senior thesis-analyzing the interactions between myogef and a component of erythrocyte cytoskeleton...
TRANSCRIPT

Fordham University
Analyzing the interactions between MYOGEF and a component of erythrocyte
cytoskeleton SPTA1
Dougan McGrath
Department of Biological Sciences
Dr. Qize Wei
May 5, 2015

2
Abstract
Erythroid spectrin, a major component of the membrane cytoskeleton of erythrocytes, plays a
central role in maintaining the shape and elasticity of red blood cells, thus conferring to
erythrocytes the capability of travelling through the circulatory system while withstanding shear
and stress in the process of circulation. The basic molecular organization of erythroid spectrin is
a heterodimer consisting of an alpha-subunit (SPTA1) and a beta-subunit (SPTB1). Mutations in
the SPTA1 or SPTB1 gene have been associated with hereditary elliptocytosis. Importantly, it
has also been shown that RhoA, a key component of Rho GTPase signaling, is localized to the
erythrocyte membrane and implicated in the regulation of erythrocyte cytoskeleton. However, it
is essentially unknown whether and how Rho GTPase signaling is implicated in regulating the
shape and elasticity of red blood cells. A previous screening in Dr. Wei’s laboratory has
identified SPTA1 as an interacting partner of MYOGEF, a guanine nucleotide exchange factor
that is capable of activating RhoA and is involved in the regulation of cell division and cell
migration. In this study, we constructed various cDNA fragments encoding different regions of
MYOGEF or SPTA1 and examined their interactions using yeast two-hybrid and in vitro pull-
down assays. Our results showed that the C-terminal region of MYOGEF interacted with the EF-
hand motifs that are situated at the C-terminal region of SPTA1. We propose that the MYOGEF-
SPTA1 interaction may lead to MYOGEF-mediated RhoA activation at the erythrocyte
membrane, thus contributing to the regulation of membrane cytoskeleton in erythrocytes.

3
Introduction
Erythroid spectrin, a major component of the membrane cytoskeleton of erythrocytes,
plays a central role in maintaining the shape and elasticity of red blood cells, thus conferring to
erythrocytes the capability of travelling through the circulatory system while withstanding shear
and stress in the process of circulation. The basic molecular organization of erythroid spectrin is
a heterodimer consisting of an alpha-subunit and a beta-subunit encoded by genes SPTA1 and
SPTA2, respectively. The lateral association between the alpha- and beta-chains forms a
spectrin dimer that in turn joins head-to-head to form a spectrin tetramer (see Figure 1). There is
an actin-binding domain at both ends of the spectrin tetramer. Therefore, both ends of the
spectrin tetramer bind to a cytoskeleton complex consisting of short actin filaments, protein 4.1,
and actin-binding proteins dematin, adducin, tropomyosin, and tropomodulin (see Figure 1).
Such interactions form a spectrin-actin network that is in turn linked to the cell membrane via
ankyrin, which binds to both spectrin and the transmembrane protein band 3 (Figure 1). In
addition, protein 4.1 binds not only to the ends of the spectrin tetramers, but also to the
cytoplasmic domain of the transmembrane protein glycophorin (see Figure 1). Therefore,
protein 4.1 also plays a critical role in linking the spectrin-actin network to the cell membrane
(Machnicka et al, 2011).
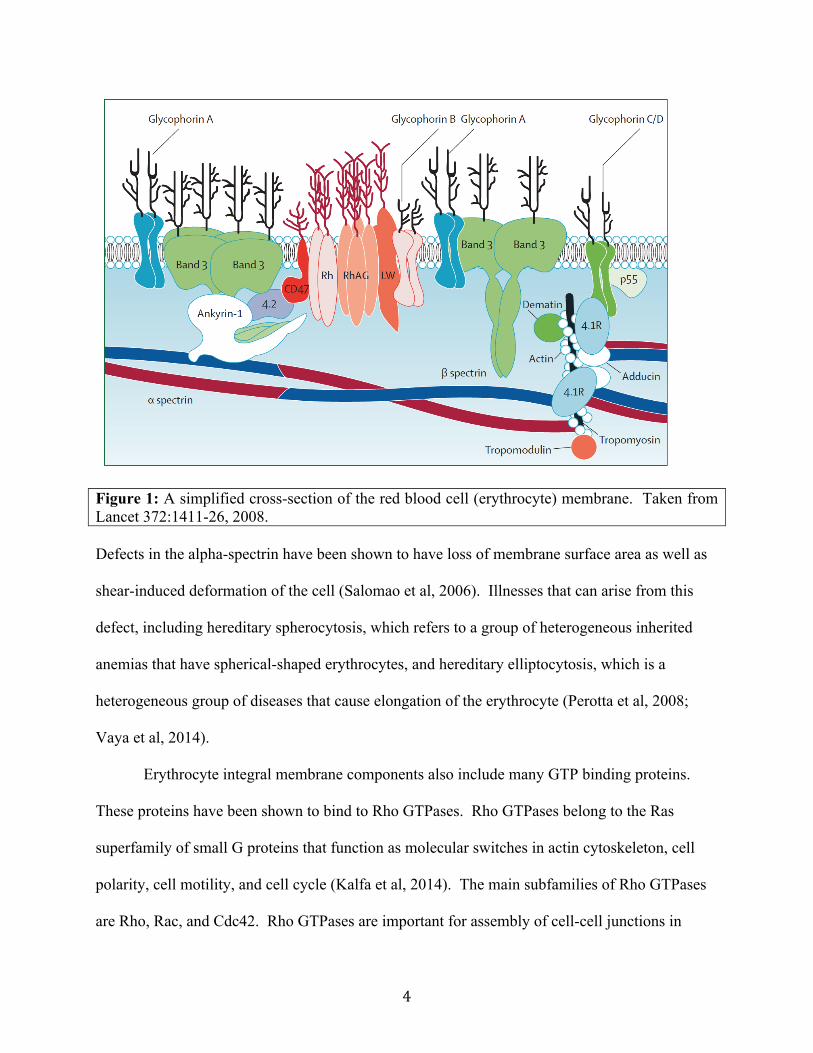
4
Figure 1: A simplified cross-section of the red blood cell (erythrocyte) membrane. Taken from Lancet 372:1411-26, 2008. Defects in the alpha-spectrin have been shown to have loss of membrane surface area as well as
shear-induced deformation of the cell (Salomao et al, 2006). Illnesses that can arise from this
defect, including hereditary spherocytosis, which refers to a group of heterogeneous inherited
anemias that have spherical-shaped erythrocytes, and hereditary elliptocytosis, which is a
heterogeneous group of diseases that cause elongation of the erythrocyte (Perotta et al, 2008;
Vaya et al, 2014).
Erythrocyte integral membrane components also include many GTP binding proteins.
These proteins have been shown to bind to Rho GTPases. Rho GTPases belong to the Ras
superfamily of small G proteins that function as molecular switches in actin cytoskeleton, cell
polarity, cell motility, and cell cycle (Kalfa et al, 2014). The main subfamilies of Rho GTPases
are Rho, Rac, and Cdc42. Rho GTPases are important for assembly of cell-cell junctions in

5
epithelial cells and tumor invasion. The three main types of Rho are RhoA, RhoB, and RhoC
(Kalfa et al, 2014). RhoA stimulates assembly of contractile actomyosin filaments and is critical
for tumor invasion in breast carcinoma. Previous studies also show how cytosolic RhoA binds to
the cytoplasmic side of erythrocyte membranes, and this interaction regulates actin filaments and
actomyosin complexes of the contractile ring (Boukharov et al, 1998). This interaction of RhoA
with the erythrocyte membrane has an effect on the erythroid spectrin on the RBCs. Many
signaling proteins and exchanges factors can mediate this interaction; in particular, myosin-
interacting guanine exchange factor (MyoGEF) activates RhoA and other Rho GTPases.
Decreased levels of MyoGEF decrease the activation of RhoA and RhoC (Wu et al, 2009).
MyoGEF also plays an important role in regulating cytokinesis and cell migration. Like
other GEFs, MyoGEF contains a Dbl homology (DH) domain and pleckstrin homology (PH)
domain. The DH domain is involved in catalyzing the exchange of GDP/GTP, while the PH
domain is involved in intracellular pathways such as ß-adrenergic and Phospholipase C (Wu et
al, 2014: Rebecchi et al, 1998). This important signaling molecule is highly expressed in
invasive breast cancer cells. Previous studies Dr. Wei’s lab has already MyoGEF interacting
with the C-terminal of Spta1, as shown in Figure 2.
Figure 2: A map of Spta1, consisting of 2415 amino acid residues and its specific domains, and its interaction with MyoGEF. Previous studies determine MyoGEF interacting with the C

6
terminal half of Spta1, specifically, between amino acid residues 2040-2415.
The aim of this study is to characterize the interaction between Spta1 and MyoGEF, previously
identified through yeast 2-hybrid and in vitro pulldown assays. The data collected indicates the
C-terminal of Spta1, specifically the EF-hand, is important for the interaction with the C-
terminal half of MyoGEF.
Experimental Procedures
High-Fidelity Polymerase Chain Reaction (Hi-Fi PCR) System
The Hi-Fi PCR system was used to amplify cDNA fragments corresponding to the C-
terminal half of mSpta1, amino acid residues 2040-2256, as well as amino acid residues 2040-
2340. The following reaction mixture was used to amplify the desired fragments: 0.5µL
template DNA, 10µL 5x PCR HiFi buffer, 1µL appropriate primer, 1µL appropriate reverse
primer, 2µL 10mM dNTP mix, 0.8µL HiFi enzyme, and 34.7µL ddH2O. The samples were then
placed in PCR system at the following conditions: denatured at 94˚C for 30 seconds, annealed at
48˚C for 30 seconds, and elongated at 68˚C for 1 minute. This process continued for 30 cycles.
The samples were then purified via PCR purification protocol (Qiagen).
TOPO Cloning Reaction for PCR Products and Transformation into Competent Cell
Each purified PCR product was then cloned into pCR8-GW-TOPO entry vector. The
following reaction mixture was used: 3µL of PCR product, 1µL salt solution, 1µL ddH2O, and
1µL pCR8-GW-TOPO vector. This reaction mixture incubated for 20 minutes at room
temperature and then placed on ice. 2µL of reaction mixture was added to 25µL of TOPO One
shot competent cells, and incubated on ice for 30 min. The samples were then heat shocked at
42˚C for 40sec, and then placed on ice for 2 min. 250µL of SOC medium was added to each

7
sample, and then placed in shaker at 225rpm for 1 hour at 37˚C. The samples were then plated
on Luria-Bertani (LB)-spectinomycin agar plates. These plates were incubated overnight at
37˚C. The cultures were then harvested and grown in 4mL of LB broth with 4µL of
spectinomycin. These liquid bacteria cultures shook overnight 250rpm at 37˚C. Plasmids were
then extracted by QIAprep Miniprep protocol (Qiagen). The fragments’ length were confirmed
by EcoRI enzyme digestion and then the fragments’ orientations were confirmed by EcoRV
enzyme digestion. The fragments were also sequenced to confirm length and orientation.
LR Recombination Reaction
Once the plasmids were confirmed, they were then cloned into the destination vector
pACT-2.2-gtwy. The following reaction mixture was used: 3.5µL plasmid, 0.5µL pACT-2.2-
gtwy vector, and 1µL clonase II mix. The reaction mix was then incubated at room temperature
for 2 hours. Then 0.5µL of proteinase k was added and subsequently incubated at 37˚C for
10min and then placed on ice. Reaction mixes were then transformed into competent cells as
described previously. The only difference is that the destination vector is ampicillin-resistant.
Positive cloning was confirmed through EcoRI and XhoI enzyme digestions.
Yeast 2-Hybrid- Preparation of Competent AH109 Yeast Cells
A yeast 2-hybrid protocol (Clontech) was used to determine if the clones purified from
the PCR system interact with C-terminal half MyoGEF. The yeast strain AH109 was used in this
protocol. After AH109 was streaked on a YPDA agar plate and incubated until colonies
appeared, one colony was inoculated into 3mL of YPDA medium. This was incubated in the
shaker overnight at 30˚C. 10µL of the culture was transferred to 100mL of YPDA and incubated
at 30˚C shaking at 230rpm for 16-20hr. The culture was then centrifuged for 5min and the cell
pellet was resuspended in 200mL of YPDA. The culture was incubated at 30˚C until the optical

8
density (OD) was 0.4-0.5. Then 50mL of the cells were transferred to a separate tube, then
centrifuged and resuspended in 1.5mL of 1.1X TE/LiAc. The solution was then centrifuged for
15sec and the cell pellet was resuspended in 300µL of 1.1X TE/LiAc.
Yeast 2-Hybrid- Transformation of Competent AH109 Yeast Cells
The transformation mix was then set up with the following mixture: 5µL of bait plasmid,
5µL of prey plasmid, 5µL Herring Testes Carrier DNA, 50µL Competent Cells, 0.5mL
PEG/LiAc. The mixtures were mixed by vortexing and then incubated at 30˚C for 30min; the
cells were mixed every 10min. 20µL of DMSO was added and then placed in 42˚C water bath
for 15min; vortexed every 5min. The mixtures were centrifuged for 15sec to pellet the yeast
cells. The supernatant was removed and pellet was resuspended in 1mL of YPD. The solution
was then centrifuged for 15sec, the supernatant was discarded, and was then resuspended in
400µL of 1x TE buffer.
Yeast 2-Hybrid: Plating the Transformation Mixture
200µL of the transformation mixture was plated on each of the SD selected medium:
(SD/-Trp-Leu) and (SD/-Trp-Leu/-Ade/-His). The plates were incubated at 30˚C upside down
until colonies were formed.
Yeast 2-Hybrid: Restreak colonies and confirmation of bait-prey interaction by colony-lift filter
assay
After noticeable colonies grew on the (SD/-Trp-Leu/-Ade/-His), the colonies were re-
streaked on another (SD/-Trp-Leu/-Ade/-His) plate and incubated at 30˚C until colonies are
formed. The colony-lift filter assay was performed (Clontech). A filter paper was presoaked in
2.5mL of Z buffer/X-gal solution in a clean 100-mm plate. Then a dry filter is placed over the
surface of the plate of yeast patches, and the filter was rubbed with forceps to ensure the yeast

9
colonies stuck to the filter. This filter was then submerged in liquid nitrogen for about 10sec.
The filter paper thawed at room temperature and, with colony side up, placed on presoaked filter
and incubated at room temperature until signs of blue patches or dry filter paper.
In vitro Pulldown Assay: TnT Transcription/Translation systems
In vitro pulldown assay was done to determine protein interactions of Spta1 and
MyoGEF. In this part of the In vitro pulldown assay, the positive clones have been previously
isolated, and the protein for each clone is expressed through this process. The reaction mixture
consists of the following: 20µL TnT Quick Master Mix (Sp6), 0.5µL Methionine, 1µL plasmid,
and 3.5µL Nuclease-Free Water. The components were spun down and then incubated for
90min at 30˚C. During this incubation period, the GST- beads were washed with 500µL BC100
binding buffer.
In vitro Pulldown Assay
The following reaction mixture was made: 10µL of in vitro translated proteins, 20µL
GST-beads, 150µL 2x BC buffer, 120µL ddH2O, and 0.5µL 1M DTT. These contents were
incubated overnight at 4˚C. The beads were washed with BC100 buffer 3 times and then
resuspended in 1xSDS load buffer to elute the bound proteins.
In vitro Pulldown Assay: Western Blot
The pulldown samples were separated on 8% SDS-PAGE gel at 200volts for 1hr. The
gel was then transferred to nitrocellulose membrane. Western Blot was run at 15volts for
40mins. The membrane was then covered with Ponceaus in order to check if there were proteins
attached to the membrane. Then water and TBST buffer were used to wash membrane. Once
the membrane is cleared of Ponceaus, blocking buffer was added to membrane in container for
30min. Then the primary antibody was added: GFP(1:1000, R). With the primary antibody, the

10
membrane soaked overnight at 4˚C. After three washes, the membrane was incubated with the
secondary antibody, anti-rabbit (1:5000), for 1hr at 24˚C.
Results
C-terminal half of MyoGEF interacts with Spta1 2040-2340 only
This round of yeast 2-hybrid was performed in order to see which of cloned Spta1
plasmids interact with MyoGEF, specifically whether the plasmids interact with the N-terminal
or the C-terminal of MyoGEF. The first yeast 2-hybrid test that was performed revealed Spta1
amino acid residues 2040-2340 interacted with the C-terminal half of MyoGEF, as shown below
in Figure 3. The results also indicated there was no interaction between the N-terminal half of
MyoGEF with Spta1 2040-2340. There was also no interaction between Spta1 between 2040-
2256 with both the N-terminal and C-terminal half of MyoGEF.
Figure 3. (left) This is the result of the colony lift filter assay. The blue yeast colonies (as shown in rectangle) are the amino acid residues 2040-2340 of Spta1 interacting with C-terminal of MyoGEF. The blue colonies indicate a positive interaction between the two proteins, whereas the lack of blue indicates no interaction between the two proteins.

11
GST- Pulldown Assay indicates no interaction with Spta1 2040-2256 or 2040-2340
GST-pulldown assay was performed in order to confirm the yeast 2-hybrid results, shown
in Figure 4. This assay tested the same Spta1 fragments as the first round of yeast 2-hybrid
results. The only difference is that this assay wanted to confirm the interaction through in vitro
binding between the Spta1 fragments and MyoGEF amino acid residues 501-790, the C-terminal
of MyoGEF. The N-terminal of MyoGEF was not tested through in vitro pulldown assay. The
results in Figure 4 show that there was no interaction between Spta1 2040-2256, Spta1 2040-
2340 with MyoGEF 501-790.
Figure 4. (left) Western Blot results from GST- pulldown assay. Lanes 1-3 are the results of Spta1 2040-2256. Lanes 4-6 are the results of Spta1 2040-2340. Lanes 3 and 6 show no interaction between MyoGEF 501-790 and both Spta1 fragments.

12
Yeast 2-Hybrid results show Spta1 may interact with MyoGEF between 501-790
The second round of yeast 2-hybrid testing was performed in order to determine which
part of the MyoGEF C-terminal interacts with Spta1 C-terminal. Previous studies in Dr. Wei’s lab
have shown interaction with MyoGEF 501-790 and C-terminal of Spta1. Therefore this yeast 2-
hybrid testing determined the possible interaction with C-terminal half of Spta1 and truncated
fragments of MyoGEF’s C-terminal. Table 1 shows that MyoGEF 501-790 interacted with the Spta1
C-terminal, but there was no interaction between Spta1 C-terminal and MyoGEF 501-752 or 501-
590. These results suggest that the region between 752-790 is the region where the interaction
occurs.
Bait Plasmids Prey X-Gal filter assay
GAL4-BD-MyoGEF 501-790 GAL4-AD-Spta1 Positive
GAL4-BD-MyoGEF 501-590 GAL4-AD-Spta1 Negative
GAL4-BD-MyoGEF 501-752 GAL4-AD-Spta1 Negative
Table 1. Summary of yeast 2-hybrid results shown above. The results show that there is an interaction between MyoGEF 501-790 and the C-terminal of Spta1 and no interaction between MyoGEF 501-752 and C-terminal of Spta1, suggesting that the region of MyoGEF 752-790 is important.
Discussion
In this study, we have shown that the C-terminal half of MyoGEF is important in the
interaction with the C-terminal of Spta1. Specifically, it seems that MyoGEF’s amino acid
residues 752-790 are important with the interaction of Spta1’s amino acid residues 2256-2340.
These results from the yeast 2-hybrid seem to indicate that the EF hand, a calcium-binding motif
shown in Figure 2, is important in the MyoGEF-Spta1 interaction. Our findings are consistent

13
with previous data collected, because Dr. Wei’s laboratory showed that the Spta1’s amino acid
residues 2040-2415 are important in the interaction with MyoGEF’s amino acid residues 501-
790. But there were some contradictory results, as shown with the GST- In vitro Pulldown
Assay because there was no interaction with Spta1 2040-2256 and Spta1 2040-2340 with
MyoGEF 501-590. One possible explanation for this lack of interaction can be due to
posttranslational modifications, which can occur during the process of yeast 2-hybrid protocol,
but will not occur during in vitro binding.
Further experimentation is necessary in order to determine if the EF hand is actually
important in this interaction. This determination can be accomplished through further cloning of
Spta1’s C-terminal. Along with characterizing the interaction of MyoGEF-Spta1, the effects of
this interaction will be analyzed. It is not well known how exactly this interaction will affect the
formation of erythrocytes. The effects of decreased levels of MyoGEF in erythrocytes is
unknown, it is only known that MyoGEF activates RhoA, which is present in erythrocytes
cytoskeleton. Further, this research is the important beginning to see how MyoGEF-Spta1
interaction can affect the formation of erythrocytes and the maintenance of erythrocyte
cytoskeleton as it travels throughout the circulatory system.

14
Works Cited
Boukharov, Andrey A., and Carl M. Cohen. "Guanine Nucleotide-dependent Translocation of
RhoA from Cytosol to High Affinity Membrane Binding Sites in Human Erythrocytes."
Jstor.org. Joural of Biochemistry, 1998. Web.
Hall, A., and C. D. Nobes. "Rho GTPases: Molecular Switches That Control the Organization
and Dynamics of the Actin Cytoskeleton." Philosophical Transactions of the Royal
Society B: Biological Sciences 355.1399 (2000): 965-70. Web.
Heasman, Sarah J., and Anne J. Ridley. "Result Filters." National Center for Biotechnology
Information. U.S. National Library of Medicine, Sept. 2008. Web. 27 Apr. 2016.
Kalfa, Theodosia A., and Yi Zheng. "Rho GTPases in Erythroid Maturation." ResearchGate.
NIH, May 2014. Web. 27 Apr. 2016.
Korsgren, C., and S. E. Lux. "The Carboxyterminal EF Domain of Erythroid -spectrin Is
Necessary for Optimal Spectrin-actin Binding." Blood 116.14 (2010): 2600-607. Google
Scholar. Blood Journal, 22 Apr. 2016. Web.
Machnicka, B., R. Grochowalska, D. M. Bogusławska, A. F. Sikorski, and M. C. Lecomte.
"Spectrin-based Skeleton as an Actor in Cell Signaling." Cellular and Molecular Life
Sciences. SP Birkhäuser Verlag Basel, 23 May 2011. Web. 27 Apr. 2016.
Perrotta, Silverio, Patrick G. Gallagher, and Narla Mohandas. "Hereditary Spherocytosis."
Lancet 372 (2008): 1411-426. Print.
Rebecchi, M. J., and S. Scarlata. "PLECKSTRIN HOMOLOGY DOMAINS: A Common Fold
with Diverse Functions." Annual Review of Biophysics and Biomolecular Structure Annu.
Rev. Biophys. Biomol. Struct. 27.1 (1998): 503-28. Web.

15
Salomao, Marcela, Xiuli An, Xinhua Guo, Walter B. Gratzer, Narla Mohandas, and Anthony J.
Baines. "Mammalian Alpha1-Spectrin Is a Neofunctionalized Polypeptide Adapted to
Small Highly Deformable Erythrocytes." Jstor.org. PNAS, 17 Jan. 2006. Web.
Vaya, Ampara, Marta Suescun, Amparo Pardo, and Oscar Fuster. "Erythrocyte Deformability
and Hereditary Elliptocytosis." Jsor.org. Clinical Hemorheology and Microcirculation,
2014. Web.
Wu, D., M. Asiedu, and Q. Wei. "Result Filters." National Center for Biotechnology
Information. U.S. National Library of M3edicine, 2009. Web. 27 Apr. 2016.
Wu, Di, Meng Jiao, Shicheng Zu, Christopher C. Sollecito, Kevin Jimenez-Cowell, Alexander J.
Mold, Ryan M. Kennedy, and Qize Wei. "Intramolecular Interactions between the Dbl
Homology (DH) Domain and the Carboxyl-terminal Region of Myosin II-interacting
Guanine Nucleotide Exchange Factor (MyoGEF) Act as an Autoinhibitory Mechanism
for Regulation of MyoGEF Functions." Jstor.org. Journal of Biological Chemistry, 26
Aug. 2014. Web.