identification of a contact between arginine-180 of the catabolite
TRANSCRIPT

Proc. Nadl. Acad. Sci. USAVol. 87, pp. 4717-4721, June 1990Genetics
Identification of a contact between arginine-180 of the catabolitegene activator protein (CAP) and base pair 5 of the DNA sitein the CAP-DNA complex
(protein-DNA interaction/heli-turn-heix motif/cyclic AMP receptor proteinl/Escherichia coli transcription)
XIAOPING ZHANG AND RICHARD H. EBRIGHT*Department of Chemistry and Waksman Institute, Rutgers University, New Brunswick, NJ 08855
Communicated by Jon Beckwith, March 30, 1990 (received for review January 26, 1990)
ABSTRACT We have used site-directed mutagenesis toreplace amino acid 1 of the recognition a-helix of the catabolitegene activator protein (CAP), Arg-180, with glycine and withalanine. Substitution of Arg-180 of CAP eliminated specificitybetween G-C, APT, C'G, and TEA at base pair 5 of the DNAvhalf-site. The effect was position-specific: substitution of Arg-180 did not eliminate specificity between G-C, A-T, C-G, andTEA at base pair 7 of the DNA half-site. We conclude, inagreement with the model for the structure of the CAP-DNAcomplex [Weber, I. & Steitz, T. (1984) Proc. Natl. Acad. Sci.USA 81, 3973-3977; and Ebright, R., Cossart, P., Gicquel-Sanzey, B. & Beckwith, J. (1984) Proc. Natl. Acad. Sci. USA81, 7274-7278], that Arg-180 of CAP makes a specificity-determining contact with base pair 5 of the DNA half-site in theCAP-DNA complex. The identification of the contact by Arg-180 in this report, in conjunction with the identification of thecontact by Glu-181 in a previous report [Ebright, R., Cossart,P., Gicquel-Sanzey, B. & Beckwith, J. (1984) Nature (London)311, 232-235], provides information sufficient to define theorientation of the helix-turn-helix motif ofCAP with respect toDNA in the CAP-DNA complex.
The Escherichia coli catabolite gene activator protein (CAP;also referred to as the cAMP receptor protein, CRP) is asequence-specific DNA-binding protein involved in tran-scription regulation; CAP functions by binding to specificDNA sites located at or near promoters (1, 2). The three-dimensional structure of CAP has been determined to 2.5-Aresolution by x-ray diffraction analysis (3). The protein is adimer of two identical subunits, each of which is 209 aminoacids long and contains a helix-turn-helix DNA-bindingmotif (see refs. 4 and 5). A detailed model has been proposedfor the structure of the CAP-DNA complex (refs. 6-10; Fig.1). In the model, the CAP-DNA complex is twofold sym-metric: one subunit of the CAP dimer interacts with one halfof the DNA site; the other subunit of the CAP dimer interactsin a twofold symmetry-related fashion with the other half ofthe DNA site. One contact between an amino acid of thehelix-turn-helix motif of CAP and a base pair of the DNAhalf-site has been identified experimentally (7, 11, 12): i.e.,amino acid 2 of the recognition a-helix of CAP, Glu-181, hasbeen shown to contact base pair 7 of the DNA half-site.The model for the structure of the CAP-DNA complex
predicts that amino acid 1 of the recognition a-helix of CAP,Arg-180, makes a specificity-determining contact with basepair 5 of the DNA half-site (refs. 6 and 7; Fig. 1B). The modelpredicts that the guanidinium side chain of Arg-180 makeshydrogen bonds with the guanine N7 atom and the guanine 06atom of the canonical base pair, G-C, at base pair 5 of the
DNA half-site. Note that only G'C (not A'T, COG, or TEA) canmake the two putative hydrogen bonds.We have tested experimentally the role of Arg-180 in
specificity at base pair 5 of the DNA half-site. Our approachwas to replace Arg-180 by an amino acid unable to contactbase pair 5, and, then, to ask whether this replacement affectsspecificity at base pair 5. This general approach has beendesignated the "loss-of-contact approach" and has been usedsuccessfully in investigation ofamino acid-base pair contactsby CAP, lac repressor, A repressor, A Cro, AraC protein, theo70 subunit of E. coli RNA polymerase, and the Q.H subunitofBacillus subtilis RNA polymerase (12-20). To eliminate theability of amino acid 180 to contact base pair 5, we havereplaced Arg-180 by glycine, which has no side chain, and byalanine, which has a short, one-carbon side chain.We have used site-directed mutagenesis to construct two
substituted CAP variants: [Gly111CAP and [Alal&]CAP. Wehave investigated the profiles of specificity ofwild-type CAP,[Gly1801CAP, and [Alal&ICAP with respect to base pairs 5and 7 of the DNA half-site. We have found that replacementof Arg-180 ofCAP by glycine or alanine eliminates the abilityof CAP to distinguish between canonical and noncanonicalbase pairs at base pair 5 of the DNA half-site. The effect isposition-specific; replacement of Arg-180 by glycine or ala-nine does not eliminate the ability of CAP to distinguishbetween canonical and noncanonical base pairs at base pair7 of the DNA site. These results indicate that Arg-180 ofCAPcontrols specificity at base pair 5. We conclude, in agreementwith the model in Fig. 1 (6-10), that Arg-180 of CAP makesa specificity-determining contact with base pair 5 in theCAP-DNA complex.
MATERIALS AND METHODSBacterial Strains. The E. coli K-12 strains constructed in
this work (Table 1) are derivatives of strain XA102 (Alac-proABXI I I argEam metB ara rpoB nal Su-2; ref. 21). The crpand strA markers are from strain CA8445 (HfrH Acrp-45Acya-854 strA thi; ref. 22). The fnr-1 and zci::TnlO markersare from strain ECL323 (fnr-J zci::TnIO AlacUJ69 araD139motA strA thi; ref. 23). The fnr-J marker was included toprevent Fnr-dependent transcription of lacPJ(-68T;-55A)(see ref. 24).
Strains XAE400, XAE451, XAE452, XAE453, XAE471,XAE472, and XAE473 were constructed as follows. StrainXAE14 (XA102 Acrp-45 strA fnr-J zci::TnJO pHA5) waslysogenized with Ai434placS and with derivatives of Ai434-placS having substitutions at base pairs -68 and -55, or basepairs -66 and -57, of lacPJ (Table 2); lysogens wereidentified by blue colony color on 5-bromo-4-chloro-3-indolylP-D-galactoside indicator plates. The resulting lysogens were
Abbreviation: CAP, catabolite gene activator protein.*To whom reprint requests should be addressed at: WaksmanInstitute, Rutgers University, New Brunswick, NJ 08855.
4717
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

4718 Genetics: Zhang and Ebright
FIG. 1. (A) Model of the CAP-DNA complex (ref. 9; see also refs. 6-8, 10). Each subunit of the CAP dimer interacts with one half of thetwofold-symmetric consensus DNA site. The DNA site in the CAP-DNA complex is bent; in the present model, there is one 20° kink betweenbase pairs 1 and 2 of the DNA half-site and one 350 kink between base pairs 6 and 7 of the DNA half-site. CAP is illustrated in a ribbonrepresentation; cAMP and DNA are illustrated in Dreiding representations. The helix-turn-helix motif of each subunit ofCAP is illustrated ingreen. (B) Model for the interaction between the helix-turn-helix motif ofCAP and nucleotide pairs 4-9 of the DNA half-site. The model predictsthat Arg-180 contacts base pair 5 of the DNA half-site. The model predicts that Glu-181 contacts base pair 7 of the DNA half-site. These aminoacids and the bases they are predicted to contact are illustrated in green. The model predicts that several additional amino acids contact DNAbase-pair edges (Thr-182 and Arg-185) and/orDNA phosphates [Thr-168, Arg-169, Gln-170, Ser-179 (possibly through a water molecule), Thr-182(possibly through a water molecule), and Lys-188]. These amino acids are illustrated in yellow. Coordinates for the structure of CAP were fromref. 3. The coordinates were obtained from the Brookhaven Protein Data Base (accession code PDB3GAP). Coordinates for the structure ofDNA were generated using the program DNA-FIT-MAN, written by J. Warwicker (Yale University, New Haven, CT). Molecular modeling wasperformed on an Evans & Sutherland PS390 graphics system, using the software package INSIGHT (BIOSYM).
grown in liquid culture in the absence of ampicillin selection;isolates that had lost plasmid pHA5 were identified by redcolony color on maltose/tetrazolium indicator plates. Theisolates utilized were established to be single-copy lysogens,based on segregation patterns and on levels of CAP-independent P-galactosidase expression.
Bacteriophage. A list of bacteriophage used or constructedin this work is presented in Table 2. M13mp2-IacPI(-68C;
-SSG) and M13mp2-lacPI(-68T;-SSA) were constructedfrom M13mp2 by site-directed mutagenesis (see method inref. 26). Derivatives of Ai434placS having substitutions atbase pairs -68 and -55 of lac were constructed by homol-ogous recombination between Ai434placS-PIL162 and theappropriate derivatives of M13mp2 (see method in ref. 27).Recombinants were identified by light blue plaque color on5-bromo-4-chloro-3-indolyl P3-D-galactoside indicator plates
Proc. Natl. Acad. Sci. USA 87 (1990)

Proc. Natl. Acad. Sci. USA 87 (1990) 4719
Table 1. E. coli K-12 strains constructed in this studyStrain GenotypeXAE400 XA102 Acrp45 strA fnr-J zci::TnlO Ai434placSXAE451 XA102 Acrp45 strA fnr-1 zci::TnJO Ai434placS-Pl(-68A;-SST)XAE452 XA102 Acrp45 strA fnr-1 zci::TnlO Ai434placS-PJ(-68C;-SSG)XAE453 XA102 Acrp45 strA fnr-I zci::TnJO Ai434placS-PI(-68T;-SSA)XAE471 XA102 Acrp45 strA fnr-I zci::TnlO Ai434placS-PI(-66A;-57T)XAE472 XA102 Acrp45 strA fnr-1 zci::TnlO Ai434plac5-PJ(-66C;-57G)XAE473 XA102 Acrp45 strA fnr-1 zci::TnlO Ai434placS-PJ(-66T;-57A)To analyze the profile of specificity of CAP variant X, the plasmid encoding X was introduced into
each of the strains (see Materials and Methods). For example, the set of strains constructed to analyzethe specificity of wild-type CAP carried pHA5; these strains are designated XAE400/CRP, XAE451/CRP, etc.
and were verified by nucleotide sequence determination ofthe lac promoter region.
Plasmids Encoding CAP and CAP Derivatives. The plasmidencoding wild-type CAP (pHA5) has been described (28).Plasmids encoding [Gly'8']CAP (pTK180G) and [Ala180]CAP(pTK180A) were constructed by use of site-directed muta-genesis to introduce substitutions into M13mpl8-CRP (seemethod in ref. 26), followed by subcloning of the 3.5-kilobaseBamHI-BamHI crp fragment into the BamHI site of plasmidpBR322 (29). For each plasmid, the nucleotide sequence ofthe complete crp structural gene and promoter was verified.Plasmids were constructed and maintained in strain CA8445(HfrH Acrp45 Acya-854 strA thi; ref. 22).In Vivo DNA Binding. Experimentation. Seven E. coli K-12
"tester strains" were used: XAE400, XAE451, XAE452, XAE-453, XAE471, XAE472, and XAE473 (Table 1). Each testerstrain has two important components. (i) Acrp45, a deletionof the gene that encodes wild-type CAP (22). Acrp45 is >5kilobases long; it deletes all homology to the 3.5-kilobase crpinsert of the plasmids used in this study (30). (ii) lacZ, thegene that encodes 3-galactosidase. In tester strain XAE400,lacZ is placed under the control of the wild-type lacPl DNAsite for CAP. In the remaining six tester strains, lacZ is placedunder the control of derivatives of the lacPJ DNA site forCAP that have G-C -* A-T, G-C -- C-G, and G-C -* T-Asubstitutions at base pair 5 of each DNA half-site, or G-C -*A-T, G-C -- C-G, and G-C -* T-A substitutions at base pair 7of each DNA half-site (sequences in Fig. 2). The lacZ gene ispresent on a Ai434plac5 prophage stably integrated into thebacterial chromosome at attA.To analyze the profile of specificity of CAP variant X, the
plasmid encoding X was introduced into each of the seventester strains. Note that each of the resulting plasmid-bearingstrains was identical except for two base pairs in lacPJ.
Table 2. Bacteriophage used in this study
Bacteriophage
Ai434plac5Ai434placS-PlL162Ai434plac5-PI(-68A;-SST)Ai434plac5-PI(-68C;-SSG)Ai434plac5-PI(-68T;-55A)Ai434plac5-PJ(-66A;-57T)Ai434plac5-Pl(-66C;-57G)Ai434plac5-P1(-66T;-57A)M13mp2M13mp2-lacPI(-68A;-SST)M13mp2-lacPI(-68C;-55G)M13mp2-lacPI(-68T;-55A)M13mpl8-CRPM13mpl8-180GM13mpl8-180A
Source or
ref.
1310This workThis workThis work10101025T. KunkelThis workThis work10This workThis work
Therefore, in each ofthe resulting plasmid-bearing strains theintracellular concentration of CAP was equal.For each plasmid-bearing strain, the differential rate of
P-galactosidase synthesis was determined by the method ofMiller (31). Cultures were grown in LB medium (31) con-taining ampicillin at 100 ,g/ml. Data were corrected forbackground-i.e., for the differential rate of 3-galactosidasesynthesis in the absence of CAP or CAP variant.Data reduction. Ratios of equilibrium dissociation con-
stants, KD1/KD,2, were calculated from the differential ratesof B-galactosidase synthesis as described in ref. 10 (see alsorefs. 13 and 14). Briefly,
0 = Z/ZM, [1]
where 0 denotes the fractional occupancy of lacPJ by CAP;Z denotes the measured differential rate of f3-galactosidasesynthesis; and ZM denotes the maximum differential rate off3-galactosidase synthesis (equal to 5000 for the tester strainsused, determined separately; see ref. 10). And
KDJ1 02(1-01)KD,2 01(1-02)'
[2]
where KDJ1 denotes the equilibrium dissociation constant forinteraction of CAP variant X with the DNA site in strain 1,and KD.2 denotes the equilibrium dissociation constant forinteraction of the identical CAP variant X with the DNA sitein strain 2.
This method makes the assumption that, for a givenfractional occupancy of lacPI, CAP variant X stimulates,B-galactosidase synthesis to the same extent as does wild-type CAP. In most instances, this assumption is valid. Inprevious work, we have measured ratios of equilibriumdissociation constants exhibited by wild-type CAP and bymore than 20 substituted CAP variants-both in vivo, usingthe method above, and in vitro, using the nitrocellulose filterbinding assay (ref. 10; R.H.E., X.Z., A. Gunasekera, M.Smith, and T. Kunkel, unpublished results). We have foundgood agreement between the in vivo results and the in vitroresults.The method above yields high-precision data for 0 = 0.002
to 0 = 0.95 (precision of estimate of 0 typically within 10%).Where 0 < 0.002, the value 0 = 0.002 was used to calculatea minimum estimate for the ratio KD/KDp+.
RESULTSWe have measured the profiles of specificity of wild-typeCAP, [Gly1"1CAP, and [Alal80]CAP with respect to G-C,A-T, C-G, and T-A at base pairs 5 and 7 of the DNA half-site.Fig. 2 shows the DNA sites used as ligands in the analysis.They consist of the wild-type lacPJ DNA site for CAP, threederivatives of the lacPJ DNA site for CAP that have G-C -)
A-T, G-C -- C-G, and G-C -- T-A substitutions at base pair 5
Genetics: Zhang and Ebright

4720 Genetics: Zhang and Ebright
1 2 3 4 5 S 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 2
A A - T G T G A - - - - - - T C A C A - T TT T - A C A C T - - -a- - - A G T G T - A A
-70 45
LACP1 T A - T G T G A - - - - - - T C AA T - A C A C T - - -a- - - A G T
LACP1(-68A; -55T)
LACP1(-68C; -55G)
LACP1(-68T; -55A)
LACP1(-66A; -57T)
LACP1(-66C; -57G)
LACP1(-66T; -57A)
AT
C
G
T
A
CT - A TGA - T A
T
A
GC
AT
A
G
C
AT
of each DNA half-site, and three derivatives of the lacPlDNA site for CAP that have G-C -b APT, G-C -k COG, and G-C
-- TEA substitutions at base pair 7 of each DNA half-site. Thedata were obtained using the quantitative in vivo DNAbinding assay described in ref. 10 (see also refs. 13 and 14).In Tables 3 and 4, the data are expressed as the ratioKD/KDP+, i.e., as the equilibrium dissociation constant forthe interaction of CAP variant X with a substituted lacPJDNA site, divided by the equilibrium dissociation constantfor the interaction of the identical CAP variant X with thewild-type lacPI DNA site.
Specificity at Base Pair 5 of the DNA Half-Site. Wild-typeCAP exhibits robust specificity for G-C at base pair 5 of theDNA half-site (Table 3). Wild-type CAP interacts 200-fold,2000-fold, and 30-fold more strongly with the wild-type lacPJDNA site for CAP than it interacts with the derivatives of thelacPJ DNA site that have, respectively, APT, C-G, and T-A atbase pair 5 of each DNA half-site. These values correspondto specificity free energies of -3.2 kcal/mol, -4.7 kcal/mol,and -2.1 kcal/mol.
In contrast, [Gly180]CAP and [Ala']CAP exhibit essen-tially no specificity between G-C and A-T, COG, or T-A at basepair 5 of the DNA half-site (Table 3). [Gly'80]CAP and[Ala180]CAP interact essentially equally with each of the fourDNA sites. (In fact, [Gly180]CAP and [Ala1I]CAP exhibit a
borderline-significant preference for the derivative of thelacPJ DNA site that has the noncanonical base pair T-A atbase pair 5 of the DNA site.) These results demonstrate a
significant change in the energetics of specificity at base pair5 of the DNA half-site upon substitution of Arg-180.
Specificity at Base Pair 7 of the DNA Half-Site. As a control,we have asked whether the observed change in the energeticsof specificity upon substitution of Arg-180 ofCAP is position-specific. To do this, we have measured the profiles ofspecificity of wild-type CAP, [Gly180]CAP, and [Ala180]CAPwith respect to G-C, A*T, C-G, and T-A at a second position
FIG. 2. DNA sites referred to in thisstudy. (Upper) The consensus DNA sitefor CAP (9, 12, 32). The site is 22 base pairslong and has perfect twofold sequencesymmetry. The twofold axis is located be-tween base pairs 11 and 12. (Lower) Thewild-type lacPl DNA site for CAP [basepairs -72 to -51 with respect to the startpoint of the lacPI promoter (33)]. Thesymmetric A-T, C-G, and TEA substitutionsat base pair 5 of the DNA half-site, and thesymmetric APT, C-G, and TEA substitutionsat base pair 7 of the DNA half-site, areindicated beneath the sequence.
within the DNA half-site, i.e., at base pair 7 of the DNAhalf-site.
Wild-type CAP exhibits robust specificity for G'C at basepair 7 of the DNA half-site (refs. 10, 12; Table 4). Wild-typeCAP interacts >5000-fold, >5000-fold, and 700-fold morestrongly with the wild-type lacPI DNA site for CAP than itinteracts with the derivatives of the IacP1 DNA site thathave, respectively, ANT, COG, and T*A at base pair 7 of eachDNA half-site.
Within the limits of the data, [Gly'80]CAP and [Ala']CAPretain wild-type specificity (i.e., retain specificity for G-C) atbase pair 7 of the DNA half-site (Table 4). [Gly1"]CAP and[Ala'80]CAP interact at least 1 order of magnitude morestrongly with the wild-type lacPJ DNA site for CAP than withthe derivatives ofthe lacPJ DNA site that have ANT, C-G, andT-A at base pair 7 of each DNA half-site. It has not beenpossible to quantify with more precision the specificities of[Gly'80]CAP and [Ala1"]CAP with respect to base pair 7 ofthe DNA half-site (due to the low DNA-binding affinities of[Gly180]CAP and [Alal80ICAP; R.H.E., A. Gunasekera, andP. S. Pendergrast, unpublished results). Nevertheless, thequalitative pattern is clear.
DISCUSSIONReplacement of Arg-180 of CAP by glycine or alanine elim-inated specificity at base pair 5 of the DNA half-site but didnot eliminate specificity at base pair 7 of the DNA half-site.These results indicate that Arg-180 of CAP controls speci-ficity at base pair 5 of the DNA half-site. We conclude, inagreement with the model in Fig. 1 (6-10), that Arg-180 ofCAP makes a specificity-determining contact with base pair5 of the DNA half-site in the CAP-DNA complex.The results do not exclude the possibility that Arg-180 of
CAP controls specificity at base pair 5 of the DNA half-siteindirectly-i.e., without a direct contact. For example, an-other protein functional group could make the contact with
Table 3. In vivo data: Recognition of DNA sites symmetrically altered at positions 5 and 18
Wild-type CAP [Gly180]CAP [Ala'80]CAPDNA site Z KD/KD.P Z KD/KDP+ Z KD/KD.p+
P+ (5G;18C) 4500 [1] 80 [1] 150 [1]5A;18T 260 200 230 0.3 550 0.35C;18G 25 2000 320 0.2 780 0.25T;18A 1100 30 980 0.06 1500 0.07
PI denotes the wild-type lacP1 DNA site; substituted derivatives ofthe lacPJ DNA site are identifiedby the position and sequence of the substitutions (sequences in Fig. 2). Tester strains XAE400,XAE451, XAE452, and XAE453 were used. Values ofZ are presented for reference only and have beencorrected for CAP-independent f3-galactosidase expression.
CAP CONSENSUS
Proc. Natl. Acad. Sci. USA 87 (1990)
Proc. Natl. Acad. Sci. USA 87 (1990) 4721
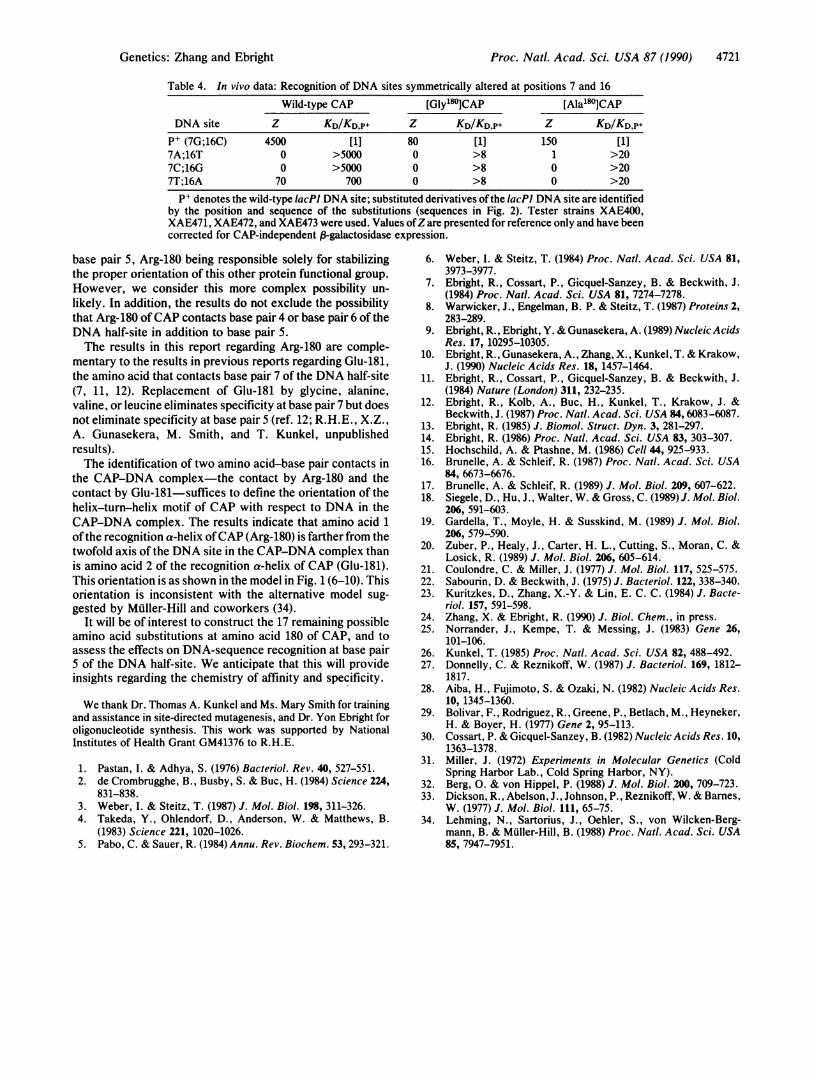
Table 4. In vivo data: Recognition of DNA sites symmetrically altered at positions 7 and 16Wild-type CAP [Gly'80]CAP [Ala180lCAP
DNA site Z KD/KDp+ Z KD/KD.P+ Z KD/KD.p+P+ (7G;16C) 4500 [1] 80 [1] 150 [1]7A;16T 0 >5000 0 >8 1 >207C;16G 0 >5000 0 >8 0 >207T;16A 70 700 0 >8 0 >20
PI denotes the wild-type IacPl DNA site; substituted derivatives ofthe lacPJ DNA site are identifiedby the position and sequence of the substitutions (sequences in Fig. 2). Tester strains XAE400,XAE471, XAE472, and XAE473 were used. Values ofZare presented for reference only and have beencorrected for CAP-independent 3-galactosidase expression.
base pair 5, Arg-180 being responsible solely for stabilizingthe proper orientation of this other protein functional group.However, we consider this more complex possibility un-likely. In addition, the results do not exclude the possibilitythat Arg-180 of CAP contacts base pair 4 or base pair 6 of theDNA half-site in addition to base pair 5.The results in this report regarding Arg-180 are comple-
mentary to the results in previous reports regarding Glu-181,the amino acid that contacts base pair 7 of the DNA half-site(7, 11, 12). Replacement of Glu-181 by glycine, alanine,valine, or leucine eliminates specificity at base pair 7 but doesnot eliminate specificity at base pair 5 (ref. 12; R.H.E., X.Z.,A. Gunasekera, M. Smith, and T. Kunkel, unpublishedresults).The identification of two amino acid-base pair contacts in
the CAP-DNA complex-the contact by Arg-180 and thecontact by Glu-181-suffices to define the orientation of thehelix-turn-helix motif of CAP with respect to DNA in theCAP-DNA complex. The results indicate that amino acid 1ofthe recognition a-helix ofCAP (Arg-180) is farther from thetwofold axis of the DNA site in the CAP-DNA complex thanis amino acid 2 of the recognition a-helix of CAP (Glu-181).This orientation is as shown in the model in Fig. 1(6-10). Thisorientation is inconsistent with the alternative model sug-gested by Muller-Hill and coworkers (34).
It will be of interest to construct the 17 remaining possibleamino acid substitutions at amino acid 180 of CAP, and toassess the effects on DNA-sequence recognition at base pair5 of the DNA half-site. We anticipate that this will provideinsights regarding the chemistry of affinity and specificity.
We thank Dr. Thomas A. Kunkel and Ms. Mary Smith for trainingand assistance in site-directed mutagenesis, and Dr. Yon Ebright foroligonucleotide synthesis. This work was supported by NationalInstitutes of Health Grant GM41376 to R.H.E.
1. Pastan, I. & Adhya, S. (1976) Bacteriol. Rev. 40, 527-551.2. de Crombrugghe, B., Busby, S. & Buc, H. (1984) Science 224,
831-838.3. Weber, I. & Steitz, T. (1987) J. Mol. Biol. 198, 311-326.4. Takeda, Y., Ohlendorf, D., Anderson, W. & Matthews, B.
(1983) Science 221, 1020-1026.5. Pabo, C. & Sauer, R. (1984) Annu. Rev. Biochem. 53, 293-321.
6. Weber, I. & Steitz, T. (1984) Proc. Natl. Acad. Sci. USA 81,3973-3977.
7. Ebright, R., Cossart, P., Gicquel-Sanzey, B. & Beckwith, J.(1984) Proc. Natl. Acad. Sci. USA 81, 7274-7278.
8. Warwicker, J., Engelman, B. P. & Steitz, T. (1987) Proteins 2,283-289.
9. Ebright, R., Ebright, Y. & Gunasekera, A. (1989) Nucleic AcidsRes. 17, 10295-10305.
10. Ebright, R., Gunasekera, A., Zhang, X., Kunkel, T. & Krakow,J. (1990) Nucleic Acids Res. 18, 1457-1464.
11. Ebright, R., Cossart, P., Gicquel-Sanzey, B. & Beckwith, J.(1984) Nature (London) 311, 232-235.
12. Ebright, R., Kolb, A., Buc, H., Kunkel, T., Krakow, J. &Beckwith, J. (1987) Proc. Natl. Acad. Sci. USA 84, 6083-6087.
13. Ebright, R. (1985) J. Biomol. Struct. Dyn. 3, 281-297.14. Ebright, R. (1986) Proc. Natl. Acad. Sci. USA 83, 303-307.15. Hochschild, A. & Ptashne, M. (1986) Cell 44, 925-933.16. Brunelle, A. & Schleif, R. (1987) Proc. Natl. Acad. Sci. USA
84, 6673-6676.17. Brunelle, A. & Schleif, R. (1989) J. Mol. Biol. 209, 607-622.18. Siegele, D., Hu, J., Walter, W. & Gross, C. (1989) J. Mol. Biol.
206, 591-603.19. Gardella, T., Moyle, H. & Susskind, M. (1989) J. Mol. Biol.
206, 579-590.20. Zuber, P., Healy, J., Carter, H. L., Cutting, S., Moran, C. &
Losick, R. (1989) J. Mol. Biol. 206, 605-614.21. Coulondre, C. & Miller, J. (1977) J. Mol. Biol. 117, 525-575.22. Sabourin, D. & Beckwith, J. (1975) J. Bacteriol. 122, 338-340.23. Kuritzkes, D., Zhang, X.-Y. & Lin, E. C. C. (1984) J. Bacte-
riol. 157, 591-598.24. Zhang, X. & Ebright, R. (1990) J. Biol. Chem., in press.25. Norrander, J., Kempe, T. & Messing, J. (1983) Gene 26,
101-106.26. Kunkel, T. (1985) Proc. Nail. Acad. Sci. USA 82, 488-492.27. Donnelly, C. & Reznikoff, W. (1987) J. Bacteriol. 169, 1812-
1817.28. Aiba, H., Fujimoto, S. & Ozaki, N. (1982) Nucleic Acids Res.
10, 1345-1360.29. Bolivar, F., Rodriguez, R., Greene, P., Betlach, M., Heyneker,
H. & Boyer, H. (1977) Gene 2, 95-113.30. Cossart, P. & Gicquel-Sanzey, B. (1982) Nucleic Acids Res. 10,
1363-1378.31. Miller, J. (1972) Experiments in Molecular Genetics (Cold
Spring Harbor Lab., Cold Spring Harbor, NY).32. Berg, 0. & von Hippel, P. (1988) J. Mol. Biol. 200, 709-723.33. Dickson, R., Abelson, J., Johnson, P., Reznikoff, W. & Barnes,
W. (1977) J. Mol. Biol. 111, 65-75.34. Lehming, N., Sartorius, J., Oehler, S., von Wilcken-Berg-
mann, B. & Muller-Hill, B. (1988) Proc. Natl. Acad. Sci. USA85, 7947-7951.
Genetics: Zhang and Ebright