structure of rickettsia quintana cultivated in vitro the lousestructure of rickettsia quintana...
TRANSCRIPT

JOURNAL OF BACrERIOLOGY, Feb., 1965Copyright © 1965 American Society for Microbiology
Vol. 89, No. 2Printed in U.S.A.
Fine Structure of Rickettsia quintana CultivatedIn Vitro and in the Louse
SUSUMU ITO AND J. W. V'INSONDepartment of Anatomy, Harvard Medical School, and Department of Microbiology, Harvard School of
Putblic Health, Boston, Massachusetts
Received for publication 21 September 1964
ABSTRACT
ITO, SUSUMU (Harvard Medical School, Boston, Mlass.), AND J. W. \'INSON. Finestructure of Rickettsia quintana cultivated in vitro and in the louse. J. Bacteriol. 89:481-495. 1965.-Usually rod-shaped, Rickettsia quintana cells measure about 0.2 to 0.5 ,wide and up to 1.6 IA long. The rickettsiae have both an outer cell wall, about 80 A thick,and a plasma membrane, about 70 A thick, each of which is trilaminar. Occasional vesic-ular invaginations of the plasma membrane occur. The nuclear material, distributed inirregular zones throughout the cytoplasm, appears as a loose network of fine fibrilswhen postfixed with uranyl acetate and as thick strands or clumps after routine OS04 fixa-tion. The cytoplasm is densely packed with numerous granules, presumably ribosomes,about 150 A in diameter. Histochemical studies revealed the presence of both ribonucleicand deoxyribonucleic acids. During binary fission, a constricting furrow is formed bythe cell wall and plasmalemma. No difference in fine structure was observed betweenR. quintana propagated on cell-free media and in the louse.
Rickettsiae constitute a grotul) of microorgan-isms regarded as intermediate between bacteriaand viruses. Although their morphological andphysiological characteristics more closely resem-ble those of certain bacteria, they depend, likeviruses, on living host cells for their multiplica-tion. With the exception of Rickettsia quintana,rickettsiae are obligatory intracellular parasites.R. quintana, the etiological agent of trench fever,multiplies extracellularly in the gut of its arthro-pod vector, Pediculus humanus var. corporis(Wolbach, Todd, and Palfrey, 1922); and, re-cently, Vinson and Fuller (1961) and Vinson,Varela, and Molina (in preparation) reportedthe cultivation of R. quintana on cell-free media,as well as in lice, in cells cultivated in vitro, andin the yolk sac of the developing chick embryo.
Although studies on the fine structure ofseveral rickettsiae have been reported by variousinvestigators (Plotz et al., 1943; Shepard andWyckoff, 1946; Stoker, Smith, and Fiset, 1956;Van Rooyen and Scott, 1949; Wissig et al., 1956),comparable studies on R. quintana have not beenpublished. The present study attempts to defineand compare the fine structure of R. quintanacultivated on blood-agar and in the louse.
MATERIALS AND METHODS
Two lines of the Fuller strain of R. quintanawere used: (i) microorganisms isolated directly onblood-agar from the blood of a trench fever patient
and passaged seven times on this medium, and (ii)microorganisms isolated from the patient's bloodin lice and subsequently passaged in series in theyolk sac of the chick embryo, in cells cultivatedin vitro, in lice, and on blood-agar (Vinson andFuller, 1961). Lice were infected with rickettsiaefrom the first line. For the examination of micro-organisms propagated on blood-agar, both linesof rickettsia were used.
R. quintana propagated on blood-agar. Coloniesof R. quintana, propagated on blood-agar by themethod of V'inson and Fuller (1961), were sus-pended in a solution of phosphate, glutamate, andsucrose (pH 7.0), referred to hereafter as sucrose-PG (Bovarnick, Miller, and Snyder, 1950). Thecolonies were sedimented by centrifugation at450 X g for 15 min. In some cases the rickettsiae were washed twice in sucrose-PG. The re-sulting pellet was partially disrupted, and theseparate fragments were fixed in each of the follow-ing fixatives: 1%o OS04 buffered with V-eronalacetate (R-K fixation) as recommended by Ryterand Kellenberger (1958); s-collidine-bufferedOS04 at pH 7.2 to 7.4 (Bennett and Luft, 1959),containing 5% sucrose; phosphate-buffered OSO4according to the method of Millonig (1962); and4% glutaraldehyde (Sabatini, Bensch, and Barr-nett, 1963) buffered with s-collidine and postfixedwith OS04 (with the same buffer). The rickettsiaewere dehydrated in a graded series of cold ethanolsolutions, and the microorganisms were recoveredbetween each ethanol solution by centrifugationfor 3 to 5 min. Embedments in Epon, Vestopal,and Araldite were made by standard, recom-
481
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

ITO AND VINSON
mended procedures. Since Epon appeared to besatisfactory for these studies, it was used for mostof the observations. The other plastics gavecomparable results.
In addition to the study of sectioned rickett-siae, intact microorganisms cultivated on blood-agar and suspended in sucrose-PG were examinedafter negative staining with phosphotungstic acid(Valentine and Horne, 1962).R. quintana propagated in the louse. Young
adult female lice, from a laboratory strain main-tained on rabbits and known to be free frommicroorganisms resembling rickettsiae, were
inoculated intrarectally, by the technique ofWeigl (1920), with R. quintana suspended insucrose-PG. These microorganisms were from theseventh passage on blood-agar. The lice were fedto repletion once daily on a normal rabbit, andbetween feedings were maintained at 32 C. Theprogress of infection was followed in each louseby daily microscopic examination of Giemsa-stained smears of its feces. Lice used for electronmicroscopy were fasted for 48 hr before killing.Each louse was then placed on a clean slide near
a drop of fixative. After excision of the terminalabdominal segment, the abdominal contents weresqueezed out into the fixative by immobilizing thethorax and compressing the abdomen from thethorax posteriorly with a needle. The abdominalviscera were then placed in fresh fixative andtreated as were the pellets described above,except that centrifugation was unnecessary. Somespecimens were obtained by cutting off the ter-minal abdominal segment and placing the wholeabdomen into the fixative. After fixation, theviscera were dissected from the exoskeleton andembedded as one piece. The louse tissues wereembedded in the same series of materials as werethe rickettsial suspensions. Sections were cut withglass knives, picked up on celloidin grids, stainedwith uranyl acetate followed by lead citrate(Reynolds, 1963), or by lead citrate alone, andexamined with an electron microscope. Normalcontrol lice were processed by the same methods.
Light-microscope observations were made onthick (0.5 to 1.5 pu) sections of the Epon-embeddedmaterials. Sections were stained with alkalinetoluidine blue (Trump, Smuckler, and Benditt,1961), periodic acid Schiff (PAS) reaction, or theFeulgen stain. Whole microorganisms from par-tially disrupted colonies were washed in saltsolution, serially diluted, and air-dried on gelatin-coated slides. They were then stained by theGim6nez (1964) stain. Other slides were fixed inCarnoy's solution (glacial acetic acid-absoluteethanol, 1:3) for 15 min, washed with water, andtreated with deoxyribonuclease (0.2 mg/ml inphosphate buffer at pH 7.5 containing 0.03 M
MgSO4) for 1 to 5 hr at 37 C, or with ribonuclease(0.2 mg/ml in distilled water) for 2 hr at 37 C.Some preparations were digested with both nu-
cleases. The Feulgen stain for deoxyribonucleicacid (DNA), toluidine blue (0.5% at pH 3.0) forcytoplasmic basophilia, and acid-fast green (0.5%)
for proteins were used to stain the test prepara-tions.
RESULTS
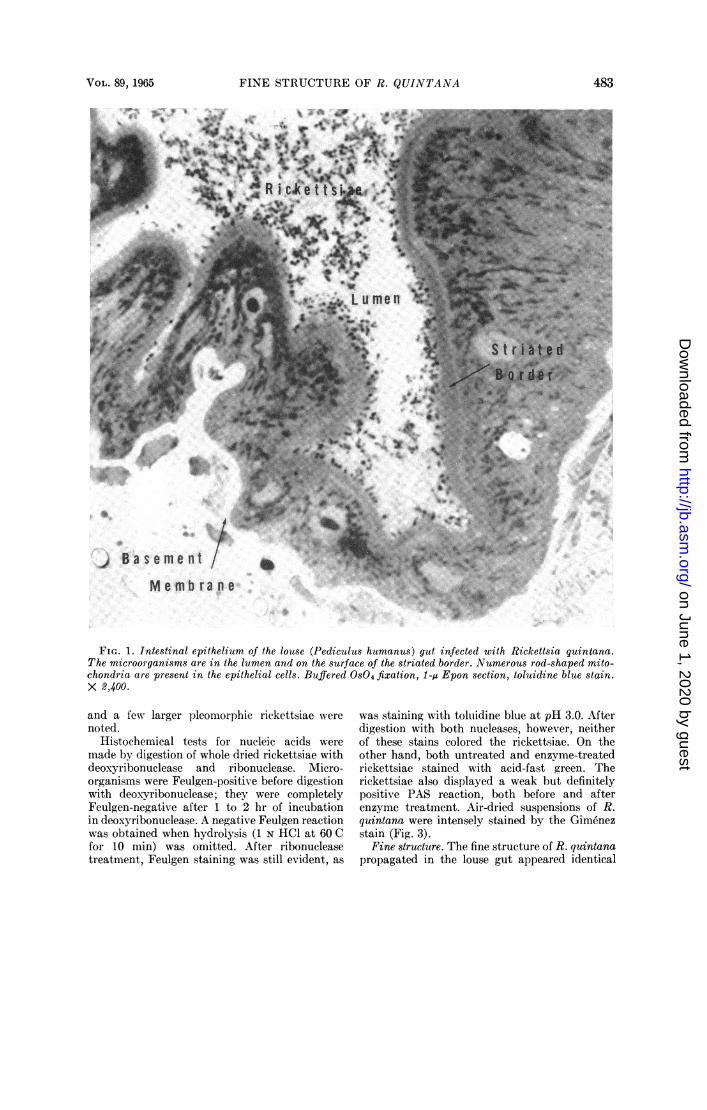
Light microscopy. Fixed louse gut embeddedin Epon, sectioned between 0.25 and 1.5 u, andstained with toluidine blue was examined in alight microscope to determine the extent of infec-tion and the distribution of the rickettsiae (Fig.1). Numerous rickettsiae are present in the lumenof the gut as well as on the free surface of thestriated border of the lining epithelial cells.The intestinal epithelium is formed of a single
layer of columnar to low columnar cells whoselateral boundaries cannot be clearly distinguishedwith the light microscope. The cells contain anumber of mitochondria and lightly stainednucleus with a prominent nucleolus. The basalsurface of the epithelium rests on an unusuallythick basement membrane, the structure ofwhich appears to differ from the usual basementmembrane of vertebrates in failing to stain withthe PAS reaction. It also failed to stain withtoluidine blue.When the intestinal lumen was distended with
blood, the epithelium became attenuated and insome areas did not exceed 1 ,u in thickness. Mostof these observations were limited to fasted licebecause the high density of hemolyzed blood inthe lumen rendered observation of the rickettsiaedifficult.
In heavily infected lice, the rickettsiae werefound in association with the free surface of amajority of the epithelial cells and in the lumenof the gut, where they tended to be aggregatedinto tightly packed clusters of varying size. Insome animals, the intestine contained bacterialcontaminants intermingled with the rickettsiae.These bacteria were distinctly different fromthe rickettsiae in size, shape, and staining charac-teristics. Observations were limited to rickettsiaein lice free from contaminating microorganisms.
R. quintana cultivated on blood-agar grew asmicrocolonies composed of densely packed organ-isms. These colonies, which measured 200 ,u ormore in diameter, were circular in outline andappeared translucent with reflected light. Intransverse section the colony had a convex freesurface and a flat area in contact with the surfaceof the blood-agar (Fig. 2). The central part ofthe colony was up to 40 ,u thick. No notablemorphological differences were found amongrickettsiae situated in different areas of thecolony.
Light microscopy revealed little internal detailof the rickettsiae. A central constriction furrowwas observed in some of the larger bacillary forms,
482 J. BACTERIOL.
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
FINE STRUCTURE OF R. QUINTANA
a.F g J li~~~~~f
-SRws:.<,%W%+A ~4;X FAt'^r'~~~~~41
M e m b r a n e
FIG. 1. Intestinal epithelium of the louse (Pediculus humanus) gut infected with Rickettsia quintana.The microorganisms are in the lumen and on the surface of the striated border. Numerous rod-shaped mito-chondria are present in the epithelial cells. Buffered OS04 fixation, 1-Au Epon section, toluidine blue stain.X 2,400.
and a few larger pleomorphic rickettsiae werenoted.
Histochemical tests for nucleic acids weremade by digestion of whole dried rickettsiae withdeoxyribonuclease and ribonuclease. Micro-organisms were Feulgen-positive before digestionwith deoxyribonuclease; they were completelyFeulgen-negative after 1 to 2 hr of incubationin deoxyribonuclease. A negative Feulgen reactionwas obtained when hydrolysis (1 N HCI at 60 Cfor 10 min) was omitted. After ribonucleasetreatment, Feulgen staining was still evident, as
was staining with toluidine blue at pH 3.0. Afterdigestion with both nucleases, however, neitherof these stains colored the rickettsiae. On theother hand, both untreated and enzvme-treatedrickettsiae stained with acid-fast green. Therickettsiae also displayed a weak but definitelypositive PAS reaction, both before and afterenzyme treatment. Air-dried suspensions of R.quintana were intensely stained by the Gimenezstain (Fig. 3).
Fine structure. The fine structure of R. quintanapropagated in the louse gut appeared identical
483VOL. 89, 1965
MP A
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

ITO AND VINSON
FIG. 2. Section of three colonies of Rickettsia quintana cultivated in vitro. The microorganisms aredensely packed, homogeneous aggregates. Between and around the colonies are clusters of parttially disruptedaggregates of rickettsiae. The inset shows part of a colony at higher magnification. Buffered OS04 fixation,0.5-,u Epon section, toluidine blue stain. X 1,200; inset, X 3,400.
FIG. 3. Suspension of Rickettsia quintana cul-tivated in vitro, dried on a slide, and stained by theGimenez method. X 1,900.
with that of rickettsiae cultivated in vitro. Vari-ous procedures of preparation )roduced some
differences in the general appearance of therickettsiae. The fixation technique suggested by
Ryter and Kellenberger (1958) yielded resultscomparable to those with bacteria that are wellpreserved bv this method. Therefore, the descrip-tion of the internal structure of R. quiintana isbased on microorganisms fixed by this method.
In thin sections the microorganisms are seenas circular or rod-shaped profiles, about 0.2 to0.5,u wide and up to 1.6 ,u in length. The limitingboundary is formed by two clearly defined struc-tures: the rickettsial cell wall, which surroundsthe whole microorganism, and the underlyingplasma membrane, which limits the cytoplasm.(The term "rickettsial cell wall" will be used todescribe the outer limiting structure surroundingR. quintana since the wall structure is similar tothe gram-negative bacterial cell wall.) Sectionsperl)endicular to the wall and plasmalemma revealtheir total thickness and their interspace to beabout 200 to 250 A (Fig. 7, 8, 11). The cell wall isabout 80 A thick, and the plasma membrane isabout 70 A. Each of these membranes is tri-laminar and resembles the unit membranedescribed by Robertson (1959). The (lear spaceseparating the cell wall from the plasmalemmavaries from 50 A to several times this width.
484 J. BACTERIOL.
-- --- --- ----
lk4k
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

FINE STRUCTURE OF R. QUINTANA
The contours of the rickettsiae are generallysmooth, with few irregularities. Some specimens(Fig. 9, 10) show wrinkled or bulbous outpocket-ings of the cell wall. These may have been due toinadequacies of the preparation technique, todegeneration, or to suboptimal growth conditions.The outer surface of the cell wall is sometimescoated with a fuzzy material of moderate opacity(Fig. 7, 8). Since it is not a consistent component,it may be adsorbed extraneous material ratherthan an integral part of the cell wall. Adjacentmicroorganisms are separated by a space of lessthan 100 A. Thus, if a permanent external coatingis present, it must be a very thin layer indeed,and not distinctly shown in these preparations.The plasmalemma has occasional vesicular
inpocketings measuring up to 150 m,u in diameter(Fig. 8, 9). These invaginations resemble thepinocytotic vesicles or vacuoles seen in animalcells. Their contents are of low electron densityand are continuous with the structureless clearspace between the plasma membrane and the cellwall. At the site of these invaginations, the cellwall is uninterrupted and does not parallel theinfolded plasmalemma.The cytoplasmic surface of the plasma mem-
brane may be adjacent to areas containing eitheran array of fine filaments or dense granules, 150 Ain diameter. The distribution and number of thesegranules vary from one rickettsia to another.They may be in linear array (Fig. 7) or groupedin irregular clusters. Although these granules aresomewhat smaller than the usual ribosomes foundin mammalian cells, they probably represent theribonucleoprotein component of this organism.The interstices between the aggregations of
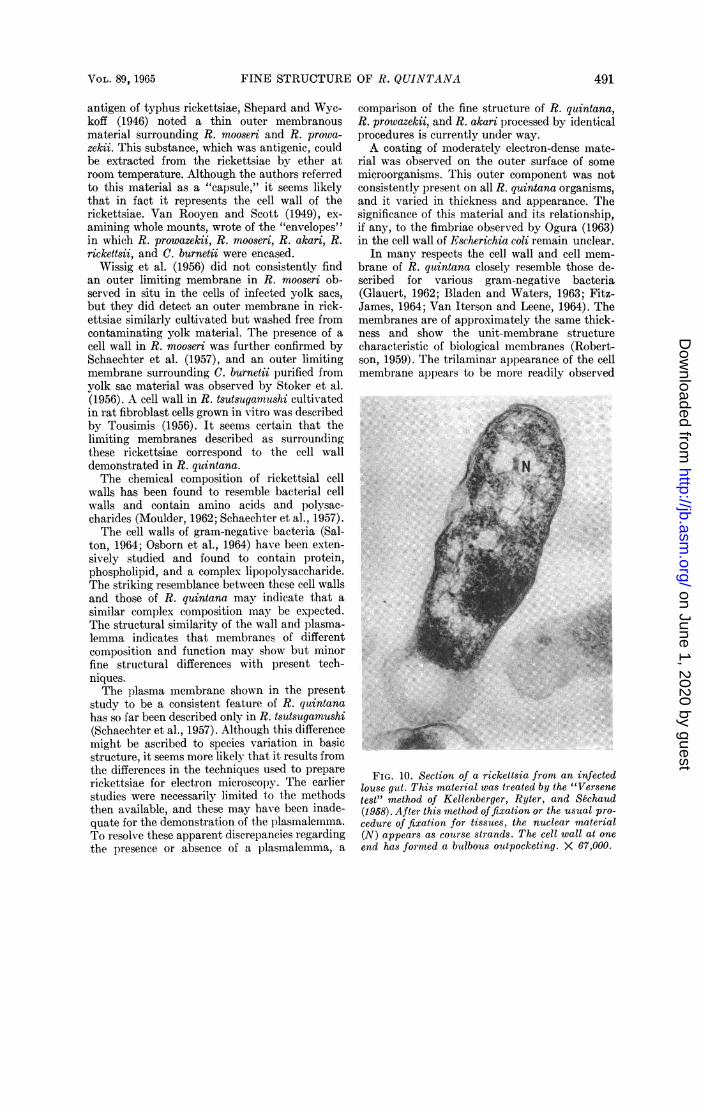
the ribosomal particles are occupied by a mesh-work of fine filamentous strands of about 20 to50 A in diameter (Fig. 7, 8, 13, 14). These struc-tures were observed after R-K fixation (Ryterand Kellenberger, 1958), which involves post-fixation in uranyl acetate. After routine fixationmethods, which are ordinarily satisfactory foranimal tissues, or after application of the so-called"Versene test" for DNA (Kellenberger, Ryter,and Sechaud, 1958), the cytoplasm was preservedinstead as a network of coarse strands and clumpsof dense material (Fig. 5, 6, 10). These differingresponses to fixation suggest that the filamentousareas of cytoplasm comprise the DNA componentof R. quintana. No other structural componenthas been regularly encountered in the cytoplasmof these rickettsiae.
In addition to the more commonly encounteredforms just described, rickettsiae thought to beundergoing binary fission were frequently found(Fig. 5, 6, 11, 12). A furrow is formed by the
apparent constriction of both the cell wall andthe plasma membrane (midway along the length)of the microorganism. These binary forms werefound only in the longest rod-shaped rickettsiae,ranging up to 1.6 A in length. Dividing rickettsiaeshowed no distinct alteration of their internalstructure during this process. No specializedstructure was detected within the cell whichmight explain the mechanism of fission. There issome suggestion of modification of the limitingmembranes, which appear as annular profiles atthe constriction furrow (Fig. 11). Rickettsiaewith shallow furrows, which probably indicatethe initial stages of fission, and later stages leadingto complete cleavage into two microorganisms,have been observed. These varied forms weremore readily observed in negatively stained wholemicroorganisms. In such preparations the ob-servation of cleavage did not depend uponfortuitous orientation of the rickettsiae, as itdoes in sections, since they all tended to lie flaton the supporting membrane of the specimengrid. Negatively stained preparations revealsurface features of the plasma membrane and thecell wall but little of the internal structures (Fig.9, 12). The only cytoplasmic component demon-strable by this technique is an occasional vesicularinfolding of the plasma membrane (Fig. 9).
Electron microscopy of the infected lousemidgut revealed varying numbers of rickettsiaein the lumen. Some were apparently free in thelumen while others were intimately associatedwith the plasma membrane of the intestinalepithelial cells (Fig. 4, 5). This close associationdid not appear to involve any alteration of thefine structure of either the host cell or the micro-organism. In lice with a low or moderate degreeof infection, the preferential association of numer-ous rickettsiae with certain cells was noted.Although these cells had a striated border inwhich the microvilli were short, sparse, and dis-ordered, their cytoplasm appeared normal (Fig.5). They are interpreted as cells which willundergo exfoliation. Some dislodged epithelialcells with broken cell membranes were found inthe lumen. These cells often contained rickettsiaein their interior. R. quintana was never observed,however, in the cytoplasm of intact, normalintestinal cells.
DISCUSSION
It has long been known that R. quintana nor-mally proliferates in the lumen of the louse gutand not within the intestinal epithelial cells (Wol-bach et al., 1922). The present light-microscopeobservations are in agreement with the earlierstudies, and the fine-structure study has further
VOL. 89, 1965 485
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

ITO AND VINSON
LUMEN
xR
7N, .~~~~~~~~~.
..B
L.,..s,>,,
B L
PERIVISCERALHAEMOCOELE
FIG. 4. Intestinal epithelium of the louse. The lumen contains several rickettsiae (R); some are in closeassociation with the microvilli which form the striated border (SB) . The dense mitochondria in the epithelialcells have a different internal structure from the extracellular microorganisms. A basement lamnina (BL)or membrane of considerable thickness borders the base of the epithelium. X 9,500.
confirmed the exclusive extracellular localizationof R. quintana in the louse intestine.
All workers who have examined rickettsiaewith the electron microscope have noted a limit-ing membrane surrounding the microorganisms,
although several different terms have been usedto describe this structure. Plotz et al. (1943) firstdescribed this limiting membrane in R. prowa-zekii, R. mooseri (R. typhi), R. rickettsii, andCoxiella burnetii. In an ear-ly report on the soluble
486 J. BACTERIOL.
e
.Jf
ibil .kk C
,:i. 1.
,:.
..:.1 :,!.,;t
;, it
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

AL
.41
sn:s;,::. ;;U,
a O i.Mf.;!,
... .
s.:t:
q,n.. T
Xt
rf
w:.,
.. .........
...Owl:
.. .....
......
yo
FIG. 5. Apical parts of two intestinal epithelial cells of the louse. On the cell to the right, the usually
well-developed striated border is markedly disrupted. Rickettsiae are more often in intimate association
with such cells than with more normal cells, as illustrated along the lower margin. Note the diplobaccillaryform with a division furrow (arrow). X 19,000.
487
..... ..1.......
..... ...
-t
.: i: .:.:
.1 vN!:.:.,.W...i ;
ii, lk .. Ki-, Ifz.. ib IN.
P -, ..iW. I r ..:
.14',
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

i
4
A
FIG. 6. Electron microggraph through the central part of a colony of Rickettsia quintana grown in vitro.Circular profiles represent transverse sections; elongate forms are sectioned longitudinally. Note the con-constriction furrows across the centers of several rickettsiae. With the exception of a few pleomorphic forms,the colony is remarkably homogeneouts. X 24,000.
488
rqp,e PO .,I
.4, .-.IL.A*I
itA I
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

p~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.Nstc~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~4j,,;:.~~~~~~~~~~~NmI0tisoi;:~~~~~~~~~~~~~~~~~~~~~~-N,! W X E~~~~~~~J
Kw ;; * iibi | iF ¢ ; *~~~~~~~~~~~~~~~~~~WMtf~~~~~~~~~~~~~~~~~~~~~~ i;'S;'-it;_ls .:: i9_,j? :: :': _' Awll.:
Asit: B' ... _ . . tF : ; 'rJ ~~~~,iue ,, )l,, , ^,; ^ ......... W. .........|'is.;e,,,,,.,,.._..t'...... vs- ';'; r K 's.: 'tS'." .si'.. -- t:'~~~~~~~~~~~~~~~~-1of
*dgb>,,;, '';,.; , ' 's~~~~~~~~~~Ai,
FIG. 7. Thin sections through several Rickettsia quintana cells from the intestine of an infected louse.The detailed structure of the microorganism is well shown after R-K fixation. Both cell wall (CW) and plasmamembrane (PM) are trilaminar when viewed on edge. The arrow indicates an oblique section of a vesicularinpocketing of the plasma membrane. Granules presumed to represent ribosomes (R) are distributed inclusters throughout the cytoplasm. The nuclear component (N) is represented as areas with fine filamentsbetween the clusters of ribosomes. X 125,000.
489
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

FIG. 8. Transverse section of a rickettsia showing plasma membrane (PM) infoldings which form vesicles(arrows). These infoldings are simple invaginations of the plasma membrane and have a content of low elec-tron density. The upper vesicle is sectioned tangentially. The trilaminar limiting membranes, the filamentousnature of the nuclear component, and dense granules presumed to be ribosomes are also apparent. >X 190,000.
FIG. 9. Intact rickettsia negatively stained with phosphotungstic acid (PTA). The cell wall is the outerlimiting contour of the organism. At the upper left the plasma membrane has separated a considerable dis-tance from the cell wall. The profiles marked by the arrows represent the vesicular infoldings of the plasmamembrane. The vesicles at the right appear to be continuous with the plasmalemma. X 95,000.
40n
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
FINE STRUCTURE OF R. QUINTANA
antigen of typhus rickettsiae, Shepard and Wyc-koff (1946) noted a thin outer membranousmaterial surrounding R. mooseri and R. prowa-zekii. This substance, which was antigenic, couldbe extracted from the rickettsiae by ether atroom temperature. Although the authors referredto this material as a "capsule," it seems likelythat in fact it represents the cell wall of therickettsiae. Van Rooyen and Scott (1949), ex-amining whole mounts, wrote of the "envelopes"in which R. prowazekii, R. mooseri, R. akari, R.rickettsii, and C. burnetii were encased.
Wissig et al. (1956) did not consistently findan outer limiting membrane in R. mooseri ob-served in situ in the cells of infected yolk sacs,but they did detect an outer membrane in rick-ettsiae similarly cultivated but washed free fromcontaminating yolk material. The presence of acell wall in R. mooseri was further confirmed bySchaechter et al. (1957), and an outer limitingmembrane surrounding C. burnetii purified fromyolk sac material was observed by Stoker et al.(1956). A cell wall in R. tsutsugamushi cultivatedin rat fibroblast cells grown in vitro was describedby Tousimis (1956). It seems certain that thelimiting membranes described as surroundingthese rickettsiae correspond to the cell walldemonstrated in R. quintana.The chemical composition of rickettsial cell
walls has been found to resemble bacterial cellwalls and contain amino acids and polysac-charides (Moulder, 1962; Schaechter et al., 1957).The cell walls of gram-negative bacteria (Sal-
ton, 1964; Osborn et al., 1964) have been exten-sively studied and found to contain protein,phospholipid, and a complex lipopolysaccharide.The striking resemblance between these cell wallsand those of R. quintana may indicate that asimilar complex composition may be expected.The structural similarity of the wall and plasma-lemma indicates that membranes of differentcomposition and function may show but minorfine structural differences with present tech-niques.The plasma membrane shown in the present
study to be a consistent feature of R. quintanahas so far been described only in R. tsutsugamushi(Schaechter et al., 1957). Although this differencemight be ascribed to species variation in basicstructure, it seems more likely that it results fromthe differences in the techniques used to preparerickettsiae for electron microscopy. The earlierstudies were necessarily limited to the methodsthen available, and these may have been inade-quate for the demonstration of the plasmalemma.To resolve these apparent discrepancies regardingthe presence or absence of a plasmalemma, a
comparison of the fine structure of R. quintana,R. prowazekii, and R. akari processed by identicalprocedures is currently under way.A coating of moderately electron-dense mate-
rial was observed on the outer surface of somemicroorganisms. This outer component was notconsistently present on all R. quintana organisms,and it varied in thickness and appearance. Thesignificance of this material and its relationship,if any, to the fimbriae observed by Ogura (1963)in the cell wall of Escherichia coli remain unclear.
In many respects the cell wall and cell mem-brane of R. quintana closely resemble those de-scribed for various gram-negative bacteria(Glauert, 1962; Bladen and Waters, 1963; Fitz-James, 1964; Van Iterson and Leene, 1964). Themembranes are of approximately the same thick-ness and show the unit-membrane structurecharacteristic of biological membranes (Robert-son, 1959). The trilaminar appearance of the cellmembrane appears to be more readily observed
J.!
FIG. 10. Section of a rickettsia from an infectedlouse gut. This material was treated by the "Versenetest" method of Kellenberger, ftyter, and &chaud(1958). After this method offixation or the usual pro-cedure of fixation for tissues, the nuclear material(N) appears as courise strands. The cell wall at oneend has formed a butlbous outpocketing. X 67,000.
VOL. 89, 1965 491
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

FIG. 11. (top) Longitudinal section of a diplobacillary formn of Rickettsia quintana. T'he constrictionfurrow formed by the cell wall and the plasma membrane is almost complete. Although the furrow usuallybisects the microorganism, adequal division as illustrated here is fairly common. X 105,000.
FIG. 12. (bottom) Dividing rickettsia negatively stained with PTA. This intact organism is compar ablewith the section shown in Fig. 1 1. The slightly larger size is probably due to the flattening of the microorganismduring drying. The excessive enlargement causing the limiting membranes to be distinct in the lower righthalf is also a consequence of specimen preparation. X 86,000.
492
a,
e4t;
4i."I ;.,;!,
h:
.9
.W-lkl, ..:!%-1--tii
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

FINE STRUCTURE OF R. QUINTANA
FI(.. 13. (left) Section of rickettsia illustrating plecmnorphism. Though relatively infrequent, micro-organzisms with varying (legrees of pleomorphism are found in tmIost samples. Such rickettsiae are usuallylatge and less electron-dense than the more common rod-shaped forms. X 59,000.
Fi(n. 14. (right) Same as Fig. 13. X 55,000.
in R. quintana than in bacteria because the cyto-plasm in the former is less densely packed withelectron-opaque material.The vesicular or vacuolar invaginations of the
plasmalemma of R. quintana are unusual. Suchinfoldings of the membrane have not been com-monly observed in gram-negative bacteria(Glauert, 1962); however Lee (1960) describedsmall intrusions of the plasma membrane intothe cytoplasm of a sl)herol)last of E. coli. AMorecomplex membranous components (chondroids,membranous bodies, or mesosomes) have beendescribed in numerous gram-positive bacteria(Glauert, 1962). These structures, which differfrom the simple vesicular infoldings seen inR. quiintana, are concentric lamellar bodies whichseem to have been formed by repeated infoldingsof the plasma membrane. A similar system ofinternal membranes has been observ-ed in Listeriamonocytogenes (Edwards, 1963).The mechanism of cell division can only be
suoggested by the examination of static prepara-tions. In the present studlies, eonstrictions involv-ing both plasma membrane and cell wall and
approximately bisecting the long axis of themicroorganism have been observed in varyingstages of development from shallow furrows toalmost complete cleavage. Binary fission is gen-erally acce)ted as the mechanism of division forrickettsiae (Moulder, 1962), and Schaechter et al.(1957) observed this process in R. rickettsii bymeans of phase microscopy in cells cultivated invitro. Such a graded series of cleavage stages aswe have seen, as well as the numerous diplobacil-lary forms encountered, substantiates theseobservations and establishes that binary fissionis, in fact, the mode of reproduction in R. quin-tana. The fine structure of rickettsiae undergoingdivision is quite similar to that of E. coli de-scribed by Conti and Gettner (1962).The presence of DNA in rickettsiae has been
suggested by the histochemical studies of Ris andFox (1949), who detected Feulgen-positive stain-ing of R. prowazekii. Moreover, Smith and Stoker(1951) found that 9.7% of the dry weight of C.burnetii was DNA. The present study shows thatR. quintana is Feulgen-positive and that deoxy-ribonuclease abolishes this staining reaction.These results verify the presence of DNA in R.
493Vo L. 89, 1965
OF-, ,:, V
I
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

ITO AND VINSON
quintana. The fine structure of the DNA compo-nent in R. quintana appears similar to that foundin bacteria. When the microorganism is fixedaccording to the method of Ryter and Kellen-berger (1958), the nuclear material appears as anirregular central zone of lower electron density,loosely filled with fine fibrils (50 A or less indiameter). Furthermore, the appearance of theseareas after ordinary fixation or after the "Versenetest" of Kellenberger shows the characteristiccoarse strands typical of inadequately fixedDNA.The presence of ribonucleic acid (RNA) in R.
quintana is suggested by toluidine blue stainingat pH 3.0 after deoxyribonuclease digestion andby complete lack of staining after treatment withboth nucleases. Biochemical analysis of C.burnetii by Smith and Stoker (1951) showed thatsome 4.3% of its dry weight is RNA. In theelectron micrographs of R. quintana, discretegranules about 150 A in diameter are common andare presumed to represent the ribosomes.No consistent difference in the fine structure
or staining characteristics was seen between R.quintana propagated in the louse gut and on cell-free medium. The etiological agent of trench feveris the only rickettsia pathogenic for man to becultivated in vitro. It is hoped that the elucida-tion of the fine structure of R. quintana will serveas a basis for a comparative examination of theother rickettsiae and that, combined with physio-logical and biochemical studies, it will extend ourunderstanding of the relationships existing amongthe members of this group of microorganisms.
ACKNOWLEDGMENTSThis study was supported by Public Health
Service grant GM10182 and by the Office of TheSurgeon General, Department of the Army, withthe sponsorship of the Commission on RickettsialDiseases of the Armed Forces EpidemiologicalBoard.The technical assistance of Sylvia Sawyer and
Ann Shortell in cultivating the rickettsiae and ofGail Brassil in performing the histochemicalreactions is gratefully acknowledged.
ADDENDUM IN PROOFStructures resembling the cell wall and plasma
membrane described here were recently reportedin Coxiella burnetii (Anacker et al., J. Bacteriol.88:1130-1138, 1964). The surface of this rickettsiawas shown to have "four distinct lines which de-lineate a structure or structures approximately200 A thick." The authors designated this as the"multiple external layers" and did not attempt tointerpret them as the cell wall and plasmalemma.The appearance and dimensions of these limitingmembranes are almost identical to those of R.quintana, and probably represent the equivalentstructures.
LITERATURE CITEDBENNETT, H. S., AND J. H. LUFT. 1959. s-Collidine
as a basis for buffering fixatives. J. Biophys.Biochem. Cytol. 6:113-115.
BLADEN, H. A., AND J. F. WATERS. 1963. Electronmicroscopic study of some strains of Bacteriodes.J. Bacteriol. 86:1339-1344.
BOVARNICK, M. R., J. C. MILLER, AND J. C.SNYDER. The influence of certain salts, aminoacids, sugars, and proteins on the stability ofrickettsiae. J. Bacteriol. 59:509-522.
CONTI, S. F., AND M. E. GETTNER. 1962. Electronmicroscopy of cellular division in Escherichiacoli. J. Bacteriol. 83:544-550.
EDWARDS, M. R. 1963. Electron microscopic ob-servations of the morphological characteristicsof Listeria monocytogenes, p. 210-220. In M1. L.Gray [ed.], Proc. 2nd Symp. Listeric Infection,Montana State College, Bozeman.
FITZ-JAMES, P. 1964. Thin sections of dividingNeisseria gonorrhoeae. J. Bacteriol. 87:1477-1482.
GIMP,NEZ, D. F. 1964. Staining rickettsiae in yolksac cultures. Stain Technol. 39:135-140.
GLAUERT, A. MI. 1962. The fine structure of bac-teria. Brit. Med. Bull. 18:245-250.
KELLENBERGER, E., A. RYTER, AND J. SfiCHAUD.1958. Electron microscopic study of DNA con-taining plasma. II. Vegetative and maturephage DNA as compared with normal bacterialnucleoids in different physiological states. J.Biophys. Biochem. Cytol. 4:671-678.
LEE, S. 1960. Fixation of E. coli spheroplasts forelectron microscopy. Exp. Cell Res. 21:252-255.
MILLONIG, G. 1962. Further observations on aphosphate buffer for osmium solutions in fixa-tion. Proc. Intern. Conf. Electron Microscopy,5th, 2:8.
MOULDER, J. W. 1962. The biochemistry of intra-cellular parasitism. The University of ChicagoPress, Chicago.
OGURA, M. 1963. High resolution electron micros-copy on the surface structure of Escherichiacoli. J. Ultrastruc. Res. 8:251-263.
OSBORN, M. J., S. M. ROSEN, L. ROTHFIELD, L. D.ZELEZMILK, AND B. L. HORECKER. 1964. Lipo-polysaceharide of the gram-negative cell wall.Science 145:783-789.
PLOTZ, H., J. E. SMADEL, T. F. ANDERSON, ANDL. A. CHAMBERS. 1943. Morphological structureof rickettsiae. J. Exp. Med. 77:355-358.
REYNOLDS, E. S. 1963. The use of lead citrate athigh pH as an electron-opaque stain in electronmicroscopy. J. Cell Biol. 17:208-211.
Ris, H., AND J. P. Fox. 1949. The cytology ofrickettsiae. J. Exp. Med. 89:681-686.
ROBERTSON, J. D. 1959. The ultrastructure ofcell membranes and their derivatives. Biochem.Soc. Symp. 16:3-43.
RYTER, A., AND E. KELLENBERGER. 1958. Etudeau microscope 6lectronique de plasmas conte-nant de l'acide d6soxyribon-nucleique. I. Lesnucleoides des bact6ries en croissance active.Z. Naturforsch. 13:597-599.
494 J. BAICTERIOL.
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

FINE STRUCTURE OF R. QUINTANA
SABATINI, D. D., K. BENSCH, AND R. J. BARRNETT.1963. Cytochemistry and electron microscopy.The preservation of cellular ultrastructure andenzymatic activity by aldehyde fixation. J.Cell Biol. 17:19-58.
SALTON, M. R. J. 1964. The bacterial cell wall.Elsevier, Amsterdam.
SCHAECHTER, M., A. J. TousIMIs, Z. A. COHN.H. ROSEN, J. CAMPBELL, AND F. E. HAHN.1957. Morphological, chemical, and serologicalstudies of the cell walls of Rickettsia mooseri.J. Bacteriol. 74:822-829.
SHEPARD, C. C., AND R. W. G. WYCKOFF. 1946.The nature of the soluble antigen from typhusrickettsiae. Public Health Rep. 71:761-767.
SMITH, J. D., AND M. G. P. STOKER. 1951. Thenucleic acids of Rickettsia burnetii. Brit. J.Exp. Pathol. 32:433-441.
STOKER, M. G. P., K. M. SMITH, AND P. FISET.1956. Internal structure of Rickettsia burnetiias shown by electron microscopy of thin sec-tions. J. Gen. Microbiol. 15:632-635.
TOUSIMIS, A. J. 1956. Intracellular Rickettsiatsutsugamushi in tissue culture cells as studiedby ultrathin sectioning and electron micros-copy. J. Appl. Phys. 27:1392-1393.
TRUMP, B., E. A. SMUCKLER, AND E. P. BENDITT.1961. A method for staining epoxy sections for
light microscopy. J. Ultrastruct. Res. 5:343-348.
VALENTINE, R. C., AND R. W. HORNE. 1962. Anassessment of negative staining techniques forrevealing ultrastructure, p. 263-278. In R. J. C.Harris [ed.], The interpretation of ultrastruc-ture. Academic Press, Inc., New York.
VAN ITERSON, W., AND W. LEENE. 1964. A cyto-chemical localization of reductive sites in agram-negative bacterium. J. Cell Biol. 20:377-388.
VAN ROOYTEN, C. E., AND G. D. SCOTT. 1949.Electron microscopy of typhus rickettsiae. Can.J. Res. 26E:250-253.
VINSON, J. W., AND H. S. FULLER. 1961. Studieson trench fever. I. Propagation of rickettsia-like microorganisms from a patient's blood.Pathol. Microbiol. 24(Suppl.):152-166.
WEIGL, R. 1920. Untersuchungen und Experi-mente an Fleckfieberlausen. Die Technik derRickettsiae-Forschung. Beitr. Klin. Infektion-skrankh. 8:353-376.
WISSIG, S. L., L. G. CARO, E. G. JACKSON, ANDJ. E. SMADEL. 1956. Electron microscopic ob-servations on intracellular rickettsiae. Amer. J.Pathol. 32:1117-1133.
WOLBACH, S. B., J. L. TODD, AND F. W. PALFREY.1922. The etiology and pathology of typhus.Harvard University Press, Cambridge, Mass.
VOL. 89, 1965 495
on June 1, 2020 by guesthttp://jb.asm
.org/D
ownloaded from