studies in Animal Locomotion 99
Symbols used.
a = the angle between the surface of the body and its direction of motion.9 = the angle between the surface of the body and a line drawn at right angles to
the longitudinal axis of movement (ab) of the fish.9m = the value of 9 for any particular point on the body when this point is crossing
the axis ab.ab = the longitudinal axis movement of the fish.cd = an axis at right angles to the longitudinal axis of movement, i.e. an axis of
transverse movement.A*= the pitch or wave-length of the muscular waves on the body.
OJ = the amplitude of the muscular waves on the body.P = the pressure normal to the surface of the body.F = the factional force tangential to the surface of the body.T •= the forward component of the force P.D = the transverse component of the force P.X = the transverse component of the force F.Y = the forward component of the force F.V = the velocity of forward movement of the
fish.Vg = the transverse velocity of any given
segment.VR = the resultant velocity of any given seg-
ment and the surrounding water.As long as the fish does not move forward,
the direction of movement (relative to theearth) is to the side and backwards (Fig. 18);but when the fish is travelling forwards thedirection of motion of AB is to the side andforwards. During this movement water isdisplaced, and the water displaced at theleading surface (AB in Fig. 19) must eitherflow along the surface of the body or over thedorsal and under the ventral fins. Since thebody presents a relatively flat surface to thewater, some at least of the water must bedeflected along the surface of the body in abackward direction, i.e. in the direction OA.Let the mass of water so deflected be m gm. ^ Zper sec. If the original velocity of this water axis ab. The segment AB is moving along the(relative to the body) is Frcm. per sec. (= OE, ^ ^ ^ ^ t ?J%( y) p ( ^ ^ ^ ^ t ^ t h e ?J%Fig. 19), this Can be resolved into two com- inclination (0m) is FOd. The normal pressureponents, one (OF) being parallel to the body °f
Dthe ^ J °" t h e w.at
fer is proportional to
r ' v ' 1 , 1 1 OP and the fnctional force is proportionaland the other (EF) normal to the body. to OF-OH.

ioo J. GRAY
After encountering the body, the component EF is lost in respect to m gm. ofwater per sec., hence there has been a loss of normal momentum by the water ofm , EF gm./cm. per sec.: this momentum must be gained by the body and itrepresents the pressure of the water on the body in a direction at right angles toits surface. Now EF = Vr sin a (where a. is the angle between the body andits direction of motion), hence the normal pressure of the body (P) on thewater is m Vr sin a and the component of this force along the axis of longitudinal
A
\1
A/
Jo?'
Jh
K
N
MN
/ t
/ /
a/'
A
A'
N
M
/
/
1A
KN
M
I,
A
N
K
ML 4 L 5 L 6
Fig. 20. Showing the effect of an increase in forward speed on the value of the angle a. r, fishstationary; the forward velocity of the segment has a negative value of MN. 2, forward velocity offish = backward velocity of segment. 3 and 4, forward velocity giving reduced but positive valuesfor a. 5, forward velocity giving zero value for a. 6, forward velocity giving negative value for a,giving negative thrust. AIN —- forward velocity of segment; LAf -= trans, velocity of segment;MK = the pitch of the segment which is a function of the wave length of the muscular waves;cd = trans, axis of movement.
movement is the corresponding propulsive thrust. As long as the body moves at anangle to its own path of motion, there must therefore be a tendency for the fish tomove bodily through the water. As soon as the fish begins to move, however, twoevents occur: (i) the angle a diminishes, and (2) frictional forces are generated at itssurface. The diminution of the angle a is seen in Fig. 20, where it can be seen thatthere is a value for the rate of forward progression which is such that a = 0°, and atthis point the propulsive thrust must be zero; at the moment it may be noted thatthe faster is the forward speed of the fish the smaller is the angle of attack and thesmaller is the normal pressure of the fish against the water.

Studies in Animal Locomotion 101
As long as the fish is in motion, its movements will be resisted by factionalforces which are due to the disturbances set up in the water in the neighbourhoodof the body. These forces act along all surfaces and their direction is tangential tothe direction of motion of the surface, so that when water is moving past the bodyof the fish, the velocity of the water is reduced. Thus in Fig. 19 if FO be therelative tangential velocity of m gm. of water struck by the fish per sec., and if OHbe the relative velocity of this water after passing over the segment, then the waterhas lost momentum equivalent to m (FO-OH) gm./cm. per sec. in the direction ofOH. This momentum is gained by the body and represents the frictional force (F)acting in the same direction. Thus the net effect of moving a segment of the bodythrough the water at an angle to its own direction of motion is to impress on thebody two forces—one normal to the surface (P)1 and the other tangential to thesurface (F). The longitudinal resultant of these two forces represents, if the present
K
analysis is correct, the net propulsive thrust which drives the fish against theresistance of the water.
If forward motion at a uniform velocity is to take place, the resultant forcesacting on the segment of the body when measured in any direction must be zero.The conditions under which this will occur can be seen by resolving the forces Pand F along the longitudinal (ab) and transverse (cd) axes of movement respectively.Thus in Fig. 21 let LM = Vo = the velocity at which the segment AB is travelling,along the' transverse axis of movement. Let MN = V = the velocity of forwardmovement of the fish. Then the segment inclined at an angle 9 to LM is travellingalong LN with a velocity LN = Vr — VFo
a + V2,* and the angle between the body1 The existence of a force normal to the surface of the body, and the reduction of the longitudinal
component produced by an increase in the angle of inclination (0) of the body, was pointed out byBreder (1926).
• This is the flow of the water relative to the body if the water is stationary in respect to theearth.

102 J. GRAY
and its path of motion is a. The force P (which depends on the value of a, seep. ioo) and the force F can both be resolved along ab and cd. The propulsive thrustalong ab is T - Y and the resisting force along cd is D + X. Now if these forces areto have no resultant they must be compensated by equal and opposite forcesoperating on the segment. These latter forces are (i) the force exerted by themuscles, (ii) the resistance exerted by other parts of the body. Whether or not astructure (e.g. the dorsal and ventral fins, the skull, etc.) is exerting a propulsivethrust there will always be a frictional force at its surface, and as such structures maybe moving in any forward direction it follows that the frictional forces can always beresolved into components acting along ab or cd. Uniform motion will ensue, there-fore, when the forces along cd are collectively equal and opposite to the forceexerted by the muscles, and when the forces acting along ab are equal and oppositeto the longitudinal components of all the forces developed by other parts of the fishwhich resist the forward motion of the segment. It follows that when a fish (whichis initially at rest) begins to move its body in such a way that the leading surface isinclined backwards at an angle to the axis (ab) it will move forward with increasingvelocity until the angle (a) between the leading surfaces and their direction ofmotion is reduced to such a value that the net propulsive thrust is exactly equaland opposite to the effect of the frictional forces acting on the body.
It will be remembered that the value of 6 varies for different phases of themovement of a single segment (see Fig. 14) and for different regions of the body(Fig. 14). For regions lying towards the middle of the body of the fish shown inFig. 14 the value of 8m (when the segment is passing the longitudinal axis of move-ment) is about 500. As the segment moves away from this axis the value increasesto 900, and as this increase occurs, the transverse velocity falls. It is clear fromFig. 21 that a rapid fall probably occurs in the thrust and in the work done as thevalue of 8 increases—and for high values, a very weak and very inefficient thrustremains. It has also been shown that the value of 8 varies for different regions alongthe body of the fish. The thrust and therefore the useful work done by a segmentthus depend on its position in the body as well as on the particular phase of its owncycle, so that the total thrust exerted by the whole fish represents the sum of thethrusts exerted by all the segments of the body, all of which have different valuesof 8 and may have different values of a. Before examining these phenomena ingreater detail it is convenient to consider the relationship which exists betweenthe values of the angles 8 and a on the one hand and the form and properties of themuscular waves, which pass from one segment to another, on the other. This will bedone in a subsequent paper.
The analysis given above assumes that the relative velocity of the body and thesurrounding water is the resultant of the transverse velocity of the body and itsforward velocity through the water; in other words, that the water which encountersthe body is at rest relative to the earth. It is unlikely that this condition isstrictly fulfilled, since the anterior regions of a fish such as that of a mackerel mayinfluence the rate of flow of the water past the segments lying more posteriorly; inthis case the velocity with which these segments encounter the water may not be

Studies in Animal Locomotion 103
simply the resultant of the transverse and longitudinal motions of the fish itself. Atpresent there is no means of determining the exact flow of water past the fish, and itis necessary to assume that disturbances of this type are comparatively small.
III. SUMMARY.
1. The waves of muscular contraction which pass along the body of a swimmingeel occur also in other fish. The waves vary greatly in speed of propagation,amplitude and frequency. The speed of propagation of the waves is too low to becontrolled by the rate of conduction of a simple nervous impulse.
2. The movements executed by a localised area on the surface of the body aresuch that each area moves in a direction transverse to the line of forward movement.During these movements the leading surface of the body is inclined backwardstowards the tail and at an angle to the path of motion of the area concerned. Theangle of inclination and the angle made with the path of motion vary with (a) dif-ferent regions of the body, and (b) with different phases in the motion of each region.
3. Each point on the body travels in a horizontal figure of 8 relative to a transverseaxis which is moving forward at the same average velocity as the whole fish. A seg-ment of the body at the mid-point of its transverse motion is travelling forwards at arate slightly less than that of a segment at the extreme position of its transverse move-ments. These movements are the mechanical result of the inextensibility of the body,and they effect significant changes in the angle between the surface of the body andits direction of movement.
4. The movements of each part of the body are shown to be such as to generatea forward thrust which drives the fish forwards against the resistance of the water.The magnitude of the forward thrust depends among other things on (a) the anglewhich the surface of the fish makes with its own path of motion, and (b) on theangle between the surface of the fish and the axis of forward movement of the wholefish, (c) on the velocity of transverse movement of the body.
5. The propulsive properties of each segment of the body are greatest as thesegment is crossing the axis of forward movement.
REFERENCES.BREDER, C. M. (1926). Zoologica, 4, 159.GRAY, J. (1930). Proc. Roy. Soc. B, 107, 313.MAREY, E. J. (1894). he Movement. Paris.PETTIGREW, J. B. (1873). Animal Locomotion. London.
EXPLANATION OF PLATES.
PLATE I.Fig. 2. Successive positions of a young eel (Anguilla vulgaris) (7 cm. long) during a period of 1 sec.The photographs were taken at 009 sec. intervals. The side of each square is 1 in. The passage ofthe muscular waves is marked by black dots and crosses. The dark line represents the pigmentedmid-dorsal line of the transparent animal. Note the well-defined curvature of the body.

104 J- GRAY
Fig. 3. Successive positions of a butterfish (C. gunnelltu) in i sec. The photographs were taken at0-05 sec. intervals. The side of each square ia 3 in. Note the 9mall amplitude of transverse movementof the head.Fig.4. Successive positions of a butterfish in J sec. The photographs were taken at 005 sec. intervals.The side of each square is 1 in. The passage of the waves is marked by dots or by crosses. Notethat the tail is almost at right angles to the path of motion of the fish when it is crossing thelongitudinal axis in photographs 1 and 10.
PLATE II.
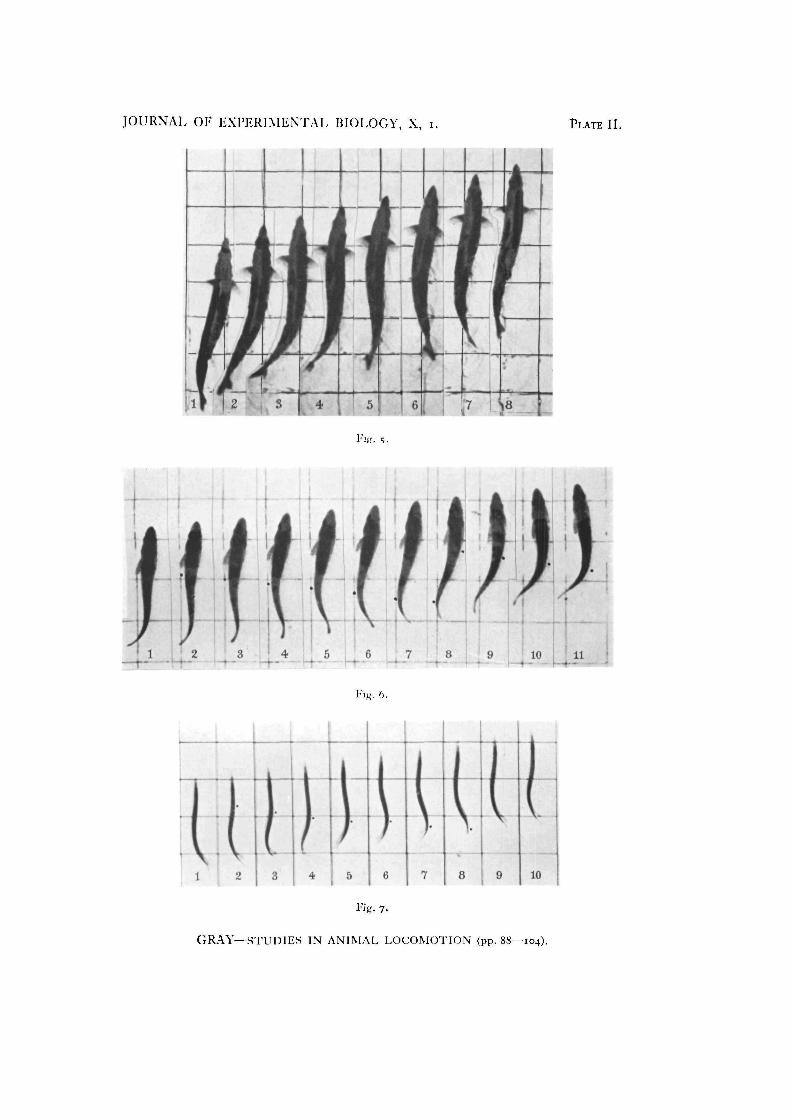
Fig. 5. Successive positions of a mackerel (Scomber tcombrui) within a period of 0035 sec. Theinterval between each photograph was 005 sec. and the grid shown has 3 in. squares. The grid hasbeen inked over in the photograph—note the disturbance of the water in the neighbourhood of thefish. Note also the rapid rate of propagation of the muscular waves and the high forward speed ofthe fish.Fig. 6. Successive positions of a whiting (Gadus merlangus) within a period of 05 sec. Intervalbetween each photograph 005 sec. Scale 3 in. The wave crests are marked by dots. Note that thepitch angle of the tail is greater than that of the butterfish (Fig. 4), but less than that of the mackerel(Fig. 5) or of Ammodytes (Fig. 7).Fig. 7. Successive positions of a sand-eel (Ammodytes lanccolatus) within a period of 05 sec. Theinterval between each photograph is approx. 005 sec. Scale 3 in. Note the relatively high pitch of thebody as compared to the eel shown in Fig. 2. Note also the much greater forward velocity in com-parison to Fig. 2. The forked appearance of the tail is due to the shadow cast on the bottom of thetank.
PLATE III.
Fig. 8. Dogfish. Note the large amplitude of the movements of the body and tail. A wave crest ismarked by a black dot in photographs 3-8. Interval between the photographs 010 sec. Scale 3 in.Fig. 9. Rockling (Onos). Note that the transverse movements are almost completely confined to thetail. Note that the angle of inclination (9) of the body is distinctly steeper than in the dogfish andthat the frequency of the movements is higher. Interval between the photographs 005 sec. Scale3 in.
PLATE IV.
Fig. 10. An eel moving backwards. In 10 sec. the fish has moved back about 3 in. Note the passageof the waves from the tail towards the head of the fish: note also the large amplitude of the waves.Interval between each photograph approx. o-1 sec.Fig. 11. A young glass-eel which is stationary and yet exhibits curvature of the body. Compare withphotograph 4 in Fig. 2. The form of the waves is approximately the same as when the waves aremoving and the fish is in motion. Total period 05 sec. Scale 1 in.

JOURNAL OF EXPERIMENTAL ISIOLOGY, X, i- PLATE I.
1 2 31 4 1 51 61 7 1 8 9 10' 11
«- .1-
1 • U-U
5 i 6 i i 7 i 8 , 9 1 0 11
Fig. 4»
GRAY—STUDIES IN ANIMAL LOCOMOTION (pp. 88—104).

JOURNAL OF EXPERIMENTAL RIOLOGY, X, i. PLATE II.
Fig. 7.
GRAY—STUDIES IN ANIMAL LOCOMOTION (pp. 88—104).


JOURNAL OF EXPERIMENTAL BIOLOGY, X, i
12
PLATE III.
00
GRAY—STUDIES IN ANIMAL LOCOMOTION (pp. 88—104).

JOURNAL OF EXPERIMENTAL BIOLOGY, X, i. PLATE IV.
iV
d
Fig. 10.
3 4 5 6 7 .• • i i
Fig. Ii.
GRAY—STUDIES IN ANIMAL LOCOMOTION (pp. 88—104).
