steroidogenesis and apoptosis in the mammalian ovary
TRANSCRIPT

Steroids 68 (2003) 861–867
Steroidogenesis and apoptosis in the mammalian ovary
Abraham Amsterdam∗, Iris Keren-Tal, Dorit Aharoni, Ada Dantes,Abigail Land-Bracha, Eli Rimon, Ravid Sasson, Liron Hirsh
Department of Molecular Cell Biology, Weizmann Institute of Science, Rehovot 76100, Israel
Abstract
Ovarian cell death is an essential process for the homeostasis of ovarian function in human and other mammalian species. It ensuresthe selection of the dominant follicle and the demise of excess follicles. In turn, this process minimizes the possibility of multiple embryodevelopment during pregnancy and assures the development of few, but healthy embryos. Degeneration of the old corpora lutea in eachestrous/menstrual cycle by programmed cell death is essential to maintain the normal cyclicity of ovarian steroidogenesis. Although thereare multiple pathways that can determine cell death or survival, crosstalk among endocrine, paracrine and autocrine factors, as well asamong protooncogenes, tumor suppressor genes, survival genes and death genes, plays an important role in determining the fate of ovariansomatic and germ cells. The establishment of immortalized rat and human steroidogenic granulosa cell lines and the investigation of purepopulations of primary granulosa cells allows systematic studies of the mechanisms that control steroidogenesis and apoptosis in granulosacells. We have discovered that during initial stages of granulosa cell apoptosis progesterone production does not decrease. In contrast, wefound that it is elevated up to 24 h following the onset of the apoptotic stimuli exerted by starvation, cAMP, p53 or TNF-� stimulation, beforetotal cell collapse. These observations raise the possibility for an alternative unique apoptotic pathway, one not involving mitochondrialCyt C release associated with the destruction of mitochondrial structure and steroidogenic function. Using mRNA from apoptotic cellsand affymetrix DNA microarray technology we discovered that granzyme B, a protease that normally resides in T cytotoxic lymphocytesand natural killer cells of the immune system is expressed and activated in granulosa cells. Thus, the apoptotic signals could bypassmitochondrial signals for apoptosis, which can preserve their steroidogenic activity until complete cell destruction. This unique apoptoticpathway assures cyclicity of estradiol and progesterone release in the estrous/menstruous cycle even during the initial stages of apoptosis.© 2003 Elsevier Inc. All rights reserved.
Keywords: Ovary; Granulosa cells; Granztme B
1. Introduction
Ovarian cell death is a crucial event in maintaining ovar-ian homeostasis in mammals. It ensures that in every es-trus/menstrous cycle only one or very few follicle-enclosedoocytes will reach the stage of a Graafian follicle and willovulate. This is essential in the prevention of multiple em-bryos during pregnancy. The rest of the follicles are gradu-ally eliminated during the fertility period of the female. Theapoptotic process of the old corpora lutea is essential formaintaining the cyclicity and the release of progesteroneduring the estrous/menstrous cycle (reviewed in[1–5]).There are several factors that may control apoptosis ofgranulosa cells. In each stage of the cycle about 50% of thelarge preantral and antral follicles will be in the process ofapoptotic death[6]. In most antral follicles apoptosis willinitiate at inner layers of the membrana granulosa, border-
∗ Corresponding author. Tel.:+972-8-9343713; fax:+972-8-9344125.E-mail address: [email protected] (A. Amsterdam).
ing the follicle antrum (Fig. 1) Apoptosis is protected bythe basement membrane which can sequester basic fibrob-lastic growth factor (bFGF) where both laminin the maincomponent of the basement membrane, and bFGF serve assurvival factors[7,8]. Interestingly, laminin protects againstapoptosis without affecting steroidogenesis while bFGF en-hances formation of progesterone in preovulatory follicularcells [7].
Induction of apoptosis in vivo by hyposectomy of fe-male rats surprisingly elevates progesterone production inthe ovary[9]. This may suggest that the initial steps of apop-tosis enhance rather than block progesterone production. Asimilar phenomenon is found upon induction of apoptosisin highly luteinized granulosa cells (Fig. 2) and raises thequestion whether steroidogenesis and apoptosis can exist inthe same cells[10–12]. If the answer is positive, then thenext question is: how can mitochondrial integrity which isthe prime target of apoptotic signals in numerous cell typesbe preserved during the initial steps of apoptosis in ovariansteroidogenic cells?
0039-128X/$ – see front matter © 2003 Elsevier Inc. All rights reserved.doi:10.1016/j.steroids.2003.09.003

862 A. Amsterdam et al. / Steroids 68 (2003) 861–867
Fig. 1. Antral follicle of rat ovary. The oocyte in the center is a section at the level of the germinal vesicle and the nucleolus. Although the oocyteseems intact with no sign of apoptosis, high incidence of apoptosis appears in the inner layers of granulosa cells bordering the antrum (white circles).Hematoxylin–eosine staining 1000×.
1.1. The sensory world of the granulosa cell that controlssteroidogenesis and apoptosis
Increasing evidence suggests that there are multiple apop-totic stimuli in the ovarian follicle as well as in culturedgranulosa cells[3]. Some of the stimuli negate steroidoge-nesis such as, Tumor Necrosis Factor-� (TNF-�) [13], andsome of them enhance steroidogenesis such as, high levelsof intracellular cAMP[10]. Stimuli for apoptosis or sur-vival can be endocrine, paracrine or autocrine[3]. The mainsurvival factors are gonadotropins, epidermal growth fac-tor (EGF), insulin growth factor (IGF), fibroblastic growthfactor (FGF), prolactin, laminin, leptin, glucocorticoidsand estradiol (Fig. 3), while the main apoptotic signals areTNF-�, high levels of cAMP and gonadotropin releasinghormone (GnRH). Eventually, some apoptotic signals cansynergise with each other like high levels of cAMP andactivation of p53[3,14] and some survival factors such assteroid hormones and growth factors can negate apoptoticsignals evoked by p53 and TNF-� [3,13]. We conclude thatcrosstalk among the different signals determines the fate ofthe ovarian follicle. Because there are multiple paracrineand autocrine signals, it is still not completely understoodwhich are the critical factors that discriminate between thefollicles destined for elimination by apoptosis (the majorpopulation and the follicles) and between one or few ofthe follicles that will continue to develop to reach the finalstage of a Graafian preovulatory follicle.
Progesterone formation and release during the estrous/menstrual cycle may play an important role in the fertiliza-tion of the oocyte during ovulation. It has been suggestedthat early progesterone production in the preovulatory folli-cle impairs the quality of the mature egg during fertilization[15–17]. In contrast, proper timing of progesterone produc-tion and the duration of its secretion seems to be critical formaintaining functional granulosa-lutein cells, subsequent tothe LH surge and maintenance of the corpus luteum duringearly pregnancy. Progesterone interaction with its cytoso-lic receptor may play a part in the survival activity of thegranulosa-lutein cells[18].
1.2. Mechanism of action: relationship betweensteroidogenesis and apoptosis
Early studies of hypophysectomed female rats demon-strated a dramatic increase in progesterone formation andrelease from the ovarian follicle, with a progressive in-crease in atresia (apoptotic cells) which reached 20–30%of the total granulosa cell population within 48 h[9]. In-terestingly there was a drop in the formation of follicularandrostinedion, testosterone and estradiol[9] where the en-zymes catalyzing the formation of the latter hormones arelocated at the extra mitochondrial part of the cytoplasm.When immortalized granulosa cells were stimulated to un-dergo apoptosis by activation of p53 and stimulation with50�M of forskolin (FK), there was a dramatic increase in
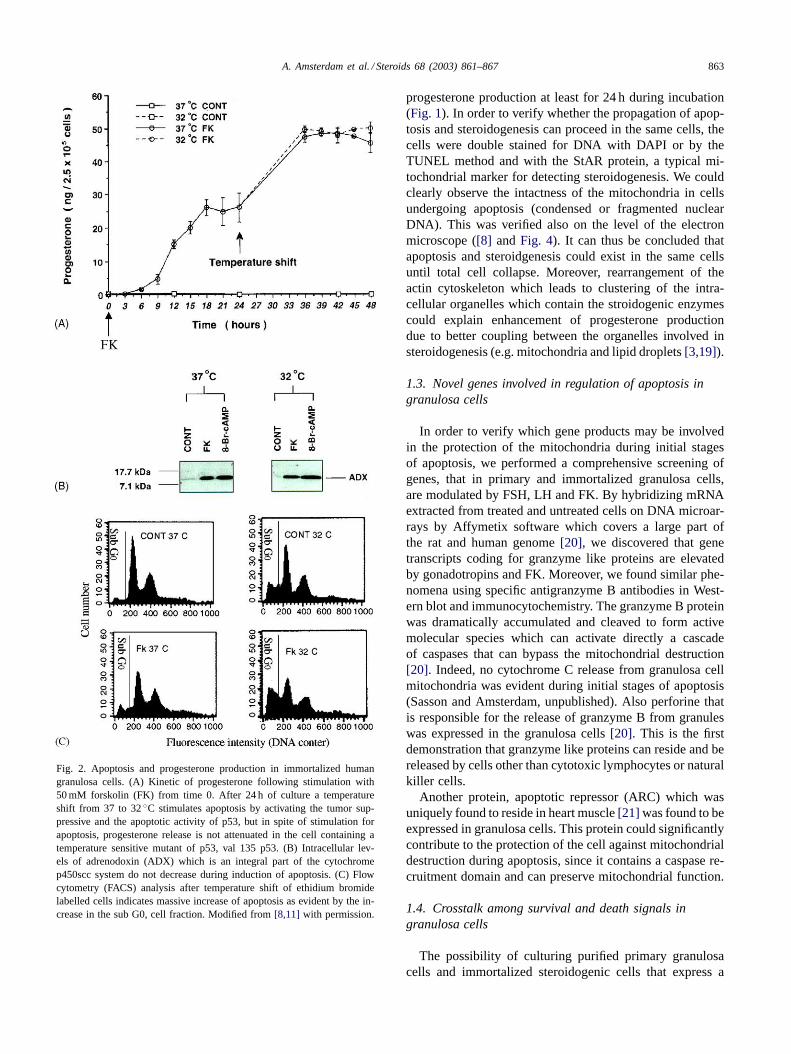
A. Amsterdam et al. / Steroids 68 (2003) 861–867 863
Fig. 2. Apoptosis and progesterone production in immortalized humangranulosa cells. (A) Kinetic of progesterone following stimulation with50 mM forskolin (FK) from time 0. After 24 h of culture a temperatureshift from 37 to 32◦C stimulates apoptosis by activating the tumor sup-pressive and the apoptotic activity of p53, but in spite of stimulation forapoptosis, progesterone release is not attenuated in the cell containing atemperature sensitive mutant of p53, val 135 p53. (B) Intracellular lev-els of adrenodoxin (ADX) which is an integral part of the cytochromep450scc system do not decrease during induction of apoptosis. (C) Flowcytometry (FACS) analysis after temperature shift of ethidium bromidelabelled cells indicates massive increase of apoptosis as evident by the in-crease in the sub G0, cell fraction. Modified from[8,11] with permission.
progesterone production at least for 24 h during incubation(Fig. 1). In order to verify whether the propagation of apop-tosis and steroidogenesis can proceed in the same cells, thecells were double stained for DNA with DAPI or by theTUNEL method and with the StAR protein, a typical mi-tochondrial marker for detecting steroidogenesis. We couldclearly observe the intactness of the mitochondria in cellsundergoing apoptosis (condensed or fragmented nuclearDNA). This was verified also on the level of the electronmicroscope ([8] andFig. 4). It can thus be concluded thatapoptosis and steroidgenesis could exist in the same cellsuntil total cell collapse. Moreover, rearrangement of theactin cytoskeleton which leads to clustering of the intra-cellular organelles which contain the stroidogenic enzymescould explain enhancement of progesterone productiondue to better coupling between the organelles involved insteroidogenesis (e.g. mitochondria and lipid droplets[3,19]).
1.3. Novel genes involved in regulation of apoptosis ingranulosa cells
In order to verify which gene products may be involvedin the protection of the mitochondria during initial stagesof apoptosis, we performed a comprehensive screening ofgenes, that in primary and immortalized granulosa cells,are modulated by FSH, LH and FK. By hybridizing mRNAextracted from treated and untreated cells on DNA microar-rays by Affymetix software which covers a large part ofthe rat and human genome[20], we discovered that genetranscripts coding for granzyme like proteins are elevatedby gonadotropins and FK. Moreover, we found similar phe-nomena using specific antigranzyme B antibodies in West-ern blot and immunocytochemistry. The granzyme B proteinwas dramatically accumulated and cleaved to form activemolecular species which can activate directly a cascadeof caspases that can bypass the mitochondrial destruction[20]. Indeed, no cytochrome C release from granulosa cellmitochondria was evident during initial stages of apoptosis(Sasson and Amsterdam, unpublished). Also perforine thatis responsible for the release of granzyme B from granuleswas expressed in the granulosa cells[20]. This is the firstdemonstration that granzyme like proteins can reside and bereleased by cells other than cytotoxic lymphocytes or naturalkiller cells.
Another protein, apoptotic repressor (ARC) which wasuniquely found to reside in heart muscle[21] was found to beexpressed in granulosa cells. This protein could significantlycontribute to the protection of the cell against mitochondrialdestruction during apoptosis, since it contains a caspase re-cruitment domain and can preserve mitochondrial function.
1.4. Crosstalk among survival and death signals ingranulosa cells
The possibility of culturing purified primary granulosacells and immortalized steroidogenic cells that express a

864 A. Amsterdam et al. / Steroids 68 (2003) 861–867
Fig. 3. Signal transduction pathways and possible crosstalk among signals that control steroidogenesis and programmed cell death in mature ovariangranulosa cells, and in immortalized steroidogenic granulosa cells. Stars indicate signals that may lead to both steroidogenesis and apoptosis; trianglesindicate signals that may function as survival factors. The increased level of tyrosine phosphorylation[35] induced either by growth factors or byinhibiting tyrosine phosphatases (Pase) via vanadate was found to synergise with cAMP-generated signals in the induction of progesterone production[36]. Synergism in progesterone production between gonadotropin-releasing hormone (GnRH) stimulation, mediated at least in part by phospholipaseD (PLD), and cAMP-generated signals was also reported recently[37]. Culturing of granulosa cells on extracellular matrix, in the form of a nativebasement membrane, enhanced progesterone production by itself and augmented cAMP-induced steroidogenesis[7,33,38,39]in primary cells, while itattenuated cAMP-induced steroidogenesis in the immortalized cells. ‘Cytoskeleton response’ refers to the rearrangement of the actin cytoskeleton, whichoccurs during both luteinization[19,32] and apoptosis of granulosa cells[3,40]. ‘Other metabolic events’ refers to upregulation of the steroidogenicenzymes, sterol carrier protein 2 (SCP2), the steroidogenic transcription factor SF1/Ad4-binding protein and the StAR protein (reviewed in[40]).DG, diacylglycerol; EGF, epidermal growth factor; IGF, insulin-like growth factor; IP-3, inositol 1,4,5-trisphosphate; PA, phosphatidic acid; PDE,phosphodiesterase; PIP 2, phosphatidylinositol 4,5-bisphosphate; PKA, protein kinase A; PKC, protein kinase C; PLC, phospholipase C; R, receptor;TPA, 12-O-tetradecanoylphorbol-13-acetate; AC, adenylate cyclase; Gs, G-stimulatory protein (modified from[3]).
temperature sensitive mutant of p53 (val 135 p53), whichbehaves as an oncogenic p53 at 37◦C and as an inducer ofapoptosis at 32◦C permits a comprehensive analysis of themolecules involved in crosstalk between signaling pathwaysfor death and life in a well defined and synchronised apop-totic system[8,10,14]. Basic FGF exerts a survival activityon p53-induced apoptosis, and cAMP augments p53-inducedapoptosis[8]. We discovered that bFGF enhances MDM2expression while cAMP attenuates MDM2 expression[3,8].Therefore, since MDM2 is known to block p53 activity itcan be concluded that MDM2 plays a pivotal role in thecrosstalk between bFGF and p53-generated signals[8,22] aswell as between cAMP-generated signals and p53 signaling[3,8]. Another interesting crosstalk between the FGF recep-tor system and the p53-mdm2 system was revealed recently.When the FGFR3 G37R mutation for human dwarfness wasintroduced into mice by a knock-in technique, the mice be-came dwarfs and the females suffered from infertility. Anal-ysis of the ovaries revealed absence of ovulation, and thusabsence of corpora lutea formation and very intensive apop-tosis in the granulosa cells that never reached stages of theGraafian follicle[23]. Interestingly, p53 intracellular levelsin the dwarf mice were very high compared to normal an-imals, while progesterone production was very low. Thesephenomena implicate malfunction of growth factor recep-
tors associated with: p53 expression, ovarian cell death andattenuation of progesterone production.
Glucocorticoids such as dexamethasone and hydrocor-tisone enhance steroidogenesis in granulosa cells whileco-stimulation with gonadotropin/cAMP enhances forma-tion of progesterone[11,13,24]. Glucocorticoids were alsofound to exert protective effects on apoptosis induced byserum deprivation, cAMP, p53 and TNF-�-induced apop-tosis [11,13,24,25]. We found that the protective effectsare exerted by up-regulation of BCL2 and/or attenuationof its degradation[13,26]. Glucocorticoids, therefore, mayplay an important role in vivo by accelerating the healingprocess of the ruptured follicle subsequent to ovulation andduring formation of the corpus luteum[27,28].
1.5. The role of cell contact and intracellularcommunication in controlling apoptosis
Granulosa cells communicate both in vivo and in vitro viagap junctions, and they establish adherence junctions thatare specialized zones of cell–cell contact[29–34]. It seemsthat the integrity of gap junctions plays an important rolein the survival of granulosa cells. This conclusion is drawnfrom the fact that gap junctions become larger and appearin higher incidence subsequent to culturing of the cells on

A. Amsterdam et al. / Steroids 68 (2003) 861–867 865
Fig. 4. Organization of intracellular organelles during apoptosis of immortalized steroidogenic granulosa cells. (A) Electron microscopic imageof anon-apoptotic immortalized human granulosa cell demonstrates numerous mitochondrial (m) and microvilli (v) on the cell surface (10,000×); (B) apoptoticcell induced by activation of p53 demonstrating numerous cytoplasmic blebs (b). Chromatin in the nucleus that acquires irregular shape is condensed(n). Mitochondria remain intact in the central body of the cell (10,000×); (C) enlargement of the central part of the cell. Mitochondria in the centralpart of the apoptotic cell contain well-defined crystae and seem intact (30,000×). (D–D′ ′) Apoptotic cell double stained with phalloidin rhodamine (red)for actin cytoskeleton and with antibodies to StAR mitochondrial protein (green). Three optical sections bottom (D), middle (D′) and top (D′ ′) part ofthe cell. Note the rearrangement of the actin cytoskeleton in the cell periphery (red) and the intactness of the steroidogenic mitochondria (green) whichare highly clustered (3000×). Modified from Amsterdam et al.[3] and Hosokawa et al.[8] with permission.

866 A. Amsterdam et al. / Steroids 68 (2003) 861–867
native ECM like bovine corneal basement membrane[31]and/or in the presence of LH, FSH or glucocorticoids[26].Following stimuli for apoptosis, integrity of the junctions isinterrupted[26], but it is not clear yet whether apoptotic sig-nals cause the breakdown of gap junctions or whether break-down of gap junctions initiates and accelerates the apoptoticprocess. Connexin 43 is a major component of granulosa cellgap junctions, and its expression is clearly elevated both bygonadotropins/cAMP[34] and glucocorticoids[26]. Adher-ence type junction size and frequency were also found to beelevated by glucocorticoids, concomitantly with the eleva-tion of cadherin expression (Sasson and Amsterdam unpub-lished). Therefore, integrity of adherence and gap junctionsmay also play a role in the resistance of granulosa cells toapoptotic signals.
2. Conclusions
1. Ovarian cell death is critical for ovarian homeostasis.2. Initial steps of this process enhance, rather than attenuate
progesterone production, probably by bypassing mito-chondrial destruction and clustering of the steroidogenicorganelles containing the factors and enzymes respon-sible for conversion of cholesterol to pregnonolone, thefirst obligatory step for steroid hormone production.
3. Novel components discovered to be expressed in granu-losa cells such as ARC and granzyme B protect the mi-tochondria during the apoptotic process, bypassing thenecessity of mitochondrial signaling for apoptosis suchas the release of cytochrome C.
4. Enhancement of progesterone release from apoptoticcells may sharpen the cyclicity of progesterone releasein the estrous/menstrous cycle. This process may play animportant role in the maintenance of the newly formedcorpus luteum, while a sharp decline of progesteroneproduction may facilitate the demise of the old corpusluteum.
Acknowledgements
We thank Dr. Fortuna Kohen for helpful discussion. Thiswork was supported by grants from the Yad Avraham Centerfor Cancer Research and the Center for Scientific ExcellenceResearch supported by a grant from La Fondation Raphaelet Regina Levy at the Weizmann Institute of Science, Re-hovot Israel. AA is the incumbent of the Joyce and Ben B.Eisenberg professorial Chair of Molecular Endocrinologyand Cancer Research at the Weizmann Institute of Science.
References
[1] Amsterdam A, Selvaraj N. Control of differentiation, transformation,and apoptosis in granulosa cells by oncogenes, oncoviruses, andtumor suppressor genes. Endocr Rev 1997;18:435–61.
[2] Amsterdam A, Dantes A, Hosokawa K, Schere-Levy CP, Kotsuji F,Aharoni D. Steroid regulation during apoptosis of ovarian follicularcells. Steroids 1998;63:314–8.
[3] Amsterdam A, Gold RS, Hosokawa K, Yoshida Y, Sasson R, JungY, et al. Crosstalk among multiple signaling pathways controllingovarian cell death. Trends Endocrinol Metab 1999;10:255–62.
[4] Hurwitz A, Adashi EY. Ovarian follicular atresia as an apoptoticprocess: a paradigm for programmed cell death in endocrine tissues.Mol Cell Endocrinol 1992;84:C19–23.
[5] Tilly JL, Kowalski KI, Johnson AL, Hsueh AJ. Involvement ofapoptosis in ovarian follicular atresia and postovulatory regression.Endocrinology 1991;129:2799–801.
[6] Almog B, Gold R, Tajima K, Dantes A, Salim K, Rubinstein M, etal. Leptin attenuates follicular apoptosis and accelerates the onset ofpuberty in immature rats. Mol Cell Endocrinol 2001;183:179–91.
[7] Aharoni D, Meiri I, Atzmon R, Vlodavsky I, Amsterdam A.Differential effect of components of the extracellular matrix ondifferentiation and apoptosis. Curr Biol 1997;7:43–51.
[8] Hosokawa K, Aharoni D, Dantes A, Shaulian E, Schere-LevyC, Atzmon R, et al. Modulation of Mdm2 expression andp53-induced apoptosis in immortalized human ovarian granulosacells. Endocrinology 1998;139:4688–700.
[9] Braw RH, Bar-Ami S, Tsafriri A. Effect of hypophysectomy onatresia of rat preovulatory follicles. Biol Reprod 1981;25:989–96.
[10] Aharoni D, Dantes A, Oren M, Amsterdam A. cAMP-mediatedsignals as determinants for apoptosis in primary granulosa cells. ExpCell Res 1995;218:271–82.
[11] Hosokawa K, Dantes A, Schere-Levy C, Barash A, Yoshida Y, KotsujiF, et al. Induction of Ad4BP/SF-1, steroidogenic acute regulatoryprotein, and cytochrome P450scc enzyme system expression innewly established human granulosa cell lines. Endocrinology1998;139:4679–87.
[12] Breckwoldt M, Selvaraj N, Aharoni D, Barash A, Segal I, Insler V,et al. Expression of Ad4-BP/cytochrome P450 side chain cleavageenzyme and induction of cell death in long-term cultures of humangranulosa cells. Mol Hum Reprod 1996;2:391–400.
[13] Sasson R, Winder N, Kees S, Amsterdam A. Induction of apoptosis ingranulosa cells by TNF alpha and its attenuation by glucocorticoidsinvolve modulation of Bcl-2. Biochem Biophys Res Commun2002;294:51–9.
[14] Keren-Tal I, Suh BS, Dantes A, Lindner S, Oren M, AmsterdamA. Involvement of p53 expression in cAMP-mediated apoptosis inimmortalized granulosa cells. Exp Cell Res 1995;218:283–95.
[15] Lindheim SR, Chang PL, Vidali A, Ferin M, Sauer MV. The utilityof serum progesterone and inhibin A for monitoring natural-cycleIVF-ET. Assis Reprod Genet 1998;15:538–41.
[16] Fanchin R, Righini C, Olivernnes F, Ferreira AL, de Ziegler D,Frydman R. Consequences of premature progesterone elevation onthe outcome of in vitro fertilization: insights into a controversy. FertilSteril 1997;68:799–805.
[17] Urman B, Alatas C, Aksoy S, Mercan R, Isiklar A, Balaban B.Elevated serum progesterone lvel on the day of human chorionicgonadotropin administration does not adversely affect implantationrates after intracytoplasmic sperm injection and embryo transfer.Fertil Steril 1999;72:975–9.
[18] Eva Ch, Svensson MD, Markstrom E, Shao R, Andersson M, BilligH. Progesterone receptor antagonists ORG 31710 and RU 486increase apoptosis in human periovulatory granulose cells. FertilSteril 2001;76:1225–31.
[19] Amsterdam A, Aharoni D. Plasticity of cell organization duringdifferentiation of normal and oncogene transformed granulosa cells.Microsc Res Tech 1994;27:108–24.
[20] FASEB. Novel genes modulated by FSH in normal and immortalizedFSH-responsive cells: new insights into the mechanism of FSHaction. FASEB J 2003;17:1256–66.
[21] Neuss M, Monticone R, Lundberg MS, Chesley AT, Fleck E, CrowMT. The apoptotic regulatory protein ARC (apoptosis repressor

A. Amsterdam et al. / Steroids 68 (2003) 861–867 867
with caspase recruitment domain) prevents oxidant stress-mediatedcell death by preserving mitochondrial function. J Biol Chem2001;276:33915–22.
[22] Shaulian E, Resnitzky D, Shifman O, Blandino G, Amsterdam A,Yayon A, et al. Induction of Mdm2 and enhancement of cell survivalby bFGF. Oncogene 1997;15:2717–25.
[23] Amsterdam A, Kannan K, Givol D, Yoshida Y, Tajima K, Dantes A.Apoptosis of granulosa cells and female infertility in achondroplasticmice expressing mutant fibroblast growth factor receptor 3G374R.Mol Endocrinol 2001;15:1610–23.
[24] Barkan D, Jia H, Dantes A, Vardimon L, Amsterdam A,Rubinstein M. Leptin modulates the glucocorticoid-induced ovariansteroidogenesis. Endocrinology 1999;140:1731–8.
[25] Sasson R, Tajima K, Amsterdam A. Glucocorticoids protect againstapoptosis induced by serum deprivation, cyclic adenosine 3′,5′-mono-phosphate and p53 activation in immortalized human granulosa cells:involvement of Bcl-2. Endocrinology 2001;142:802–11.
[26] Sasson R, Amsterdam A. Stimulation of apoptosis in humangranulosa cells from in vitro fertilization patients and its preventionby dexamethasone: involvement of cell contact and bcl-2 expression.J Clin Endocrinol Metab 2002;87:3441–51.
[27] Amsterdam A, Sasson R. The anti-inflammatory action of gluco-corticoids is mediated by cell type specific regulation of apoptosis.Mol Cell Endocrinol 2002;189:1–9.
[28] Amsterdam A, Tajima K, Sasson R. Cell-specific regulation ofapoptosis by glucocorticoids: implication to their anti-inflammatoryaction. Biochem Pharmacol 2002;64:843–50.
[29] Amsterdam A, Josephs R, Lieberman ME, Lindner HR. Organizationof intramembrane particles in freeze-cleaved gap junctions of ratgraafian rollicles: optical-diffraction analysis. J Cell Sci 1976;21:93–105.
[30] Amsterdam A, Knecht M, Catt KJ. Hormonal regulation of cyto-differentiation and intercellular communication in cultured granulosacells. Proc Natl Acad Sci USA 1981;78:3000–4.
[31] Amsterdam A, Rotmensch S, Furman A, Venter EA, Vlodavsky I.Synergistic effect of human chorionic gonadotropin and extracellular
matrix on in vitro differentiation of human granulosa cells:progesterone production and gap junction formation. Endocrinology1989;124:1956–64.
[32] Amsterdam A, Rotmensch S, Ben-Ze’ev A. Coordinated regulationof morphological and biochemical differentiation in a steroidogeniccell: the granulosa cell model. Trends Biochem Sci 1989;14:377–82.
[33] Amsterdam A, Rotmensch S. Structure–function relationships duringgranulosa cell differentiation. Endocr Rev 1987;8:309–37.
[34] Sommersberg B, Bulling A, Salzer U, Frohlich U, Garfield RE,Amsterdam A, et al. Gap junction communication and connexin43 gene expression in a rat granulosa cell line: regulation byfollicle-stimulating hormone. Biol Reprod 2000;63:1661–8.
[35] Peluso JJ. Putative mechanism through whichN-cadherin-mediatedcell contact maintains calcium homeostasis and thereby preventsovarian cells from undergoing apoptosis. Biochem Pharmacol1997;54:847–53.
[36] Aharoni D, Dantes A, Amsterdam A. Cross-talk between adenylatecyclase activation and tyrosine phosphorylation leads to modulationof the actin cytoskeleton and to acute progesterone secretion inovarian granulosa cells. Endocrinology 1993;133:1426–36.
[37] Amsterdam A, Dantes A, Liscovitch M. Role of phospholipase-dand phosphatidic acid in mediating gonadotropin-releasing hormone-induced inhibition of preantral granulosa cell differentiation.Endocrinology 1994;135:1205–11.
[38] Furman A, Rotmensch S, Dor J, Venter A, Mashiach S, VlodavskyI, et al. Culture of human granulosa cells from an in vitrofertilization program: effects of extracellular matrix on morphologyand cyclic adenosine 3′,5′-monophosphate production. Fertil Steril1986;46:514–7.
[39] Furman A, Rotmensch S, Kohen F, Mashiach S, Amsterdam A.Regulation of rat granulosa cell differentiation by extracellularmatrix produced by bovine corneal endothelial cells. Endocrinology1986;118:1878–85.
[40] Amsterdam A, Selvaraj N. Control of differentiation, transformation,and apoptosis in granulosa cells by oncogenes, oncoviruses, andtumor suppressor genes. Endocr Rev 1997;18:435–61.