strategy changing penalty promotes cooperation in spatial prisoner’s dilemma game
TRANSCRIPT

Chaos, Solitons & Fractals 45 (2012) 395–401
Contents lists available at SciVerse ScienceDirect
Chaos, Solitons & FractalsNonlinear Science, and Nonequilibrium and Complex Phenomena
journal homepage: www.elsevier .com/locate /chaos
Strategy changing penalty promotes cooperation in spatial prisoner’sdilemma game
Qing Jin a, Zhen Wang a,b,c,⇑, Zhen Wang d,e, Yi-Ling Wang f
a School of Physics, Nankai University, Tianjin 300071, Chinab Department of Physics, Hong Kong Baptist University, Kowloon Tong, Hong Kongc Center for Nonlinear Studies and the Beijing-Hong Kong-Singapore Joint Center for Nonlinear and Complex systems (Hong Kong), Hong Kong Baptist University,Kowloon Tong, Hong Kongd School of Innovation Experiment, Dalian University of Technology, Dalian 116024, Chinae School of Software Technology, Dalian University of Technology, Dalian 116024, Chinaf School of Life Science, Shanxi Normal University, LinFen, Shanxi 041000, China
a r t i c l e i n f o a b s t r a c t
Article history:Received 16 December 2011Accepted 13 January 2012Available online 14 February 2012
0960-0779/$ - see front matter � 2012 Elsevier Ltddoi:10.1016/j.chaos.2012.01.004
⇑ Corresponding author at: Department of PhysicUniversity, Kowloon Tong, Hong Kong.
E-mail addresses: [email protected] (Z.edu.cn (Z. Wang).
Many classical studies suggest that punishment is a useful way to promote cooperation inthe well-mixed public goods game, whereas relative evidence in the research of spatialprisoner’s dilemma game is absent. To address this issue, we introduce a mechanism ofstrategy changing penalty, combining memory and penalty during the update process, intospatial prisoner’s dilemma game. We find that increasing penalty rate or memory length isable to promote the evolution of cooperation monotonously. Compared with traditionalversion, recorded penalty could facilitate cooperation better. Moreover, through examiningthe process of evolution, we provide an interpretation for this promotion phenomenon,namely, the effect of promotion can be warranted by an evolution resonance of standarddeviation of fitness coefficient. Finally, we validate our results by studying the impact ofuncertainty within strategy adoptions on the evolution of cooperation. We hope that ourwork may shed light on the understanding of the cooperative behavior in the society.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Cooperative behavior is abundant in the real world, rang-ing from microorganism groups to complex human societies[1,2]. To understand the emergence and persistence of coop-eration, it has attracted great interest in biology, physics,economics, as well as sociology [3,4]. Evolutionary gametheory has proved to be one of the most fruitful approachesto investigate this problem, using evolutionary modelsbased on the so-called social dilemmas [5,6]. Well-knownexamples of these dilemmas include public goods gamefor groups of interaction individuals [8,9,7,10], snowdrift
. All rights reserved.
s, Hong Kong Baptist
Wang), wangz@dlut.
game [11–13] and prisoner’s dilemma game [14–19] forpairwise interactions. Among these evolutionary games,the prisoner’s dilemma game has acquired most prominentachievements in theoretical and experimental studies[20–22]. In a typical prisoner’s dilemma game, two playershave a choice between cooperation and defection. They willreceive the reward R if both players cooperate, and thepunishment P if both choose defection. However, if onechooses defection against a cooperator, it is able to attainthe temptation T while the co-player obtains the sucker’spayoff S. The ranking of four payoffs satisfies T > R > P > S,from which it is clear that selfish players are forced to choosedefection, irrespective of the co-player’s selection. Thus, thesocial dilemma that leads to widespread defection appears.In order to overcome this dilemma, specific mechanismssupporting cooperation are needed (see Ref. [23] for a recentreview).

396 Q. Jin et al. / Chaos, Solitons & Fractals 45 (2012) 395–401
Over the past decades, a number of mechanisms havebeen proposed to explain the emergence of cooperation[24–26], the introduction of spatial structure being one ofthem, which is also refereed as network reciprocity[27,28]. This successful research was firstly suggested byNowak and May in their seminal paper [22]. In the spatialgame, players were situated on the nodes of networks.Each player did not interact with every other, but withits neighbors. The payoff (or fitness) of each player wasevaluated by playing the game with all its neighbors. Theevolution of individual strategy was determined throughadopting the strategy of its neighbor, provided that itsfitness was higher. Interestingly, it proved that networkreciprocity played a so significant role that cooperatorscould survive via forming compact clusters. Stimulatedby this pioneering work, a great many investigations basedon spatial structure have been extended to date [29–32](for a survey see [33]). Most notably, paradigmatic exam-ples include heterogeneous activity of players [17,34,35],reward mechanism [36], influence of noise level [29,37],preferential selection of neighbors [38], effect of expectedpayoffs [39], mobility of players [40–42], differences intime scales [43], to name but a few. While among thesemechanisms, what triggers our research interest most isthe reward mechanism. For instance, a recent work ofSzolnoki and Perc shows that moderate reward may pro-mote cooperation better than high reward [44]. Mean-while, it is prominent that the effect of punishment andreward on promoting the evolution of cooperation isdebatable, and the relative study of punishment mecha-nism mainly focuses on public goods game [45–47], whichmeans it is seldom investigated in prisoner’s dilemmagame. Motivated by these facts, it is constructive to builda simple yet meaningful mechanism that is relative withpunishment, but not completely similar to the traditionalconcept, in the prisoner’s dilemma game, where the playerwill suffer an appropriate penalty if it fails to pass its ownstrategy to its offsprings. And furthermore, the origin ofthis new idea is able to gain reasonable interpretation fromsome realistic phenomena in natural society, which will becharacterized in the next section.
On the other hand, the coevolution of game theory isbecoming a mushrooming avenue to explore the evolutionof cooperation [48–51]. Since it not only reflects the evolv-ing of strategies over time, but also characterizes the adap-tive development of the network topologies or the updaterules (for a further view see Refs. [52,53]). For instance, in[51] authors showed that cooperation could be promotedwithin a large scale when the coevolution of strategyupdating and network topology was taken into account,while the coevolution of teaching ability was identifiedas an beneficial scenario to facilitate the survival of cooper-ation [54]. Inspired by these innovations, an interestingquestion poses itself, which we aim to address in what fol-lows. Namely, if we consider the coevolution scheme aboutpenalty which is based on the above mentioned viewpointand introduces memory length that is strictly related withstrategy changing time, can this accelerate the evolution ofcooperation or not?
In the present work, we study the spatial prisoner’sdilemma game with the introduction of strategy changing
penalty involved with penalty and memory length. Beforethe game, each player holds the same fitness coefficient,which makes the game return to the traditional version.However, with the evolvement of the game, fitness coeffi-cient of individuals also evolve according to the strategychanging situation and memory length. Our main purposeis to study how this mechanism affects the evolution ofcooperation, and if it really facilitates cooperation, whatsupports the promotion phenomenon. By means of sys-tematic Monte Carlo simulations, we show that, as whatwe expected, this mechanism promotes the evolution ofcooperation. Interestingly, we find that this promotionphenomenon can be supported by a recovery effect, whichcan be well interpreted by an evolution resonance of stan-dard deviation of fitness coefficient. Moreover, we examinethe impact of different levels of uncertainty by strategyadoptions. In the remainder of this paper we will first de-scribe the considered evolutionary game, subsequentlypresent the main results, and summarize our conclusionsfinally.
2. Model
We consider an evolutionary prisoner’s dilemma gamewith players located on the sites (x) of a regular L � L squarelattice with periodic boundary conditions. The player at sitex is initially designated as a cooperator (sx = C) or defector(sx = D) with equal probability, and is given a fitness coeffi-cient /x(t) as well. Following the notation suggested in pre-vious literatures [35,37], we use the rescaled payoff matrix:the temptation to defect T = b (the highest payoff receivedby a defector if playing against a cooperator), reward formutual cooperation R = 1, and both the punishment for mu-tual defection P and the sucker’s payoff S (the lowest payoffreceived by a cooperator if playing against a defector)equaling to 0, whereby 1 6 b 6 2 ensures a proper payoffranking. The game is iterated in accordance with the MonteCarlo (MC) simulation procedure comprising the followingelementary steps. First, player x acquires its payoff Px byplaying the game with its four nearest neighbors, and thenevaluates its fitness Px via the following expression
Px ¼ /xðtÞ � Px: ð1Þ
Next, one randomly chosen neighbor, denoted by y, alsoevaluates its fitness Py in the same way. Last, player xadopts the strategy sy from the selected player y in accor-dance with the probability
Wðsy ! sxÞ ¼1
1þ exp½ðPx �PyÞ=K� ; ð2Þ
where K denotes the amplitude of noise or its inverse (1/K),the so-called intensity of selection [21]. During one fullMonte Carlo step (MCS) each player has a chance once onaverage to adopt the strategy from a randomly selectedneighbor as described above.
Of particular renown, the fitness coefficient is alsoallowed to evolve according to the following protocol. Be-fore the game player x possesses the same fitness coeffi-cient /x(t) = 1.0 to avoid preferential influence. However,if player x adopts the strategy of its neighbor y, namely,

Fig. 1. Frequency of cooperator qC in dependence on the parameter b fordifferent values of penalty rate a. Note that a = 0 recovers the traditionalversion, while the increment of a will introduce the impact of penalty intothe game. Depicted results were obtained for M = 5 and K = 0.1.
Q. Jin et al. / Chaos, Solitons & Fractals 45 (2012) 395–401 397
it fails to pass its own strategy to the offspring, it will suffera certain penalty. The similar phenomenon is ubiquitousamong the competition of biological species and other so-cial behaviors [57,58], for example, two species competefor the territory, the defected one (which corresponds tothe case of individual adopting other strategy in the game)and its offspring lose part of their initial territory, whichwould affect their activity and induce a new evaluationwith respect to the adjusted environment [59]. While inour study we mainly focus on how the strategy changingaffects the fitness evaluation. We define when the strategychanging occurs, the fitness coefficient at site x willdecrease in the following way /x(t + 1) = (1 � a) � /x(t),where a (a� 1.0) is the penalty rate, and otherwise thefitness coefficient keeps constant. At the same time, we as-sume that the step when player x incurs a penalty is re-garded as the first step. If player x keeps its strategychangeless in the subsequent M steps (called memorylength), namely, it remains indefectible during the succe-dent competitions or battles, its fitness coefficient returns1.0. However, once player x still adopts the strategy of itsneighbor in one of the subsequent M steps, that step willbe regarded as the first step over again. The above processrepeats till its fitness coefficient gets back to 1.0. This setupseems reasonable and is easily justifiable with realisticexamples. For example, it is a fact that every social or nat-ural system holds its own rules. When anyone violatesthem, he will undergo the chastisement, and even his free-dom or property is partly restricted. However, if his subse-quent performance is well in the special period, he canregain the freedom or property.
Results of Monte Carlo simulations presented belowwere obtained on populations comprising 100 � 100 to400 � 400 individuals, whereby the fraction of cooperatorsqC was determined within last 104 full steps of overall2 � 105 MCS. In order to overcome the influence of largeM value, longer simulation time was used. Moreover, sincethe penalty mechanism may introduce heterogeneousinfluence of fitness coefficient, final results were averagedover 20–40 independent runs for each set of parametervalues in order to assure suitable accuracy.
Fig. 2. Frequency of cooperator qC in dependence on the parameter b fordifferent values of memory length M. Note that with the increment ofmemory length M, cooperation can be greatly facilitated, but theenhancement of cooperation will reach a limit if memory length issufficient large. Depicted results were obtained for a = 0.1 and K = 0.1.
3. Simulation results and discussion
We start by examining the impact of penalty rate a onthe evolution of cooperation while keeping the memorylength M constant. Fig. 1 shows how qC varies in depen-dence on the temptation of defect b for different valuesof a. Evidently, a = 0 (irrespective of the value of M) recov-ers the traditional version of spatial prisoner’s dilemmagame, where cooperators die out when the value of b issmall. However, as a increases, the evolution of coopera-tion will be promoted more effectively. It can be clearly ob-served that for a = 0.02 cooperation can be facilitated to anear-complete dominant strategy when the value of b issmall. With the continuous increment of a, defectors canonly exist when the temptation to defect is sufficient large.These results suggest that when the strategy changingpenalty is taken into account, the emergence and mainte-nance of cooperation could be enhanced, and the increase
of penalty rate a could directly result in a better evolutionof cooperation, since the introduction of strategy changingpenalty may result in a heterogeneous state during theprocess of evolution.
In what follows, we explore the impact of memorylength M on the evolution of cooperation. Results pre-sented in Fig. 2 clearly shows how qC varies in dependenceon b for different values of M. What firstly attracts our curi-osity is the case of M = 0 that corresponds to the situationwhere only penalty but no memory is contained in thegame, since strategy changing penalty in present step willhave no influence on the next one. Then compared with theresults of traditional version (see black squares in Fig. 1), itis evident that better facilitation of cooperation could bewarranted, which implies that the performance of spatialreciprocity could be greatly improved [33]. As the valueof memory length M increases, the evolution of
398 Q. Jin et al. / Chaos, Solitons & Fractals 45 (2012) 395–401
cooperation fares better, namely, the survivability of coop-erators will monotonously enhance with increasing M. Inparticular, when the value of M is sufficiently large, thepromotion effect becomes saturated, that is, the fractionof cooperators converges to a deterministic value for en-ough large value of M. It is worth pointing out that for largevalue of M system needs longer time to arrive at the steadystate. Moreover, it is also notable that compared with thecase of merely punishment (M = 0), cooperation could bebetter promoted within the framework involved with thejoint influence of penalty and memory.
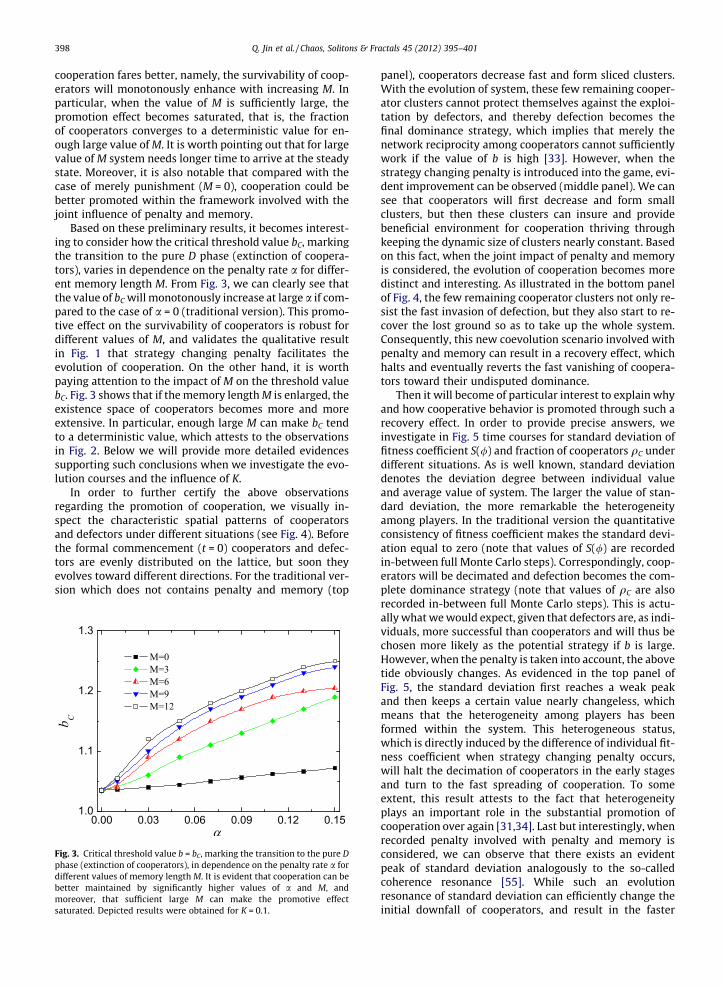
Based on these preliminary results, it becomes interest-ing to consider how the critical threshold value bC, markingthe transition to the pure D phase (extinction of coopera-tors), varies in dependence on the penalty rate a for differ-ent memory length M. From Fig. 3, we can clearly see thatthe value of bC will monotonously increase at large a if com-pared to the case of a = 0 (traditional version). This promo-tive effect on the survivability of cooperators is robust fordifferent values of M, and validates the qualitative resultin Fig. 1 that strategy changing penalty facilitates theevolution of cooperation. On the other hand, it is worthpaying attention to the impact of M on the threshold valuebC. Fig. 3 shows that if the memory length M is enlarged, theexistence space of cooperators becomes more and moreextensive. In particular, enough large M can make bC tendto a deterministic value, which attests to the observationsin Fig. 2. Below we will provide more detailed evidencessupporting such conclusions when we investigate the evo-lution courses and the influence of K.
In order to further certify the above observationsregarding the promotion of cooperation, we visually in-spect the characteristic spatial patterns of cooperatorsand defectors under different situations (see Fig. 4). Beforethe formal commencement (t = 0) cooperators and defec-tors are evenly distributed on the lattice, but soon theyevolves toward different directions. For the traditional ver-sion which does not contains penalty and memory (top
Fig. 3. Critical threshold value b = bC, marking the transition to the pure Dphase (extinction of cooperators), in dependence on the penalty rate a fordifferent values of memory length M. It is evident that cooperation can bebetter maintained by significantly higher values of a and M, andmoreover, that sufficient large M can make the promotive effectsaturated. Depicted results were obtained for K = 0.1.
panel), cooperators decrease fast and form sliced clusters.With the evolution of system, these few remaining cooper-ator clusters cannot protect themselves against the exploi-tation by defectors, and thereby defection becomes thefinal dominance strategy, which implies that merely thenetwork reciprocity among cooperators cannot sufficientlywork if the value of b is high [33]. However, when thestrategy changing penalty is introduced into the game, evi-dent improvement can be observed (middle panel). We cansee that cooperators will first decrease and form smallclusters, but then these clusters can insure and providebeneficial environment for cooperation thriving throughkeeping the dynamic size of clusters nearly constant. Basedon this fact, when the joint impact of penalty and memoryis considered, the evolution of cooperation becomes moredistinct and interesting. As illustrated in the bottom panelof Fig. 4, the few remaining cooperator clusters not only re-sist the fast invasion of defection, but they also start to re-cover the lost ground so as to take up the whole system.Consequently, this new coevolution scenario involved withpenalty and memory can result in a recovery effect, whichhalts and eventually reverts the fast vanishing of coopera-tors toward their undisputed dominance.
Then it will become of particular interest to explain whyand how cooperative behavior is promoted through such arecovery effect. In order to provide precise answers, weinvestigate in Fig. 5 time courses for standard deviation offitness coefficient S(/) and fraction of cooperators qC underdifferent situations. As is well known, standard deviationdenotes the deviation degree between individual valueand average value of system. The larger the value of stan-dard deviation, the more remarkable the heterogeneityamong players. In the traditional version the quantitativeconsistency of fitness coefficient makes the standard devi-ation equal to zero (note that values of S(/) are recordedin-between full Monte Carlo steps). Correspondingly, coop-erators will be decimated and defection becomes the com-plete dominance strategy (note that values of qC are alsorecorded in-between full Monte Carlo steps). This is actu-ally what we would expect, given that defectors are, as indi-viduals, more successful than cooperators and will thus bechosen more likely as the potential strategy if b is large.However, when the penalty is taken into account, the abovetide obviously changes. As evidenced in the top panel ofFig. 5, the standard deviation first reaches a weak peakand then keeps a certain value nearly changeless, whichmeans that the heterogeneity among players has beenformed within the system. This heterogeneous status,which is directly induced by the difference of individual fit-ness coefficient when strategy changing penalty occurs,will halt the decimation of cooperators in the early stagesand turn to the fast spreading of cooperation. To someextent, this result attests to the fact that heterogeneityplays an important role in the substantial promotion ofcooperation over again [31,34]. Last but interestingly, whenrecorded penalty involved with penalty and memory isconsidered, we can observe that there exists an evidentpeak of standard deviation analogously to the so-calledcoherence resonance [55]. While such an evolutionresonance of standard deviation can efficiently change theinitial downfall of cooperators, and result in the faster
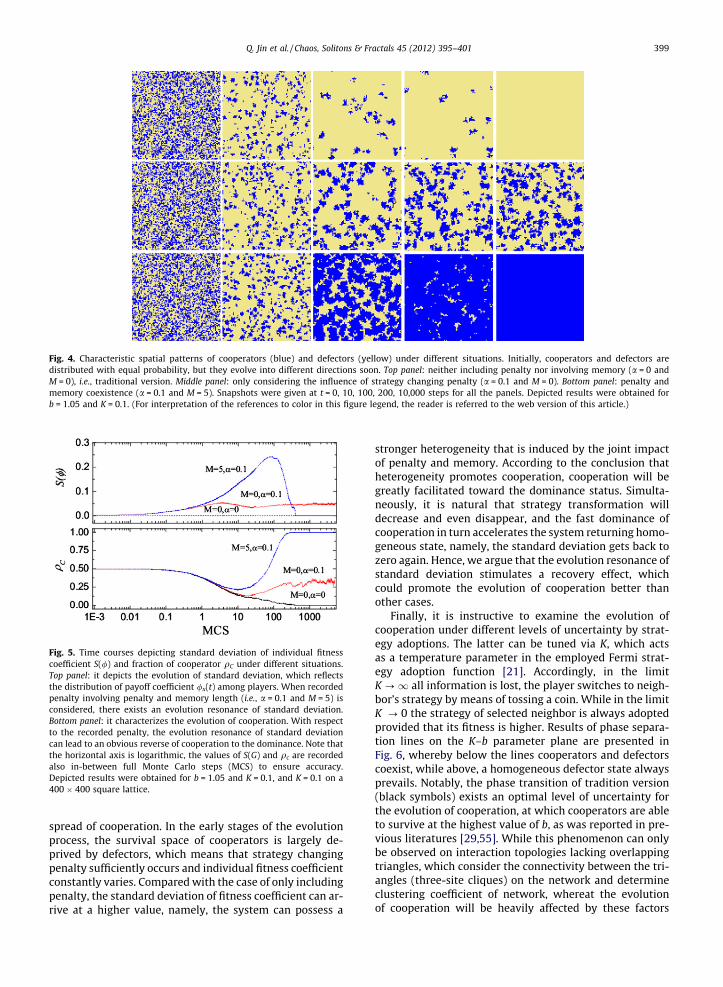
Fig. 4. Characteristic spatial patterns of cooperators (blue) and defectors (yellow) under different situations. Initially, cooperators and defectors aredistributed with equal probability, but they evolve into different directions soon. Top panel: neither including penalty nor involving memory (a = 0 andM = 0), i.e., traditional version. Middle panel: only considering the influence of strategy changing penalty (a = 0.1 and M = 0). Bottom panel: penalty andmemory coexistence (a = 0.1 and M = 5). Snapshots were given at t = 0, 10, 100, 200, 10,000 steps for all the panels. Depicted results were obtained forb = 1.05 and K = 0.1. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
Fig. 5. Time courses depicting standard deviation of individual fitnesscoefficient S(/) and fraction of cooperator qC under different situations.Top panel: it depicts the evolution of standard deviation, which reflectsthe distribution of payoff coefficient /x(t) among players. When recordedpenalty involving penalty and memory length (i.e., a = 0.1 and M = 5) isconsidered, there exists an evolution resonance of standard deviation.Bottom panel: it characterizes the evolution of cooperation. With respectto the recorded penalty, the evolution resonance of standard deviationcan lead to an obvious reverse of cooperation to the dominance. Note thatthe horizontal axis is logarithmic, the values of S(G) and qc are recordedalso in-between full Monte Carlo steps (MCS) to ensure accuracy.Depicted results were obtained for b = 1.05 and K = 0.1, and K = 0.1 on a400 � 400 square lattice.
Q. Jin et al. / Chaos, Solitons & Fractals 45 (2012) 395–401 399
spread of cooperation. In the early stages of the evolutionprocess, the survival space of cooperators is largely de-prived by defectors, which means that strategy changingpenalty sufficiently occurs and individual fitness coefficientconstantly varies. Compared with the case of only includingpenalty, the standard deviation of fitness coefficient can ar-rive at a higher value, namely, the system can possess a
stronger heterogeneity that is induced by the joint impactof penalty and memory. According to the conclusion thatheterogeneity promotes cooperation, cooperation will begreatly facilitated toward the dominance status. Simulta-neously, it is natural that strategy transformation willdecrease and even disappear, and the fast dominance ofcooperation in turn accelerates the system returning homo-geneous state, namely, the standard deviation gets back tozero again. Hence, we argue that the evolution resonance ofstandard deviation stimulates a recovery effect, whichcould promote the evolution of cooperation better thanother cases.
Finally, it is instructive to examine the evolution ofcooperation under different levels of uncertainty by strat-egy adoptions. The latter can be tuned via K, which actsas a temperature parameter in the employed Fermi strat-egy adoption function [21]. Accordingly, in the limitK ?1 all information is lost, the player switches to neigh-bor’s strategy by means of tossing a coin. While in the limitK ? 0 the strategy of selected neighbor is always adoptedprovided that its fitness is higher. Results of phase separa-tion lines on the K–b parameter plane are presented inFig. 6, whereby below the lines cooperators and defectorscoexist, while above, a homogeneous defector state alwaysprevails. Notably, the phase transition of tradition version(black symbols) exists an optimal level of uncertainty forthe evolution of cooperation, at which cooperators are ableto survive at the highest value of b, as was reported in pre-vious literatures [29,55]. While this phenomenon can onlybe observed on interaction topologies lacking overlappingtriangles, which consider the connectivity between the tri-angles (three-site cliques) on the network and determineclustering coefficient of network, whereat the evolutionof cooperation will be heavily affected by these factors
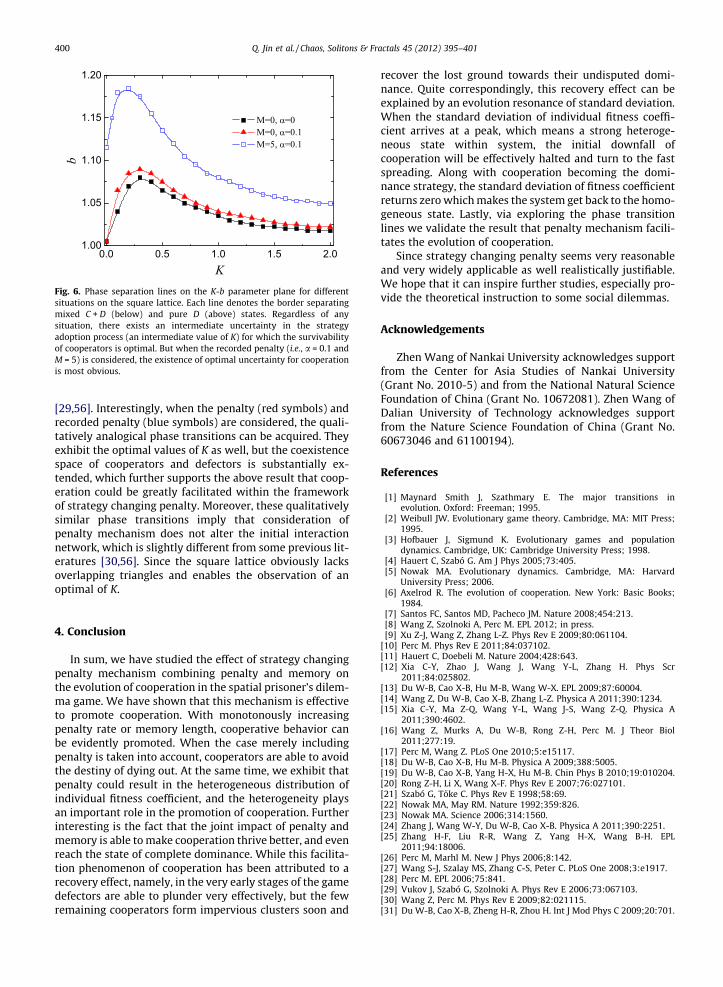
Fig. 6. Phase separation lines on the K-b parameter plane for differentsituations on the square lattice. Each line denotes the border separatingmixed C + D (below) and pure D (above) states. Regardless of anysituation, there exists an intermediate uncertainty in the strategyadoption process (an intermediate value of K) for which the survivabilityof cooperators is optimal. But when the recorded penalty (i.e., a = 0.1 andM = 5) is considered, the existence of optimal uncertainty for cooperationis most obvious.
400 Q. Jin et al. / Chaos, Solitons & Fractals 45 (2012) 395–401
[29,56]. Interestingly, when the penalty (red symbols) andrecorded penalty (blue symbols) are considered, the quali-tatively analogical phase transitions can be acquired. Theyexhibit the optimal values of K as well, but the coexistencespace of cooperators and defectors is substantially ex-tended, which further supports the above result that coop-eration could be greatly facilitated within the frameworkof strategy changing penalty. Moreover, these qualitativelysimilar phase transitions imply that consideration ofpenalty mechanism does not alter the initial interactionnetwork, which is slightly different from some previous lit-eratures [30,56]. Since the square lattice obviously lacksoverlapping triangles and enables the observation of anoptimal of K.
4. Conclusion
In sum, we have studied the effect of strategy changingpenalty mechanism combining penalty and memory onthe evolution of cooperation in the spatial prisoner’s dilem-ma game. We have shown that this mechanism is effectiveto promote cooperation. With monotonously increasingpenalty rate or memory length, cooperative behavior canbe evidently promoted. When the case merely includingpenalty is taken into account, cooperators are able to avoidthe destiny of dying out. At the same time, we exhibit thatpenalty could result in the heterogeneous distribution ofindividual fitness coefficient, and the heterogeneity playsan important role in the promotion of cooperation. Furtherinteresting is the fact that the joint impact of penalty andmemory is able to make cooperation thrive better, and evenreach the state of complete dominance. While this facilita-tion phenomenon of cooperation has been attributed to arecovery effect, namely, in the very early stages of the gamedefectors are able to plunder very effectively, but the fewremaining cooperators form impervious clusters soon and
recover the lost ground towards their undisputed domi-nance. Quite correspondingly, this recovery effect can beexplained by an evolution resonance of standard deviation.When the standard deviation of individual fitness coeffi-cient arrives at a peak, which means a strong heteroge-neous state within system, the initial downfall ofcooperation will be effectively halted and turn to the fastspreading. Along with cooperation becoming the domi-nance strategy, the standard deviation of fitness coefficientreturns zero which makes the system get back to the homo-geneous state. Lastly, via exploring the phase transitionlines we validate the result that penalty mechanism facili-tates the evolution of cooperation.
Since strategy changing penalty seems very reasonableand very widely applicable as well realistically justifiable.We hope that it can inspire further studies, especially pro-vide the theoretical instruction to some social dilemmas.
Acknowledgements
Zhen Wang of Nankai University acknowledges supportfrom the Center for Asia Studies of Nankai University(Grant No. 2010-5) and from the National Natural ScienceFoundation of China (Grant No. 10672081). Zhen Wang ofDalian University of Technology acknowledges supportfrom the Nature Science Foundation of China (Grant No.60673046 and 61100194).
References
[1] Maynard Smith J, Szathmary E. The major transitions inevolution. Oxford: Freeman; 1995.
[2] Weibull JW. Evolutionary game theory. Cambridge, MA: MIT Press;1995.
[3] Hofbauer J, Sigmund K. Evolutionary games and populationdynamics. Cambridge, UK: Cambridge University Press; 1998.
[4] Hauert C, Szabó G. Am J Phys 2005;73:405.[5] Nowak MA. Evolutionary dynamics. Cambridge, MA: Harvard
University Press; 2006.[6] Axelrod R. The evolution of cooperation. New York: Basic Books;
1984.[7] Santos FC, Santos MD, Pacheco JM. Nature 2008;454:213.[8] Wang Z, Szolnoki A, Perc M. EPL 2012; in press.[9] Xu Z-J, Wang Z, Zhang L-Z. Phys Rev E 2009;80:061104.
[10] Perc M. Phys Rev E 2011;84:037102.[11] Hauert C, Doebeli M. Nature 2004;428:643.[12] Xia C-Y, Zhao J, Wang J, Wang Y-L, Zhang H. Phys Scr
2011;84:025802.[13] Du W-B, Cao X-B, Hu M-B, Wang W-X. EPL 2009;87:60004.[14] Wang Z, Du W-B, Cao X-B, Zhang L-Z. Physica A 2011;390:1234.[15] Xia C-Y, Ma Z-Q, Wang Y-L, Wang J-S, Wang Z-Q. Physica A
2011;390:4602.[16] Wang Z, Murks A, Du W-B, Rong Z-H, Perc M. J Theor Biol
2011;277:19.[17] Perc M, Wang Z. PLoS One 2010;5:e15117.[18] Du W-B, Cao X-B, Hu M-B. Physica A 2009;388:5005.[19] Du W-B, Cao X-B, Yang H-X, Hu M-B. Chin Phys B 2010;19:010204.[20] Rong Z-H, Li X, Wang X-F. Phys Rev E 2007;76:027101.[21] Szabó G, T}oke C. Phys Rev E 1998;58:69.[22] Nowak MA, May RM. Nature 1992;359:826.[23] Nowak MA. Science 2006;314:1560.[24] Zhang J, Wang W-Y, Du W-B, Cao X-B. Physica A 2011;390:2251.[25] Zhang H-F, Liu R-R, Wang Z, Yang H-X, Wang B-H. EPL
2011;94:18006.[26] Perc M, Marhl M. New J Phys 2006;8:142.[27] Wang S-J, Szalay MS, Zhang C-S, Peter C. PLoS One 2008;3:e1917.[28] Perc M. EPL 2006;75:841.[29] Vukov J, Szabó G, Szolnoki A. Phys Rev E 2006;73:067103.[30] Wang Z, Perc M. Phys Rev E 2009;82:021115.[31] Du W-B, Cao X-B, Zheng H-R, Zhou H. Int J Mod Phys C 2009;20:701.

Q. Jin et al. / Chaos, Solitons & Fractals 45 (2012) 395–401 401
[32] Vainstein MH, Arenzon JJ. Phys Rev E 2001;64:051905.[33] Szabó G, Fáth G. Phys Rep 2007;446:97.[34] Szolnoki A, Szabó G. EPL 2007;77:30004.[35] Szabó G, Szolnoki A. Phys Rev E 2009;79:016106.[36] Jiménez R, Lugo H, Cuesta JA, Sánchez A. J Theor Biol 2008;250:475.[37] Szolnoki A, Vukov J, Szabó G. Phys Rev E 2009;80:056112.[38] Wu Z-X, Xu X-J, Huang Z-G, Wang S-J, Wang Y-H. Phys Rev E
2006;74:021107.[39] Du W-B, Cao X-B, Hu M-B, Yang H-X, Zhou H. Physica A
2009;388:2215.[40] Helbing D, Yu W-J. Proc Natl Acad Sci USA 2009;106:3680.[41] Vainstein MH, Silva ATC, Arenzon JJ. J Theor Biol 2007;244:722.[42] Sicardi EA, Fort H, Vainsteina MH, Arenzon JJ. J Theor Biol
2009;256:240.[43] Rong Z-H, Wu Z-X, Wang W-X. Phys Rev E 2010;82:026101.[44] Szolnoki A, Perc M. EPL 2010;92:38003.[45] Helbing D, Szolnoki A, Perc M, Szabó G. New J Phys 2010;12:083005.
[46] Sigmund K, De Silva H, Traulsen A, Hauert C. Nature 2010;466:861.[47] Szolnoki A, Szabó G, Perc M. Phys Rev E, in press.[48] Szolnoki A, Perc M. New J Phys 2009;11:093033.[49] Poncela J, Goómez-Gardeñes J, Floría LM, Sánchez A, Moreno Y. PLoS
One 2008;3:e2449.[50] Ebel H, Bornholdt S. Phys Rev E 2002;66:056118.[51] Cardillo A, Gómez-Gardeñes J, Vilone D, Sánchez A. New J Phys
2010;12:103034.[52] Perc M, Szolnoki A. BioSystems 2010;99:109.[53] Gross T, Blasius B. JR Soc Inter 2008;5:259.[54] Szolnoki A, Perc M. New J Phys 2008:10 043036.[55] Perc M. New J Phys 2006;8:22.[56] Szabó G, Vukov J, Szolnoki A. Phys Rev E 2005;72:047107.[57] Krakauer AH. Nature 2005;434:69.[58] Rodrigues A, Koeppl H, Ohtsuki H, Satake A. J Theor Biol 2009;258:
127.[59] Cant MA, English S. Ecol Lett 2006;17:560.