science 2002 chen 1634 5.pdf a beautiful pathway
DESCRIPTION
Science 2002 Chen 1634 5.PDF a Beautiful PathwayTRANSCRIPT

DOI: 10.1126/science.1071924, 1634 (2002);296 Science
Guoqing Chen and David V. GoeddelTNF-R1 Signaling: A Beautiful Pathway
This copy is for your personal, non-commercial use only.
clicking here.colleagues, clients, or customers by , you can order high-quality copies for yourIf you wish to distribute this article to others
here.following the guidelines
can be obtained byPermission to republish or repurpose articles or portions of articles
): October 15, 2014 www.sciencemag.org (this information is current as of
The following resources related to this article are available online at
http://www.sciencemag.org/content/296/5573/1634.full.htmlversion of this article at:
including high-resolution figures, can be found in the onlineUpdated information and services,
http://www.sciencemag.org/content/296/5573/1634.full.html#relatedfound at:
can berelated to this article A list of selected additional articles on the Science Web sites
576 article(s) on the ISI Web of Sciencecited by This article has been
http://www.sciencemag.org/content/296/5573/1634.full.html#related-urls100 articles hosted by HighWire Press; see:cited by This article has been
http://www.sciencemag.org/cgi/collection/cell_biolCell Biology
subject collections:This article appears in the following
registered trademark of AAAS. is aScience2002 by the American Association for the Advancement of Science; all rights reserved. The title
CopyrightAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. (print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by theScience
on
Oct
ober
15,
201
4w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
o
n O
ctob
er 1
5, 2
014
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
on
Oct
ober
15,
201
4w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
TNF-R1 Signaling: A Beautiful PathwayGuoqing Chen and David V. Goeddel*
Tumor necrosis factor (TNF) is a major mediator of apoptosis as well asinflammation and immunity, and it has been implicated in the pathogenesisof a wide spectrum of human diseases, including sepsis, diabetes, cancer,osteoporosis, multiple sclerosis, rheumatoid arthritis, and inflammatory bow-el diseases. The interaction of TNF with TNF receptor–1 ( TNF-R1) activatesseveral signal transduction pathways. A common feature of each pathway isthe TNF-induced formation of a multiprotein signaling complex at the cellmembrane. Over the past decade, many of the components and mechanismsof these signaling pathways have been elucidated. We provide an overview ofcurrent knowledge of TNF signaling and introduce an STKE Connections Mapthat depicts a canonical view of this process.
The anticancer activity now known as tumornecrosis factor (TNF) was first describedmore than a century ago. However, it wasn’tuntil 1984 that human TNF was purified andits encoding cDNA was cloned and ex-pressed. The subsequent availability of re-combinant TNF led to a rapid cataloging ofTNF’s pleiotropic activities.
In addition to triggering apoptosis of certaintumor cells, TNF mediates the inflammatoryresponse and regulates immune function. Inap-propriate production of TNF or sustained acti-vation of TNF signaling has been implicated inthe pathogenesis of a wide spectrum of humandiseases, including sepsis, cerebral malaria, di-abetes, cancer, osteoporosis, allograft rejection,and autoimmune diseases such as multiple scle-rosis, rheumatoid arthritis, and inflammatorybowel diseases.
In the past dozen years, the molecular detailsof signal transduction by TNF gradually havebeen unveiled. The TNF Pathway ConnectionsMap (http://stke.sciencemag.org/cgi/cm/CMP_7107) at Science’s Signal TransductionKnowledge Environment presents currentknowledge of the pathway’s componentsand connections between them (1). Today,our understanding of the TNF signalingnetwork provides a paradigm for elucidat-ing the signaling pathways utilized by otherTNF-related proteins and their receptors(2). This understanding has also led to thedevelopment of novel therapies that neutral-ize the deleterious effects of TNF for treat-ment of recalcitrant inflammatory condi-tions such as rheumatoid arthritis (3).
TNF is a homotrimer of 157 amino acidsubunits primarily produced by activated macro-phages. TNF signals through two distinct cellsurface receptors, TNF-R1 and TNF-R2. Multi-ple experimental approaches have revealed thatTNF-R1 initiates the majority of TNF’s biolog-
ical activities. The binding of TNF to TNF-R1triggers a series of intracellular events that ulti-mately result in the activation of two majortranscription factors, nuclear factor kB (NF-kB)and c-Jun. These transcription factors are respon-sible for the inducible expressionof genes important for diverse bi-ological processes, including cellgrowth and death, development,oncogenesis, and immune, in-flammatory, and stress responses.
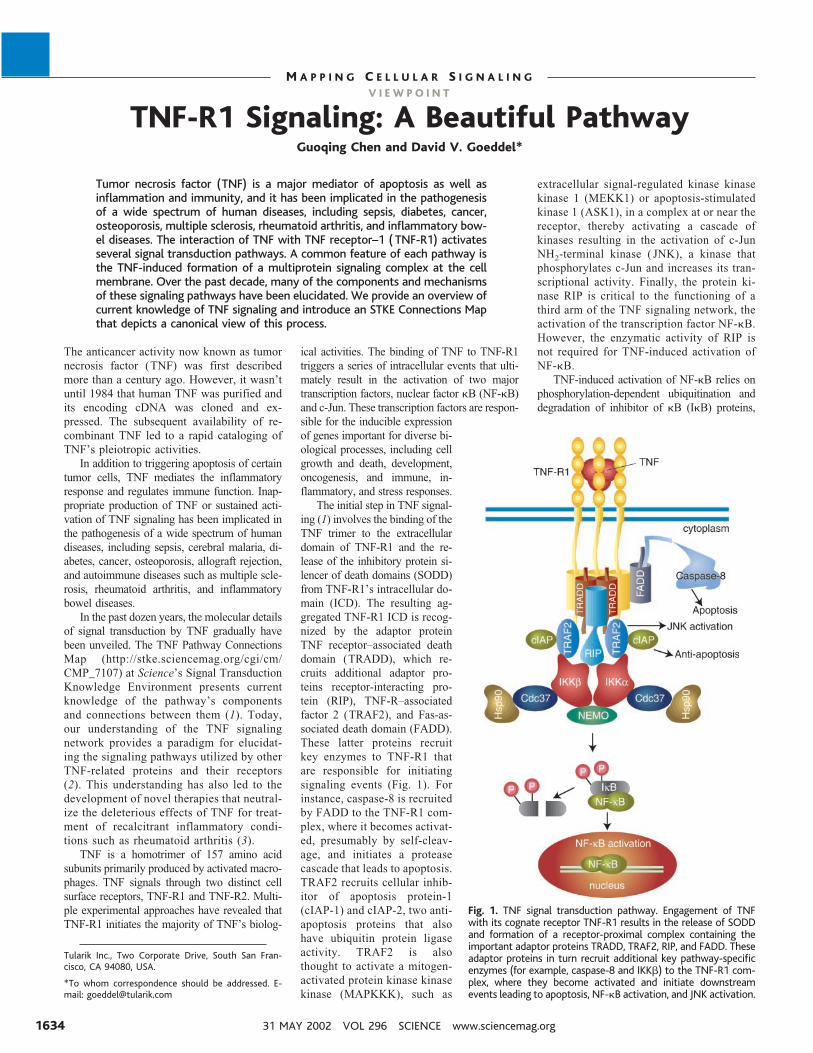
The initial step in TNF signal-ing (1) involves the binding of theTNF trimer to the extracellulardomain of TNF-R1 and the re-lease of the inhibitory protein si-lencer of death domains (SODD)from TNF-R1’s intracellular do-main (ICD). The resulting ag-gregated TNF-R1 ICD is recog-nized by the adaptor proteinTNF receptor–associated deathdomain (TRADD), which re-cruits additional adaptor pro-teins receptor-interacting pro-tein (RIP), TNF-R–associatedfactor 2 (TRAF2), and Fas-as-sociated death domain (FADD).These latter proteins recruitkey enzymes to TNF-R1 thatare responsible for initiatingsignaling events (Fig. 1). Forinstance, caspase-8 is recruitedby FADD to the TNF-R1 com-plex, where it becomes activat-ed, presumably by self-cleav-age, and initiates a proteasecascade that leads to apoptosis.TRAF2 recruits cellular inhib-itor of apoptosis protein-1(cIAP-1) and cIAP-2, two anti-apoptosis proteins that alsohave ubiquitin protein ligaseactivity. TRAF2 is alsothought to activate a mitogen-activated protein kinase kinasekinase (MAPKKK), such as
extracellular signal-regulated kinase kinasekinase 1 (MEKK1) or apoptosis-stimulatedkinase 1 (ASK1), in a complex at or near thereceptor, thereby activating a cascade ofkinases resulting in the activation of c-JunNH2-terminal kinase ( JNK), a kinase thatphosphorylates c-Jun and increases its tran-scriptional activity. Finally, the protein ki-nase RIP is critical to the functioning of athird arm of the TNF signaling network, theactivation of the transcription factor NF-kB.However, the enzymatic activity of RIP isnot required for TNF-induced activation ofNF-kB.
TNF-induced activation of NF-kB relies onphosphorylation-dependent ubiquitination anddegradation of inhibitor of kB (IkB) proteins,
Tularik Inc., Two Corporate Drive, South San Fran-cisco, CA 94080, USA.
*To whom correspondence should be addressed. E-mail: [email protected]
Fig. 1. TNF signal transduction pathway. Engagement of TNFwith its cognate receptor TNF-R1 results in the release of SODDand formation of a receptor-proximal complex containing theimportant adaptor proteins TRADD, TRAF2, RIP, and FADD. Theseadaptor proteins in turn recruit additional key pathway-specificenzymes (for example, caspase-8 and IKKb) to the TNF-R1 com-plex, where they become activated and initiate downstreamevents leading to apoptosis, NF-kB activation, and JNK activation.
31 MAY 2002 VOL 296 SCIENCE www.sciencemag.org1634
M A P P I N G C E L L U L A R S I G N A L I N G
V I E W P O I N T

which normally retain NF-kB within the cyto-plasm of unstimulated cells. The past 5 yearshave witnessed tremendous advances in our un-derstanding of this branch of the TNF signalingnetwork. Especially noteworthy was the identi-fication of the multiprotein IkB kinase (IKK)complex that mediates phosphorylation of IkB ina TNF-dependent manner (4). The core of theIKK complex consists of two catalytic subunits,IKKa and IKKb, and a regulatory subunit, NF-kB essential modulator (NEMO, or IKKg). Inaddition, the IKK complex contains a kinase-specific chaperone consisting of Cdc37 andHsp90 that plays a role in shuttling the complexfrom the cytoplasm to the membrane. The IKKcomplex is also recruited to TNF-R1, where itbecomes activated within minutes of TNF treat-ment. This activation depends on RIP, indicatingthat the IKK activation within the receptor com-plex likely occurs through a RIP-dependent in-termediate factor, perhaps a kinase. Gene knock-out studies in mice have established essentialroles for IKKb in TNF-induced activation ofNF-kB, and for NEMO in regulation of IKKcomplex activation in response to numerous up-stream signals. In contrast, IKKa plays only a
minor role in TNF-induced activation of NF-kB,but it has other important functions, such asserving as a NF-kB2/p100 kinase in B cells.
An interesting feature of the TNF signalingnetwork is the existence of extensive cross talkbetween the apoptosis, NF-kB, and JNK signal-ing pathways that emanate from TNF-R1. In theabsence of NF-kB activity, cellular susceptibil-ity to TNF- induced apoptosis increases, where-as enforced activation of NF-kB protects againstapoptosis. Similarly, TNF-induced JNKactivation is stronger and more prolonged incells lacking NF-kB, and the products of severalNF-kB–activated genes inhibit activation ofJNK by TNF. Moreover, NF-kB activationprompts the resynthesis of IkB and other inhib-itory molecules, such as the cIAPs, thereby add-ing another layer of regulation of the durationand amplitude of TNF signaling.
To date, most of the players in the TNFpathway have been validated by both biochem-ical and genetic means, thus providing a richsource of potential drug targets for the develop-ment of a new generation of anti-inflammatoryagents. However, many questions remain unan-swered. For example, what MAPKKK initiates
the kinase cascade that activates JNK, and howis this kinase recruited to TNF-R1 and activatedwithin the receptor complex in response toTNF? In the case of IKK complex activation,the possibility remains that an intermediatefactor or kinase is required between RIP andNEMO. Unraveling the molecular details ofhow the enzymes like caspase-8 and the IKKcomplex become activated within the TNF-R1complex will be key to a full understandingof the dynamic nature of TNF signaling.Finally, the molecular basis for cross talkbetween TNF-mediated apoptosis, NF-kB,and JNK signaling pathways is not wellunderstood. Deciphering these puzzles willgreatly help interpret how a specific out-come of TNF signaling is achieved in dis-tinct biological contexts.
References1. G. Chen, D. V. Goeddel, TNF Pathway, Science’s STKE
(Connections Map, as seen May 2002), http://stke.sciencemag.org/cgi/cm/CMP_7107.
2. R. M. Locksley, N. Killeen, M. J. Lenardo, Cell 104, 487(2001).
3. M. Feldmann, R. N. Maini, Annu. Rev. Immunol. 19,163 (2001).
4. S. Ghosh, M. Karin, Cell 109, S81 (2002).
V I E W P O I N T
The Fas Signaling Pathway:More Than a Paradigm
Harald Wajant*
Apoptosis and related forms of cell death have central importance in development,homeostasis, tumor surveillance, and the function of the immune system. Apopto-sis is initiated by two principal pathways. The intrinsic pathway emerges frommitochondria, whereas the extrinsic pathway is activated by the ligation of deathreceptors. This Viewpoint introduces the basic mechanisms of the extrinsic path-way, using the example of the prototypical death receptor Fas and its role inapoptosis, but it also points out the increasingly understood importance of thisreceptor as a non-apoptotic signal transducer.
Fas (also called Apo-1 or CD95) is a deathdomain–containing member of the tumor necro-sis factor receptor (TNFR) superfamily. It has acentral role in the physiological regulation ofprogrammed cell death and has been implicatedin the pathogenesis of various malignancies anddiseases of the immune system (1, 2) [see FasSignaling Pathway, http://stke.sciencemag.org/cgi/cm/CMP_7966 (3) and Fas Signaling Path-way in Cardiomyocytes, http://stke.sciencemag.org/cgi/cm/CMP_9993 (4)]. Although the Fasligand (FasL)–Fas system has been appreciatedmainly with respect to its death-inducing func-tion, it also transduces proliferative and activat-ing signals through pathways that are still poorlydefined (1, 2).
In the absence of membrane-bound ligand,inactive complexes of Fas are formed by thepre–ligand-binding assembly domain of the mol-ecule (2). Interaction with membrane-boundFasL (or agonistic antibodies) reorganizes thesecomplexes and allows the formation of a death-inducing signaling complex (DISC). The FasDISC contains the adaptor protein Fas-associat-ed death domain protein (FADD) and caspases 8and 10, which can initiate the process of apopto-sis. FasL-induced clustering of Fas, FADD, andcaspase-8 or -10 within the DISC results inautoproteolytic processing of these caspases byinduced proximity and in release of the pro-cessed active proteases (Fig. 1). In type I cells,processed caspase-8 is sufficient to directly ac-tivate other members of the caspase family,whose action on defined substrates paves theway to the execution phase of apoptosis (1). Intype II cells, proper activation of effector
caspases by Fas depends on an amplificationloop that relies on caspase-8–mediated cleavageof the pro-apoptotic Bcl-2 family member Bidand subsequent release of mitochondrial pro-apoptotic factors [for example, cytochrome cand second mitochondria-derived activator ofcaspases (SMAC, also called Diablo)] to drivethe formation of the caspase-9–activating ap-optosome. Active caspase-9 activates the exe-cutioner caspase-3, which in turn activatescaspase-8 outside the Fas DISC, thereby com-pleting a positive feedback loop (1).
Each step in Fas-mediated apoptosis can be atarget of regulatory mechanisms enabling cells toshow flexible responses to stimulation by Fas.Corresponding to the hierarchy of events in Fas-mediated apoptosis, these regulatory mecha-nisms can be specific for Fas or common todeath receptors, or they can affect the apoptoticcore machinery of the cell. The FasL gene istranscriptionally inactive in most cells. Thus,regulation of FasL expression itself, for example,by the transcription factors nuclear factor kappaB (NF-kB), activating protein 1 (AP1), or nucle-ar factor of activated T cells (NF-AT), regulatesFasL/Fas-mediated effects, such as those of ac-tivation-induced cell death of CD41 T cells (5).To a lesser extent, regulation of Fas expression isalso used to control Fas responses, for example,
Institute of Cell Biology and Immunology, Allman-dring 31, University of Stuttgart, 70 569 Stuttgart,Germany. E-mail: [email protected]
www.sciencemag.org SCIENCE VOL 296 31 MAY 2002 1635
M A P P I N G C E L L U L A R S I G N A L I N G