studies in animal locomotion - the journal of experimental biology
TRANSCRIPT

STUDIES IN ANIMAL LOCOMOTION
VIII. THE KINETICS OF LOCOMOTION OFNEREIS DIVERSICOLOR
BY J. GRAY
Zoological Laboratory, Cambridge
(Received 30 March 1938)
(With One Plate and Eight Text-figures)
THE locomotory movements of a typical polychaete worm, such as Nereis diversicolor,are of interest in that they are effected by two distinct mechanisms, (i) a series ofparapodia which act as levers comparable to the appendages of terrestrial animals,(ii) the longitudinal muscles of the body. When Nereis is moving slowly over a solidsurface, only the parapodia are active; during more rapid motion, or when theanimal is swimming through water, the movements of the parapodia are co-ordinatedwith those of the longitudinal muscles and the two mechanisms combine to give ahighly co-ordinated locomotory mechanism.
Under all circumstances the movements of one side of a segment alternate withthose of the other, and during normal forward progression the unilateral activity ofany one segment begins slightly after that of the segment situated immediatelyposterior to itself. During slow forward movement, waves of activity appear to passalternately over the parapodia of each side of the body from a point situated at theposterior end of the animal. Such a picture is, however, confusing, for no suchcentre of locomotory activity is in conformity with the fact that relatively shortfragments from any region of the body may, if suitably stimulated, exhibit a well-defined and normal locomotory rhythm. A more useful picture of the facts isderived from the observation of a Nereis, originally at rest, starting to move in aforward direction. Cinematograph records taken under such conditions show thatthe first parapodia to be active are always situated near the anterior end of theanimal, and that in a very short period of time a pattern of parapodial activityspreads posteriorly over the body by the activation of the parapodia of every fourthto eighth segment (see Text-fig. 1). As soon as one of these active parapodia beginsits effective stroke, a cycle of activity begins in the parapodium of the segment lyingimmediately anteriorly to it, and this in turn is followed by a similar movement ofthe next anterior neighbour. The original pattern of active parapodia thereby movesanteriorly and gives rise to a series of waves moving, at a relatively low velocity,from the tail of the animal towards the head. The acquisition of the fully activeambulatory state clearly involves two distinct phenomena, (i) the rapid establish-

io J. GRAY
ment of a distinct pattern of parapodial activity over the whole animal from head totail, (ii) a much slower spread of activity from each active segment to the one situatedimmediately anteriorly and ipsilaterally to itself. The number of segments lyingbetween the active parapodia on one side of the animal varies with the region of thebody examined and with the frequency of parapodial movement. In the specimensexamined, the smallest number was two and the largest number was eight. Accordingto Foxon (1936) the parapodial waves pass posteriorly along the body; such a direc-tion has never been seen during the present observations except during backwardprogression.
The propulsive action of an individual parapodium has recently been describedby Foxon (1936). A point d'appui with the substratum is effected by the tip of theparapodium when the latter is directed obliquely forwards. The body of the animalis then subjected to a forward pull by the contraction of the adductor muscles of the
^ ^ s ^ ^ -
\ .1 > ' ' ' ' j " ' '."'~ 'L ' " ' ' " ' i ' U "fr 3>• r s~ s~ s~ S'/S' A s~ \-\s~ /* s i-^A K ^
/ ' • •.« \ \ \ V
./' \ \ \ \ w\. v y\ s N .X >- .'-̂ > '>-\ }s\ 'S V.vr 1 ~ 1 ~ 1" 1 ~ 1 ~ 1" 1 ~ 1 ~ 1"I \~I"i" 1 ~yj~ \Vt* 4
' r r r v r r r r ^\r r r y YText-fig. 1. DiagramshowingthestartofslowanibulationinTVereu. Note the rapid spread (—• — •—)of the ambulatory pattern over the whole body from head to tail and the movement of this patternfrom tail to head at a much slower rate ( ).appendage; when the effective stroke is completed the parapodium faces obliquelybackwards. At the end of this phase the parapodium remains inactive until thebeginning of the next rapid preparatory stroke. During the preparatory stroke, thetip of the parapodium is lifted from the ground and the appendage swung obliquelyforwards and then downwards to effect contact with the ground; the second effectivestroke immediately follows. As pointed out by Foxon, the aciculum is retractedduring the preparatory stroke and extended during the effective stroke.
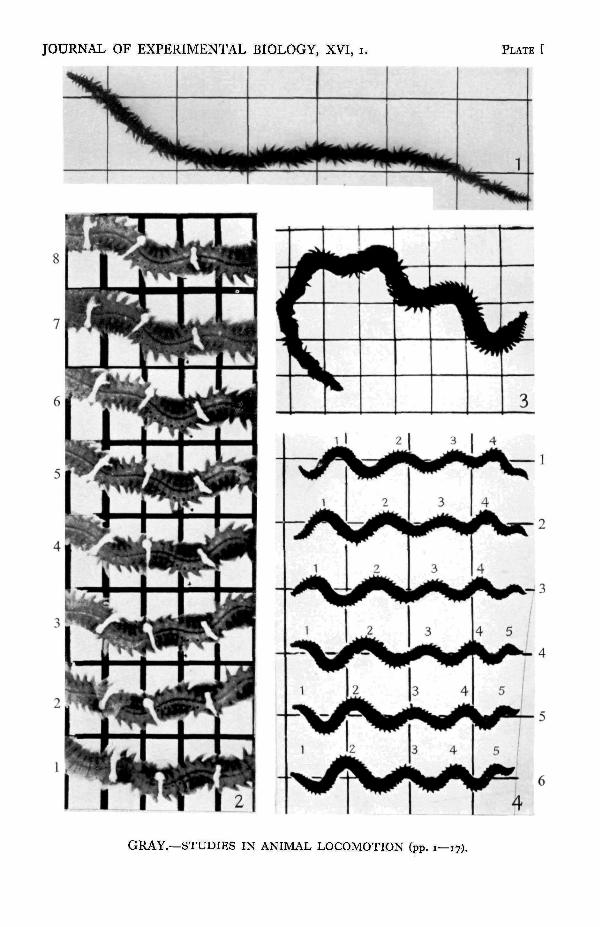
When a Nereis is creeping rapidly over the ground, clearly defined activity of thelongitudinal muscles of the body can be recognized (PI. I, fig. 2), for waves of con-traction and relaxation pass over the body from tail to head. This undulatorymuscular pattern is developed in a manner essentially similar to that described forthe pattern of parapodial activity. Very often the wave pattern appears almostsimultaneously over the whole body, but a number of photographic records indicatethat it starts at the anterior end and spreads very rapidly backwards. As in the case

Studies in Animal Locomotion I I
of the pattern of parapodial activity, the muscular waves, as soon as they appear,begin to move anteriorly at a velocity much lower than the rate of backward spreadof the undulatory pattern itself. Again, there are two phenomena, (i) the rapidspread of the undulatory pattern over the whole of the body, (ii) the spread of thispattern, at a much lower rate, from one region of the body to the one lying imme-diately anteriorly to itself.
The motion of a segment during rapid forward ambulation is illustrated byPI. I, fig. 2. The movements of particular points on the body can be observedby following the movements of the cotton ligatures tied round the body. Fourpoints may be noted: (i) the two ends of each ligature move forward alternately—one end pivoting on the other, (ii) each point on the body remains stationary relativeto the earth during the period at which the adjacent ipsilateral parapodium is
- Direction of motion of wave
Text-fig. 2. Diagram illustrating the motion relative to the ground of two points A^ and Blt onopposite sides of the body of a Nereis, when a complete ambulatory wave of longitudinal contractionpasses over the longitudinal muscles of the body. The wave, as shown, consists of fourteen segmentsand the muscular cycle is therefore conveniently divided into fourteen phases. Since each segmentattaches itself to the ground when in the fully extended state, the wave advances (relative to theground) a distance equal to fourteen times the length of a fully extended segment during the passageof the wave. The advance of the wave, relative to the ground, is shown by the points on the base lineWI-WIJ : the advance (relative to the ground) of the points At and Blt situated on the front edge ofsegment I, is shown by the dotted lines At—Au and B^—Blt respectively.
carrying out its effective stroke, (iii) the parapodium carries out its effectivestroke when the underlying circular muscles are relaxed, (iv) each point on thebody moves forward, relative to the earth, when the underlying longitudinalmuscles are contracted and when the adjacent ipsilateral parapodia are at rest atthe end of their effective strokes.
Under favourable conditions, there is no slip between the ground and a regionof fully relaxed longitudinal muscles, although there may be considerable slip of theparapodium itself over the substratum.
The motion of a typical segment can be followed diagrammatically in Text-figs. 2-4. The wave, as seen in its initial position on the right side of Text-figs. 2and 4, is arbitrarily composed of fourteen segments and is moving from right to left.During the passage of one complete wave over the surface of the body each of thefourteen segments, in turn, will be stationary, relative to the earth, when the under-lying longitudinal muscles are fully relaxed; it follows that while the wave travelsone wave-length over the body, it travels a distance {W^-W^) (relative to the

12 J. GRAY
earth) equal to fourteen times the length (/) of a fully relaxed segment (-BiQ).During the same period the points A1 and Bx at the anterior end of segment I travelalong the lines Ax-Au and Bj-Bu respectively, since segment I will occupy, relativeto the wave, the successive positions shown for each of the segments I-XTV on theright of the figure. The motion of the line A1B1 is shown in Text-fig. 3, and this canbe compared with the motion of the ligatures seen in the cinematograph record inPI. I, fig. 2.
It is clear that during the passage of one complete muscular wave over a pointsituated on the side of the animal's body, that point moves forward along an arc whichis concave towards the outside of the body. Since the distance moved by the waverelative to the body is equal to one wave-length (A) and the distance moved by thewave relative to the earth is nl, where n is the number of segments in the wave and /
Text-fig. 3. Track of the line A1B1 in Text-fig. 2 during the passage of one complete muscularwave. Note that the tracks of the threads shown in PI. I, fig. a are of the same type.
is the length of a segment when it is in contact with the ground, it follows that thedistance moved by the body relative to the earth is nl—A. So long as the wave-lengthand number of segments constituting the wave are constant the value of / depends onthe degree of shortening undergone by the side of a segment when the longitudinalmuscles are fully contracted. The greater is the degree of shortening the longer mustbe the length of a segment during maximum relaxation, and the longer is the distancetravelled by the wave (relative to the ground) during the passage of one completewave. If the body wall of the worm were flexible but not extensible the value of /would be minimal and the maximum rate of progression of the animal would be verymuch slower, for it would only be equal to the difference between the length of thebody and the sum of its constituent wave-lengths. It may be noted that the degreeof contraction of the segments in Text-fig. 2 is very much greater than is typical ofan actual Nereis: in Plate I, fig. 2, the relative rate of progression of the body to that ofthe waves is considerably less than that shown in the diagrams in Text-figs. 2 and 4.

Studies in Animal Locomotion
Direction of movement of wavt
Text-fig. 4. Diagram illustrating the motion, relative to the ground, of one side of a segment, with itsparapodium, when one complete ambulatory wave of longitudinal activity passes over the body ofNereis. The dotted lines show the motion of the points Ax, Bt and Cx during the passage over the bodyof a wave, identical in form with that shown in Text-fig. 2. The preparatory stroke (shown occurringat the n t h phase in the cycle) occurs when the longitudinal muscles begin to extend in length afterrelaxation from a period of active contraction; this phase occurs on the leading surface of the wave.
- Dn-octWa of movem«l of wavt relative to tie bead of tiio uima]
A 1 ' 1 « ,. *$ ' ; ' i '
\ i /// i \- \ \ h
i—VN ) £j / j—! ' *—f r-
/ / .-•' / /
1•«-Direction of movement of wiv, relaUve to the head of the aiisul-*Dlr«tkHi of movement of animal relative to tie (r«nd
(, 5 4 3 2 1
• 4 - DixMtwn of moTtBttt of wava rtlativc to the bmd of the •"ht1fl
v-Dirvctioa of moYsment of «n±m*J rdrtivt to the grtraod
\ J\6\ --, \WrT-\
c
J/
1/ )
}
'•—f—
1
/ / /
1/ 1
-I ./ // j ' t'
s * s 1
Text-fig. 5. Digrams showing that the relationship between the direction of motion of a bilaterallycontractile worm, and that in which waves of longitudinal contraction pass over the body, depends onwhether the body is attached to the ground during periods of maximum longitudinal contraction orduring periods of maximum longitudinal elongation. In A no attachment is effected, the centre ofgravity (CG) of the worm remaining stationary; no progression occurs. In B the worm is attached tothe ground by the posterior edge of each fully contracted segment. Note that the body moves in adirection opposite to that in which the waves of contraction pass over the body. In C attachment iseffected by the posterior edge of each fully relaxed segment. Note that the body moves, relative tothe ground, in the same direction' as that of the waves relative to the head of the animal. In all threefigures the direction, velocity, and form of the waves relative to the body are identically the same. Themovement of each segment relative to the earth is shown by the lines and the movementsof the points d'appui by the lines

14 J. GRAY
The motion of a single parapodium can be followed in Text-fig. 4, wherein theline Bx Cx represents one side of segment I. The parapodium is lifted from the groundas it completes its effective stroke and is carried forwards and inwards as the under-lying longitudinal muscles contract. The parapodium is carried forwards and out-wards as these muscles relax; it is during the early part of this period of relaxationthat the parapodium makes its preparatory stroke. It should be observed that themotive power for this type of locomotion is almost entirely derived from the longi-tudinal muscles pulling against points d'appui which are established by the bases ofthe active parapodia. The adductor muscles of the parapodia probably play a verysubordinate role in the propulsion of the animal. It may be recalled that the esta-blishment of points d'appui by the longitudinally relaxed segments of Nereis is theprecise opposite to the condition found in the earthworm (Gray & Lissmann, 1938),where fixation is effected by those segments showing complete longitudinalcontraction. Text-fig. 5 shows that this difference accounts for the fact that theearthworm progresses in a direction opposite to that in which the muscular wavespass over the body, whereas Nereis moves in the same direction as the waves.Obviously if no fixation occurred, no progression would be effected.
THE PROGRESSION OF NEREIS THROUGH WATER
The movements executed by Nereis when swimming actively through water areessentially the same as those seen during rapid locomotion over the surface of theground, although the amplitude, wave-length,and frequency of the undulatory waves passingover the longitudinal muscles are greatly in-creased, particularly in the anterior region ofthe body (PI. I, fig. 4). The details of transitionfrom ambulation to swimming cannot readilybe followed by the eye, but cinematographrecords show that the change in the form ofthe muscular waves first occurs at the anteriorend of the animal, although the rest of thebody is affected very shortly afterwards.(Text-fig. 6). Once this pattern is established itis transmitted anteriorly as during ambulation.Similarly, when a Nereis ceases to swim the that of swimming.'return to waves of shorter length and amplitudefirst begins at the anterior end of the body and then spreads backwards.
The propulsive mechanism of a swimming Nereis can be followed by a consi-deration of Text-fig. 7, which shows, diagrammatically, the track of a single para-podium, relative to the earth, when a wave (of the same form as that shown in Text-figs. 2 and 4) moves forward, relative to the ground, at the same velocity as it movesover the surface of the body; under these circumstances, of course, the bodywould remain stationary relative to the ground. It can be seen that, during its

Studies in Animal Locomotion 15
effective beat, the parapodium is moving backwards relative to the earth. A move-ment of this type creates a posteriorly directed flow of water towards the hind end ofthe animal; the water is, in fact, driven from regions immediately anterior to theparapodium in the vicinity of a leading surface of a muscular wave and is directedtowards regions lying in the vicinity of a trailing surface. This backward currentsubjects the worm to a forward thrust and under normal circumstances the animalmoves forward through the water. As the worm moves forward the track of aparapodium through the water becomes of the type shown in Text-fig. 8. It will benoted that the backward velocity of the parapodium relative to the earth is partlydue to its own movements relative to the body of the worm and partly to the effectof the longitudinal muscles of the segment. In a form such as Nephthys the parapodiaare probably passive—but they possess, owing to the movements of the underlyinglongitudinal muscles, a definite propulsive effect.
Direction of movement of wive
Text-fig. 7. Diagram showing the movement (relative to the ground) of a parapodium and of indi-vidual points (A, B and C) on the surface of a wave when the wave moves forward (relative to theground) at the same velocity as it moves over the surface of the body. The dotted line • — • — shows thetrack of the point B, situated on the posterior edge of segment XIV; the line — H 1- shows the trackof point A on segment XIV. The track of a parapodium on segment I is also shown. Note that theparapodium, when executing its effective stroke, is travelling towards the hind end of the body andbackwards relative to the ground; this motion causes a flow of water from the leading surface of thewave towards the trailing surface and so subjects the body to a forward propulsive force.
The efficiency of Nereis, as a swimming organism, is not very great, for the rateof progression of the body through the water is very small compared with thefrequency and velocity at which the muscular waves pass over the longitudinalmuscles (see PI. I, fig. 4). If Nereis possessed no parapodia, there can be no doubtthat the animal would progress through the water in a direction opposite to that ofthe muscular waves of the body. It is of some interest to note that if parapodial-likeappendages are attached to a cylinder capable of exhibiting undulatory motion (seeGray, 1936), the direction of water flow past the body of the model is opposite tothat which occurs in the absence of attached appendages, and to that in which theundulations pass over the cylinder itself.
Little or nothing is known of the mechanism responsible for muscular andparapodial co-ordination in Nereis. Removal of the supra- and suboesophagealganglia usually initiates, like other violent stimuli, a period of active swimming,after which the animal temporarily loses tone and cannot usually swim for a con-

16 J. GRAY
siderable period of time. N. virens, on the other hand, exhibits prolonged swimmingmovements when cut into relatively short lengths. Transection of the nerve cordabolishes co-ordination between the two regions of the body, and involves loss oftone by the posterior region, but the movements of each region are normal in type;removal of several adjacent parapodia does not interfere with the propagation of theparapodial or muscular waves.
After removal of the supra- and suboesophageal ganglia, Nereis displays onereflex which may be of ambulatory significance. Such preparations when left un-disturbed usually show little or no spontaneous activity, but if they are subjected to
Direction of motion of worm
IS
Text-fig. 8. Diagram showing the type of movement, relative to the ground, of a single parapodiumwhen a Nereis is swimming forwards in water during the passage of two complete waves over thesegment. Note that during the effective stroke (e.g. positions 8, 9, 10) the parapodium is movingbackwards relative both to the ground and to the muscular wave. The two upper dotted line* showthe track of the corresponding point on the other side of the body.
gentle tension by being drawn over a horizontal surface, the parapodia quicklydisplay an active and normal locomotory rhythm; if the preparation is initially activethe frequency of the rhythm is increased on applying longitudinal tension.
SUMMARY
1. Ambulation in Nereis involves two phenomena: (a) the spread, at a rapid rate,of an ambulatory pattern over the segments of the body, the pattern being propa-gated (during forward progression) from the anterior end of the animal towards thetail; (b) the transmission of this pattern, at a relatively slow rate, in an anteriordirection.
2. During rapid ambulation, the activity of the parapodia is co-ordinated withthat of the longitudinal muscles and progression is, largely, attributable to these

JOURNAL OF EXPERIMENTAL BIOLOGY, XVI, i . PLATE T
GRAY.—STUDIES IN ANIMAL LOCOMOTION (pp. 1—1

Studies in Animal Locomotion 17
muscles. Since one side of each segment is fixed to the ground when the underlyinglongitudinal muscles are fully relaxed, it follows that the animal must progress in thedirection in which the muscular waves travel over the body, and not, as in the case ofthe earthworm, in the opposite direction.
3. The mechanism of swimming is described.
REFERENCESFOXON, G. E. H. (1936). Ann. Mag. nat. Hist. (10), 18, 403.GRAY, J. (1936). J. exp. Biol., 13, 192.GRAY, J. & LISSMANN, H. W. (1938). J. exp. Biol., 15, 506.
EXPLANATION OF PLATE IFig. 1. Photograph (by Mr D. P. Wilson, Marine Laboratory, Plymouth) of Nereis diverticolorcreeping from right to left. Note the variation in the length of the parapodial waves and thecorrelation between these waves and those seen on the dorsal blood vessel, the active parapodia beingsituated on each of the leading surfaces of the muscular waves. Scale: 1 in.Fig. 2. Successive cinema-photographs of an actively creeping Nereis diversicolor moving from rightto left. The movement of the transverse axis of a segment can be followed by observing the move-ments of the three transverse threads attached to the body. Note that each point moves forwardwhen the underlying longitudinal muscles are contracted or contracting and is at rest when thesemuscles are relaxed. Scale: 1 cm.Fig. 3. Transition from ambulation to swimming in Nereis virens. Note that the anterior end of thebody shows typical swimming waves, while the posterior end still displays typical ambulatory wavesof short wave-length and smaller amplitude.Fig. 4. Successive cinema-photographs of Nereis diversicolor swimming from right to left. Note theslow rate of progression of the animal through the water compared to the speed of movement of thewaves (indicated by numerals) relative either to the body or to the ground. Scale: 1 in.
JEB -XVli