stress alters immune function and disease resistance in chinook salmon (oncorhynchus tshawytscha)
TRANSCRIPT

Stress alters immune function and disease resistance inchinook salmon (Oncorhynchus tshawytscha)A. G. Maule, R. A. Tripp, S. L. Kaattari and C. B. SchreckOregon Cooperative Fishery Research Unit, Department of Fisheries and Wildlife, and ""Department of
Microbiology, Oregon State University, Corvallis, Oregon 97331, U.S.A.
received 20 May 1988
ABSTRACT
We examined the effects of acute stress on the immunesystem and disease resistance of juvenile chinooksalmon (Oncorhynchus tshawytscha) in laboratory andclinical trials. Immune function, as measured by theability of lymphocytes from the anterior kidney togenerate specific antibody-producing cells (APC) invitro, was depressed 4 h after stress, when plasma cor-tisol levels were highest. At the same time, resistanceto the fish pathogen, Vibrio anguillarum, was alsodepressed. Compared with controls, plasma cortisoland APC of stressed fish were unchanged after 24 h,and disease resistance was enhanced as evidenced by
higher survival rate and longer mean time to death ofmortalities. After 7 days, even though numbers ofAPC were depressed, plasma cortisol concentrationand disease resistance did not differ from controls.This pattern was generally the same, independent ofthe type of stress applied: i.e. being held out of waterin a dipnet for 30 s, manipulation during hatcheryoperations for 4 h, or transportation for 9 h. Theseand earlier findings suggest that similar endocrine\x=req-\immune interactions operate in the mammalian andsalmonid systems during acute stress.Journal ofEndocrinology (1989) 120, 135\p=n-\142
INTRODUCTION
It is generally accepted that physical and psycho¬logical stress can impair immune function and lead todecreased disease resistance (Monjan & Collector,1977; Riley, 1981; Laudenslager, Ryan, Druganet al. 1983). Furthermore, neuroendocrine responsesappear to drive this stress-related immune dysfunction(Ader, 1981; Stein, Keller & Schleifer, 1985). Thesecretion of glucocorticoids, catecholamines and some
neuropeptides increases in response to stress, andthese hormones have wide-ranging and sometimesopposing effects on the immune system (Yu &Clements, 1976; Munck, Guyre & Holbrook, 1984;Spangelo, Hall & Goldstein, 1985). The adminis¬tration of glucocorticoids to animals or lymphocytecultures usually depresses the immune response.However, glucocorticoid treatment may enhance theimmune response, depending on the concentration ofthe steroid and physical state of the animal (Cupps,Edgar, Thomas & Fauci, 1984), timing of treatmentand immune assay (Cupps & Fauci, 1982), or immunefunction assayed (Goodwin & Atluru, 1986). Similarvariability in the response of the immune system afterstress has been reported. Gisler, Bussard, Mazie &Hess (1971) reported that, compared with non-
stressed controls, lymphocytes removed from mice 15min after acute stress had an increased ability togenerate antibody-producing cells (APC) to sheep redblood cells (SRBC) in vitro, but that the response was
depressed 6-72 h after stress. Okimura & Nigo (1986)and Okimura, Satomi-Sasaki & Ohkuma (1986a)demonstrated that acutely stressed mice had reducednumbers of splenic APC when immunized withSRBC, but when similarly stressed mice were immu¬nized with trinitrophenylated (TNP)-Ficoll or TNP-lipopolysaccharide (LPS) splenic APC increased. Thisvariability in response leads one to question whether a
single specific assay of immune function is an accurateindex of immune competence, the final measure ofwhich must be resistance to disease.
We previously demonstrated that artificially raisingplasma concentrations of cortisol in Pacific salmon(Oncorhynchus spp.) led to a reduction in their abilityto generate APC in vivo and their survival whenexposed to the fish pathogen Vibrio anguillarum(Maule, Schreck & Kaattari, 1987). We have alsoshown a direct action of cortisol on lymphocytes asthe addition of physiological concentrations of corti¬sol to lymphocyte cultures reduced APC (Tripp,Maule, Schreck & Kaattari, 1987). This cortisolinhibition was abrogated by the addition of super-

natants from antigen-stimulated lymphocyte cultures(Tripp et al. 1987) or recombinanl interleukin-1(Kaattari & Tripp, 1987) suggesting that cortisol isinterfering with lymphokine production or activity.It has also been shown that acute stress results inthe transient increase in circulating cortisol in sal¬mon (Strange & Schreck, 1978; Barton, Schreck &Sigismondi, 1986), and that stress and cortisol admin¬istration can cause leucocytopenia in fish (Pickford,Srivastava, Slicher & Pang, 1971; McLeay, 1973). Inthe present study, our objective was to correlatechanges in plasma cortisol concentration, the abilityof lymphocytes from the anterior kidney to generateAPC in vitro and disease resistance in acutely stressedchinook salmon (O. tshawytscha).
MATERIALS AND METHODS
Experimental animalsJuvenile spring chinook salmon (spring chinooksalmon are distinguished from fall chinook salmon byvirtue of the timing of their spawning migrations)were obtained from Eagle Creek (OR, U.S.A.) or
Abernathy (WA, U.S.A.) National Fish Hatcheriesand were maintained at either the Oregon StateUniversity/Smith Farm Experimental Hatchery orthe Fish Disease Laboratory at Corvallis, OR. Bothfacilities receive fish-pathogen-free well water at a
constant temperature of 12+1 °C. Fish were fedOregon Moist Pellet (OMP) commercial salmon foodat a rate of 1 to 2% body weight per day. Fish wereacclimated to these conditions for at least 30 daysbefore experiments were started. To distinguishgenetic influences from possible environmental fac¬tors, we conducted clinical trials at the Warm SpringsNational Fish Hatchery, Warm Springs, OR, U.S.A.and the Oregon Department of Fish and Wildlife,Round Butte Hatchery, Madras, OR, U.S.A. Thespring chinook salmon at these hatcheries were thesame genetic stock and were fed OPM, but there weredifferences in water temperature regimens. RoundButte water was constant at 13 °C, while water atWarm Springs varied seasonally (6-21 °C) anddiurnally (17-21 °C in summer).Hormone and immune assaysFish were sampled after rapid netting and transfer to alethal concentration of tricaine methanesulphonate(200 mg/1). This dose of anaesthetic inhibits stress-induced increases in plasma cortisol in salmon(Barton, Schreck, Ewing et al. 1985). After fish wereimmobilized by the anaesthetic, we severed the tailand collected blood from the caudal vasculature intoheparinized capillary tubes. Plasma was separatedfrom the blood by centrifugation, stored at
—
20 °C,
and later assayed for cortisol by a radioimmunoassaydescribed by Redding, Schreck, Birks & Ewing (1984).
The ability of lymphocytes to generate specific APCwas assessed using the in-vitro assay described byTripp et al. (1987). Briefly, anterior kidneys were
aseptically harvested and put into tissue culturemedium (TCM) composed of RPMI 1640 containingL-glutamine and bicarbonate (GIBCO, Grand Island,NY, U.S.A.) and supplemented with 10% (v/v) hybri-doma-screened fetal calf serum (B. A. Bioproducts,Walkerville, MD, U.S.A.), 100 mg gentamicin sul¬phate/1 (Sigma Chemical Co., St Louis, MO, U.S.A.),50 pmol 2-mercaptoethanol/l (MCB ManufacturingChemists, Inc., Cincinnati, OH, U.S.A.), 4-0 pmoladenosine/1, 40pmol cytosine/1, 9-0pmol thymidine/1and 9-0 pmol guanosine/1 (Sigma). Lymphocytes were
separated from surrounding tissue by gentle aspira¬tion with a 1 ml syringe and tissue debris was allowedto settle to the bottom of the test tube. Supernatantscontaining cells were collected, and cells were washedby centrifugation and resuspended to 2 IO7 cells/mlin TCM. We transferred 200 µ cell suspension to wellsof a 24-well, flat-bottom microculture plate (CorningGlass Works, Corning, NY, U.S.A.) and added 200 µ of either TCM (negative controls) or TCM containingthe antigen TNP-LPS (0-4 pg/ml; Jacobs & Morrison,1975). Cell cultures were incubated at 17 °C in an air¬tight gasbox (C.B.S. Scientific, Del Mar, CA, U.S.A.)with blood-gas mixture (10% 02, 10% C02 and 80%N2) and were fed 50 pi of feeding cocktail (Tittle &Rittenberg, 1978) every other day. After 9 days, cellswere harvested, washed by centrifugation, and resus¬pended in medium. Lymphocytes secreting anti-TNPantibodies were detected by Cunningham plaqueassay (Cunningham & Szenberg, 1968). We mixed100 pi lymphocyte suspension, 25 pi TNP-coatedSRBC (Rittenberg & Pratt, 1969) and 25 pi dilutedsteelhead trout (Salmo gairdneri) serum as a comp¬lement source and deposited the mixture in a
Cunningham slide chamber. During incubation (2 h at17 °C), anti-TNP antibody became bound to TNP-SRBC and activated the complement cascade, result¬ing in lysis of the surrounding TNP-SRBC. Thus,lymphocytes secreting anti-TNP antibody caused ahole, or plaque, in the surrounding TNP-SRBC.Plaques were counted using a low-power dissectingmicroscope, and were expressed in terms of APC permillion lymphocytes; lymphocyte numbers were deter¬mined with a Coulter Counter (Model Z.M., CoulterElectronics, Hialeah, FL, U.S.A.).Disease challenge tests
We tested disease resistance by the fish pathogenVibrio anguillarum, a marine bacterium to whichsalmon in fresh water presumably had not previouslybeen exposed. Duplicate treatment groups offish were

exposed to V. anguillarum (LS-174) using the water-borne infection method of Gould, O'Leary, Garrisonet al. (1978). Briefly, fish were exposed to 3^1 IO10 V.anguillarum bacteria in 20 litres of water for 20 min.Dead fish were removed at least every 12 h and weconfirmed that death was caused by V. anguillarum byisolating the pathogen from their kidneys. Data wereexpressed as mean time to death (MTD) and percenttotal mortality.
Laboratory experimentsTo assess the effects of stress on immune function, we
acutely stressed juvenile spring chinook salmon ontwo occasions (approximate weights: 25 and 40 g forexperiments in January and March 1986) by holdingthem in a dipnet, out of water, for 30-60 s andsampled them 4 h and 1,2,4 and 7 days after stress.In April and October 1986, we challenged similarlystressed fish (weights approximately 20 and 45 g) withV. anguillarum to determine whether the pattern seenin the APC data reflected the ability of fish to resistdisease. Groups of 25-40 fish were acclimated to 65litre cuboidal tanks for 2 weeks. We acutely stressedthree randomly selected groups of fish 7 days beforeexposing them to disease, and three groups 4 h or 1day before exposure; three groups (controls) were notstressed. All groups were subjected to the exposureprotocol but only two of three groups for each treat¬ment actually received the pathogen. The third groupserved as controls for non-disease-related mortalities(no fish in any of the controls died during the diseasechallenges). In the second experiment, we removedfour fish from each tank before exposure and col¬lected plasma for cortisol determination, and anteriorkidney lymphocytes for APC assays.
Clinical trials
We examined the effects of stressful handling duringstandard hatchery operations at Warm Springs andRound Butte hatcheries from May through October1986 and 1987. Fish weights increased from about 5 gto 60 g during these trials. Hatchery operators markor tag anadromous fish so that they can determine theproportion of juvenile fish released that survive toadulthood and contribute to the fishery. Fish markingoperations at the two hatcheries were similar, in thatall of the fish in a raceway (about 300 000) werecrowded to one end of the raceway, removed, anaes¬thetized, individually marked by hatchery workers,allowed to recover from the anaesthetic, and returnedto a raceway at 20% of the previous density (i.e.60 000 fish/raceway). At Warm Springs, 60 000 fishwere handled in 4 h, compared with several hundredfish in 45-60 min at Round Butte. Workers at Warm
Springs marked fish by clipping off the left or rightpectoral fin, whereas workers at Round Butte clippedthe adipose fin and inserted a microscopic coded-wiretag into the fish's snout. During four trials, we
sampled 25-30 fish from raceways before stress, and4 h, 1 day, 7 days and for several weeks after stress.
We collected emigrating juvenile fall chinooksalmon from the Columbia River to assess the effectsof stress on disease resistance. About 8-9 months afterfall chinook salmon are spawned, the juveniles arereleased from hatcheries and migrate to the ocean.At McNary Dam, on the Columbia River, theseemigrants are collected and transported by tank-truckor barge to a release site below Bonneville Dam, thelast downstream dam on the Columbia River. Trans¬ported fish avoid the often fatal hazards associatedwith passage through four intervening hydroelectricdams, but the fish are exposed to stresses associatedwith collection and transportation (Maule, Schreck,Bradford & Barton, 1988). To assess the effects of thismanipulation on disease resistance, we transportedtwo groups of fish to the Mark O. Hatfield MarineScience Center (HMSC), Newport, OR, U.S.A. andexposed them to V. anguillarum. In each experiment,about 500 fall chinook salmon (weights were about 9 gin June and 24 g in August 1982) were transferredfrom the holding facilities at McNary Dam to a 760litre tank mounted on a truck for transport to HMSC.The tank had a water recirculation system to aeratewater and water temperature was maintained atambient river-water temperature (16±1°C in Juneand 20 ± 1 °C in August) by adding ice as needed.Each transport run took 8-9 h and upon arrival atHMSC, 25-30 fish were stocked into each of twelve0-61 m circular tanks. Duplicate tanks of fish were
exposed immediately upon arrival, or 1 day or 8 daysafter arrival. These fish were going through, or hadgone through, a physiological transformation (smolti-fication) that would allow them to survive in themarine environment (Hoar, 1976). To control for anundetermined osmoregulatory preference, we exposedsimilar groups of fish to the pathogen in seawater andfreshwater, as described above.
Data analysesAll numerical data were subjected to analyses ofvariance and, where significant between-group differ¬ences were found, we conducted pairwise compari¬sons, using the Duncan's multiple range test at theP<005 level. Although we present APC data in termsof per cent of control, statistical analyses were donewith the raw data. Per cent mortality data from thedisease challenges were analysed using the G-teststatistic based on the chi-square distribution atP<005 (Sokal & Rohlf, 1981).
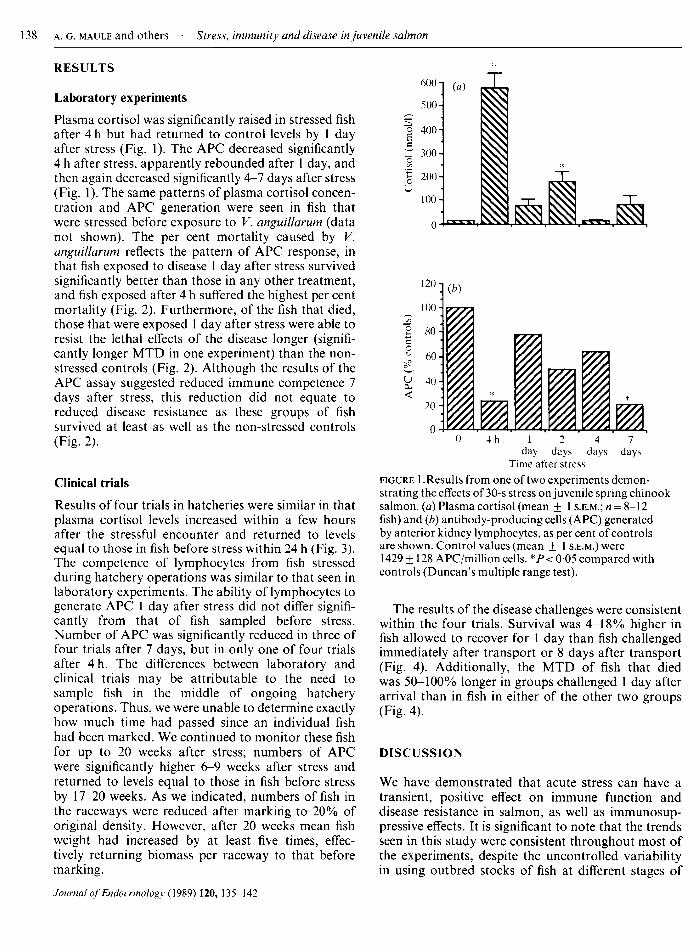
RESULTS
Laboratory experimentsPlasma cortisol was significantly raised in stressed fishafter 4 h but had returned to control levels by 1 dayafter stress (Fig. 1). The APC decreased significantly4 h after stress, apparently rebounded after 1 day, andthen again decreased significantly 4-7 days after stress(Fig. 1). The same patterns of plasma cortisol concen¬tration and APC generation were seen in fish thatwere stressed before exposure to V. anguillarum (datanot shown). The per cent mortality caused by V.anguillarum reflects the pattern of APC response, inthat fish exposed to disease 1 day after stress survivedsignificantly better than those in any other treatment,and fish exposed after 4 h suffered the highest per centmortality (Fig. 2). Furthermore, of the fish that died,those that were exposed 1 day after stress were able toresist the lethal effects of the disease longer (signifi¬cantly longer MTD in one experiment) than the non-
stressed controls (Fig. 2). Although the results of theAPC assay suggested reduced immune competence 7days after stress, this reduction did not equate toreduced disease resistance as these groups of fishsurvived at least as well as the non-stressed controls(Fig. 2).
Clinical trials
Results of four trials in hatcheries were similar in thatplasma cortisol levels increased within a few hoursafter the stressful encounter and returned to levelsequal to those in fish before stress within 24 h (Fig. 3).The competence of lymphocytes from fish stressedduring hatchery operations was similar to that seen inlaboratory experiments. The ability of lymphocytes togenerate APC 1 day after stress did not differ signifi¬cantly from that of fish sampled before stress.Number of APC was significantly reduced in three offour trials after 7 days, but in only one of four trialsafter 4 h. The differences between laboratory andclinical trials may be attributable to the need tosample fish in the middle of ongoing hatcheryoperations. Thus, we were unable to determine exactlyhow much time had passed since an individual fishhad been marked. We continued to monitor these fishfor up to 20 weeks after stress; numbers of APCwere significantly higher 6-9 weeks after stress andreturned to levels equal to those in fish before stressby 17-20 weeks. As we indicated, numbers offish inthe raceways were reduced after marking to 20% oforiginal density. However, after 20 weeks mean fishweight had increased by at least five times, effec¬tively returning biomass per raceway to that beforemarking.
600'
500'
o 400
r 300-
200
100
o BUM
4h 1 2 4 7day days days days
Time after stress
figure 1 .Results from one of two experiments demon¬strating the effects of 30-s stress on juvenile spring chinooksalmon, (a) Plasma cortisol (mean + 1 s.e.m.; = 8-12fish) and (b) antibody-producing cells (APC) generatedby anterior kidney lymphocytes, as per cent of controlsare shown. Control values (mean ± 1 s.e.m.) were1429 ± 128 APC/million cells. *P < 005 compared withcontrols (Duncan's multiple range test).
The results of the disease challenges were consistentwithin the four trials. Survival was 4-18% higher infish allowed to recover for 1 day than fish challengedimmediately after transport or 8 days after transport(Fig. 4). Additionally, the MTD of fish that diedwas 50-100% longer in groups challenged 1 day afterarrival than in fish in either of the other two groups(Fig. 4).
DISCUSSION
We have demonstrated that acute stress can have atransient, positive effect on immune function anddisease resistance in salmon, as well as immunosup-pressive effects. It is significant to note that the trendsseen in this study were consistent throughout most ofthe experiments, despite the uncontrolled variabilityin using outbred stocks of fish at different stages of

100
so
I 60oS 40S?
20
0
(ß)
ILbII¡1111 1 ¡ai Jili(b)
1- 0 4h ld 7d 0 4h ld 7d
April OctoberTime after stress
figure 2. Results of pathogenic challenges ( V. anguillarum)of spring chinook salmon before and at various times after a30-s stress. Plasma cortisol concentrations and APC dataare similar to those in Fig. 1. (a) Mean per cent mortalityand (b) mean time to death (MTD) offish that died induplicate tanks. *P < 005 compared with time zero (G-test(a) or Duncan's multiple range test (b)).
development, in various environmental settings, andafter stress lasting from 30 s to 9 h. The fact thatdisease resistance was not depressed 7 days afterstress indicates that, although the ability of anteriorkidney lymphocytes to generate APC was depressed,other immune mechanisms were not affected or were
strengthened at that time. In fact, the apparent stress-related resistance to this disease 1 day after stress mustbe the result of increased functioning of non-specificimmune mechanisms, because APC response to V.anguillarum O-antigen vaccination peaks in 16-17days in vivo (Maule et al. 1987), whereas death fromexposure to V. anguillarum occurred in 2-6 days (Figs2 and 4). The positive change in immune response andimproved disease resistance seen in salmon 1 day afterstress may be the equivalent of the enhanced immuneresponse seen in mice 15 min after stress (Gisler et al.1971). The additional time (i.e. 1 day versus 15 min)required for the response in fish will be advantageousin elucidating the mechanisms involved.
300
200
100
(¿0
LrzzzJ IZZZL4 h 1 1 6 12 19
day week weeks weeks weeksTime after stress
figure 3. Effects ofstress ofmarking procedures at WarmSpring National Fish Hatchery in 1986. Similar results wereobtained in 1987 and at Round Butte Fish Hatchery in 1986and 1987. (a) Plasma cortisol (mean ± 1 s.e.m.; = 25-32fish) in spring chinook salmon before (time = 0) and atvarious times after marking, (b) Antibody-producing cells(APC) as per cent ofcontrols. Control value (mean + 1S.E.M.) was 460 ± 78 APC/million cells. *P< 005 comparedwith time zero (Duncan's multiple range test).
We previously reported that cortisol implantsreduced disease resistance and ability to generate APCin vivo (Maule et al. 1987) and that the addition ofphysiological concentrations of cortisol to cell cul¬tures caused a dose-dependent reduction in APC invitro (Tripp et al. 1987). In those studies, however,exogenous cortisol was present throughout the 9- to17-day incubation period required to develop theresponse. In mammals, an acute cortisol treatmentcaused redistribution of lymphocytes (primarily regu¬latory cells) out of the circulation and into the bonemarrow (Cupps & Fauci, 1982). This leukocytopeniareached a maximum by 4 h and returned to normalwithin 24 h. We have shown that, during the 3^4months when coho salmon (O. kisutch) were under¬going smoltification, the number of circulating leuko¬cytes and relative numbers of splenocytes decreased as
resting plasma cortisol concentrations increased(Maule et al. 1987). Other data also demonstratedthat, when coho salmon were acutely stressed or given

June
100
S 80
g 60
SS 40
20
FW
isw
1IliCitaïïïAugust
FW SW
mi muW\t IliSiI
0 1 8I I«
18 0 1Time (days)
IIHi
figure 4. Effects of transportation on disease resistance ofemigrating juvenile fallchinook salmon. (Upper graphs) Mean per cent mortality in duplicate groups of 30fish exposed to V. anguillarum at various times after the stress of9-h transpor¬tation. Pathogenic challenges were conducted in freshwater (FW) and seawater(SW) in June and August. (Lower graphs) Mean time to death (MTD) offish thatdied. *P< 005 compared with time zero (G-test (upper) or Duncan's multiplerange test (lower)).
a single feeding of cortisol-treated food, the numbersof cells in the spleen and circulation decreased,whereas cell numbers in the thymus and anteriorkidney increased (A. G. Maule & C. B. Schreck,unpublished data). These alterations were evidentwithin 3 h and persisted for 2-3 days. Thus, in thepresent study, the subpopulations of cells harvestedfrom anterior kidneys of non-stressed control fish mayhave been different from the cells of stressed fish.
Studies with mice have also shown changes in func¬tional populations of cells within lymphoid organsafter stress. Okimura & Nigo (1986) reported thatwhen they expressed their data in terms of anti-TNPAPC per spleen, there was no significant differencebetween stressed mice and controls; however, whenAPC were expressed relative to number of cells in thespleen, stressed mice had significantly more APC thancontrols. This relation suggests that some cells notnecessary for the anti-TNP response had been select¬ively removed from the spleens of stressed mice.Subsequently, Okimura et al. (1986 ) and Okimura,Ogawa, Yamauchi & Sasaki (19866) concluded that
stress-induced increases in corticosteroids and cate¬cholamines were acting to suppress the function of cells, thus suppressing the response of cells thatrequire cell help for activation (T-dependent, TD).At the same time, cells that did not require cellhelp (T-independent, T,) were not affected or were
augmented by the stress. Although there are function¬ally heterogeneous cells in the salmon immune system,the TD-T, paradigm has not been confirmed in sal¬mon. It is known that the hormonal response offish toacute stress is similar to that of mammals in that thereis an immediate increase in circulating catecholaminesfollowed by an increase in corticosteroids (Pickering,1981). Furthermore, Tripp et al. (1987) and Kaattari& Tripp (1987) demonstrated that cortisol sup¬pression of APC in salmon appears to be caused byinhibition of an interleukin-like (interleukin-1) mole¬cule secreted by macrophage-like cells to activateantigen-specific cell precursors. In the present study,it appears that 4 h after stress cortisol had a directimmunosuppressive effect, but that changes in immunefunction and disease resistance between 1 and 7 days

after stress may be the result of other hormonallydriven effects on the immune system. These findingssuggest similar immune-endocrine interactions in themouse and salmon.
ACKNOWLEDGEMENTS
We wish to thank the staff at the Warm SpringNational Fish Hatchery and the Round Butte FishHatchery for their assistance during this study. Agen¬cies co-operating in the Oregon Cooperative FisheryResearch Unit are the U.S. Fish and Wildlife Service,Oregon State University and the Oregon Departmentof Fish and Wildlife. This paper is Oregon AgricultureExperiment Station No. 8503. Funding was providedby the National Océanographie and AtmosphericAdministration Office of Sea Grant, Department ofCommerce, grant number NA85AA-D-SG-095 andthe Bonneville Power Administration, Project 82-16,contract DE-A179-82BP3479. The use of trade namesdoes not imply endorsement of commercial productsby the authors, funding agencies or co-operators.
REFERENCES
Ader, R. (1981). Psychoneuroimmunology: New York: AcademicPress.
Barton, . ., Schreck, C. B., Ewing, R. D., Hemmingsen, A. R. &Patino, R. (1985). Changes in plasma cortisol during stress andsmoltification in coho salmon, Oncorhynchus kisutch. General andComparative Endocrinology 59,468—471.
Barton, . ., Schreck, C. . & Sigismondi, L. . (1986). Multipleacute disturbances evoke cumulative physiological responses injuvenile chinook salmon. Transactions of the American FisheriesSociety 115,246-251.
Cunningham, A. J. & Szenberg, A. (1968). Further improvements inthe plaque technique for detecting single antibody-forming cells.Immunology 14, 599.
Cupps, T. R„ Edgar, L. C, Thomas, C. A. & Fauci, A. S. ( 1984).Multiple mechanisms of cell immunoregulation in man afteradministration of in vìvo corticosteroids. Journal ofImmunology132,170-175.
Cupps, T. R. & Fauci, A. S. (1982). Corticosteroid-mediatedimmunoregulation in man. Immunological Reviews 65,133-155.
Gisler, R. H., Bussard, A. E., Mazie, J. C. & Hess, R. (1971 ).Hormonal regulation of an immune response. I. Induction of an
immune response in vitro with lymphoid cells from mice exposedto acute systemic stress. Cellular Immunology 2,634-645.
Goodwin, J. S. & Atluru, D. (1986). Mechanism of action ofglucocorticoid-induced immunoglobulin production: role oflipoxygenase metabolites of arachidonic acid. Journal ofImmunology 136, 3455-3460.
Gould, R. W., O'Leary, P. J., Garrison, R. L., Rohovec, J. S. &Fryer, J. L. (1978). Spray vaccination: a method for theimmunization offish. Fish Pathology 13,63-68.
Hoar, W. S. (1976). Smolt transformation: evolution, behavior, andphysiology. Journal of the Fisheries Research Board ofCanada 33,1234-1252.
Jacobs, D. M. & Morrison, D. C. (1975). Stimulation ofaT-independent primary anti-hapten response in vitro by TNP-
lipopolysaccharide (TNP-LPS). Journal ofImmunology 144,360-364.
Kaattari, S. L. & Tripp, R. A. (1987). Cellular mechanisms of gluco¬corticoid immunosuppression in salmon. Journal ofFish Biology31(A), 129-132.
Laudenslager, M. L., Ryan, S. M.. Drugan, R. C, Hyson, R. L. &Maier, S. F. (1983). Coping and immunosuppression: inescapablebut not escapable shock suppresses lymphocyte proliferation.Science 221, 568-570.
Maule, A. G, Schreck, C. B., Bradford, C. S. & Barton, B. A.(1988). Physiological effects ofcollecting and transportingemigrating juvenile chinook salmon past dams on the ColumbiaRiver. Transactions of the American Fisheries Society.
Maule, A. G., Schreck, C. B. & Kaattari, S. L. (1987). Changes inthe immune system of coho salmon (Oncorhynchus kisutch)during the parr-to-smolt transformation and after implantationofcortisol. Canadian JournalofFisheries andAquatic Sciences 44,161-166.
McLeay, D. J. (1973). Effects ofcortisol and dexamethasone on thepituitary-interrenal axis and abundance of white blood cell typesin juvenile coho salmon, Oncorhynchus kisutch. General andComparative Endocrinology 21,441—450.
Monjan, A. A. & Collector, M. I. (1977). Stress induced modulationof the immune response. Science 196,307-308.
Munck, ., Guyre, P. M. & Holbrook, N. J. (1984). Physiologicalfunctions of glucocorticoids in stress and their relations topharmacological actions. Endocrine Reviews 5,25—44.
Okimura, T. & Nigo, Y. (1986). Stress and immune responses. I.Suppression of cell function in restrain-stressed mice. JapaneseJournal ofPharmacology 40, 505-511.
Okimura, T., Satomi-Sasaki, Y. & Ohkuma, S. (1986a). Stress andimmune responses. II. Identification of stress-sensitive cells inmurine spleen cells. Japanese Journal ofPharmacology 40,513-525.
Okimura, T., Ogawa, M., Yamauchi, T. & Sasaki, Y. (19866). Stressand immune responses. IV. Adrenal involvement in the alterationof antibody responses in restraint-stressed mice. Japanese JournalofPharmacology 41,237-245.
Pickering, A. D. (Ed.) (1981). Stress infish. London: AcademicPress.
Pickford, G. E., Srivastava, A. K., Slicher, . M. & Pang, P. . T.(1971). The stress response in the abundance of circulating leuco¬cytes in the killifish, Fundulus heteroclitus III. The role of theadrenal cortex and a concluding discussion of the leucocyte-stress syndrome. Journal ofExperimental Zoology 111,109-118.
Redding, J. M., Schreck, C. B., Birks, E. K. & Ewing, R. D. (1984).Cortisol and its effects on plasma thyroid hormones and electro¬lyte concentrations in fresh water during seawater acclimationin yearling coho salmon, Oncorhynchus kisutch. General andComparative Endocrinology 56, 146-155.
Riley, V. (1981). Psychoneuroendocrine influences on immuno-competence and neoplasia. Science 212, 1100-1109.
Rittenberg, . . & Pratt, . (1969). Anti-trinitrophenyl (TNP)plaque assay. Primary response of BALB/c mice to soluble andparticulate immunogen. Proceedings of the Society ofBiologicalMedicine 132, 575-578.
Sokal, R. R. & Rohlf, F. J. (1981). Biometry. San Francisco: W. H.Freeman.
Spangelo, B. L., Hall, N. R. & Goldstein, A. L. (1985). Evidencethat prolactin is an immunomodulatory hormone. In Prolactin.Basic and Clinical Correlates, pp. 343-349. Eds R. M. MacLeod,M. O. Thorner & U. Scapagnini. Fidia Research Series, vol 1.Padova: Liviana Press.
Stein, M., Keller, S. E. & Schleifer, S. J. (1985). Stress and immuno-modulation: the role of depression and neuroendocrine function.Journal ofImmunology 135,827s-833s.

Strange, R. J. & Schreck, C. . (1978). Anesthetic and handlingstress on survival and cortisol concentration in yearling chinooksalmon (Oncorhynchus tshawytscha). Journal of the FisheriesResearch Board of Canada 35,345-349.
Tittle, T. V. & Rittenberg, . . (1978). Expression of IgG memoryresponse in vitro to thymus-independent antigens. CellularImmunology 35, 180-184.
Tripp, R. ., Maule, . G., Schreck, C. . & Kaattari, S. L. (1987).Cortisol mediated suppression of salmonid lymphocyte responsesin vitro. Developmental and Comparative Immunology 11,565-576.
Yu, D. T. Y. & Clements, P. J. (1976). Human lymphocyte sub¬populations: effects of epinephrine. Clinical and ExperimentalImmunology 25,472^179.