structure-activity relationships in acetylcholinesterase reactions : hydrolysis of non-ionic acetic...
TRANSCRIPT

Eur. J . Biochem. 67, 315-322 (1976)
Structure-Activity Relationships in Acetylcholinesterase Reactions Hydrolysis of Non-ionic Acetic Esters
Jaak JARV, T6nu KESVATERA, and Aavo AAVIKSAAR
Tartu State University, Chemistry Department, Tartu, and Institute of Cybernetics of the Academy of Sciences of the Estonian SSR, Biochemistry Department, Tallinn
(Received April 14. 1976)
The Michaelis-Menten parameters k,,,, Ks(app) and the second-order rate constants kll = kz/K, of acetylcholinesterase-catalyzed hydrolysis of 25 acetic esters with non-ionic leaving groups have been determined at 25 "C and pH 7.5 in 0.1 5 M KCI. A linear relationship between the substrate non- covalent binding capacity and the leaving group hydrophobicity, and a multiparameter correlation of the acetylation reaction rate constant logarithm with the leaving group inductive effect, hydro- phobicity, and steric effect, have been established. The acetyl-enzyme deacetylation rate constant has been calculated. Taken together, a fairly complete understanding of acetylcholinesterase specificity is possible.
The data are consistent with a model of the acetylcholinesterase active site, in which the catalyti- cally active groups are located at the bottom of a jaws-like slit with a limited range of hydrophobic walls that provide the sorption of the substrate leaving groups not longer than that in n-butyl acetate.
Acetylcholinesterase has been shown to hydrolyse, in addition to its specific substrate acetylcholine, many other acetic esters [l]. This provides a prospect for extensive study of the influence of substrate leaving group structure on the effectiveness of acetylcholin- esterase-catalyzed hydrolysis. A conclusion that both the substituent inductive effect and hydrophobicity are important in acetylcholinesterase reactions, can be drawn from the results of previous studies [2- 121. In this paper we are concerned with quantitative evalua- tion of these influences in the reaction series CH3C- (O)OX, where the group X includes hydrocarbon chains and various non-ionic electronegative substit- uents (see Table 1).
Up to now the analysis of enzyme kinetics data by means of correlation equations has been carried out mainly on chymotrypsin-catalyzed reactions [13 - 161. In accordance with the three-step reaction scheme
E + S+ES (1) ES 5 EA $- PI
EA -% E + P2 (2)
(3) the structure-activity relationship for the non-co- valent binding step (3) has been given by the equation
P K = PKt + qbn, (4) ___
Enzymes. Acetylcholinesterase or acetylcholine acetyl-h ydrolase (EC 3.1.1.7); chymotrypsin (EC 3.4.21 .I); trypsin (EC 3.4.21.4).
where K, = k-l/kl, while E is enzyme, S substrate, ES the enzyme-substrate complex, EA the acyl-en- zyme, PI and P2 the first and second products. For the acylation reaction (2), as well as for the deacylation (3), the multiparameter correlation equation
log k2 = log k2" + @* O* + van + 6 E, ( 5 )
can be applied. The parameters e*, q and 6 in the equations are the intensity factors of the inductive effect, hydrophobicity and steric effect, respectively. The constants cr* and E, have their usual meanings as in physical organic chemistry [17]. The hydrophobicity constants n have been introduced by Hansch [18,19].
Under the conditions where the product and substrate inhibition phenomena could be left out of consideration, the reaction sequence (1 - 3) would be applicable to the acetylcholinesterase-catalyzed hydro- lysis of esters [20]. According to the reaction scheme the meaning of the constants kcat and Ks(app) (K, = K, if k2 < k-l) in the Michaelis-Menten equation
depends on the values of the ratio kz/kj as follows :
(7)

316 Acetylcholinesterase Specificity
Although for some substrates the possible values of k2/k3 have been quoted [20-221, the question of the limiting step in the acetylcholinesterase-catalyzed hydrolysis of carboxylic esters is still not clear. Therefore the only meaningful steady-state kinetic parameter a priori applicable in the structure-activity correlation analysis of acetylcholinesterase reactions is the second-order rate constant kll = kZ/K,. By combining Eqns (4) and (5), the following relationship is obtained for krr:
The value of k3 in Eqns (7) and (8) remains unchanged for the substrates with various leaving groups. This allows us to evaluate k s on the basis of the steady- statekinetic data for the systematic series of substrates CH3C(0)OX, as demonstrated below.
MATERIALS AND METHODS
Chemicals
Acetylcholinesterase from cobra Naju naja oxiana venom was used after gel filtration on a Sephadex G-75 column. The specific activity of the enzyme prepara- tions was about 200 IU (1 IU = 1 pmol acetylcholine min-' x mg protein-'). Enzyme stock solutions were made in 0.15 M KC1. These solutions could be kept at +4 "C for several weeks without any change in activity. The operational normality of the solutions was calculated from the initial rate of the enzymic hydrolysis of acetylcholine (2.3 mM) at 25 "C at pH 7.5 in 0.15 M KC1, using the molecular activity of the enzyme a, = 6.33 x lo3 s-l [23].
The acetic esters were obtained from Reakhim (USSR) or synthesized by common methods from the appropriate alcohols or bromides [24]. All esters were rectified before using. The boiling points and refractive indexes corresponded to literature values [25 - 281. The purity of the esters with boiling points below 150 "C (atmospheric pressure) was checked by gas- liquid chromatography.
The compounds CH~C(O)O(CHZ),SCZH~ (n = 2-4) which have not been described earlier, were prepared analogously to CH~C(O)O(CHZ)ZSCH~ [29] and had satisfactory elemental analyses and 13C nuclear magnetic resonance spectra.
Substrate stock solutions were made in 0.15 M KCI immediately before use. The amount of the acetic acid liberated on complete hydrolysis of the esters was in agreement with substrate concentrations based on the weight of the samples.
Acetylcholine chloride from Merck was used after recrystallization from absolute ether/alcohol mixture. Other reagents used were of analytical grade.
Kinetic Measurements The rates of the enzymic hydrolysis of acetic
esters were followed titrimetrically (Radiometer Titri- graph TTTlc/SBR2c/ABUI, Denmark) at pH 7.5 and 25 "C in 0.15 M KCl. 0.01 M KOH solution was used in titration, which was performed in a closed vessel with argon atmosphere in order to avoid the absorption of atmospheric COz. The reaction mixture consisted of 5 - 30 ml substrate solution to which 0.01 - 0.5 ml of the enzyme solution was added. In determining the initial rates of hydrolysis the reaction was followed during 4-5 min. Data processing was based on Eqn (6), the kcat and K,(,,,) values were calculated by the statistical method of Wilkinson [30]. The second- order rate constants were calculated from the pseudo- first-order rate constants obtained by the differential method of Rudakov [31] from the first-order kinetic curves at low substrate concentration, [S] < Ks(,pp). The reactions were followed for at least three half- life periods and three different enzyme concentrations were used for each substrate. Product inhibition was shown to be negligible at the concentrations used.
Substituent Constants ,for Structure-Activity Correlurions
The substituent constants are listed in Table 1. In taking into account the inductive effect of electro- negative substituents we have used Taft o* constants [17,32]. For the alkyl groups the constants modified by Palm et al. [33] were used. In Palm's modification an infinitely long polymethylene chain is considered as the reference substituent with o* = 0. For - CH3 and -C2H5 o* > 0 and for other alkyl groups O* is equal to zero within the error limits. This enables one to treat the alkyl groups and electronegative substituents in a common system, since the o* values of both types of groups approach the same limit as the inductive effect attenuates [33].
The steric constants E, were chosen according to the isostericity principle [34], the oxygen atom in the alcohol portion of the esters was taken to be equivalent to a CHz group.
The hydrophobicity constants were calculated additively from the 'aliphatic' 71 values [18,19].
Calculations
All calculations in the statistical treatment of experimental data were performed on a Nairi 2 com- puter (we are indebted to Dr H. Kiis for the programs).
RESULTS
Kinetic Constants
The values of k ~ r , k,,, and KlCapp) are listed in Table 1. The data indicate that the second-order

J . Jarv, T. Kesvatera, and A. Aaviksaar 317
Table 1. Kinetic constantsfhi. the acctylcholinesteru.ce-cuta1~~~~~ hydi.olj1si.s of acetic esters C H s C ( 0 ) O X in 0.15 M KCI ut p H 7.5 und 25 ' C The k,,, and Kqc,,,, were determined from the initial rate measurements. The second-order rate constants k j ~ were determined under the pseudo- first-order conditions where [S] < Ks(app). The values for rr* were taken from other work 132,331; CJ* for CHzCsCH was calculated from the kinetic data in [36]. The values for z werc calculated by the additive scheme [18, 191; I[ for NO2 was taken from other work (351. L values for the groups OX were taken to be equal to those of the CHzX groups [34]
Com- X pound
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 I8 19 20 21 22 23 24 25
M - l s - '
0.10 f 0.02 0.23 0.04
5.23 fe;'O.10 5.49 i-3 0.17 4.78 * 0.18 5.60 * 0.20
21.00 * 0.55 2;09 k 0.06 0.052 0.003 0.49 & 0.02 5.19 f 0.25
0.81 4 ,0.05
200.0 & 8.6
10.02 <0.02 530 10 22.5 & 1.0
6.46 & 0.19 1.26 k 0.09
16.10 k 0.90 10.44 k 0.52 25.2 k 1.0 4.60 0.23 4.65 k 0.20 5.34 k 0.24
S - '
-
2.0 0.9 3.7 * 0.4 9.11 f 0.45
10.5 k 3.2 8.3 If: 3.0
32.4 If: 2.0 17.5 , f 2.5
-
5.00 f 0.30
0.582 0.066 3.16 f 0.15
-
-
-
93.7 f 1.0 48.0 k 2.2 49.0 f 2.2 10.0 f 2.7
35.5 k 2.0 35.5 _+ 3.3 33.4 * 2.0
-
6.50 +_ 0.70 8.52 F 0.43
M -
10 * 5 5.5 f 2.1 1.74 _+ 0.36 1.28 f 0.24 1.7 f 1.0
i8.140 +_ -0.005 0.6&0 & 0.0.57 2.33 .' $ 0.10
1.80 * 0.10 0.64 0.03
-
-
-
-
0.38 * 0.02 2.35 f 0.10 7.25 & 0.20 8.03 k 0.60
4.75 f 0.27 1.48 k 0.11 0.64 f 0.02 1.50 0.30 1.49 k 0.35
-
0.20 0.50 0.07 1 .OO 0 1.50 0 2.00 0 2.50 0 3.00 0 3.50 0 2.98 0 2.30 0 1.80 0 1.30 0 1.80 0 2.51 0 1.98 0 2.48 0.60 2.13 0.56 1.45 0.55 0.94 0.23 1.23 0.62 1.31 0.39 1.39 0.22 1.95 0.25 2.26 0.08 2.45 0.03 2.95
0.07 0.36 0.39 0.40 0.30 0.40 0 40 0.40 0.43 0.35 0.93 0.93 0.98 1.74 1.74 0.38 0.44
-0.60 20.20
0.40 0.48 0.44 0.38 0.40 0.40
rate constants k I l = kz /K, , obtained under the pseudo- first-order conditions are considerably more precise than the constants calculated as the ratio of k,,, to K,(,,,,. For some esters of low reactivity and poor solubility a separate determination of kcat and Ks(app, was not possible by the conventional methods used in the present work. In the case of nitroethyl acetate the enzymic reaction was accompanied by rapid spon- taneous hydrolysis of the compound.
Range of Hydrophobic Binding Site
In the study of the topography of acetylcholin- esterase active site hydrophobic region for the binding of the substrate leaving group, a subseries of alkyl acetates, in which the hydrophobicity is the only changing substituent parameter, was used. The plot of log k11 versus 7t for the alkyl acetates is presented in Fig. 1. The relationship splits into two separate straight lines which have a point of intersection at 7t = 2. However, it is evident from the data that the substituent hydrophobicity is not critical for the breaking point of log k r l versus n plot. The straight
0 1 .O 2.0 3.0 4.0 li
Fig. 1. Plot of log k,, ugninst n ,for the acetykholin~sterase- catnlyzed hydrolysis of alkyl acetates CH3C(O)OX. (@) Acetic esters of primary alcohols; (0) acetic esters of secondary alcohols. The numbers in the figure refer to the substrates listed in Table 1

318 Acetylcholinesterase Specificity
line with a slope 1.65 (see Table 2), which can be expressed by a simplified form of Eqn (9),
logkr1 = c + qn:, (10) includes the substrates with the n: values considerably greater than 2, if the length of the alkyl substituent does not exceed that of the n-butyl group. The rate constants of the compounds with longer alkyl chains remain practically unchanged although the substituent hydrophobicity increases. We take these results as an indication of the limited size of the enzyme hydrophobic binding site the length of which corre- sponds to that of the n-butyl group in the alcohol moiety of acetate. The results obtained here agree with earlier data [7-91 on the initial velocities of enzymic hydrolysis of alkyl acetates by cobra venom, pigeon brain and horse erythrocyte acetylcholin- esterases which also reveal a break at n-butyl acetate.
Leu v ing Group lnduc t ive Effect
In the quantitative estimation of the role of inductive effect in acetylcholinesterase-catalyzed hy- drolysis, the second-order rate constants of the esters with electronegative substituents were used. Provided that the steric effect remains constant, Eqn (9) can be simplified to
Taking into consideration the limited range of the hydrophobic binding area, we used in the correlation only the substrates with the leaving groups not longer than that of n-butyl acetate. For the compounds that satisfy this requirement (compounds 1, 2, 16-21 in Table l), no mutual statistically relevant inter-relation- ship exists between a* and 71 ( r = 0.371), and so the
data can be processed by a two-parameter equation. The results of the correlation by Eqn (1 1) are given in Table 2. It is evident that the constants C and cp obtained for the substrates with electronegative leaving groups coincide with the corresponding values for the alkyl acetate subseries (substrates 3 , 4, 8-10 in Table 1). This serves as a proof of similarity in the interaction of the electronegative and alkyl substituents with the acetylcholinesterase surface. It should be noted that log kIl for phenyl acetate, the only aromatic compound investigated, fits the correlation by Eqn (11) and therefore the phenyl group must also bind in the hydrophobic slit.
Relutionship between Binding and Acetylation
The more than 100-fold variation of the k,,, values in Table 1 indicates that enzyme deacetylation cannot be the rate-limiting step and that at least for the esters from the lower reactivity side of the series the rate constant kZ must be sufficiently less than k3,
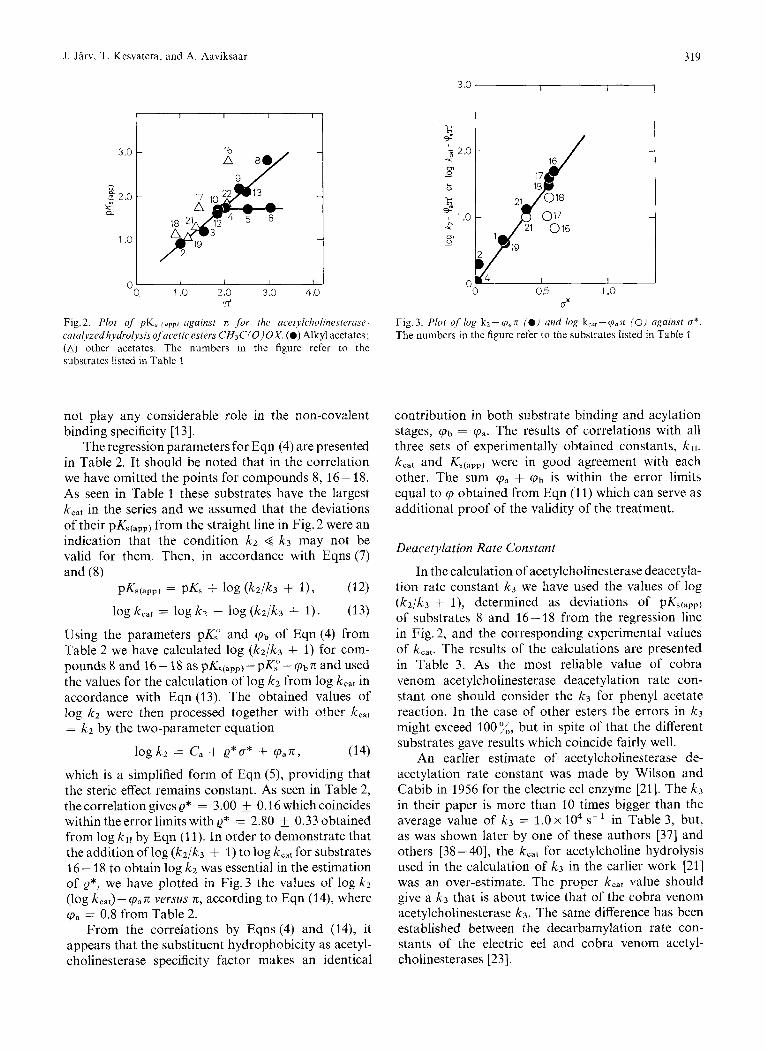
so that k,,, = kz, Ks(app) = K,. It makes a study of the relationship between binding and reaction specificities in terms of Eqns (4) and (5) possible. In order to check the applicability of Eqn (4) for acetylcholinesterase all the values of pKs(app) obtained have been plotted against TC in Fig. 2. For alkyl acetates with the pKs(app) indicated separately as filled circles we get the relation- ship of the same form as with log k11 in Fig. 1. The values of pK,(,,,, of the substrates with the leaving group alkyl substituents longer than n-butyl group remain constant, and the pKs(app) for the compounds with the appropriate length of the chain give a straight line which includes the substrates with electronegative substituents as well. It is consistent with the main generalizing idea of Eqn (4) that electronic effects do
Table 2. Structure-activity correlations .fov the acetylcholinestera.se-cuta!vzed hydrolysis of acetic esters, CH,C(OjOX The numbers of the compounds included in the analyses refer to the substrates in Table I . The values of correlation coefficients r and standard deviations s are given
Equation Constants Y S Compounds used in correlation
logkrl = c + cpn (13) cp = 1.65 k 0.07 0.997 0.076 3 ,4 ,8-10 C = 1.43 S_ 0.15
log kir = C + Q * U * + c p p n (14) e* = 2.80 k 0.33 0.994 0.152 1 ,2 , 16-21 q = 1.64 i 0.16 C = 1.55 k 0.15
PK, = PK: + vbn (4) ( ~ b = 0.88 k 0.06 0.983 0.087 2-4,9,10,12,13,19,21 pK," = 0.06 k 0.11
log k2 = C, + Q * U * + <pspn (17) Q* = 3.00 f 0.16 0.991 0.112 2-4, 10, 11, 16-19,21 cp. = 0.80 * 0.10 C, = 1.40 k 0.16
(9) p = 1.61 _+ 0.10 6 = 1.07 k 0.27 lor k z = 1.84 +_ 0.19
log k i i = log kfi + Q * U * + pi^ + 6Es e* = 3.00 _+ 0.33 0.9992 0.217 1-4,8-13, 16-21
J. Jarv, T. Kcsvatera, and A. Aaviksaar 319
0 1 .0 2.0 3.0 4.0 T t
0
Fig.2. Plot of pK, ( u p p ) against n for the acetylcholinesterasr- Fig.3. Plot of log kz-cp,n (e) and log kcnr-cpun (0) ugainst o*. catalLzedh~vdrolysisofacetic esters CH3C(O)OX. (e) Alkyl acetates; (A) other acetates. The numbers in the figure refer to the substrates listed in Table 1
The numbers in the figure refer to the substrates listed in Table 1
not play any considerable role in the non-covalent binding specificity [13].
The regression parameters for Eqn (4) are presented in Table 2. It should be noted that in the correlation we have omitted the points for compounds 8, 16- 18. As seen in Table 1 these substrates have the largest k,,, in the series and we assumed that the deviations of their pK,(,,,) from the straight line in Fig. 2 were an indication that the condition kz < k3 may not be valid for them. Then, in accordance with Eqns (7) and (8)
PKs(app) = pKs + 1% (k2/k3 + I),
log kcat = log kz - log (k2/k3 + 1).
(12)
(13) Using the parameters pK," and (pb of Eqn (4) from Table 2 we have calculated log (k2/k3 + 1) for com- pounds 8 and 16-18 aspK,~,,,)-ppK,"-(pbn andused the values for the calculation of log k2 from log k,,, in accordance with Eqn (13). The obtained values of log k2 were then processed together with other k,,, = kz by the two-parameter equation
logkz = Ca + Q*O* + (pan, (14) which is a simplified form of Eqn ( 5 ) , providing that the steric effect remains constant. As seen in Table 2, the correlation gives Q* = 3.00 0.16 which coincides within the error limits with Q* = 2.80 +_ 0.33 obtained from log by Eqn (1 1). In order to demonstrate that the addition of log (k2/k3 + 1) to log k,,, for substrates 16 - 18 to obtain log k2 was essential in the estimation of e*, we have plotted in Fig. 3 the values of log k2 (log kcat)- (pan versus z, according to Eqn (14), where (pa = 0.8 from Table 2.
From the correlations by Eqns (4) and (14), it appears that the substituent hydrophobicity as acetyl- cholinesterase specificity factor makes an identical
contribution in both substrate binding and acylation stages, (pb = (pa. The results of correlations with all three sets of experimentally obtained constants, k ~ , k,,, and Ks(,,,) were in good agreement with each other. The sum (pa + (Pb is within the error limits equal to (p obtained from Eqn (1 1) which can serve as additional proof of the validity of the treatment.
Deucetylution Rate Constant
In the calculation of acetylcholinesterase deacetyla- tion rate constant k3 we have used the values of log (kZlh-3 + l), determined as deviations of pK,(,,,, of substrates 8 and 16- 18 from the regression line in Fig. 2, and the corresponding experimental values of kcat. The results of the calculations are presented in Table 3. As the most reliable value of cobra venom acetylcholinesterase deacetylation rate con- stant one should consider the k3 for phenyl acetate reaction. In the case of other esters the errors in k3 might exceed loo%, but in spite of that the different substrates gave results which coincide fairly well.
An earlier estimate of acetylcholinesterase de- acetylation rate constant was made by Wilson and Cabib in 1956 for the electric eel enzyme [21]. The k3 in their paper is more than 10 times bigger than the average value of k3 = 1.0 x lo4 s-' in Table 3, but, as was shown later by one of these authors [37] and others [38 - 401, the kcat for acetylcholine hydrolysis used in the calculation of k3 in the earlier work [21] was an over-estimate. The proper k,,, value should give a k3 that is about twice that of the cobra venom acetylcholinesterase k3. The same difference has been established between the decarbamylation rate con- stants of the electric eel and cobra venom acetyl- cholinesterases [23].

3 20 Acetylcholinesterase Specificity
Table 3. Calculation oy the acetylcholinesterase deacetylation rute constant k3
The values of log (k2/k3 + 1) were found as pK,c,,,,-pK,”--cpbn
Substrate 1% (kz/k3 + 1) x k3
S-l
CH3C(O)OC6H5 0.86 1.1 CH~C(O)OCHZSC~H~ 0.35 0.9 CH3C(O)OCHzC=CH 0.30 1 .0 CH~C(O)OCZH~C(CH~)~ ~ 0 . 2 -0.9
Steric Effect
Low reactivity of the secondary and tertiary alcohol esters (see Table 1) refers to the necessity of considering the steric effect. The data in Fig. 1 show that log k,, obtained for these esters are significantly lower than required by Eqn (10). However, in the plot of PK,(~,,, versus n the points for these substrates lie on the same straight line with other esters (see Fig.2). This proves that only the acetylation step (2) is sensitive to the steric effect and the second-order rate constants kI1 may be used in determining the constant 6.
The results of the total three-parameter correlation of all k1, values obtained in this work are given in Table 2. It is evident from the data that the inclusion of the secondary alcohol esters into the correlation does not alter the Q and cp values which had been obtained from Eqns (10) and (11) as special cases of Eqn (9) for the selected subseries of the substrates.
DISCUSSION
It can be concluded from the results that the three-step reaction scheme (1 - 3) agrees with the kinetic data of the acetylcholinesterase-catalyzed hydrolysis of acetic esters with non-ionic leaving groups. The rate constant k3 for the deacetylation reaction (3) was determined to be 1.0 x lo4 s-’ at 25 “C and pH 7.5 and thus k2 and Ks values for all acetic esters could be evaluated on condition that the reaction mechanism remains unchanged within the series.
For phenyl acetate, kzlk3 was equal to 6 and it was the only substrate for which the deacetylation reaction may be considered to be the rate-limiting step. For most alkyl acetates kz was significantly less than k3 and so the formation of acetyl enzyme was the rate- limiting step in their hydrolysis as it has been assumed in earlier studies [22,41].
The independence of binding effectiveness and reactivity of the acetates with moieties longer than n-butyl in their leaving part from the substituent hydrophobicity seems to be consistent with the model
3 ’.O t
0 1 .o 2.0 2.0 - 1 .o f i x - fii(CH313
Fig. 4. Plot of log [k:r/k,y:CH3~.] ugainst xx - x‘,,,,, for the acetj.1- choi ines terase-catu l~~~~ hydro1j.sr.r of CH3CIO) OC2H4X. X stands
for N(CH3)3-,,Hn (0) and N(CHj)*R (0); R is a normal hydro- carbon radical; (0) denotes the reference compound, acetylcholine
+ +
of acetylcholinesterase active site in which the jaws- like slit with the catalytic site at the bottom opens into the reaction medium. In order to demonstrate that the leaving groups of acetylcholine-like ionic substrates and alkyl acetates bind at the same hydrophobic slit the logarithms of the relative second- order acetylation rate constants for the substrates
CH3C(O)OC2H4X with X = -N(CH3)3-,,Hn [21]
and -N(CH3)zR [42], where R stands €or a normal alkyl group have been plotted against nx - ~ - & ( c . H ~ ) ~
in Fig.4. The relationship is in good agreement with that for alkyl acetates in Fig. 1.
On the other hand, a well-defined difference be- tween acetylcholine derivatives and alkyl acetates in their interactions with acetylcholinesterase active sur- face has been found in the present work. In contrast to the interaction with acetyl p-methylcholine (I) [43] the enzyme did not exhibit stereoselectivity in the case of see-butyl acetate (11) which has an asym- metric centre similar to that of compound (I). The enzymic hydrolysis of sec-butyl acetate under the
0 II * +
I CH3
I
H3C - C - 0 - CH - CH2 - N(CH3)3
0 II *
I CH3
H3C-C-O-CH-CH2 -CH3
IT

J. Jiirv, T. Kesvatera. and A. Aaviksaar 321
conditions where [S] @ Kc(app) was kinetically first- order, and the amount of acid released corresponded to the weighed concentration of the substrate. Thus acetylcholinesterase appears to be more sensitive to the stereochemical factors of ionic substrates that may be caused by the interaction of these compounds with the ‘anionic point’.
The equality of substituent hydrophobic influence intensity factors in binding and acetylation stages has been forinerly discovered in the reactions of chymotrypsin with the substrates [44] and organo- phosphorus quasisubstrates [13] with varying acyl portions. In the present work the equality of V)b and qd has been demonstrated to hold in the case of another serine enzyme, acetylcholinesterase, and con- cerning the substrates with varying leaving portion. In accordance with the explanation given elsewhere [I31 we assume that the influence of hydrophobic interaction on the rate of the bond-breaking step of acetylcholinesterase acetylation is determined by en- zyme molecule conformational change that consists of ‘shutting‘ the hydrophobic slit in the active centre. The shutting process should be accompanied by a structural perturbation with a positive free energy change which is a part of the total activation energy. An additional contribution of the negative sign (in the free energy scale) hydrophobic interaction in the transition state as a result of the shutting of the slit makes the activated complex with the closed slit by the term qaz energetically more favourable than the activated complex with open slit. In the Michaelis complex and the acetyl-enzyme, as well as in the free enzyme, the Conformation with the closed slit is ener- getically unfavourable because it is accompanied by the above-mentioned structural perturbation. There- fore the corresponding equilibria for these forms are largerly shifted in favour of the conformation with the open slit [13].
In the studied interval of (T* values, the acetyl- cholinesterase-catalyzed hydrolysis of esters was char- acterized by positive @*. This agrees with the generally accepted views on the mechanism of a serine hydrolase action that involves nucleophilic attack on the carbonyl carbon of esters. Apparently, the acetylation step is assisted by several catalytic groups of the active site [45] which may lead to notable differences in the activated complex structure of the enzymic reaction compared with alkaline hy- drolysis. The significant difference in the @* values of the reactions refers to this. In the alkaline hydrolysis of acetates @* is about 2 [33,46] while Q* = 3 has been found for the acetylcholinesterase-catalyzed hydro- lysis. The steric effects in the alkaline and enzymic hydrolysis of the acetic esters cannot be compared at present since no reliable 6 was available for the former reaction.
The contributions of substituent inductive and steric effects and hydrophobicity in the acetylcholin- esterase-catalyzed hydrolysis of non-ionic acetates, CH3C(O)OX, are hopefully considered to be appli- cable in further research on quantitative characteriza- tion of the role of acetylcholinesterase anionic site in the reaction with charged substrates.
REFERENCES
1.
2. 3.
4.
5. 6. 7.
8. 9.
10. 11.
12.
13.
14 15.
16.
17
18.
19
20. 21.
22.
23.
24.
25.
26 27
28 29
30 31
Cohen, J . A. & Oosterbaan, K. A. (1963) in Handbuch der Experimentellen Pharmucologie, Erg.-Werk XV (Koclle, G. B., ed.) pp. 299 - 373, Springer-Verlag, Berlin-Gottingen-Heidel- berg .
Adams, D. H. (1949) Biochim. Biophys. Acta, 3, 1 - 14. Adams, D. H. & Whittaker, V. P. (2949) Biochim. Biophys.
Adams, D. H. & Whittaker, V. P. (1950) Biochim. Biophys.
Wilson, I . B. (3952) J . Riol. Chem. 197, 215-225. Bergman, F. & Segal, R. (1954) Biochetn. J . 58, 692-698. Mounter, L. A. & Whittaker, V. P. (1950) Biochez. .I. 47,
Mounter, L. A. (1951) Biochem. J . SO, 122-128. Whittaker, V. P. (1953) Biochem. .I. 54, 660-664. Aldridge, W. N. (1954) Chem. Ind. 473-476. Kitz, R. J., Ginsburg, S. & Wilson, I . R . (1967) Mol. Phannu-
Fukuto, T. R. & Mctcalf, R. L. (1956) J . Agric. Food (‘hem. 4,
Aaviksaar, A., Paris, J . & Palm. V. (1971) Org. Reucf. 8, 817-
Hansch, C. & Coats, E. (1970) J . Phurm. Sci. 59, 731 -743. Dupaix, A, , Bechet, J.-J. & Roucous. C. (1973) Biochemistry,
Klyosov, A. A. (1971) in Proc. Sth Anniversary Meeting Lah. Bioorg. Chem., Lonionosov Strile University, Moscow (Belo- zersky, A. N. & Bogdanov, A. A., eds) pp. 263 - 271, Lomo- nosov State University, Moscow.
Taft. R. W., Jr (1956) in Steric Effects in Organic C‘hemistri, (Newman, M . S., ed.), Chapter 13, Wiley, New York.
Leo, A,, Hansch, C. & Elkins, D. (1971) Chem. Rev. 71, 525- 616.
Fujita. T., Iwasa, J. & Hansch, C. (1964) J . Am. Chem. Soc. 86,
Krupka, R. M . (1964) Can. J . Biochem. 42, 677-693. Wilson, I . €3. & Cabib, E. (1956) .I. A m . Chem. Sot.. 78, 202-
Hellenbrand. K. & Krupka, R. M. (1970) Biocheniistrv, 9,
Jarv, J . L., Aaviksaar, A. A, , Godovikov, N . N., Langel, U . L. &Pas t , U . E. (1976) Biokhirniya, 41, 827-835.
Weygand-Hilgetag (1964) Orgunisch-chrmi.rche E.xppcrirnentieu- kunst vol. 3, 3rd edn (Hilgetag, G. & Martini, A., eds) pp. 442-452. Johann Ambrosius Barth, Leipzig.
Heilbron, I . & Bunbury, H. M., eds (1946) Dictionary of’Orgtmic Compounds vol. 1-3, 2nd edn, Eyre, London.
Birch, A. J. (1949) J . Chem. Soc. 2721-2722. Bohme, H., Fisher, H. & Frank, R. (1949) Lidig.7 Ann. Chem.
Gold, M. H. ( 3 946) J . A m . Chem. Soc. 68, 2545 - 2549. Challenger, F. & Sinipson, M. I. (1948) J . Chem. Soc. 1591 -
Wilkinson, G. N. (1961) Biochem. J . 80, 324-330. Rudakov, E. S. (1960) Kinetika i Kutaliz, I , 177- 187.
A c ~ u , 3, 358-366.
Actu, 4 , 543- 558.
525 - 530.
C o l . 3, 255- 232.
930-935.
832.
12.2559-2566.
5175- 51 80.
207.
4665 - 4672.
”
563, 54-72.
1597.

322 J. Jarv, T. Kesvatera, and A. Aaviksaar : Acetylcholinesterase Specificity
32. Koppel, I. A,, Karelson, M. M. & Palm, V. A. (1973) Org.
33. Palm, V. A,, Piissa, T. O., Nummert, V. M. & Talvik, I. V.
34. Talvik, I . V. & Palm, V. A. (1971) Org. React. 8, 445-462. 35. Fastrez, J. & Fersht, A. R. (1973) Biochemistry, 12, 1067-
36. Grob, C. A. & Schlageter, M. G. (1974) Helv. Chim. Acta, 57,
37. Wilson, 1. B. & Harrison, M. A. (1961) J . Biol. Chem. 236,
38. Berman, J . D. (1973) Biochemistry, 12, 1710- 1715. 39. Rosenberry, T. L., Chang, H. W. & Chen, Y. T. (1972) J .
React. 10,497-514.
(1973) Org. React. 10, 243 - 261.
1074.
509- 51 1.
2292 - 2295.
Biol. Chem. 247, 1555-1565.
40. Mooser, G., Schulman, H. & Sigman, D. S. (1972) Biochem-
41. Krupka, R. M. (1964) Biochemistry, 3, 1749- 1754. 42. Brestkin, A. P., Rozengart, E. V., Soboleva, L. N. & Khromov-
43. Beckett, A. H., Harper, N . J . & Clitherow, J . W. (1963) J .
44. Berezin, I . V., Kazanskdya, A. A., Klyosov, A. A. & Martinek,
45. Brestkin, A. P. & Rozengart, E. V. (1965) Nature (Lond.) 205,
46. Babayeva, L. G., Bogatkov, S. V., Kruglikova, R. I. & Un-
istry, 11, 1595-1602.
Borisov, N. V. (1975) Biokhimiya, 40, 95- 102.
Pharm. Pharmacol. 15,362 - 371.
K. (1971) FEBS Lett. 15, 125- 128.
388- 389.
kovsky, B. V. (1974) Org. React. 11, 465-416.
J. J i b , Orgaanilise keemia kateeder, Tartu Riiklik Ulikool, Kingissepa 14/16, Tartu, U.S.S.R. 202400
A. Aaviksadr and T. Kesvatera, Eesti NSV Teaduste Akadeemia Kiiberneetika Instituudi biokeemia sektor, pk 670, Tallinn. U.S.S.R. 200026