soil-borne diseases of sitka spruce seedlings in a forest nursery
TRANSCRIPT

Vol. 45, Part 4
Trans. Brit. mycol. Soc. 45 (4), 433-448 (1962).
December 1962
MYC·45
SOIL-BORNE DISEASES OF SITKA SPRUCESEEDLINGS IN A FOREST NURSERY
By E. W. BUXTON, INDU SINHA AND VALERIE WARD
Rothamsted Experimental Station, Harpenden, Herts
(With Plate 9 and 8 Text-figures)
Seedlings of Sitka spruce (Picea sitchensis Carr.) growing at Old Kenningtonforest nursery, Oxford, showed a succession of troubles in 1959: damping-off inspring, browning of leaves and roots in summer, and stunting without rootdamage in autumn, all on experimental plots treated with formalin each yearfor 5 years. Plants grew normally on adjacent plots treated only once withformalin.
The microfloras of the rhizosphere and of seedling roots were examined atrz-day intervals from early May to October. Only thirteen genera of fungiwere identified in plots treated annually with formalin; the most prevalent fungiwere Pythium spp., Fusarium oxysporum, F. roseum, Penicillium spp. and Trichoderma viride. The frequency in the rhizosphere of Pythium spp., numbers ofbacteria and to a lesser extent those of F. oxysporum, decreased as the seasonadvanced, whereas Trichoderma viride, Penicillium spp., F. roseum and Bacilluscereus var. mycoides increased greatly.
Pathogenicity tests with ninety-six isolates of fungi and bacteria showed thatdamping-off was caused by Pythium spp. and browning by F. oxysporum; someisolates of F. oxysporum caused stunting, as also did F. roseum. Isolates of Pythiumand Fusarium made early in the season were more pathogenic than those isolatedlater.
The prolific rhizosphere microflora in the formalin-treated plots may resultpartly from the introduction offungi and bacteria on seed, which by plating onagar media was shown to be contaminated. However, many isolates of Pythiumand Fusarium and a few of Gliocladium roseum, Mortierella and Penicillium fromformalin-treated soil also tolerated high levels of formalin in agar media.Isolates of F. osysporum from adjacent soils untreated with formalin were lesstolerant, suggesting that tolerance may have developed as a result of repeatedtreatment.
Members of the Chemistry Department, Rothamsted ExperimentalStation, began experiments in 1945 to see how mineral fertilizers, croprotation and soil sterilization affected the growth of Sitka spruce seedlingsin forest nurseries in southern England (Benzian, 1961). Among theseedlings, grown in close stands year after year on the same ground, suchtroubles as pre-emergence and immediate post-emergence damping-off,browning, stunting, root-rot and die-back became severe in some plots inthe older nurseries such as that at Old Kennington, Oxford. None of thesymptoms seems to have been caused by the fertilizer treatments, and evidence increasingly indicated that many resulted from attacks by soil-bornemicro-organisms. The strongest evidence for this supposition is thatapplying formalin or chloropicrin to the plots usually alleviates the troubletemporarily.
During 1958, seedlings were severely stunted on plots at Kennington
Vol. 45, Part 3 was issued 20 September 1962
28

434 Transactions British Mycological Societythat had been treated with formalin each spring for the previous 5 years.This situation called for investigation for at least two reasons: first,formalin should have prevented damage from soil-borne micro-organisms;secondly, the symptoms were not associated with nutritional treatments.
At Old Kennington nursery, a 40-plot factorial experiment (no. K82 ),consisting of a randomized block of plots, each 1 sq.yd. and separatedfrom each other by 18 in. pathways, was designed to test accumulatedeffects of various levels of phosphate, potassium and magnesium onseedling growth. The nutritional problems can be studied only when thesoil is sterilized, because without this the seedlings do not grow wellenough to give typical responses; formalin is therefore applied to the wholeexperiment every February. Seed is sown in early March, and numbers ofplants and their height are recorded at regular intervals.
The work described here was done to seek the cause ofthe disorders andto investigate the apparently anomalous occurrence of soil-borne diseasesin formalin-treated soil.
Ware up (1951, 1952) showed that formalin applied to forest nurserysoils eliminated the pathogenic fungi usually responsible for damping-off.The paradoxical situation at Kennington, where repeated formalin treatment had failed to do this, suggested the following approaches: (I) analysisof the root-surface and rhizosphere microfloras of seedlings, (2) tests ofprevalent fungal species for pathogenicity, (3) tests for acquired toleranceof formalin by soil micro-organisms, (4) tests to see whether any rootsurface and rhizosphere fungi originated from contaminated seed.
DISEASE SYMPTOMS
Examination of the Kennington experiments at frequent intervalsduring 1958 and 1959 showed that the Sitka troubles were of three kinds:(I) damping-off at and before seedling emergence, (2) browning and dieback during late summer, (3) stunting without discoloration, whichbecame increasingly apparent as the season advanced.
443316
Percentageemergence
No. plants emergedfrom 4000 seeds
17581314654
FormalinChloropicrinNo treatment
Damping-off
Damping-off in seedling conifers has been reported to be caused byPythium ultimum, P. debaryanum, Rhizoctonia sp. or Fusarium oxysporum(Gifford, 1911; Hartley, 1921; Roth & Riker, 1943a, b, c; Tint, 1945;Warcup, 1951, 1952; Vaartaja & Cram, 1956; Griffin, 1958). At Kennington, as elsewhere, damped-off Sitka seedlings either never brokethrough the soil surface or grew for only a few days and collapsed afterbecoming weakened at the stem base. Roots were withered and the plantscould be easily removed from the soil (PI. 9, fig. I).
In an additional experiment at Kennington (K89), plots were treatedonce, during 1959, with formalin or chloropicrin, and this partiallycontrolled damping-off:

Sitka spruce diseases. E. W. Buxton et al. 435Germination percentage of Sitka seeds is normally low and even in
plots treated with formalin was only 46 %, but without fungicide treatmentitwas only about one-third of the 44 %emergence. PI. 9, figs. 2 and 4,show the differences between treated and untreated plots in this experiment. Griffin (1958) found that damping-off of conifer seedlings wasrestricted to near-neutral or alkaline soils, and Vamos (1954) observedthat the disease was worse in clay than in sandy soils. The soil at Kennington is a heavy silty loam of pH 6·8.
Browning
Seedlings turned brown during July and August, especially after prolonged periods of high soil temperature (Text-fig. 1 and PI. 9, figs. 2 and 4).Browning has been ascribed to several fungi and to the nematode Haplo-
26 Aug.
12
11...... 10
~ 9
8
76
5
432
1OL-_L.- -L. --I
262422
~ 20~ 18
~ 16coQ. 14..,~ 12 - . .-------0~10~
10 8 __---~6 ~-
4 •2OL-L.- -L. ---l
13 July
lamus sp. The first symptoms are yellowing of the leaf tips, resemblingsigns of magnesium deficiency, but, unlike this condition, the chlorosisdevelops rapidly and the whole plant soon turns pale brown. The stemsbecome wiry and the roots discoloured (PI. 9, fig. 3). The root cortexturns dark brown and later black, and shreds away from the vascularcylinder, which also ultimately turns brown and is sometimes flecked withbright red patches.
Stunting
Stunted seedlings occurred in patches unrelated to nutritional treatments, thus indicating a localized soil-borne and probably microbialcause. Affected plants remained green and their roots showed no lesions.There was no evidence to suggest that stunting was related to damping-offor browning; plants smaller than average at an early stage were no more
28-2

436 Transactions British Mycological Societylikely to turn brown later than were tall or average-sized plants (Textfig. 2). Text-fig. 3 shows the distribution of stunted plants at Kenningtonin 1958.
II III IV
10
9
8
7
6
5
4
3
2
VS vs VT T
T vs VS VS
T S VT S
T A VS VS
VT A A S
VS S VS A
VT VT T VS
T T T VS
VT T VS T
T S VT VT
Text-fig. 3. Distribution of stunting in plots of Sitka seedlings on Expt. K 82 at OldKennington, 1958. S = Stunted, > 0'5 in. from mean; VS = very stunted, > 1'0 in.from mean; A = of average height; T= taller than mean by> 0'5 in.; VT = tallerthan mean by > 1'0 in,
THE RHIZOSPHERE MICROFLORA OF SITKA SEEDLINGS
To identify the micro-organisms prevalent at different times during thegrowing season on or near roots of seedlings, rhizosphere soil and pieces ofroots were plated on agar media at fourteen I2-day intervals from earlyMay to October 1959. On each sample occasion, ten plants were taken atrandom from each of eight plots from the forty-plot experiment K82.Four of the eight plots had produced healthy plants in 1958, and four hadproduced stunted plants. Samples were also taken from plots in experiment K89. Soil was also sampled at some distance from roots to comparerhizosphere and soil microfloras.
Soil adhering to roots, forming a layer about I mm. deep, was regardedas rhizosphere soil. This was washed from each plant in 10 ml. steriledistilled water, the resulting soil suspension diluted to II1000, and o· I ml.

Sitka spruce diseases. E. W. Buxton et al. 437of the dilution was spread over an agar medium consisting of (g./l.): yeastextract, 5; sucrose, 20; NaNOa, 2; KH2P04, 1; KCI, 0'5; MgS04 • 7H20,0'5; FeS04, 0'01; agar, 20; adjusted to pH 5'5 with phosphoric acid.Roots were washed in tap water for 1 min. and ten 0'5 em. root lengthsfrom each plant were plated and incubated at 25° C. for 1 day, followedby 5 days in daylight at room temperature. At each sampling time,800 root fragments and 80 aliquots of rhizosphere soil suspension wereplated. The soil suspensions were filtered and dried to estimate the dryweight of soil. A total of 1320 plants from experiment K82 and about500 from K89 were treated in this way, and rhizosphere and root surfaceorganisms were estimated from 3860 agar plates.
RESULTS
Few soil-borne fungi could be expected from plots that had been treatedwith formalin. Warcup (1951, 1952) showed that formalin-treated forestnursery soilwas slowlyre-colonized and contained fewfungi even 18monthsafter treatment, whereas up to fifty genera were represented in untreatedsoil. At Kennington, thirteen genera were recorded during the 8 monthsafter formalin treatment (Table 1).
Table 1. Rhieosphere androot surface micro-organisms isolatedfrom Sitka seedlings
ActinornycetesAspergillus niger van Teigh.Bacillus cereus var. mycoidesBacteria spp.Cephalosporium coremioides RailloCylindrocarpon radicicola Wollenw.Fusarium oxysporum Schlect. ex Fr.F. roseum LinkGliocladium roseum Bain.Mortierella spp.
Mucorhiemalis WehrnerPenicillium spp.P. lilacinum ThornP. luteum ZukalPhoma spp.Pythium spp.Rhizopus stolonifer (Ehr.) LindSporobolomyces sp.Stysanus stemonites (Pers.) CordaTrichoderma viride Pers. ex Fr.
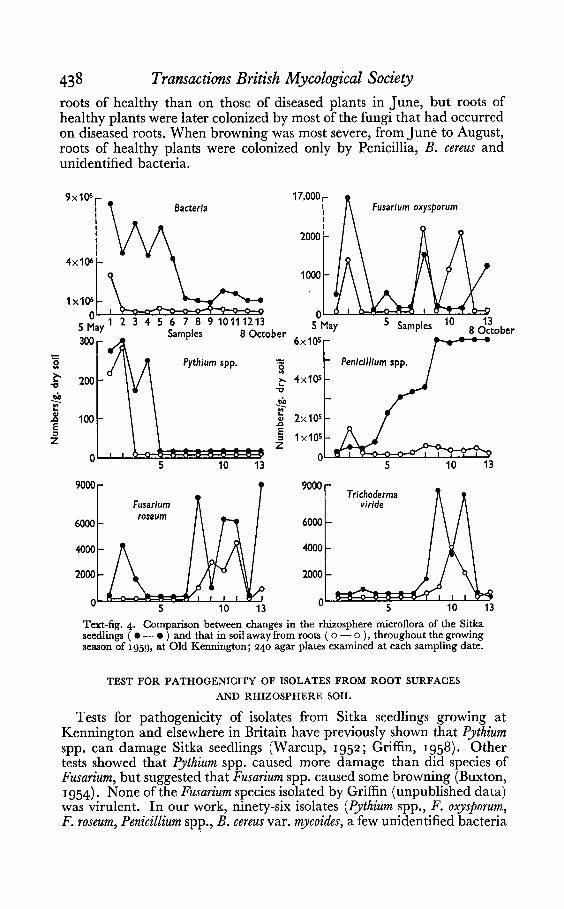
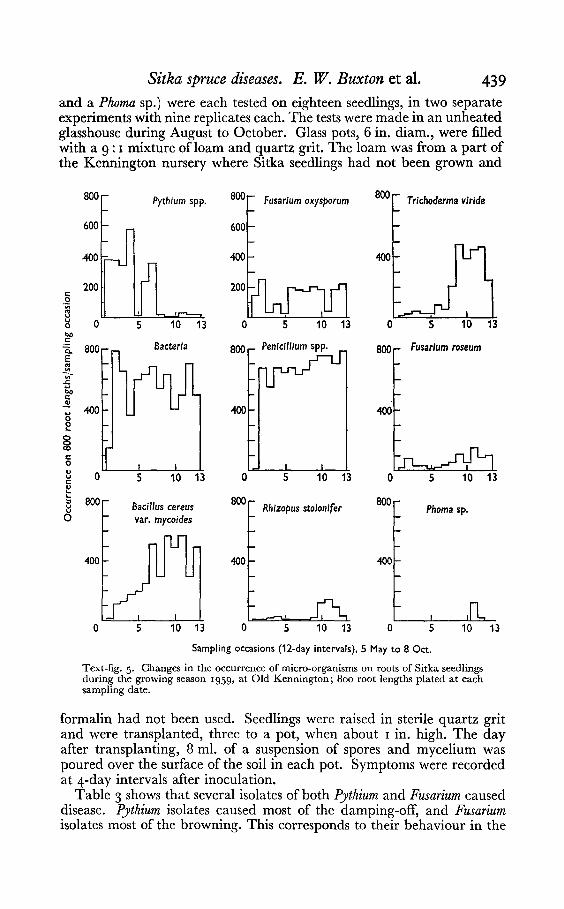
Few specieswere isolated very early in the season from either rhizospheresoil or root surfaces (Text-figs, 4 and 5). The numbers of some microorganisms fluctuated both on roots and in the rhizosphere as the seasonadvanced. For example, Trichoderma viride and Bacillus cereus var. mycoideswere rare at first on root surfaces but abundant later. Most speciesoccurred more often in the rhizosphere than in soil away from roots.
Both Fusarium oxysporum and Pythium spp. were common on roots atmost stages of plant growth up to mid-August, although F. oxysporum wasless common than Pythium spp. early in the season. Pythium spp. wereisolated mainly from damped-off plants, but F. oxysporum always frombrowned plants. During July, Pythium spp. began to disappear from rootsand had almost gone by October. They were not found in rhizospheresoils after June. Fusarium roseum increased towards the end of the seasonboth in the rhizosphere and on root surfaces. Warcup (1952) examinedroots of Sitka seedlings on six occasions between late May and midAugust, and found similar changes in the prevalence of Pythium andFusarium species. Table 2 shows that there were fewer micro-organisms on
438 Transactions British Mycological Society
roots of healthy than on those of diseased plants in June, but roots ofhealthy plants were later colonized by most of the fungi that had occurredon diseased roots. When browning was most severe, from June to August ,roots of healthy plants were colonized only by Penicillia, B. cereus andunidentified bacteria.
13105
.~ Penicillium spp.
t- 4x10s~
~
t 2x1OS.c
5 1x10Sz
Pythium spp.
9x1Q6 r 17.000rI Bacteria I Fusarium oxysporumI II II II 2000 'III
4x1Q61000
1x 1Q60 234 5 6 7 8 9 10111213 0
5 May 1Samples 8 October
5 May300 6x10s
'2t 200
~~
..8 100E
"Z
9000
6000
2000
Fusariumroseum
9000Trichoderma
viride
6000
2000
5 10
Text-fig. 4. Comparison between changes in the rhizosphere microflora of the Sitkaseedlings ( • -. ) and that in soil away from roots ( 0 - 0 ), throughout the growingseason of 1959, at Old Kennington ; 24 0 agar plates examined at each sampling date.
TEST FOR PATHOGENICITY OF ISOLATES FROM ROOT SURFACES
AND RHIZOSPHERE SOIL
Tests for pathogenicity of isolates from Sitka seedlings growing atKennington and elsewhere in Britain have previously shown that Py thiumspp. can damage Sitka seedlings (Warcup, I 952; Griffin, 1958). Othertests showed that Py thium spp. caused more damage than did species ofFusarium, but suggested that Fusarium spp. caused some browning (Buxton,1954). None of the Fusarium species isolated by Griffin (unpublished data)was virulent. In our work, ninety-six isolates (Py thium spp. , F. oxysporum,F. roseum, Penicilliumspp., B. cereus var. my coides, a few unidentified bacteria
Sitka spruce diseases. E. W. Buxton et a1. 439and a Phoma sp.) were each tested on eighteen seedlings, in two separateexperiments with nine replicates each. The tests were made in an unheatedglasshouse during August to October. Glass pots, 6 in. diam., were filledwith a 9 : I mixture of loam and quartz grit. The loam was from a part ofthe Kennington nursery where Sitka seedlings had not been grown and
800 Pythium spp. 800 Fusarium oxysporum 800 Trichoderma viride
o
400
800 Fusarium roseum
400
600
400
10 13
Bacteria
400
~
600
400
200s.;;;G~bOc:
Q.. 800E'"..!:!..'"..c:li'oc:..2
RhizopUS stolonifer
~eo1:1c:~
5 800~o
10 13
Bacillus cereusvar, mycoides
o
800
5 10 13
800Phoma sp.
400
o 5 10 13
400
o 5 10 13
Sampling occasions (12-day intervals), 5 May to 8 Oct.
Text-fig. 5. Changes in the occurrence of micro-organisms on roots of Sitka seedlingsduring the growing season 1959, at Old Kennington; 800 root lengths plated at eachsampling date.
formalin had not been used. Seedlings were raised in sterile quartz gritand were transplanted, three to a pot, when about I in. high. The dayafter transplanting, 8 ml. of a suspension of spores and mycelium waspoured over the surface of the soil in each pot. Symptoms were recordedat 4-day intervals after inoculation.
Table 3 shows that several isolates of both Pythium and Fusarium causeddisease. Pythium isolates caused most of the damping-off, and Fusariumisolates most of the browning. This corresponds to their behaviour in the

Table 2. Incidence of micro-organisms on roots of browned and healtfry Sitka seedlings (expressed as percentage of ~0
roots yielding each organism: mean of 80 root lengths)Bacillus
Tricho- cereusFusarium Pythium Rhieopus Fusarium derma var.
Bacteria oxyspMUtn spp, Penicillia sp. roseum oiride mycoides OthersBrowned plants
6th sample (27 Jan.) 81"8 36'4 1'3 59'1 2'3 ° 0 2,6 0~13th sample (19 Sept.) 56'3 40 '6 15"6 93'8 0 12'5 21'9 53'1 0~
Healthy plants
~6th sample (27 Jan.) 87'5 0 0 70 0 0 0 56'3 0r gth sample (19 Sept.) 58'4 16·6 8'4 100 8'4 18,8 16·6 79'1 0 ....
~.
Table 3. Results of inoculation tests of isolates from root surfaces and rhizospheres ~
of Sitka seedlings (as % diseased plants, at four stages) a·~Pythiumspp. Fusarium oxysporum Fusarium roseum
Days after Days after Days after~inoculation inoculation inoculation
Sample Date , , , ("), \ , I c:::>no. isolated 4 9 14 19 4 9 14 19 4 9 14 19 B'"'
5 May 26 64 (10)* (-)* (-) otiI 37 59 - - - - - - - - ~ .2 16 May 47 75 83 84 ?) - - - - (-) - - - - (-) ~
3 27 May 43 64 79 83 3) 10 16 28 37 (4) - - - - (-) .......4 7 June 43 67 81 82 (5) 47 86 92 92 (2) - - - - (- ) V)5 17 June - - - - (-) 20 43 63 65 (3) - - - - (- ) c:::>
(")6 27 June 12 25 44 46 (3) 16 19 40 52 (4) - - - - (-) ~.
7 9 July II 33 33 39 (I) 0 39 55 55 (I) 3 36 42 42 (2) Q8 18 July - - - - (-) 0 28 37 37 (4) 2 24 33 36 (3)9 29 July - - - - (-) 0 9 19 21 (6) II II 22 22 (I)
10 7 Aug, - - - - (-) 4 II 30 32 (4) 5 5 19 31 (2)II 19 Aug. - - - - (-) 0 5 24 27 (2) - - - - (-)12 3 Sept, - - - - (-) I 22 35 40 (6) 0 3 14 20 (2)13 19 Sept. - - - - (-) 0 30 38 44 (5) 0 0 31 39 (2)
* Figures in parentheses are the number of isolates tested on each occasion; (-) = no isolates available,

Sitka spruce diseases. E. W. Buxton et al. 441plots, where Pythium spp. were more prevalent on roots early in the seasonwhen there was damping-off, whereas Fusarium spp., especially F. roseum,were abundant later.
One unidentified bacterium killed the plants very soon after inoculation. Neither Phoma species nor any isolate of Penicillium nor Bacillus cereus,which were abundant on roots throughout most ofthe season, were pathogenic. Text-fig. 6 shows differences in the virulence of Pythium andFusarium isolates obtained at different times of the year.
••
18••• ••
12 • Fusarium oxysporum
• .,•• ••• .,8
• I••• • I·• •4 • •• •••• •0 5 10 13
•
• •
Pythium spp.
•
8
I· • •
.. :.,..I'
12 •• •
18
'" 4...e •'""ii.."
~ 0'"~'0'0.. 18III..,E'"Z Fusarium roseum
12
•• •8 •
••4 •• •
OL...._-~--""=-~I:-5 10 13
Sampling occasions (12-day intervals)
Text-fig. 6. Results of pathogenicity tests with root surface and rhizosphere isolatestaken at different times from Sitka seedlings during 1959.
Some Fusaria and a few Pythia were not pathogenic. Isolates ofPythium taken early in the season were more pathogenic than those takenlater; the Fusaria showed a similar but less clear trend. Decreasinglypathogenic Pythium isolates gave way to pathogenic Fusaria as the seasonadvanced, again suggesting that most of the early damping-off was causedby Pythium and the later browning by Fusarium species. It would beinteresting to see how inoculation with combined isolates of Fusarium andPythium would affect seedlings, for both could presumably infect seedlingssimultaneously in the field.
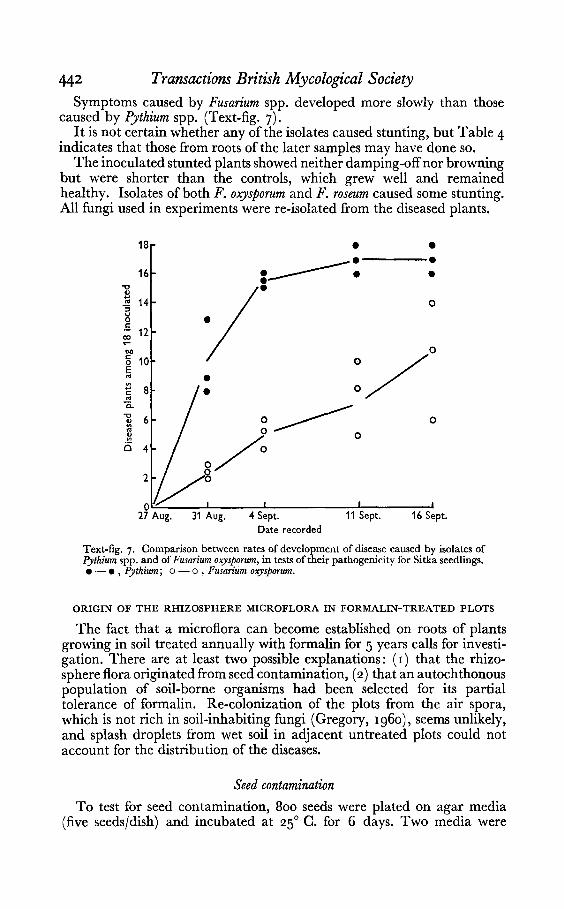
442 Transactions British Mycological SocietySymptoms caused by Fusarium spp. developed more slowly than those
caused by Pythium spp. (Text-fig. 7).It is not certain whether any of the isolates caused stunting, but Table 4
indicates that those from roots of the later samples may have done so.The inoculated stunted plants showed neither damping-offnor browning
but were shorter than the controls, which grew well and remainedhealthy. Isolates of both F. oxysporum and F. roseum caused some stunting.All fungi used in experiments were re-isolated from the diseased plants.
o
o
16 Sept.11 Sept.4 Sept.Date recorded
-0
~"3g.S 12co.....
• e.-----e/=-----. .
;':: :/0]6 o~
L o~ 0
2//01.:.::-__--1 ......L -'- -4
27 Aug. 31 Aug.
Text-fig. 7. Comparison between rates of development of disease caused by isolates ofPythium spp. and of Fusarium oxysporum, in tests of their pathogenicity for Sitka seedlings.• - • , Pythium; 0 - 0 , Fusarium osysporum.
ORIGIN OF THE RHIZOSPHERE MICROFLORA IN FORMALIN-TREATED PLOTS
The fact that a microflora can become established on roots of plantsgrowing in soil treated annually with formalin for 5 years calls for investigation. There are at least two possible explanations: (I) that the rhizosphere flora originated from seed contamination, (2) that an autochthonouspopulation of soil-borne organisms had been selected for its partialtolerance of formalin. Re-colonization of the plots from the air spora,which is not rich in soil-inhabiting fungi (Gregory, 1960), seems unlikely,and splash droplets from wet soil in adjacent untreated plots could notaccount for the distribution of the diseases.
Seed contamination
To test for seed contamination, 800 seeds were plated on agar media(five seeds/dish) and incubated at 25° C. for 6 days. Two media were

Table 4. Isolates tifFusarium oxysporum and F. roseum that causedstunting, without discoloration, in pathogenicity tests on Sitka seedlings
Sample Plot Species of Mean heightno. no. pathogen of plants (in.)
9 8/9 F. oxysporum 0'969 8/4 F. oxysporum 0'70
10 5/4 F. oxysporum 0·6510 7/8 F. roseum 0·60II 5/5 F. oxysporum 0'9212 6/9/2 F. roseum 0'5412 8/4/1 F. oxysporum 0'5012 8/4/2 F. oxysporum 1'0013 8/4- Fusarium sp. 0'7 013 8/13/1 F. oxysporum 0·82
Controls (no inoculum) 1'39
Table 5. Isolates from Sitka seeds (treated with red lead or untreated) onDox +yeast and on Dox + rose-bengal agar media*
BacillusTricho- cereus Asper- Morti-
Fusarium Pythium Rhizopus derma var, gillus erella FusariumBacteria oxysporum spp. Penicillia sp. viride mycoides niger sp. roseum Others
TreatedDox 162 0 0 150 22 14 7 4 0 0 0Rose-bengal 0 0 0 189 2 35 0 7 2 0 0
UntreatedDox 160 0 0 175 46 13 0 10 0 4- 0Rose-bengal 8 I 0 157 15 45 a 4 3 0 5
* Figures represent number of seeds bearing micro-organisms, out of a total of 800 seeds plated: 200 of both treated and untreated onDox-l-yeast, and on rose-bengal agar.
Col)
~~
~~.....
~~
~
r~
fl.
tUJ

444 Transactions British Mycological Societyused: Dox-yeast, as already described, and Czapek-Dox + rose bengal,which suppresses bacteria and thereby allows more fungi to grow.
In forest nurseries, Sitka seed is often dusted with red lead, so both thisand untreated seed were tested. The microfloras that developed fromdusted and untreated seed were similar (Table 5).
The predominant seed-borne organisms were bacteria, Penicillium spp.,Aspergillus niger, Cylindrocarpon radicicola and Trichoderma viride. F. oxysporumwas found once, F. roseum on four seeds and Pythium spp. on none (PI 9,fig. 5). However, seed sown in sterile quartz sand and watered withsterile water often gave rise to seedlings that damped-off at an early stage.Both F. osysporum and Pythium spp. were isolated from these, stronglysuggesting that seeds contaminated by these fungi were not revealed byplating.
Table 6. Tolerance of different levels offormalin in laboratory tests ofsixty-sixisolates from rhirosphere of seedling Sitka spruce. Growth after 5 days (+);intolerance ( - )
Formalin concentrations'sNo. isolates in each I \
tolerance category Nil N/4 N/2 N 2N 4N 8N
Pythium sp.I + + + + + + +I + + + + +I + + + +I + + +3 + + +2 + +
Fusarium oxysporumII + + + + + + +14 + + + + + +6 + + + + +
F. roseum5 + + + + + + +3 + + + + + +I + + + + +
Penicillium sp.2 + + + + + + +
Mortierella sp.I + + + + + +
Gliocladium sp.I + + + + +
Bacillus cereusI + + + + + +
Bacteria2 + + + + +
ActinomyceteI + + + +
F. oxysporumFrom plots not treated with formalin (as controls)
4 + + + + +3 + + + +2 + + +
• N = 6% solution (v/v) at 0'1 ml.j'dish of 15 ml. agar.

'"u"c:o"8ul>OL1cuu..uc,
l>OC
~ 100el>O
'"uC~ SOuOJ
~ 20OJeuu..~
'"uCo
"8ul>OL1cu~
~
100
Text-fig. 8. Variation in tolerance of a range of formalin concentrations shown byPythium spp., Fusarium osysporum and F. roseum isolated from rhizospheres of Sitkaseedlings at different times of the year.

Transactions British Mycological Society
Tolerance offormalin by isolates
When Warcup (1951) treated forest-nursery soil with I gal.jsq.yd, of4 % formalin, fungi in the layer down to 5 in. below the surface wereeither eliminated or reduced to very low numbers. At Kennington,experiment K82 had received I gal.(sq.yd. of6 %formalin each Februaryfor 5 years, yet a rhizosphere microflora was present and seedling diseasesoccurred. This suggested that a formalin-tolerant microflora might havearisen. Isolates of each of the three pathogens (nine of Pythium spp., fortyof F. oxysporum, and nine of F. roseum) and a few others were tested forformalin tolerance. Formalin was incorporated in Czapek-Dox agar atthe rate of 0'1 ml.(dish of 15 ml. medium. Taking as standard solution (N)6 % (vlv) of a 38 % formaldehyde solution, this procedure gave thesame concentration as that obtained in a 4 in. layer of field soil at 50 %water-holding capacity. Tolerance offormalin was tested using concentrations of 8N, 4N, 2N, N, N(2 and N(4; agar without formalin was used as acontrol. Table 6 shows the extent to which the isolates within each speciesdiffered in tolerance.
Only one Pythium isolate grew at 8Nand six failed to grow at concentrations greater than N(2. All but two of the isolates of F. oxysporum grewat 2N and eleven grew, although very little, at 8N. Tolerance reachedlevels of at least 4N in eight isolates of F. roseum and in the isolates ofPenicillium, Mortierella, Gliocladium, B. cereus and two unidentified bacteria.Nine isolates of F. oxysporum from an adjacent plot untreated with formalinwere less tolerant than isolates of the same species made from experimentK82, treated annually for 5 years.
Text-fig. 8 shows that the ability to tolerate the higher concentrations offormalin seemed to depend on the time of year when the isolates werefound in the rhizosphere.
Pythia isolated early in the season were less tolerant than those isolatedlater. F. oxysporum isolated in mid-season (samples 6, 7, 8) tolerated thehighest level better than the cultures isolated very early or very late, butmany isolates made early in the season tolerated concentrations 4Nand zx.Text-fig. 8 also shows that F. oxysporum isolates were in general moretolerant than were Pythium isolates. F. roseum also showed a high degree oftolerance, and a peak tolerance near the 8th sample.
DISCUSSION
Our results show how complex is the problem of seedling disorders inSitka spruce. First, there are at least three primary pathogens involved,each causing a specific syndrome and each important at a different time.Secondly, there is the question of how these pathogens, and other membersof the soil microflora, come to be established in soil repeatedly treated witha fungicide. Are they survivors that have become selected for theirunusual resistance to formalin? Or are they introduced into the treatedsoil on the seed?
A situation in which several pathogenic fungi succeed each other on onehost, each existing as strains of different virulence, is not unusual. In the

Sitka spruce diseases. E. W. Buxton et al. 447experimental plots at Kennington, however, it seems that the strainswithin each genus that invaded the roots early in the season were morevirulent than those that came later. This could have various explanations.Virulence may be correlated with ability to re-colonize sterile soil, or withability to survive formalin treatment. The fact that soil treated only oncewith formalin was largely free from seedling disorders, which were common on soil treated annually, implies that virulence may be correlatedwith resistance to formalin. Soils treated with repeated doses offormalinpresent a highly selective environment for any formalin-tolerant strains offungi. Little is known of adaptation by fungi to toxic materials applied tosoil, but the higher levels of disease in the plots repeatedly treated withformalin than in those given a single dose indicate that tolerance may havedeveloped.
The equally likely possibility that a rhizosphere microflora may beestablished from seed-borne contaminants needs further investigation.However rarely a potentially pathogenic fungus may occur on seed, onceintroduced into partially sterile soils it suffers little or no competitionwhile establishing itself on the root. Seed disinfection, no less than croprotation, and a more thorough testing of the effects of formalin and othertoxins on the soil microflora, should be considered as a way of minimizingthe Sitka seedling disorders.
We thank the members of the Research Staffof the Forestry Commissionfor permission to examine experimental plots, and the staff of the ForestryCommission Nursery, Kennington, Oxford, for their generous assistance inrecording symptoms among seedlings. We are also greatly indebted toMiss B. Benzian, Chemistry Department, Rothamsted, for much help andadvice during the work. The photographs were taken by members of thePhotography Department, Rothamsted.
REFERENCES
BENZIAN, B. (1961). Nutrition problems in forest nurseries. ]. Sci. Fd Agric. 10, 637-644.BUXTON, E. W. (1954). Rep. Rothamst. expo Sta. for 1954, p. 94.GIFFORD, C. M. (19 I I). The damping-off of coniferous seedlings. Bull. Vt agric. Exp. Sta.
no. 157, pp. 140 - 171.GREGORY, P. H. (1960). Outdoor aerobiology. Endeavour, 19, 223-228.GRIFFIN, D. M. (1958). Influence of pH on the incidence of damping-off. Trans. Brit.
mycol. Soc. 41, 483-490.HARTLEY, C. (1921). Damping-off in forest nurseries. Bull. U.S. Dep. Agric. no. 934.ROTH, L. F. & RIKER, A. J. (I943a). Life history and distribution of Pythium and
Rhizoctonia in relation to damping-off of red pine seedlings. ]. agric. Res. 67, 129-I48.ROTH, L. F. & RIKER, A. J. (I943b). Influence of temperature, moisture and soil
reaction on the damping-off of red pine seedlings by Pythium and Rhizoctonia.]. agric. Res. 67, 273-293.
ROTH, L. F. & RIKER, A. J. (I 943c). Seasonal development in the nursery of dampingoff of red pine seedlings caused by Pythium and Rhizoctonia. ]. agric. Res. 67, 417-43 I.
TINT, H. (1945). Studies in the Fusarium damping-off of conifers. III. Relation oftemperature and sunlight to the pathogenicity of Fusarium. Phytopathology, 35,498-5 10•
VAARTAJA, O. & CRAM, W. H. (1956). Damping-off pathogens of conifers and ofCaragana in Saskatchewan. Phytopathology, 46, 391-397.

Transactions British Mycological SocietyVAMOS, R. (1954). A Feny6csemete dolese. [Damping-off of pine seedlings.] Erda, 3,
34-4°·WARCUP,J. H. (1951). Effect of partial sterilization by steam or formalin on the fungus
flora of an old forest nursery soil. Trans. Brit. mycol. Soc. 34, 519-532.W ARCUP, J. H. (1952). Effect of partial sterilization by steam or formalin on damping
off of Sitka spruce. Trans. Brit. mycol. Soc. 35, 248-262.
EXPLANATION OF PLATE 9
Fig. I. Damping-off in Sitka seedlings taken from the field. Left, damped-off seedling; right,healthy seedling.Fig. 2. Differences in emergence of Sitka seedlings on plots treated once with 1 gal./sq.yd.of6%formalin, and on untreated plots (plot on far left and the three on right of photograph weretreated; the two remaining plots were untreated).Fig. 3. Browning of Sitka seedlings, taken from the field in July. Left, browned seedling,showing blackening and shredding of root cortex; right, healthy seedling from the same experimental plot.Fig. 4. Differences in emergence and plant height on a plot treated with chloropicrin (rear) andon an untreated plot (foreground). Photographed August, 1959.Fig. 5. Fungi growing from Sitka seed plated on Dox-agar, Left, Penicillia and Trichodermaviride; right, Penicillia and one colony of Fusarium roseum.
(Accepted for publication 17 October 196I)

Trans. Brit. mycol. Soc. Vol. 45, Plate 9
"
" ,
3
(Facing p. 448)