selection of galactose-fermenting streptococcus ... · exhausted, and catabolite repression was...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, JUIY 1984, p. 186-1910099-2240/84/070186-06$02.00/0Copyright C) 1984, American Society for Microbiology
Selection of Galactose-Fermenting Streptococcus thermophilus inLactose-Limited Chemostat Cultures
TERENCE D. THOMAS- AND VAUGHAN L. CROW
New1 Zealainld Dairy Researchl Inistititte, Palmerston Nolrtli, Nell, Zealan(d
Received 14 February 1984/Accepted 24 April 1984
Stock cultures of Streptococcus thermophilus are essentially galactose negative (Gal-). Although bothgalactose I-phosphate uridyl transferase and uridine-5-diphospho-glucose 4-epimerase are present, suggestingthat the genes for the Leloir pathway exist, cells cannot induce high levels of galactokinase. Therefore, galactoseis largely excreted when cultures are grown on lactose, and most strains cannot be readily adapted to grow onfree galactose. Gal- cultures were grown in a chemostat under lactose limitation in which high concentrationsof residual galactose were present. Under this selection pressure, Gal' organisms eventually took over theculture with all four strains examined. Gal' cells had induced galactokinase, and three of the four strains grewon free galactose with doubling times of 40 to 50 min. When Gal' organisms were grown on lactose in batchculture, the galactose moiety was only partially utilized while lactose was still present. As lactose wasexhausted, and catabolite repression was lifted, the Leloir pathway enzymes (especially galactokinase) wereinduced and the residual galactose fermented. Neither phospho-o-galactosidase activity nor the enzymes of theD-tagatose 6-phosphate pathway were detected in S. thiermophilus. In contrast to Streptococcus cremoris andStreptococcus lactis, fermentation was homolactic with galactose in batch cultures and with lactose limitation inthe chemostat. When mixed Gal+-Gal- cultures were repeatedly transferred in milk, the Gal- cells became thedominant cell type. The Gal- phenotype of stock cultures probably reflects their prolonged maintenance inmilk.
Streptococci play a vital role in many commercial milkfermentations. Selected strains are deliberately added as
"starters," and their primary role is to ferment lactose tolactic acid. Lactose fermentation by the mesophilic lalcticstreptococci (S. criemori.s and S. lactis) has been extensivelystudied. In these organisms. the phosphoenolpyruvate-de-pendent phosphotransferase system for lactose uptake andphospho-p-galactosidase are prerequisites for rapid homo-lactic lactose fermentation (2. 13). Both glucose and galac-tose 6-phosphate moieties are metabolized simultaneously.via the glycolytic and D-tagatose 6-phosphate pathways,respectively (7). Lactic streptococci also grow on free galac-tose, using the Leloir or the D-tagatose 6-phosphate pathwayor both (17).
Streptococcus thelwrmiophilius is used in commercial milkfermentations in which the temperatures reached duringproduct manufacture are relatively high (up to -55°C). Thisorganism is normally used as part of a mixed starter culture.the other component being a Lactobacillis sp. (e.g., L.bulgaricus for yogurt, L. helvleticits for Swiss-type cheese).Compared with the mesophilic starter streptococci, relative-ly little is known about sugar fermentation by S. tlieri-toplhi-lias. Lactose and sucrose are fermented more readily thantheir component monosaccharides (15. 18. 22). Lactose istaken up as the free sugar and split with 3-galactosidase.When S. the)iinoplilius strains grow in the presence of excesslactose, only the glucose portion of the lactose molecule isfermented (9, 18). Thus. galactose accumulates in fermentedmilk products when this organism is used. In yogurt, inwhich high acid concentrations limit the fermentation, freegalactose remains (12). In contrast, the free galactose pro-
duced in the early stages of Swiss cheese manufacture islater fermented by L. hlelv'eticis (8, 20).When grown in batch culture with limited lactose, some S.
* Corresponding author.
186
th2ernHiophlilius strains utilize the galactose moiety. usuallyincompletely (18). In similar cultures, the maximum freegalactose concentration attained has been only slightly lessthan the decrease in lactose concentration, indicating thatsparing galactose utilization occurs while lactose is stillpresent and continues after lactose exhaustion (15). Thissuggests that enzymes for galactose metabolism are present,but at low activity. Attempts have been made to adapt S.thleir-,nophilus strains to grow on galactose in batch culture.but few details have been reported (6. 11. 18). In preliminaryexperiments, we found that some S. tielwiniopliilus strainsgrew (after 2 to 5 days of incubation at 370C) in brothmedium containing galactose. However, this ability to growon galactose was lost on subculture. The aim of the presentstudy was to obtain galactose-utilizing organisms by usingthe highly selective environments provided by chemostatculture (3, 5).
MATERIALS AND METHODS
Organisms and growth media. S. tiler,nophiliis stock cul-tures were originally obtained from the following sources:CNRZ strain 368 from the National Agricultural ResearchInstitute, Jouy-en-Josas, France: NCDO strain 821 from theNational Institute for Research in Dairying. Reading, En-gland; strain TS2 from CSIRO Dairy Research Laboratory,Melbourne, Australia; strain MC from G. W. Reinbold,Iowa State University, Ames. Stock cultures were stored at-75°C in J8 broth medium. Batch cultures were grown in J8broth which had an initial pH of 6.8 and consisted of (perliter): lactose, 5 g, unless otherwise specified: beef extract(BBL Microbiology Systems, Cockeysville, Md.). 2 g; poly-peptone (BBL), 5 g: phytone peptone (BBL). 2 g: yeastextract (BBL) 2 g8 Na2HPO4, 5.5 g: KH2PO4. 5 g: ascorbicacid, 0.5 g; MgCl1 * 6H,O, 0.2 g. The medium was auto-claved in two parts, and the lactose-MgCl. part was addedaseptically to the bulk medium. For chemostat cultures, thismedium was modified as follows (per liter): polypeptone
Vol. 48. No. 1
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from

GALACTOSE-FERMENTING S. THERMOPHILUS 187
(BBL). 2 g: Na,HPO4, 1.7 g: KH2PO4. 1.7 g. Milk wasprepared by reconstituting antibiotic-free nonfat milk pow-der (10 g of powder per 100 ml of water).Growth conditions. Experimental batch cultures were
grown at 42°C under static conditions.Chemostat cultures were grown in a vessel with a 500-ml
working capacity (Series III Fermentor. LH Engineering.Stoke Poges, England). The temperature was maintained at42°C. and the pH was controlled at 6.45 ± 0.05 by automaticaddition of 0.1 M NaOH. Anaerobic conditions were main-tained by passing a 95% N,-5% CO, mixture through theheadspace of the stirred culture. which was grown initially inthe baitch mode. After batch growth had stopped. mediumwas pumped into the fermentor at the appropriate dilutionrate (D. proportion of culture volume replaced per hour).and the culture was allowed to reach equilibrium (i.e.. wasgrown for at least 10 mean generation times at each D) beforebeing sampled.Enzvme assays. Cells in the appropriate growth condition
were harvested. washed in 20 mM phosphate buffer (pH 6.5,containing 50 mM NaCl. 10 mM MgCI,. and 1 mM dithioer-ythritol). and disrupted in the same buffer by shaking 5 ml ofsuspension (-6 mg [dry weight] of bacteria per ml) for 2 minat 0 to 5°C with 3 ml of glass beads (75 to 150 p.m diameter.Sigma type 1-w) in a Mickle disintegrator. Debris wasremoved by centrifugation at 35,000 x g for 5 min. Cell-freeextracts (-2 mg of protein per ml) were stored on ice, andenzyme assays were completed within 2 h from preparationof extracts. All enzymes assayed were stable (>959%) for atleast 4 h. except for the glucokinase activity from strain MCGal'. in which up to 40% of the activity was lost within 1 hof cell extract preparation.
All enzyme assays were carried out at 25°C. Uridine-5'-diphospho-(UDP)-glucose 4-epimerase (EC 5.1.3.2) was as-sayed by a two-step procedure. In step 1. the bacterialextract was incubated for 0. 5. and 10 min in an assay system(500 [1I) containing 1 mM UDP-galactose in 100 mM triethan-olamine hydrochloride buffer (pH 7.5). The reaction wasstopped by heating in a boiling water bath for 2 min. In step2. the amount of UDP-glucose formed was determined byadding 150 pl. of sample (or dilution) to 400 pL. of 0.5 Mglycine-NaOH buffer (pH 8.6) and 50 pl. of 0.1 M NAD'.The reaction was started by the addition of UDP-glucosedehydrogenase (20 p.1. 0.05 U) and was followed at 340 nmwith a Gilford model 250 spectrophotometer. Both UDP-glucose:ot-D-galactose 1-phosphate uridyltransferase (EC2.7.7.12, galactose-lP uridyltransferase) and cx-D-glucose1,6-diphosphate:ot-D glucose 1-phosphate phosphotransfer-ase (EC 2.7.5.1, phosphoglucomutase) assays shared thefollowing standard reaction mixture: 100 mM triethanol-amine hydrochloride buffer (pH 7.5), 10 mM MgCl2, 1 mMNADP+, 0.25 mM glucose 1,6-diphosphate, glucose 6-phos-phate dehydrogenase (5 U), and bacterial extract. Thegalactose-1P uridyltransferase assay system contained anadditional coupling enzyme (phosphoglucomutase, 3 U) andthe two substrates (UDP-glucose and galactose 1-phos-phate), each at a final concentration of 1 mM. For the assayof phosphoglucomutase activity, 5 mM glucose 1-phosphatewas added to the standard reaction mixture.
Galactokinase (EC 2.7.1.6) and glucokinase (EC 2.7.1.2)were assayed spectrophotometrically by incubation of cellextract with carbohydrate and ATP and coupling ADPproduction to NADH oxidation via pyruvate kinase andlactate dehydrogenase. The reaction mixture contained (in a1-ml volume): 100 mM triethanolamine hydrochloride buffer(pH 7.8), 5 mM MgCl2, 2 mM ATP, 2 mM phosphoenolpy-
ruvate. 0.25 mM NADH. nonlimiting amounts of pyruvatekinase (3 U). and lactate dehydrogenase (4 U). The appropri-ate carbohydrate (final concentration. 10 mM) was addedlast to allow correction for NADH oxidase and myokinaseact ivitv.The enzymes of the D-tagatose 6-phosphate pathway
(galactose 6-phosphate isomerase. tagatose 6-phosphate ki-nase. and D-tagatose 1.6-diphosphate aldolase) and phospho-,3-galactosidase activities were assayed as described byCrow et al. (1). Lactate dehydrogenase was assayed asdescribed by Thomas and Crow (15). except that the routineassay temperature was 25°C rather than 42°C.
Sugar utilization rate. The rate of sugar utilization bynongrowing cells \vas measured with a pH-stat apparatus(A. M. Fordyce. V. L. Crow. and T. D. Thomas, submittedfor publication). Culture samples (40 ml) were removed fromthe chemostat and centrifuged. The cells were washed twiceand suspended in 10 ml of 20 mM phosphate buffer (pH 6.45,containing 50 mM NaCI and 10 mM MgCI.) at -2 mg (dryweight) of bacteria per ml. Portions (2 ml) of this suspensionwere placed in a Radiometer pH-stat apparatus (Fordyce etal., submitted for publication). The stirred suspension wasadjusted to pH 6.50 and maintained at 420C under anatmosphere of 0-free N,. Galactose (100 p.I1 1 M). glucose(100 p.1l 1 M), or lactose (100 p.1. 0.5 M) was added, and therate of alkali (0.10 M NaOH) addition for pH control at 6.50was recorded. From this rate. the rate of sugar utilizationwas calculated (Fordyce et al., submitted for publication).
Proportion of galactose-utilizing organisms. Culture dilu-tions containing 20 to 50 CFU were plated on J8 medium(containing 0.5% lactose and 1% agar) and incubated for 1day at 37°C in an atmosphere of H2-CO. All of the colonieson these plates (at least 50 per sample) were transferred to J8agar containing 0.5% galactose. After incubation for 2 daysat 37°C in an H.-CO. atmosphere. the proportion of galac-tose-utilizing organisms in the population was determined.Some stock cultures showed slight growth on J8 agar with orwithout 0.5% galactose (Gal-- cells). In contrast. galactose-utilizing organisms were clearly distinguishable (Gal cells).To examine the stability of galactose-utilizing cultures
maintained in lactose-excess media (see Table 3). cultureswere initiated in autoclaved reconstituted skim milk (10%solids) and in J8 broth (5% lactose) by inoculating with Gal'and Gal- cells in the ratio of 99:1. This was achieved byadding. to 10 ml of medium. 0.1 ml of culture grown in J8broth containing 0.5% galactose (Gal' cells) and 0.1 ml ofdiluted (10-2) stock culture grown in J8 broth containing 1%,rlactose (Gal- cells). Cultures were then incubated at 37°Cand transferred every 12 h (0.1% inoculum) into fresh media.The proportion of Gal' cells in the population was deter-mined as described above.Attempts to break up the streptococcal chains by blending
culture dilutions were unsuccessful. Blending (Ultra-TurraxJanke & Kunkel GmbH. Staufen. West Germany) actuallyresulted in a decrease in CFU per milliliter with most strains.apparently due to cell lysis. Therefore. the proportion ofGal' cells in the population will be overestimated in somecultures (the chains formed by the different strains containedan average of up to -5 cells in the 12-h cultures. as judgedfrom phase-contrast microscopy).
Other procedures. Samples taken for sugar analyses wereimmediately heated for 2 min at 100°C to inactivate any cell-free ,B-galactosidase (15). Sugar concentrations and fermen-tation products were determined by enzymatic analyses (15:Fordyce et al.. submitted for publication). Bacterial densitywas determined turbidimetrically (15). and bacterial dry
VOL. 48, 1984
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from
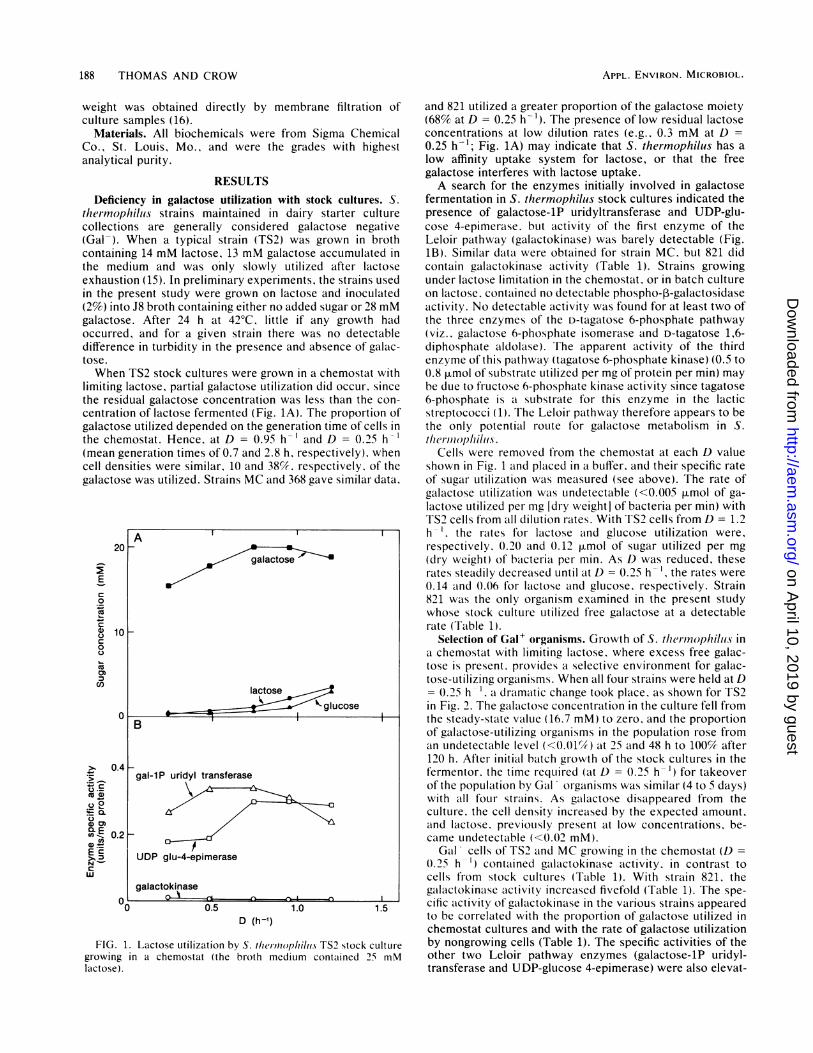
188 THOMAS AND CROW
weight was obtained directly by membrane filtration ofculture samples (16).
Materials. All biochemicals were from Sigma ChemicalCo., St. Louis, Mo., and were the grades with highestanalytical purity.
RESULTS
Deficiency in galactose utilization with stock cultures. S.therrnophilius strains maintained in dairy starter culturecollections are generally considered galactose negative(Gal-). When a typical strain (TS2) was grown in brothcontaining 14 mM lactose, 13 mM galactose accumulated in
the medium and was only slowly utilized after lactoseexhaustion (15). In preliminary experiments, the strains usedin the present study were grown on lactose and inoculated(2%) into J8 broth containing either no added sugar or 28 mMgalactose. After 24 h at 420C, little if any growth hadoccurred, and for a given strain there was no detectabledifference in turbidity in the presence and absence of galac-tose.When TS2 stock cultures were grown in a chemostat with
limiting lactose, partial galactose utilization did occur, sincethe residual galactose concentration was less than the con-
centration of lactose fermented (Fig. 1A). The proportion ofgalactose utilized depended on the generation time of cells in
the chemostat. Hence, at D = 0.95 h-' and D = 0.25 h-1(mean generation times of 0.7 and 2.8 h, respectively), whencell densities were similar, 10 and 38%. respectively, of thegalactose was utilized. Strains MC and 368 gave similar data,
E
0
0
0
Cu-
CD,
1
>, 0'
.-4-00)
co004~..L
-
.
0)
Ec>"N
-O 0.5 1.0
D (h-')
FIG. 1. Lactose utilization by S. tlier,0oplhiluii TS2 stock culturegrowing in a chemostat (the broth medium contained 25 mMlactose).
and 821 utilized a greater proportion of the galactose moiety(68% at D = 0.25 h-'). The presence of low residual lactoseconcentrations at low dilution rates (e.g.. 0.3 mM at D =
0.25 h-'; Fig. 1A) may indicate that S. thermophilus has alow affinity uptake system for lactose, or that the freegalactose interferes with lactose uptake.A search for the enzymes initially involved in galactose
fermentation in S. thermophilius stock cultures indicated thepresence of galactose-1P uridyltransferase and UDP-glu-cose 4-epimerase. but activity of the first enzyme of theLeloir pathway (galactokinase) was barely detectable (Fig.IB). Similar data were obtained for strain MC, but 821 didcontain galactokinase activity (Table 1). Strains growingunder lactose limitation in the chemostat, or in batch cultureon lactose. contaitned no detectable phospho-3-galactosidaseactivity. No detectable activity was found for at least two ofthe three enzymes of the D-tagatose 6-phosphate pathway(viz., galactose 6-phosphate isomerase and D-tagatose 1,6-diphosphate aldolase). The apparent activity of the thirdenzyme of this pathway (tagatose 6-phosphate kinase) (0.5 to0.8 [tmol of substrate utilized per mg of protein per min) maybe due to fructose 6-phosphate kinase activity since tagatose6-phosphate is a substrate for this enzyme in the lacticstreptococci (1). The Leloir pathway therefore appears to bethe only potential route for galactose metabolism in S.tlhernophiliis.
Cells were removed from the chemostat at each D valueshown in Fig. 1 and placed in a buffer, and their specific rateof sugar utilization was measured (see above). The rate ofgalactose utilization was undetectable (<0.005 p.mol of ga-lactose utilized per mg [dry weight] of bacteria per min) withTS2 cells from all dilution rates. With TS2 cells from D = 1.2h-'. the rates for lactose and glucose utilization were,respectively. 0.20 and 0.12 ILrmol of sugar utilized per mg(dry weight) of bacteria per min. As D was reduced, theserates steadily decreased until at D = 0.25 h the rates were0.14 and 0.06 for lactose and glucose, respectively. Strain821 was the only organism examined in the present studywhose stock culture utilized free galactose at a detectabler-ate (Table 1).
Selection of Gal' organisms. Growth of S. th'Ierinophilis ina chemostat with limiting lactose. where excess free galac-tose is present, provides a selective environment for galac-tose-utilizing organisms. When all four strains were held at D= 0.25 h -1 a dramatic change took place, as shown for TS2in Fig. 2. The galactose concentration in the culture fell fromthe steady-state value (16.7 mM) to zero, and the proportionof galactose-utilizing organisms in the population rose froman undetectable level (<0.01%) at 25 and 48 h to 100% after120 h. After initial batch growth of the stock cultures in thefermentor. the time required (at D = 0.25 h I) for takeoverof the population by Gal organisms was similar (4 to 5 days)with Call four strains. As galactose disappeared from theculture, the cell density increased by the expected amount,and lactose, previously present at low concentrations, be-came undetectable (<0.02 mM).
Gal' cells of TS2 and MC growing in the chemostat (D =
0.25 h i) contained galactokinase activity, in contrast tocells from stock cultures (Table 1). With strain 821, thegalactokinase activity increased fivefold (Table 1). The spe-cific activity of galactokinase in the various strains appearedto be correlated with the proportion of galactose utilized inchemostat cultures and with the rate of galactose utilizationby nongrowing cells (Table 1). The specific activities of theother two Leloir pathway enzymes (galactose-1P uridyl-transferase and UDP-glucose 4-epimerase) were also elevat-
A I
galactoe
10 ,
lactose_-glucose
Bo _
.4-gal-1 P uridyl transferase
2
UDP glu-4-epimerase
galactokinase
0-
APPL. ENVIRON. MICROBIOL.
2
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from

GALACTOSE-FERMENTING S. 7HERMOPHILUS 189
TABLE 1. Enzyme specific activities in S. thermtioplhilis strains growing under lactose limitation in a chemostat"
Enzyme sp act" Residual Galactosegalactose utilization
Strain Gal-IP in rate withGalactokinase uridyl- UDP glu-4- Phospho- Glucokinase chemostat nongrowing
transferase epimerase glucomutase (mM) cells'
TS2 0.01 0.26 0.19 0.68 1.63 16.8 <0.005TS2 Gal' 0.12 1.10 0.70 0.73 1.43 0.2 0.15MC ND" 0.34 1.12 0.56 0.80 15.0 <0.005MC Gal' 0.11 1.60 3.31 0.47 0.21 0.1 0.17821 0.05 0.68 0.43 0.79 1.99 8.6 0.07821 Gal+ 0.24 2.70 1.70 0.89 2.43 0.0 0.13
D 0.25 h growth medium contained 25 mM lactose.Specific activities are expressed as micromoles of substrate utilized per milligram of protein per minute.Micromoles of galactose utilized per milligram (dry weight) of bacteria per minute (see the text).
"ND, Not detectable (<0.005).
ed (three- to fivefold) in the Gal organisms. whereas thespecific activities of phosphoglucomutase and glucokinase.with the exception of glucokinase from MC cells, wereessentially unchanged (Table 1). Extracts from TS2 Gal'cells growing in the chemostat. where galactose was com-pletely utilized. contained no detectable activity for at leasttwo of the three enzymes of the D-tagatose 6-phosphatepathway or phospho-4-galactosidase.Growth of Gal' organisms in batch culture. (i) On lactose.
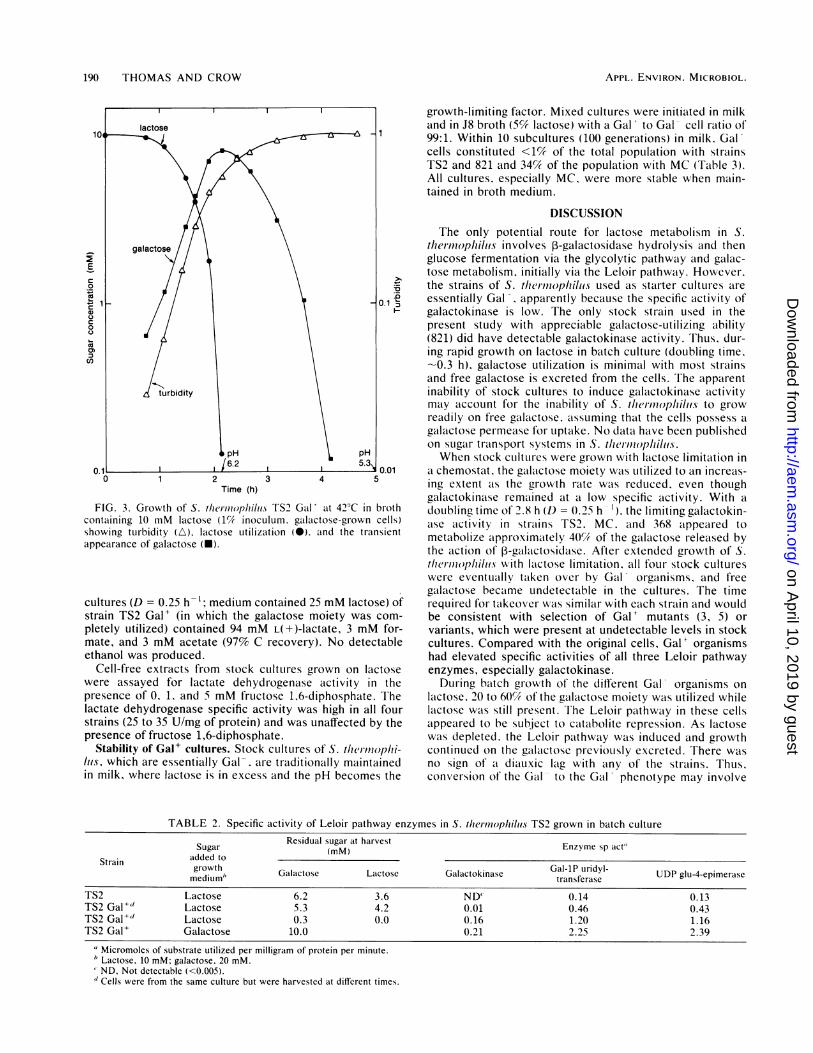
Growth of TS2 Gal in batch culture with limited lactose (10mM) is shown in Fig. 3. The growth rate was exponential(doubling time. --20 min) until the lactose concentration fellbelow - 3 mM. Galactose was released into the mediumand. with TS2. reached a maximum concentration of 8 mM.After lactose exhalustion. growth continued (on galactose) ata steadily declining rcate (Fig. 3). Galactose was undetectable(<0.02 mM) after 5 h. and no glucose WXas detected in theculture at any stage. The molar growth yield for strain TS2Gal growing on lactose was -66 g (drv weight) of bacteriaper mol of lactose utilized compared with an apparent valueof 36 for the stock culture. The other three Gal' organismsgave similalr results. except that with 821. the maximumconcentration of galactose reached was considerably lower(-4 mM). indicating that this organism had a greater capaci-ty for galactose utilization while lactose was still present.For each strain. the initial growth rates of Gal* and Galorganisms on lactose were similar. When grown in batchculture with excess lalctose (e.g.. in milk). the TS2 Gal'strain stopped growing due to the high acid concentrationproduced. and galactose utilization wvas minimal (data notshown).The specific activities of Leloir pathwvay enzymes in TS2
grown in batch cultures are shown in Table 2. In TS2 Gal'cells, the galactokinase activity was barely detectable whilelactose was still present (4.2 mM). In these cells. the specificactivities of galactose-lP uridyltransferase and UDP-glucose4-epimerase were threefold higher than in cells of the stockculture (Table 2). After lactose exhaustion and subsequentgrowth on the galactose moiety, the specific activities of allLeloir pathway enzymes, especially galactokinase, weremarkedly elevated. All three enzyme specific activities wereat their maximum in TS2 Gal' cells grown on galactose(Table 2).
(ii) On galactose or glucose. t'he Gal derivatives of strainsTS2. MC. 368, and 821 grew exponentially at 42'C in J8broth on galactose (20 mM) with doubling times of 51. 38.149. and 49 min. respectively. When grown on glucose (100mM) at 42°C. the Gal' organisms had doubling times similar
to those of the stock cultures. varying between 20 and 34 minfor the different strains.
Fermentation products. When group N streptococci (S.crenloris and S. l(atis) were grown in a chemostat tinderlactose limitation (14. 16). or in batch culture on galactose(13. 17). these "homolactic" organisms diverted fermenta-tion product from lcactate to formate. acetate. and ethainol. Itwas therefore of interest to examine the fermentation prod-ucts from S. tlwer-ImIophlillus growing under these conditions.During growth on either galactose ('0 mM) or lactose (10mM) in batch culture at 42'C. all Gal strains remainedessentially homolactic. with >95% of the fermented sugarbeing converted to L( (-lactate. Similarly. growth of eitherstock cultures or Gal' organisms in the chemostat (D = 0.25h-t) with lactose limitation resulted in homolactic fermenta-tion. Small quantities of formate and acetate were produced,in approximately equimolar amounts, by all strains growingat D values from 0.25 to 1.5 h-t. Typically, chemostat
E
0
COmI=
a)C;0n0
000
CCuCD
100
.75
cn
a)
C+-R
0 50 100 150Time (h)
FIG. 2. Selection of galactose-fer-menting S. tltErnophli/lu. TS2 inlatctose-limited chemostat cultures (I) = ).25 h the broth mediumcontained 25 mM lactose). Syrmbols indicate galactose concentration(rmM) (*) and percent Gal cells (C).
VOL. 48, 1984
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from
190 THOMAS AND CROW
E
0 /
0.10
c
0
c)
trbidity
pH pH
0.1 ~~~~~6.2 5.301
0 1 2 3 4 5Time (h)
FIG. 3. Growth of S. themonpitilus TS2 Gal' at 420C in brothcontaining 10 mM lactose (1% inoculum. galactose-grown cells)showing turbidity (A). lactose utilization (@). aind the transientappearance of galactose (*).
cultures (D = 0.25 h-'; medium contained 25 mM lactose) ofstrain TS2 Gal' (in which the galactose moiety was com-pletely utilized) contained 94 mM L(+)-lactate, 3 mM for-mate, and 3 mM acetate (97% C recovery). No detectableethanol was produced.
Cell-free extracts from stock cultures grown on lactosewere assayed for lactate dehydrogenase activity in thepresence of 0, 1, and 5 mM fructose 1,6-diphosphate. lThelactate dehydrogenase specific activity was high in all fourstrains (25 to 35 U/mg of protein) and was unaffected by thepresence of fructose 1,6-diphosphate.
Stability of Gal' cultures. Stock cultures of S. tlie-niiophli-lhis, which are essentially Gal-, are traditionally maintainedin milk, where lactose is in excess and the pH becomes the
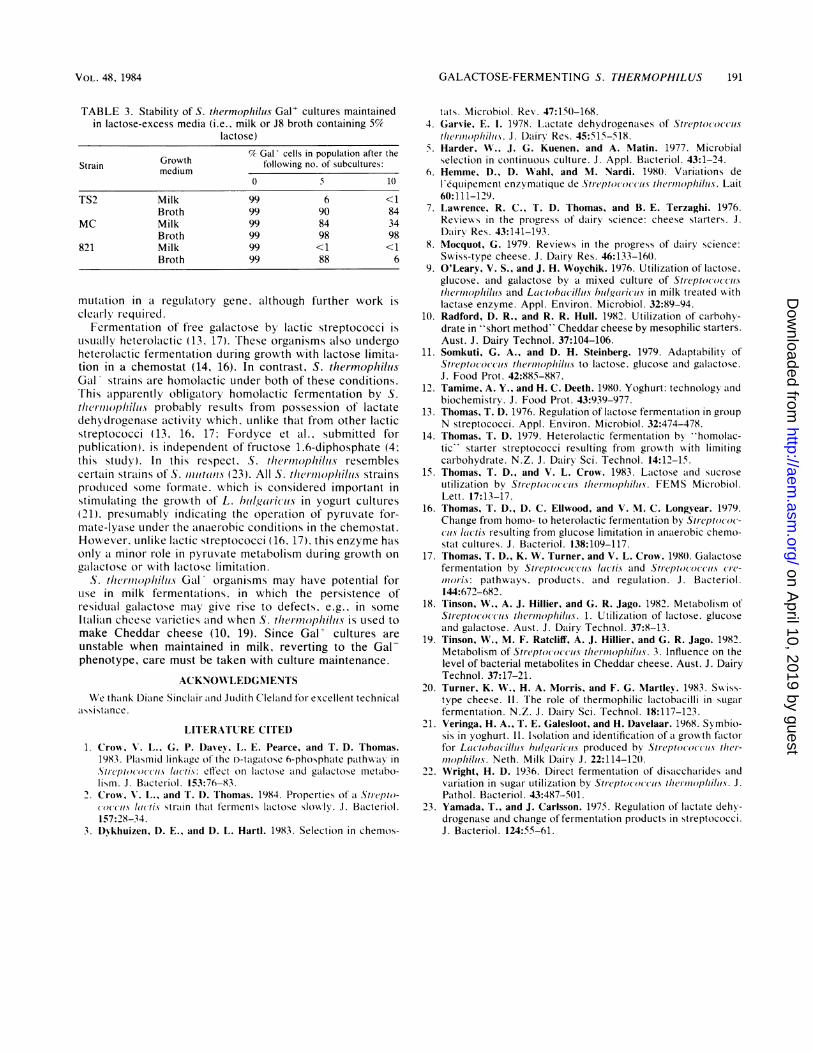
growth-limiting factor. Mixed cultures were initiated in milkand in J8 broth (5% lactose) with a Gal' to Gal cell ratio of99:1. Within 10 subcultures (100 generations) in milk. Gal'cells constituted <1% of the total population with strainsTS2 and 821 and 34% of the population with MC (Table 3).All cultures, especially MC, were more stable when main-tained in broth medium.
DISCUSSIONThe only potential route for lactose metabolism in S.
thelriopliillis involves 3-galactosidase hydrolysis and thenglucose fermentation via the glycolytic pathway and galac-tose metabolism, initially via the Leloir pathway. However.the strains of S. tiewrItnopiliius used as starter cultures areessentially Gal -. apparently because the specific activity ofgalactokinase is low. The only stock strain used in thepresent study with appreciable galactose-utilizing ability(821) did have detectable galactokinase activity. Thus. dur-ing rapid growth on lactose in batch culture (doubling time.-0.3 h), galactose utilization is minimal with most strainsand free galactose is excreted from the cells. The apparentinability of stock cultures to induce galactokinase activitymay account for the inability of S. tlhermnophlilius to growreadily on free galactose. aSSuming thatt the cells possess a
galactose permease for uptake. No data have been publishedon Sugar transport systems in S. thlermophlilits.When stock cultul-es were grown with lactose limitation in
a chemostat. the galactose moiety was utilized to an increas-ing extent as the growth rate was reduced, even thoughgalactokinase remained at a low specific activity. With adoubling time of 2.8 h (D = 0.25 h '). the limiting galactokin-ase activitv in stralins TS2. MC. and 368 appeared tometabolize approximiately 40% of the galactose released bythe action of 3-galactosidase. After extended growth of S.thlriniophiliis with lactose limitation, all four stock cultureswere eventually taken over by Gal' organisms. and freegalactose became undetectable in the cultures. The timerequired for takeover w\as similar with each strain and wouldbe consistent with selection of Gal' mutants (3, 5) orvariants, which were present at undetectable levels in stockcultures. Compared with the original cells, Gal' organismshad elevated specific activities of all three Leloir pathwayenzymes, especially galactokinase.
During batch growth of the different Gal organisms on
lactose. 20 to 60c of'the galactose moiety was utilized whilelactose was still present. The Leloir pathway in these cellsappeared to be sub'ject to catabolite repression. As lactosewas depleted. the ILeloir pathway was induced and growthcontiniled on the galactose previously excreted. There wasno sign of a diaIuxic latg with any of the str-ains. Thus,conversion of the Gal- to the Gal' phenotype may involve
TABLE 2. Specific activity of Leloir pathway enzymes in S. thedIoplilis TS2 grown in batch culture
Sugar Residual sugar at harvest Enzyme sp act"added to (MM)
Strain growth GtLcoelt Gal-IP uridyl-UD 1 4
medium" GtnlfctoseLactose Galactokinase transferae UDP glu-4-epimeraseTS2 Lactose 6.2 3.6 N D' 0.14 0.13TS2 Gal+" Lactose 5.3 4.2 0.01 0.46 0.43TS2 Gal+"' Lactose 0.3 0.0 0.16 1.20 1.16TS2 Gal+ Galactose 10.0 0.21 2.25 2.39
Micromoles of substrate utilized per milligram of protein per minute.Lactose, 10 mM; galactose. 20 mM.ND, Not detectable (<0.005).
"Cells were from the same culture but were harvested at different times.
APPL. ENVIRON. MICROBIOL.
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from
GALACTOSE-FERMENTING S. THERMOPHILUS 191
TABLE 3. Stability of S. thermoplhiluis Gal' cultures maintainedin lactose-excess media (i.e., milk or J8 broth containing 55%
lactose)
Growth % Gal cells in population after theStrain medium following no. of subcultures:
0 5 10
TS2 Milk 99 6 <1Broth 99 90 84
MC Milk 99 84 34Broth 99 98 98
821 Milk 99 <1 <1Broth 99 88 6
mutation in ac regulatory gene. although further work is
clearly required.Fermentation of free galactose by lactic streptococci is
usually heterolactic (13. 17). TFhese organisms also undergoheterolactic fermentation during growth with lactose limita-tion in a chemostat (14, 16). In contrast, S. thermophiliusGal strains are homolactic under both of these conditions.This apparently obligatory homolactic fermentation by S.thermuophilits probably results from possession of lactatedehydrogenase activity which. unlike that from other lacticstreptococci (13. 16. 17: Fordyce et al.. submitted forpuLblication). is independent of fructose 1.6-diphosphate (4:this study). In this respect. S. thermophilus resemblescertain strains of.S. inumnts (23). All S. thermuoplhilus strainsproduced some formate. which is considered important in
stimulakting the growth of L. hldguri(cius in yogurt cultures(21). presumably indicating the operation of pyruvate for-
mate-lyalse tinder the anaerobic conditions in the chemostat.However. unlike lactic streptococci (16. 17). this enzyme hasonly a minor role in pyruvate metabolism during growth on
galactose or with lactose limitation.S. therinophiluts Gal' organisms may have potential for
use in milk fermentations, in which the persistence ofresidual gal-actose may give rise to defects. e.g.. in some
Italiln cheese varieties and when S. thermophilius is used tomake Cheddar cheese (10, 19). Since Gal' cultures are
unstable when maintained in milk, reverting to the Gal-phenotype, care must be taken with culture maintenance.
ACKNOWN'LEDGNIENT'S
We thank Diane Sinclair and JLudith Cleland for excellent technical
aIssistance.LIIERATIURE CITED
1. Crow, V. L., G. P. I)avev. L. E. Pearce, and T. D. Thomas.1983. Plasmid linkage ot the D-tagatose 6-phosphate pathway in.Str(ptococcu.s loctiA effect on lictose and galactose metibo-lism. J. Bacteriol. 153:76-83.
2. Crow, V. L., and T. 1). homas. 1984. Properties of a Strepto-cocci.% lacti.s strain that fer-nients liictose slowly. J. Bacteriol.157:28-34.
3. Dvkhuizen, D. E., and D. L. Hartl. 1983. Selection in chemos-
tats. Microbiol. Rev. 47:150-168.4. Garvie, E. 1. 1978. Lactate dehydrogenases of Streptococcus
tlhierophilus J. Dairy Res. 45:515-518.5. Harder, WV.. J. G. Kuenen, and A. Matin. 1977. Microbial
selection in continuouS culture. J. AppI. Bacteriol. 43:1-24.6. Hemme, D., D. VW'ahl, and M. Nardi. 1980. Variations de
equipement enzvmnatique de Stlreptococci.s tlherti- oplhilus. Lait60:111-129.
7. Lawrence, R. C., r. D. Thomas, and B. E. Terzaghi. 1976.Reviews in the progress of dairy science: cheese starters. J.Dairv Res. 43:141-193.
8. Mocquot, G. 1979. Reviews in the progress of dairy science:Swiss-tvpe cheese. J. Dairv Res. 46:133-16t).
9. O'Leary, V. S., and J. H. Woychik. 1976. Utilization of lactose.glucose. and galactose by a mixed culture of Streptococcu(sthiermnoplhilus and Lactohacililus bulgaricus in milk treated withlactase enzyme. AppI. Environ. Microbiol. 32:89-94.
10. Radford, D. R., and R. R. Hull. 1982. Utilization of carbohy-drate in "short method" Cheddar cheese by mesophilic starters.Aust. J. Dairy Technol. 37:104-106.
11. Somkuti, G. A., and D. H. Steinberg. 1979. Adaptability ofStreptococcus tilielr)oplhili.s to lactose, glucose and galactose.J. Food Prot. 42:885-887.
12. Tamime, A. Y., and H. C. Deeth. 1980. Yoghurt: technology andbiochemistry. J. Food Prot. 43:939-977.
13. Thomas, T. D. 1976. Regulation of lactose fermentation in groupN streptococci. Appl. Environ. Microbiol. 32:474-478.
14. Thomas, T. D. 1979. Heterolactic fermentation by homolac-tic starter streptococci resulting from growth with limitingcarbohydrate. N.Z. J. Dairy Sci. Technol. 14:12-15.
15. Thomas, T. D., and V. L. Crow. 1983. Lactose and sucroseutilization by St eptococcu.s tlieimt'iopliiliu.s. FEMS Microbiol.Lett. 17:13-17.
16. Thomas, T. D., D. C. Ellwood, and V. M. C. Longyear. 1979.Change from homo- to heterolactic fermentaition by Streptococ-its latctis resulting from glucose limitation in anaerobic chemo-
stat cultures. J. Bacteriol. 138:109-117.17. Thomas, T. D., K. W. Turner, and V. L. Crow. 1980. Galactose
fermentation by Streptococcu.s (lactis and .Streptococcus crc-m/oris: pathways. products. and regulation. J. Baicteriol.144:672-682.
18. Tinson, W., A. J. Hillier, and G. R. jago. 1982. Metabolism ofStreptococcius tlterniophilus. 1. Utilization of lactose. glucoseand galactose. Aust. J. Dairy Technol. 37:8-13.
19. Tinson, VV., M. F. Ratcliff, A. J. Hillier, and G. R. Jago. 1982.Metabolism of Streptococcius thermophiluts. 3. Influence on thelevel of bacterial metabolites in Cheddar cheese. Aust. J. DairyTechnol. 37:17-21.
20. Turner, K. W., H. A. Morris, and F. G. Martley. 1983. Swiss-type cheese. 11. The role of thermophilic lactobacilli in sugarfermentation. N.Z. J. Dairy Sci. Technol. 18:117-123.
21. Veringa, H. A., T. E. Galesloot, and H. Davelaar. 1968. Symbio-sis in yoghurt. 11. Isolation and identification of a growth fiactorfor Lactohacillius buhlgaricits produced by Streptococcusl titer-Imiop/lii/us. Neth. Milk Dairy J. 22:114-12(.
22. Wright, H. D. 1936. Direct fermentation of disaccharides aindvariation in sugar utilization by Streptococcuts thermuophil/s. J.Pathol. Biacteriol. 43:487-501.
23. Yamada, T., and J. Carlsson. 1975. Regulation of lactate dehv-drogenase and change of fermentation products in streptococci.J. Bacteriol. 124:55-61.
VOL. 48, 1984
on April 10, 2019 by guest
http://aem.asm
.org/D
ownloaded from