running head: genetic control of border-like cell release ... · 34 rucha karve, frank...
TRANSCRIPT

Running Head: Genetic control of border-like cell release 1 Corresponding author: Anjali S. Iyer-Pascuzzi 2
915 W. State Street, West Lafayette, IN 47907 3 765-494-1443 4 [email protected] 5
Research Area: Genes, Development and Evolution 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
25 26 27 28 29 30 31
Plant Physiology Preview. Published on May 24, 2016, as DOI:10.1104/pp.16.00453
Copyright 2016 by the American Society of Plant Biologists
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

2
The transcription factor NIN-LIKE PROTEIN 7 (NLP7) controls border-like cell release in 32 Arabidopsis 33 Rucha Karve, Frank Suárez-Román, and Anjali S. Iyer-Pascuzzi 34 Purdue University, Department of Botany and Plant Pathology, 915 W. State Street, West 35 Lafayette, IN 47907 36 37 SUMMARY 38 A transcription factor regulates the release of border-like cells from the root cap by repressing 39 the expression of cell wall loosening enzymes. 40 41 Author contributions: RK performed most of the experiments, FS-R provided technical 42 assistance; RK and ASI-P designed the experiments and analyzed the data, ASI-P supervised the 43 experiments and wrote the article with input from all authors. 44 45 46 47 48 49 50 51 52 53 54 Financial Source: This work was funded by Purdue University. 55 Corresponding Author: Anjali S. Iyer-Pascuzzi, [email protected] 56 57 58 59 60 61 62
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

3
ABSTRACT 63 The root cap covers the tip of the root and functions to protect the root from 64
environmental stress. Cells in the last layer of the root cap are known as border cells, or border-65 like cells (BLCs) in Arabidopsis (Arabidopsis thaliana). These cells separate from the rest of the 66 root cap and are released from its edge as a layer of living cells. BLC release is developmentally 67 regulated, but the mechanism is largely unknown. Here we show that the transcription factor 68 NIN-LIKE PROTEIN 7 (NLP7) is required for the proper release of BLCs in Arabidopsis. 69 Mutations in NLP7 lead to BLCs that are released as single cells instead of an entire layer. NLP7 70 is highly expressed in BLCs, and is activated by exposure to low pH, a condition that causes 71 BLCs to be released as single cells. Mutations in NLP7 lead to decreased levels of cellulose and 72 pectin. Cell wall loosening enzymes such as CELLULASE5 (CEL5), and a pectin lyase-like gene, 73 as well as the root cap regulators SOMBRERO and BEARSKIN1/2, are activated in nlp7-1 74 seedlings. Double mutant analysis revealed that the nlp7-1 phenotype depends on the expression 75 level of CEL5. Mutations in NLP7 lead to an increase in susceptibility to a root-infecting fungal 76 pathogen. Together, these data suggest that NLP7 controls the release of BLCs by acting through 77 the cell wall loosening enzyme CEL5. 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

4
INTRODUCTION 94 The root cap surrounds the root tips of nearly all angiosperms, gymnosperms, and 95
pteridophytes (Barlow, 2003), and is fundamental to root-environment interactions. This tissue is 96 critical to plant health as it protects the delicate root meristem (Barlow, 2003), senses gravity 97 (Blancaflor et al., 1998, 1999), and water (Eapen et al., 2005), and secretes antimicrobial 98 compounds and secondary metabolites that defend the root against soil-borne pathogens and 99 abiotic stress (Hawes et al., 1998; Hawes et al., 2000; Vicre et al., 2005; Driouich et al., 2013; 100 Watson et al., 2015). The root cap is composed of two cell types that surround the meristem – 101 the lateral root cap, which flanks the meristem, and the columella root cap located at the root tip 102 (Figure 1). 103
In Arabidopsis, the columella is composed of five to six layers of rectangular cells 104 (Benfey and Scheres, 2000) (Figure 1). New columella cells are produced from columella stem 105 cells, and as they age they progressively differentiate from amyloplast-filled cells that sense 106 gravity to mucilage-secreting cells that separate from the rest of the root cap and are released 107 from its edges (Vicre et al., 2005; Durand et al., 2009; Bennett et al., 2010; Driouich et al., 2013; 108 Bennett et al., 2014). In Arabidopsis, cells of the columella root cap are released as a layer of 109 living cells that are attached to each other as well as to one or two lateral root cap cells (Figure 1, 110 Vicre et al., 2005; Durand et al., 2009; Kumpf and Nowack, 2015). Together, the columella and 111 lateral root cap cells that are released in this last layer are called ‘border-like cells’ (BLCs) 112 (Vicre et al., 2005; Kumpf and Nowack, 2015). Despite their simultaneous release and physical 113 attachment, the mechanism of release for columella BLCs and lateral root cap BLCs is very 114 different. BLCs of the lateral root cap transition to programmed cell death at the time of their 115 release (Fendrych et al., 2014). In contrast, BLCs of the columella do not undergo programmed 116 cell death (Fendrych et al., 2014; Kumpf and Nowack, 2015) and remain alive for at least 72 117 hours after release (Plancot et al., 2013). How columella BLC release occurs is not well 118 understood. 119
Border cells and BLCs secrete root exudates including antimicrobial compounds, and are 120 an important part of the root’s defense against both biotic and abiotic rhizosphere stresses 121 (Hawes et al., 1998; 2000; Vicre et al., 2005; Driouich et al., 2013; Plancot et al., 2013; Watson 122 et al., 2015). Both border cell and BLC release require cell wall degrading enzymes (Hawes and 123 Lin, 1990; Wen et al., 1999; Vicre et al., 2005; Durand et al., 2009). In Arabidopsis, BLC release 124
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

5
occurs through the action of enzymes that modify components of the cell wall, including 125 cellulases and pectin methylesterases (Bouton et al., 2002; del Campillo et al., 2004; Durand et 126 al., 2009). Mutants deficient in cell wall homogalacturonan (HG), a major component of pectin 127 (Wolf et al., 2009), release BLCs as single cells instead of an intact layer (Bouton et al., 2002; 128 Durand et al., 2009). BLC release also depends on the proper degradation of cellulose, as a 129 mutant defective in CELLULASE5 (CEL5) has a sticky root cap, with extra root cap cells stuck 130 to the root tip (del Campillo et al., 2004). Further, a triple mutant defective in the transcription 131 factors SOMBRERO (SMB), BEARSKIN1 (BRN1) and BRN2 has an extremely sticky root cap 132 with masses of cells remaining attached, and has reduced CEL5 expression (Bennett et al., 2010). 133 Although cell wall loosening enzymes are known to be necessary for BLC release, how their 134 expression is controlled to ensure release of an intact layer of BLCs is not clear. 135 Here we show that the transcription factor NIN-LIKE PROTEIN 7 (NLP7) is required for 136 the release of an intact layer of BLCs in Arabidopsis (Arabidopsis thaliana). Low pH stress 137 causes the release of BLCs as single cells from the root tip of wild type (WT) plants, and a 138 mutation in NLP7 significantly enhances this single cell release in both standard pH (pH 5.7) and 139 low pH (pH 4.0) conditions. NLP7 encodes an Arabidopsis homolog of the Nodule INception 140 (NIN) transcription factor from Lotus japonicas, and has been previously described in 141 Arabidopsis for its role in nitrate signaling (Castaings et al., 2009; Marchive et al., 2013). NLP7 142 expression is activated by low pH conditions and is highly expressed in BLCs. The root of the 143 nlp7 mutant shows a decrease in cellulose and pectin content. Gene expression of SMB and 144 BRN1/2 is activated in the mutant, as is the expression of several cell wall loosening enzymes, 145 such as CEL5, XYLOGLUCAN ENDOTRANSGLUCOSYLASE (XTH5), and a PECTIN LYASE-146 like gene (PL). Double mutant analysis reveals that the nlp7 phenotype depends on the 147 expression level of CEL5. Consistent with a role for BLCs in biotic stress, mutations in NLP7 148 lead to increased susceptibility to the soil-borne fungus Fusarium oxysporum f. sp. conglutinans 149 (Foc). Together our data show that NLP7 expression maintains pectin and cellulose levels in the 150 root, and represses the expression of CEL5, XTH5, PL, SMB, and BRN1/2, thereby preventing 151 single cell border-like cell release. 152 153 Results 154 Low pH stress promotes single cell BLC release in Arabidopsis 155
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

6
We previously found that among five different cell types in the Arabidopsis root, low pH 156 conditions most significantly affected gene expression in the columella root cap (Iyer-Pascuzzi et 157 al., 2011). Given that BCs are important for tolerance to Al toxicity (Li et al. 2000), a major 158 problem in acid soils (Kochian et al., 2015), we reasoned that BLCs may be affected by low pH 159 conditions. Under standard conditions (pH 5.7), at 7 days after imbibition (dai) over 85% of WT 160 Arabidopsis roots release BLCs as intact layers of physically attached cells, and less than 15% of 161 WT roots release BLCs as single cells (Figure 2A, G). In contrast, examination of BLCs in plants 162 grown under low pH stress (pH 4.0) showed that only 45% of WT roots release BLCs as intact 163 layers, while approximately 55% of roots release BLCs as single cells (Figure 2B, G). Low pH 164 stress therefore promotes the release of BLCs as single cells in Arabidopsis. 165 166 NLP7 is required for columella border-like cell adhesion 167
To search for genes that may play a role in BLC release, we examined a time-course of gene 168 expression in whole roots after exposure to low pH (Iyer-Pascuzzi et al., 2011). We reasoned that 169 genes co-expressed with known regulators of the low pH response may be fundamental to low 170 pH – induced phenotypes. K-means clustering of 1848 genes differentially expressed throughout 171 the time-course (Iyer-Pascuzzi et al., 2011) identified one cluster that contained the transcription 172 factor, (TF) Sensitive TO Proton Rhizotoxicity (STOP1). Genes in this cluster were activated 6 - 173 12 hours after exposure to low pH stress (Supplemental Figure 1). Mutations in STOP1 lead to 174 hypersensitivity to acidic pH and severe root growth inhibition under low pH (Iuchi et al., 2007). 175 We hypothesized that the 10 other TFs in this cluster may also play a role in the low pH 176 response. Because we were specifically interested in the BLC phenotype, we examined the root 177 cap in mutant lines for each of these 10 TFs. 178
One of these lines (SALK_026134), with a T-DNA insertion in NIN-Like Protein 7 (NLP7), 179 showed defective BLC release (Figure 2C, D, G). Quantitative RT-PCR (qRT-PCR) confirmed 180 that NLP7 was activated by low pH (Supplemental Figure 2). Examination of the BLCs in 181 SALK_026134 (hereafter nlp7-1) seedlings showed that under standard pH, approximately 44% 182 of roots released BLCs as single cells, compared to just 12% in WT plants and 19% in roots of 183 the complemented line pNLP7:NLP7:GFP/nlp7-1 (Figure 2G). Under low pH conditions, which 184 loosen cells in the root cap and promote single cell BLC release, over 75% of nlp7-1 mutant 185 roots released BLC as single cells, compared to 55% of WT plants and 42% of 186
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

7
pNLP7:NLP7:GFP/nlp7-1 roots (Figure 2G). These results show that NLP7 is necessary for 187 BLC adhesion. In addition to the border-like cell phenotype, the nlp7-1 plant is smaller than WT, 188 with a shorter root and fewer elongated lateral roots (Supplemental Figure 3). 189
We identified another T-DNA insertion mutant line (SALK_114886) within the last exon of 190 the coding region of NLP7. This allele, nlp7-2, expressed the NLP7 transcript in the root at 85% 191 of WT levels (15% reduction) (Supplemental Figure 4). Because nlp7-2 expressed NLP7 at near 192 WT levels and did not have a defective BLC phenotype, this line was not further pursued. Thus 193 the complemented line, pNLP7:NLP7:GFP/nlp7-1 was used in addition to nlp7-1 for our 194 experiments. 195 196 NLP7 is strongly expressed in BLCs 197
The above results suggested that NLP7 functions in BLC release and may be important for 198 BLC adhesion. To further understand the role of NLP7 in BLC release, we examined the 199 expression of NLP7. Plants expressing pNLP7:GUS showed that NLP7 is strongly expressed in 200 the columella root cap and maturation zone of the root (Castaings et al., 2009) (Figure 3A). 201 Examination of the translational fusion of pNLP7:NLP7:GFP demonstrated that NLP7 is 202 expressed in the BLCs under both standard and low pH conditions (Figure 3B and C, standard 203 pH shown; low pH in Supplemental Figure 5). Previous work showed that NLP7 localizes to 204 both the cytoplasm and nucleus (Castaings et al., 2009; Marchive et al., 2013). We observed 205 NLP7 in the cytoplasm and the nucleus of BLCs, and in the nucleus of cells in the root 206 differentiation zone, in both standard (Figure 3B, C, D) and low pH conditions (Supplemental 207 Figure 5). 208
209 Mutations in NLP7 lead to reduced pectin, homogalacturonan, and cellulose content 210
Because the nlp7-1 mutant had altered BLC release, and BLC release in Arabidopsis 211 depends on cellulose and pectin content (del Campillo et al., 2004; Durand et al., 2009), we 212 analyzed the levels of cell wall polysaccharides in the nlp7-1 mutant. We first stained roots of 213 nlp7-1, WT, and the pNLP7:NLP7:GFP/nlp7-1 complemented line with the histochemical stain 214 ruthenium red, which stains unesterified (acidic) pectin (Sterling, 1970; Sabba and Lulai, 2002). 215 Roots of the nlp7-1 mutant showed less pectin compared to WT at standard pH (Figure 4, top 216 panel). At low pH, pectin levels in the WT root decrease and are similar to those in the nlp7-1 217
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

8
mutant (Figure 4, bottom panel). The pectin content does not further decrease in the root of nlp7-218 1 at low pH, possibly because levels in the mutant are already low (Figure 4). 219
Homogalacturonan (HG), a major component of pectin, is the primary polysaccharide 220 responsible for cell wall adhesion in BLCs of Arabidopsis (Durand et al., 2009). To gain further 221 insight into the pectin deficiency in nlp7-1, we next tested whether HG was altered in the nlp7-1 222 mutant using whole mount immunolocalization with the HG-specific antibodies JIM5 and JIM7; 223 (Knox et al., 1990; Willats et al., 2000; Clausen et al., 2003; Durand et al., 2009). JIM5 and 224 JIM7 antibodies recognize different patterns of methyl esterification on HG (Willats et al., 2000; 225 Clausen et al., 2003). JIM5 binds to both partially methyl-esterified HG and un-esterified HG, 226 while JIM7 binds only to partially methyl-esterified HG but does not bind un-esterified HG. 227 Consistent with our ruthenium red results, the root cap of the nlp7-1 mutant showed significantly 228 less JIM5 and JIM7 antibody labeling compared to WT at standard pH (Figure 5, Supplemental 229 Figure 6 for negative control with no primary antibody). To ensure that the differences in JIM5 230 and JIM7 labeling between WT and nlp7-1 were not due to differences in antibody penetration 231 between the two genotypes, we used a positive control. Anti-CDPK32 (Calcium Dependent 232 Protein Kinase 32) showed no difference in labeling between roots of WT and nlp7-1 233 (Supplemental Figure 6), suggesting that the differences observed between WT and nlp7-1 for 234 JIM5 and JIM7 are indeed the result of decreased pectin in nlp7-1. At low pH, the root tip of 235 nlp7-1 had decreased JIM7 labeling compared to WT, but there was no significant difference in 236 labeling between WT and nlp7-1 with the JIM5 antibody (Supplemental Figure 7). 237
Cellulose, formed by ß-1,4-linked glucan chains, is another major component of plant 238 cell walls. Cellulose degradation is necessary for proper BLC release (del Campillo et al., 2004). 239 We therefore examined cellulose levels in nlp7-1 seedlings grown at standard pH using an acid 240 hydrolysis assay (Dubois et al., 1956). Compared to WT, the nlp7-1 mutant contains 241 significantly less cellulose (Figure 6). Together, these experiments demonstrate that the nlp7-1 242 mutant has decreased levels of the cell wall polysaccharides pectin and cellulose. This suggests 243 that defective BLC release in nlp7-1 is due to defects in cell adhesion. 244
245 Transcriptional profiling of nlp7-1 roots identifies cell wall loosening enzymes 246 Given that BLCs were released as single cells in nlp7-1, we hypothesized that NLP7 may 247 function in cell adhesion in the root cap. We used microarray analysis to investigate whether 248
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

9
NLP7 modulated this process. Transcriptional profiling of whole roots of 5 dai nlp7-1 plants 249 identified 194 genes differentially expressed (FDR < 0.0001, FC > 1.5) compared to WT 250 (Supplemental Table 1). One hundred sixty genes were upregulated, and 34 were repressed in the 251 root of nlp7-1. Significant gene ontology (GO) categories (Provart et al., 2003) for all 252 differentially expressed genes, as well as the upregulated and downregulated genes separately, 253 are listed in Supplemental Table 2. Highly significant GO categories for biological process 254 included transport (P = 7.68 x 10-8), response to abiotic/biotic stimulus (P = 1.33 x 10-6), 255 response to stress (P = 3.99 x 10-6), and cell organization/biogenesis (P = 1.72 x 10-3). For 256 molecular function GO categories, transporter activity (P = 7.35 x 10-4), and DNA/RNA binding 257 (P = 5.60 x 10-4) were most significant. In the cellular component category, the category with the 258 highest significance (other than unknown) was the Golgi apparatus (P = 5.29 x 10-5). This was 259 interesting because cell wall polysaccharides such as hemicelluloses are synthesized in the Golgi 260 and then transported by vesicles to the cell wall. Genes encoding proteins operating in the cell 261 wall were present but not significant (P = 0.059). 262 Previously, Marchive et al., (2013) identified direct transcriptional targets of NLP7 in 263 whole nlp7-1 seedlings that had been resupplied with nitrate for 10 minutes after a period of 264 nitrate starvation. We examined whether any genes differentially expressed in the nlp7-1 root 265 were also direct targets of NLP7 in their work and identified 14 genes (Supplemental Table 3). 266 This number is not higher likely because of the differences in growth and environmental 267 conditions in the two datasets: 13 day old whole nlp7-1 seedlings grown in liquid medium under 268 N starvation followed by resupply in Marchive et al., (2013), and 5 day old nlp7-1 roots grown 269 on agar with full nutrient supply in our study. Several of the genes that overlapped between the 270 two datasets had roles in nutrient transport (Supplemental Table 3). 271 Because of our interest in the nlp7-1 border-like cell release phenotype, we focused on 272 cell wall-related processes, and identified 11 genes related to the cell wall in our dataset. These 273 included CELLULASE5 (CEL5), XYLOGLUCAN ENDOTRANSGLUCOSYLASE/ 274 HYDROLASE5 (XTH5), 2 exostosins, and a PECTIN LYASE-like gene (PL) (Supplemental Table 275 4). All but one of the 11 genes was upregulated in nlp7-1. 276
We next used the Arabidopsis root expression map (Brady et al., 2007) to examine the cell 277 type specific expression pattern of these 11 genes. Due to the defective root cap phenotype, we 278 looked for genes highly expressed in the columella and/or lateral root cap. Two genes, CEL5 279
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

10
(At1g22880) and PL (At1g65570), were highly expressed in the columella root cap 280 (Supplemental Figure 8), while another, XTH5 (At5g13870), was highly expressed in the lateral 281 root cap. 282
We used qRT-PCR to confirm the expression of these three genes in nlp7-1 (Figure 7). We 283 also examined expression of QUASIMODO1 (QUA1) and QUA2. Although these genes were not 284 identified in our microarray analysis, QUA1 and QUA2 encode proteins that control pectin 285 biosynthesis and are important for proper border-like cell release (Bouton et al., 2002; Mouille et 286 al., 2007; Durand et al., 2009). As shown in Figure 7A, consistent with our microarray results, 287 CEL5, PL, and XTH5 expression were upregulated in the nlp7-1 root, while QUA1 and QUA2 288 remained unchanged. 289
We next used mutant analysis to determine whether mutations in CEL5, XTH5, or PL altered 290 border-like cell release. Mutations in each of these genes lead to increased root cap layers 291 (Figure 7B). The cel5 mutant had the ‘stickiest’ root cap, and we focused on this gene for further 292 analysis. Mutations in CEL5 have been previously shown to lead to defective border-like cell 293 release in Arabidopsis under standard growth conditions (del Campillo et al., 2004). CEL5 294 encodes an endo-1,4-ß-D-glucanase that hydrolyzes the ß-1,4-linked glucan chains that comprise 295 cellulose, and CEL5 expression is highly enriched in the columella compared to other cell types 296 within the root (Supplemental Figure 8) (del Campillo et al., 2004). As previously observed (del 297 Campillo et al., 2004), analysis of the cel5 mutant (T-DNA insertion line SALK_079921), 298 showed that BLC are not properly released and extra cells stick to the root cap when grown 299 under standard conditions (del Campillo et al., 2004). We searched for additional cel5 alleles, but 300 found no other lines with significantly reduced CEL5 transcript (Supplemental Figure 9). Since 301 WT plants show an increase in release of single BLCs when grown under low pH conditions 302 (Figure 2), we used confocal microscopy to examine the root cap of the cel5 mutant after 303 exposure to low pH. In contrast to WT, when the cel5 mutant was grown at low pH, the root cap 304 retained its ‘stickiness’ - BLCs adhered to the tip of the cel5 root, and intact root cap layers were 305 clearly observed adjacent to the root tip (Figure 7C, D). Only 14% of cel5 roots showed single 306 border-like cell release at low pH compared to 55% of WT plants at low pH. Thus even in a ‘root 307 cap loosening’ environment, cells of the cel5 root cap do not correctly detach. 308
309 Single border-like cell release in nlp7-1 depends on the expression level of CEL5 310
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

11
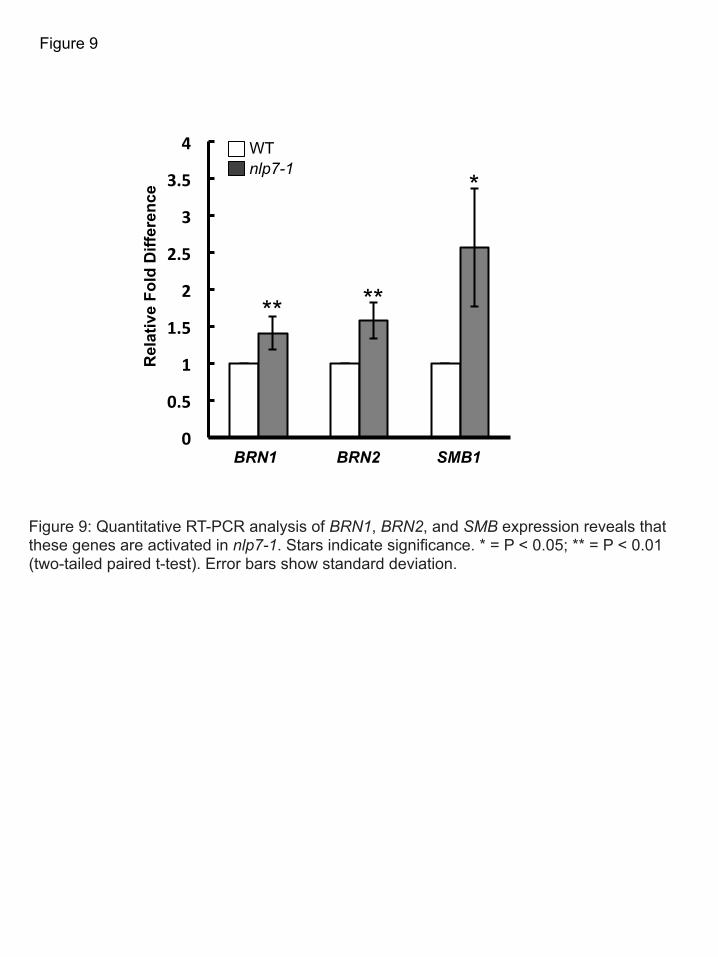
Because NLP7 encodes a transcription factor, and expression of CEL5 is activated in the nlp7-1 311 mutant, we hypothesized that CEL5 may be required for the nlp7-1 phenotype. To test this, we 312 generated the nlp7-1 cel5 double mutant. In roots of the double mutant, BLC release occurs in 313 layers, similar to WT plants (Figure 8A), and the average number of root cap layers is not 314 different compared to WT (Figure 8B). Out of 13 double mutant lines examined, none showed 315 single BLC release. Thus, the single BLC release phenotype observed in nlp7-1 depends on the 316 expression level of CEL5. 317 318 Expression of root cap regulators is altered in nlp7-1 319 The NAC-domain family transcription factors SMB, BRN1, and BRN2 redundantly regulate root 320 cap maturation and cell wall modifications in the root cap (Willemsen et al., 2008; Bennett et al., 321 2010). The smb-3 brn1-1 brn2-1 triple mutant has a very ‘sticky’ root cap, similar to that of cel5 322 (del Campillo et al., 2004; Bennett et al., 2010). We used qRT-PCR to assess expression levels 323 of these genes in the root of the nlp7-1 mutant. We found that expression of SMB, BRN1, and 324 BRN2 is activated (Figure 9). Expression of CEL5 is reduced in the smb-3 brn1-1 brn2-1 triple 325 mutant (Bennett et al., 2010). Thus in mutants with a ‘sticky’ columella root cap, CEL5 326 expression is reduced, while in the nlp7-1 mutant, which releases BLCs as single cells, CEL5 327 expression is activated. 328 329 Mutations in NLP7 lead to increased susceptibility to the soil-borne fungus Fusarium oxysporum 330 f. sp. conglutinans (Foc) 331 We hypothesized that the impaired border-like cell release in the nlp7-1 mutant may lead 332 to increased susceptibility to soil-borne pathogens. To test this, we inoculated the soil-borne 333 fungal pathogen Foc directly on to the tip of nlp7-1 and WT roots. Foc first invades root systems, 334 then enters the vasculature and eventually causes wilting of the aboveground portion of the plant 335 (Tjamos and Beckman, 1989). As shown in Figure 10A and B, nlp7-1 has more Foc growth 336 around the root tips than WT or the complementation line (Supplemental Figure 10) due to 337 increased susceptibility to Foc. Also, fewer nlp7-1 roots grew through the Foc inoculation site, 338 (Figure 10C). Further, nlp7-1 showed increased anthocyanin accumulation in the shoots 339 compared to WT at 5 days after infection (Figure 10D). Thus, in keeping with a role for BLCs in 340 plant defense, mutations in NLP7 lead to increased susceptibility to a root-invading pathogen. 341
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

12
342 DISCUSSION 343
Collectively, our data support a model in which the transcription factor NLP7 regulates BLC 344 release by suppressing expression of cell wall loosening enzymes as well as key transcription 345 factors involved in root cap maintenance and maturation (Figure 11). This promotes cellulose 346 and pectin maintenance, preventing single-cell BLC release in WT plants. Whether NLP7 347 directly regulates these genes, or does so indirectly through the transcription factors SMB, BRN1 348 or BRN2, is not clear. 349
In WT plants, environmental conditions that lead to cell wall loosening, such as those 350 encountered under low pH, result in an increase in NLP7 expression. Increased NLP7 expression 351 suppresses cell wall loosening by repressing expression of cell wall modification enzymes, 352 which results in proper cell adhesion under stressful conditions. In the nlp7-1 mutant, an increase 353 in expression of cell wall modification enzymes in the root cap coupled with a decrease in HG 354 and cellulose leads to release of BLCs as single cells. 355
Border cells and BLCs are known for their protective roles in plant defense (Hawes et al. 356 2000; Watson et al., 2015). For example, border cells in pea protect the root tip from infection by 357 the pathogenic fungus Nectria haematococca (Gunawardena and Hawes, 2002), while those of 358 maize exude a compound that promotes branching in the arbuscular mycorrhizal fungus 359 Gigaspora gigantean (Nagahashi and Douds, 2004). In Medicago truncatula, border cells have 360 elevated levels of defense compounds (Watson et al., 2015). In Arabidopsis, BLCs perceive and 361 activate defense signaling in response to microbe-associated molecular patterns, such as 362 flagellin22 and peptidoglycan (Plancot et al., 2013). Consistent with a role for BLCs in plant 363 defense mechanisms, mutations in NLP7 lead to an increase in susceptibility to the soil-borne, 364 root-infecting fungal pathogen Foc. The increased release of single BLCs from the root cap and 365 the decrease in pectin and cellulose content may facilitate fungal entrance and increase 366 colonization of the mutant root. Given the ubiquity of soil-borne pathogens (Lewis and 367 Papavizas, 1991; Lumsden et al., 1995), this suggests that NLP7 may act to protect the root from 368 biotic stress. 369
The low pH-induced release of BLCs as single cells may be a result of changes to the 370 cell wall polysaccharide pectin (Koyama et al., 2001) (Figure 4). Low pH changes the 371 availability of nutrients in growth media and soils, causing a major decrease in the availability of 372
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

13
Ca++ (Truog, 1946). Arabidopsis roots grown in minimal media at low pH had decreased viability 373 after 2 hours, but this effect was ameliorated if the minimal media was supplemented with Ca++ 374 (Koyama et al., 2001). Ca++ is a divalent cation with a large ionic radius that can cross-link and 375 stabilize pectic polysaccharides (Carpita and Gibeaut, 1993). Although our plants are grown on 376 full nutrient agar, since low pH decreases the availability of Ca++, the low pH-induced release of 377 single BLCs in WT plants may be due to a decrease in available Ca++, and thus a decrease in 378 pectin stabilization. This phenotype is exacerbated at low pH in the nlp7-1 mutant, in which the 379 expression of cell wall loosening enzymes, including PL, is activated. 380
NLP7 is part of the RWP-RK transcription factor family, so named for the conserved 381 RWPYRK protein motif present in all members (Schauser et al., 2005; Chardin et al., 2014). 382 RWP-RK proteins have been best described for their roles in nitrogen (N) regulated pathways 383 (Ferris and Goodenough, 1997; Schauser et al. 1999; Borisov et al. 2003; Schauser et al. 2005; 384 Camargo et al., 2007; Lin and Goodenough, 2007; Marsh et al., 2007). In Arabidopsis, NLPs, 385 including NLP7, are “master regulators” of the nitrate response (Castaings et al., 2009; Konishi 386 and Yanagisawa, 2013; Marchive et al., 2013; Chardin et al., 2014). Members of this family bind 387 to a nitrogen response cis-element to control nitrate-regulated transcription (Konishi and 388 Yanagisawa, 2013). The NLP7 protein moves from the cytoplasm to the nucleus after resupply 389 of nitrate to N-starved seedlings (Marchive et al., 2013), and nitrate sensing is defective in the 390 nlp7 mutant (Castaings et al., 2009). In addition to its role in the N response, nlp7 mutants have 391 high drought tolerance (Castaings et al., 2009), suggesting that this transcription factor has 392 additional unexplored roles in environmental stress responses. 393
Here we show that NLP7 controls BLC release by regulating cell wall loosening in the root 394 cap. The role of NLP7 in BLC release appears to be primarily through suppression of the 395 enzymes that control cellulose and pectin degradation. For example, a pectin-lyase like gene, 396 which degrades pectin, is upregulated in the nlp7-1 mutant, but there is no change in expression 397 of the pectin biosynthetic enzymes QUA1 and QUA2. Interestingly, cell wall loosening and 398 degradation is an important part of the nodulation process, and recent work has demonstrated 399 that NIN, a NLP7 homolog, binds to the promoter and activates the expression of a pectate lyase 400 required for nodulation in L. japonicus (Xie et al., 2012). Together, these results suggest that the 401 regulation of cell wall modification may be a common feature of NIN and NLPs, yet employed 402 in different developmental contexts. It will be intriguing to determine whether additional 403
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

14
members of the NLP family have roles in BLC release or cell wall modification in different 404 tissues. 405 406 MATERIALS and METHODS 407 Plant material and growth conditions 408 Seeds of Arabidopsis thaliana wild type (COL-0), nlp7-1 (Salk_026134), nlp7-2 409 (SALK_114886), cel5 (Salk_079921), xth5 (SALK_057512), and a pectin lyase-like gene 410 (GK100C05) were obtained from the Arabidopsis Biological Resource Center stock center 411 (ABRC, Columbus, OH). Homozygous nlp7-1 plants were backcrossed once to Col-0. The seeds 412 were surface sterilized in 50% bleach and stratified at 4°C for 48 h. The seeds were plated on 413 standard Murashige and Skoog (MS; pH = 5.7) or on low pH MS (pH = 4.0) plates. The 414 seedlings were grown in a growth chamber at 22°C under 50% relative humidity with a 16 hour/ 415 8 hour day/night schedule. Unless otherwise noted, plants in all experiments were examined 7 416 days after imbibition, and error bars in figures show standard deviation. 417 To generate complementation lines, 3kb upstream of the NLP7 start site (“NLP7 418 promoter”) was first cloned into vector pDONRP4-P1 using Gateway technology (Life 419 Technologies, Carlsbad, CA) to generate pENTR:pNLP7. The coding region of At4g24020 420 (NLP7) was amplified from cDNA and cloned into Gateway vector pDONR221 to generate 421 pENTR:NLP7. The vectors pENTR:pNLP7, pENTR:NLP7, and GFP in pDONRP2R-P3 were 422 recombined with pDESTR4-R3 using Multisite Gateway cloning technology (Life Technologies, 423 Carlsbad, CA ). The final construct of pNLP7:NLP7:GFP was transformed into Arabidopsis by 424 the floral dip method (Clough and Bent, 1998). Transgenic plants were confirmed by PCR using 425 GFP specific primers, confocal microscopy for GFP expression, and qRT-PCR for NLP7 426 expression. The seeds of pNLP7:GUS from (Castaings et al., 2009) were obtained as a gift from 427 Dr. Anne Krapp (INRA, France). 428 429 RNA Isolation, cDNA synthesis and quantitative real time PCR 430 For RNA extraction, seedlings were either grown continuously on control MS plates (pH = 5.7) 431 or transferred to low pH 4.0 plates for 24 hours after initial growth on control MS as specified 432 for each experiment. Whole seedlings or only roots were frozen in liquid N2 and stored at -80°C 433 until further use at the time specified for each experiment. Total RNA was isolated using plant 434
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

15
RNA purification kit (Norgen Biotek Corp, Ontario, Canada). On column DNase treatment was 435 performed with DNAse I in DNase digestion buffer (Omega bio-tek, Norcross, GA). The cDNA 436 was synthesized using 1μg of total RNA with oligo (dT) primers using AMV first strands cDNA 437 synthesis kit (New England Biolabs, Boston, MA) according to the manufacturers’ protocol. The 438 cDNA was diluted 1:5 (v/v) and 2 µl of diluted cDNA was used for quantitative RT-PCR (qRT-439 PCR). The qRT-PCR was performed using Sso Fast Eva Green Supermix (Bio-Rad, Hercules, 440 CA) according to the manufacturer’s instructions. The efficiency of each primer pair set was 441 evaluated using a cDNA dilution series and the relative expression was calculated using the 442 delta-delta Ct method. Gene expression was calculated relative to AT1G13320 (Czechowski et 443 al., 2004) as described in (Iyer-Pascuzzi et al., 2011). A total of three to five independent 444 biological replicates, each with three technical replicates, were performed to calculate relative 445 fold difference in gene expression. Primers used are listed in Supplemental Table 5. In all qRT-446 PCR figures, error bars show standard deviation. 447 448 Microarray sample preparation and analysis 449 Sample preparation and microarray analysis was performed as in (Iyer-Pascuzzi et al., 2011). 450 Briefly, seedlings of WT (Col-0) and nlp7-1 were grown on MS pH 5.7, for 5 days and roots cut 451 just below the root-hypocotyl junction. Cut roots were immediately placed into RLT buffer 452 (Qiagen) and RNA extracted using the RNeasy plant mini kit (Qiagen). Two biological replicates 453 were performed. Probes for array analysis were prepared using the one-cycle amplification 454 protocol from Affymetrix. Samples were hybridized to ATH1 microarrays by Expression 455 Analysis Inc (Durham, NC). All arrays were normalized and differentially expressed probesets 456 identified using a mixed-model ANOVA Perl script as in (Levesque et al., 2006). Differentially 457 expressed probesets were identified using an |1.5| fold change cut-off and a FDR (q-value) of 1 x 458 10-4 as in (Iyer-Pascuzzi et al., 2011). Heat maps were created using TMV microarray software 459 (www.tm4.org). 460 461 Gus and Ruthenium Red Staining 462 The seedlings expressing pNLP7:GUS were grown on control MS plates (pH = 5.7) for 6-8 d. 463 The seedlings were transferred to either low pH plates (pH 4.0) or to control plates (pH 5.7). 464 After 24 hours, seedlings were incubated in GUS staining buffer containing; 50 mM NaPO4, 465
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

16
pH7.0, 2.5 mM K3CN6, 2.5 mM K4CN6, 0.1%, Triton X-100 and 2.5 mM X-gluc for 1.5h at 466 30°C, and washed with the GUS staining buffer without X-gluc. To analyze pectin content in the 467 roots of WT and nlp7-1 seedlings, 0.05% (w/v) solution of Ruthenium red (Sigma-Aldrich, St 468 Louise, MO) was prepared and seedlings were stained for 10 min as described by (Durand et al., 469 2009). The pictures were taken with Nikon Eclipse 800 under 20X magnification. 470 471 Confocal microscopy and immunofluorescence 472 Seeds of WT (Col-0), nlp7-1, cel5, and nlp7-1 cel5, xth5, and pl were sterilized as described and 473 seedlings were grown for 7 d. Seedlings were stained with propidium iodide (Sigma-Aldrich, St 474 Louise, MO) for about 1 min, mounted on a glass slide, and roots caps were visualized under a 475 confocal microscope. For all the experiments, images were taken using Nikon A1Rsi confocal 476 microscope and images were edited using NIS elements software (Nikon, Tokyo, Japan). 477
For immunofluorescence experiments, 7 dai WT and nlp7-1 seedlings were fixed in 4% 478 N-N dimethylformamide for 30 min followed by three washes with 1X Phosphate Buffer Saline 479 (PBS). To prevent non-specific antibody binding, seedlings were incubated in blocking solution 480 (3% non- fat dairy milk in 1X PBS) for 30 min followed by three washes with 1X PBS. The 481 seedlings were incubated overnight with primary antibody JIM5 or JIM7 (CarboSource, Athens, 482 GA) diluted 1:6 in 1X PBS with 3% non-fat dairy milk in 1X PBS, washed with TPBS (0.05% 483 Tween in 1X PBS) for 20 min with 4X five min washes, and then incubated with FITC-484 conjugated secondary antibody diluted 1:50 in TPBS for 1h at room temperature. Finally, the 485 seedlings were washed 4X with TPBS for a total of 20 min followed by a last wash with PBS. 486 Seedlings were left in PBS until confocal microscopy was done. A ‘no primary antibody’ control 487 was used in each biological replicate as a negative control to check for specificity of antibodies. 488 As a positive control, both WT and nlp7-1 roots were labelled with anti-Calcium Dependent 489 Protein Kinase 32 (anti-CDPK32, custom made from Yenzym Antibodies, LLC, California). The 490 images of roots were taken using Nikon confocal A1Rsi with Z-stacks sectioning and a 3D 491 image was reconstructed using NIS elements. Two independent biological replicates were 492 performed. Z-stack projections for WT and nlp7-1 were composed of similar numbers of slices, 493 and each slice was 5 μm. For fluorescence measurements, the maximum intensity projections 494 were generated using NIS elements ND2 viewer, and fluorescence intensities were measured 495 using ImageJ software. For measuring fluorescence, the area between the root tip and 200 μm 496
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

17
upwards was selected in each root and the outline for the area of the root was drawn in Image J. 497 The corrected total fluorescence (CTF) was calculated as CTF = Integrated Density - (Area of 498 selected root tip X Mean fluorescence of background readings). Average of at least nine roots 499 with similar number of Z-stack sections was used for each WT and nlp7-1 for each antibody. 500 Error bars show standard deviation. 501 502 Quantification of root cap layers 503 Root cap layers were counted from the root cap images taken using a confocal microscope. 504 Number of root cap layers were counted from the columella initials to the last attached layer of 505 BLCs still adhered to the primary root. A minimum of 30 roots was examined per genotype with 506 at least two biological replicates. 507 508 Cellulose assay 509 Cellulose content in the nlp7-1 mutant and WT was determined by the acid hydrolysis method 510 modified from (Dubois et al., 1956). Briefly, the roots of 7 dai nlp7-1 and WT were ground in 511 liquid N2 and 3.5 mg each of the freeze dried powder was used for further analysis. The ground 512 tissue was treated with trifluroacetic acid (TFA) for 90 min at 1200C. To determine the cellulose 513 content as a glucose derivative, the Phenol-Sulphuric Acid hydrolysis method was performed 514 and the glucose concentration was determined using a standard curve of 0-1000 nmols of glucose 515 at 500 nm. The entire experiment was repeated twice. Error bars show standard deviation. 516 517 Pathogen assay 518 nlp7-1 and WT plants were grown on standard MS plates for 7 days. The root tips of nlp7-1 and 519 WT were inoculated with 2 µl of the 1X108 microconidia suspension in sterile water of Fusarium 520 oxysporum f. sp. conglutinans (Foc; NRRL #38297). Foc cultures were grown on Potato 521 Dextrose Agar plates for 3 - 4 days to obtain conidia. The plates inoculated with Foc were 522 scanned at 24 and 48 hours post inoculation (hpi). Mycelial growth was measured at 48 hpi using 523 Image J. The area of mycelial growth around each root of WT and nlp7-1 was selected in image 524 J and measured. Root growth inhibition was measured by counting the number of roots passing 525 through the infection site. Anthocyanin concentrations were measured by standard anthocyanin 526 spectrophotometric assay as described by (Sims and Gamon, 2002). Briefly, shoot tissue of the 527
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

18
infected plants was harvested in liquid N2 and ground in acidified methanol (99% Methanol and 528 1% HCl). The extracts were centrifuged at 4000g for 5 min at 4ºC and the supernatant was used 529 for spectrophotometry. Absorbance for anthocyanin was measured at 530 nm. The total 530 anthocyanin content was calculated by accounting for degraded chlorophyll content. 531 532 Accession Numbers 533 Sequence data from this article can be found in the EMBL/GenBank data libraries under 534 accession number(s) GSE63474. The reviewer access link can be found here: 535 http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?token=klqroscqxfwbfgx&acc=GSE63474. 536 537 SUPPLEMENTAL DATA 538 Supplemental Figure 1: Time-course of gene expression of genes in the STOP1 cluster after 539 exposure to pH 4.6. 540 Supplemental Figure 2: qRT-PCR of NLP7 expression in whole roots 24 hours after exposure to 541 pH 4.0 or pH 5.7 shows that NLP7 is activated by low pH. 542 Supplemental Figure 3: nlp7-1 mutant phenotype in the whole plant. 543 Supplemental Figure 4: Levels of NLP7 transcript in nlp7-1 (SALK_026134) and nlp7-2 544 (SALK_114886) mutants and in the complemented line. 545 Supplemental Figure 5: NLP7 localization at low pH. 546 Supplemental Figure 6: Negative and positive controls for immunolabeling experiment. 547 Supplemental Figure 7: Immunolabeling of 5 dai root tips of WT and nlp7-1 seedlings grown at 548 pH 4.0. 549 Supplemental Figure 8: Heat map of root cell-type specific expression of 11 genes with roles in 550 cell-wall related processes and differentially expressed in nlp7-1 roots 551 compared to WT. 552 Supplemental Figure 9: Semi-quantitative RT-PCR for reduction in CEL5 transcript in two 553 SALK T-DNA insertion lines. 554
Supplemental Figure 10: nlp7-1 and the complementation line (pNLP7:NLP7:GFP) 48 hpi with 555 Foc. 556
557 Supplemental Table 1: 194 genes differentially expressed between whole roots of nlp7-1 and WT 558 Col-0, ATH1 microarray, pH 5.7. 559
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

19
560 Supplemental Table 2: GO categories of the genes differentially expressed in the nlp7-1 root. 561 562 Supplemental Table 3: Genes differentially expressed in the nlp7-1 root and direct targets of 563 NLP7 after nitrate resupply in Marchive et al. 2013. 564 Supplemental Table 4: 11 genes with roles in cell wall related processes and differentially 565 expressed in the nlp7-1 root. 566 Supplemental Table 5: Primers used in this study 567 568 Supplemental Methods 569 570 ACKNOWLEDGEMENTS 571 We thank members of the Iyer-Pascuzzi lab for critical reading of the manuscript, members of 572 the Carpita lab for technical help with the cellulose acid hydrolysis assay, Anne Krapp for use of 573 the pNLP7:GUS line, and Philip Benfey for supporting early experiments with NLP7. This work 574 was supported with Purdue University Start-up funds. 575 576 FIGURE LEGENDS 577 Figure 1: The Arabidopsis root cap. Border-like cells are the last layer at the tip of the root 578 (arrow) and contain cells derived from both columella and lateral root caps. Stars show the living 579 columella border-like cells. White dashed line illustrates where border-like cell separation occurs 580 to release an intact layer of cells. 581 582 Figure 2: Low pH causes single BLC release in Arabidopsis. A and B: WT root grown at 583 standard (A) and low pH (B). C and D: nlp7-1 root grown at standard (C) and low pH (D). E and 584 F: Roots of the complemented line (pNLP7:NLP7:GFP in nlp7-1) grown at standard (E) and low 585 pH (F). No GFP is seen in E and F because the GFP filter was not used for these images. Blue 586 arrows point to BLCs released as single cells (B, C, D) or loose BLC adhesion (F). G: Graph 587 showing the percentage of roots with BLCs released as single cells in WT, nlp7-1 and the NLP7 588 complemented line. Different letters indicate significant differences by analysis of variance 589 (ANOVA) and Tukey’s HSD test. Scale bar indicates 50 μm and is the same for all panels. n > 590 50 for each genotype and each condition. 591
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

20
592 Figure 3: NLP7 expression in root. A) Transgenic plants expressing pNLP7:GUS. NLP7 is 593 expressed in the columella root cap as well as the elongation and maturation zone of the root. 594 Scale bar = 100 μm. 20X magnification. B) and C) nlp7-1 mutant complemented with 595 pNLP7:NLP7:GFP showing that NLP7 is expressed in B) cytoplasm and C) nucleus of the BLCs. 596 D) NLP7 expression in the nucleus of the cells in differentiation zone of root. White arrow 597 indicates the nucleus and the scale bar indicates 50 µm. B, C and D are 40X magnification. 598 599 Figure 4: Mutations in NLP7 lead to a decrease in pectin in roots at both standard and low pH. 600 Ruthenium red staining of WT, nlp7-1, and complemented line (pNLP7:NLP7:GFP/nlp7-1). All 601 roots 7 dai. Scale bar indicates 100 µm. Magnification 20X. The experiment was repeated twice 602 with n > 10 for each genotype and treatment in each replicate. 603 604 Figure 5: Mutations in NLP7 lead to decreased levels of HG in roots. A) and C) WT, B) and D) 605 nlp7-1. Whole-mount immunolabeling of WT and nlp7-1 roots grown at pH 5.7. JIM5 (top 606 panel) and JIM7 (middle panel). E) and F) Average corrected total fluorescence (CTF) for 607 labeling intensity of JIM5 (left) and JIM7 (right). nlp7-1 roots show significantly (P < 0.005) 608 decreased labeling for each. Scale bar indicates 50 μM. Similar numbers of slices per z-stack 609 were used for WT and nlp7-1 for each antibody, with each slice 5 μm (see materials and methods 610 for details). The area extending from the root tip to 200 μM was selected for fluorescence 611 quantification. The experiment was repeated twice and the average of at least 9 seedlings for 612 each genotype and each antibody is shown. Magnification 40X. 613 614 Figure 6: Mutations in NLP7 decrease cellulose content in roots. Acid hydrolysis assay showing 615 decreased cellulose content in 5 dai nlp7-1 roots grown at standard pH. ** = P < 0.001 (two-616 tailed paired t-test). Bars indicate standard deviation for two biological replicates. 617 618 Figure 7: NLP7 regulates border-like cell release by altering expression of cell wall modifying 619 enzymes. A) Relative expression levels of cell wall modifying enzymes in nlp7-1 mutant by 620 qRT-PCR. B) Total number of root cap layers in various mutants involved in cell wall 621 modification. C) and D) BLCs adhere more tightly to the root cap of the cel5 mutant at pH 4.0. 622
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

21
C) 7 dai WT root growing at pH 4.0 showing release of single border-like cells (white arrow). D) 623 7 dai cel5 mutant growing at pH 4.0 showing cells adhering to the root cap (white arrow) and 624 several released root cap layers adjacent to the root cap (white arrowheads). Scale bar indicates 625 50 µm. C and D are 40X magnification. * = P < 0.05 and ** = P < 0.001. 626 627 Figure 8: nlp7-1 mutant phenotype depends on expression of CEL5. A) Arrows point to border-628 like cells or cell layers (in cel5). Border-like cells of the nlp7-1 mutant are released as single 629 cells, while those of the cel5 mutant stick to the root cap. The double mutant phenocopies WT. 630 B) Average number of root cap layers in each genotype in A. Scale bar indicates 50 μm. n > 13 631 for each genotype. 40X magnification. Letters indicate significant differences by ANOVA and 632 Tukey’s HSD test. 633 634 Figure 9: Quantitative RT-PCR analysis of BRN1, BRN2, and SMB expression reveals that these 635 genes are activated in nlp7-1. Stars indicate significance. * = P < 0.05; ** = P < 0.01 (two-tailed 636 paired t-test). Error bars show standard deviation. 637 638 Figure 10: Mutations in NLP7 lead to increased susceptibility to Fusarium oxysporum f. sp 639 conglutinans (Foc) infection. A) Foc growth around the root of root-tip inoculated WT and nlp7-640 1 at 48 hours post inoculation (hpi). B) Quantification of the area of Foc growth around the root 641 tip at 48 hpi. nlp7-1 shows significantly more growth of Foc around the root tip. P < 0.05. C) 642 Percentage of roots growing through the inoculation site. D) Graph showing total anthocyanin 643 content from the shoots of WT and nlp7-1 at 6 days post inoculation. Letters indicate significant 644 difference using ANOVA and Tukey’s HSD test. 645 646 Figure 11: Model for NLP7 control of border-like cell release. See text for details. 647 648 LITERATURE CITED 649 Barlow P (2003) The Root Cap: Cell Dynamics, Cell Differentiation and Cap Function. J Plant 650
Growth Regul 21: 261-186 651 652 Benfey PN, Scheres B (2000) Root development. Curr Biol 10: R813-815 653 654
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

22
Bennett T, van den Toorn A, Sanchez-Perez GF, Campilho A, Willemsen V, Snel B, 655 Scheres B (2010) SOMBRERO, BEARSKIN1, and BEARSKIN2 regulate root cap 656 maturation in Arabidopsis. Plant Cell 22: 640-654 657
658 Bennett T, van den Toorn A, Willemsen V, Scheres B (2014) Precise control of plant stem cell 659
activity through parallel regulatory inputs. Development 141: 4055-4064 660 661 Blancaflor EB, Fasano JM, Gilroy S (1998) Mapping the functional roles of cap cells in the 662
response of Arabidopsis primary roots to gravity. Plant Physiol 116: 213-222 663 664 Blancaflor EB, Fasano JM, Gilroy S (1999) Laser ablation of root cap cells: implications for 665
models of graviperception. Adv Space Res 24: 731-738 666 667 Borisov AY, Madsen LH, Tsyganov VE, Umehara Y, Voroshilova VA, Batagov AO, Sandal 668
N, Mortensen A, Schauser L, Ellis N, et al (2003) The Sym35 gene required for root 669 nodule development in pea is an ortholog of Nin from Lotus japonicus. Plant Physiol 670 131: 1009-1017 671
672 Bouton S, Leboeuf E, Mouille G, Leydecker MT, Talbotec J, Granier F, Lahaye M, Hofte 673
H, Truong HN (2002) QUASIMODO1 encodes a putative membrane-bound 674 glycosyltransferase required for normal pectin synthesis and cell adhesion in Arabidopsis. 675 Plant Cell 14: 2577-2590 676
677 Brady SM, Orlando DA, Lee JY, Wang JY, Koch J, Dinneny JR, Mace D, Ohler U, Benfey 678
PN (2007) A high-resolution root spatiotemporal map reveals dominant expression 679 patterns. Science 318: 801-806 680
681 Camargo A, Llamas A, Schnell RA, Higuera JJ, Gonzalez-Ballester D, Lefebvre PA, 682
Fernandez E, Galvan A (2007) Nitrate signaling by the regulatory gene NIT2 in 683 Chlamydomonas. Plant Cell 19: 3491-3503 684
685 Carpita NC, Gibeaut DM (1993) Structural models of primary cell walls in flowering plants: 686
consistency of molecular structure with the physical properties of the walls during 687 growth. Plant J 3: 1-30 688
689 Castaings L, Camargo A, Pocholle D, Gaudon V, Texier Y, Boutet-Mercey S, Taconnat L, 690
Renou JP, Daniel-Vedele F, Fernandez E, et al (2009) The nodule inception-like 691 protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J 57: 426-435 692
693 Chardin C, Girin T, Roudier F, Meyer C, Krapp A (2014) The plant RWP-RK transcription 694
factors: key regulators of nitrogen responses and of gametophyte development. J Exp Bot 695 65: 5577-5587 696
697 Clausen MH, Willats WG, Knox JP (2003) Synthetic methyl hexagalacturonate hapten 698
inhibitors of anti-homogalacturonan monoclonal antibodies LM7, JIM5 and JIM7. 699 Carbohydr Res 338: 1797-1800 700
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

23
701 Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated 702
transformation of Arabidopsis thaliana. Plant J 16: 735-743 703 704 Czechowski T, Bari RP, Stitt M, Scheible WR, Udvardi MK (2004) Real-time RT-PCR 705
profiling of over 1400 Arabidopsis transcription factors: unprecedented sensitivity 706 reveals novel root- and shoot-specific genes. Plant J 38: 366-379 707
708 del Campillo E, Abdel-Aziz A, Crawford D, Patterson SE (2004) Root cap specific 709
expression of an endo-beta-1,4-D-glucanase (cellulase): a new marker to study root 710 development in Arabidopsis. Plant Mol Biol 56: 309-323 711
712 Driouich A, Follet-Gueye ML, Vicre-Gibouin M, Hawes M (2013) Root border cells and 713
secretions as critical elements in plant host defense. Curr Opin Plant Biol 16: 489-495 714 715 Dubois M, Gilles K, Hamilton J, Rebers P, Smith F (1956) Colorimetric Method for 716
Determination of Sugars and Related Substances. Anal. Chem 28: 350-356 717 718 Durand C, Vicre-Gibouin M, Follet-Gueye ML, Duponchel L, Moreau M, Lerouge P, 719
Driouich A (2009) The organization pattern of root border-like cells of Arabidopsis is 720 dependent on cell wall homogalacturonan. Plant Physiol 150: 1411-1421 721
722 Eapen D, Barroso ML, Ponce G, Campos ME, Cassab GI (2005) Hydrotropism: root growth 723
responses to water. Trends Plant Sci 10: 44-50 724 725 Fendrych M, Van Hautegem T, Van Durme M, Olvera-Carrillo Y, Huysmans M, Karimi 726
M, Lippens S, Guerin CJ, Krebs M, Schumacher K, et al (2014) Programmed cell 727 death controlled by ANAC033/SOMBRERO determines root cap organ size in 728 Arabidopsis. Curr Biol 24: 931-940 729
730 Ferris PJ, Goodenough UW (1997) Mating type in Chlamydomonas is specified by mid, the 731
minus-dominance gene. Genetics 146: 859-869 732 733 Gunawardena U, Hawes MC (2002) Tissue specific localization of root infection by fungal 734
pathogens: role of root border cells. Mol Plant Microbe Interact 15: 1128-1136 735 736 Hawes MC, Brigham LA, Wen F, Woo HH, Zhu Y (1998) Function of root border cells in 737
plant health: pioneers in the rhizosphere. Annu Rev Phytopathol 36: 311-327 738 739 Hawes MC, Gunawardena U, Miyasaka S, Zhao X (2000) The role of root border cells in 740
plant defense. Trends Plant Sci 5: 128-133 741 742 Hawes MC, Lin HJ (1990) Correlation of Pectolytic Enzyme Activity with the Programmed 743
Release of Cells from Root Caps of Pea (Pisum sativum). Plant Physiol 94: 1855-1859 744 745
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

24
Iuchi S, Koyama H, Iuchi A, Kobayashi Y, Kitabayashi S, Kobayashi Y, Ikka T, Hirayama 746 T, Shinozaki K, Kobayashi M (2007) Zinc finger protein STOP1 is critical for proton 747 tolerance in Arabidopsis and coregulates a key gene in aluminum tolerance. Proc Natl 748 Acad Sci USA 104: 9900-9905 749
750 Iyer-Pascuzzi AS, Jackson T, Cui H, Petricka JJ, Busch W, Tsukagoshi H, Benfey PN 751
(2011) Cell identity regulators link development and stress responses in the Arabidopsis 752 root. Dev Cell 21: 770-782 753
754 Knox JP, Linstead PJ, King J, Cooper C, Roberts K (1990) Pectin esterification is spatially 755
regulated both within cell walls and between developing tissues of root apices. Planta 756 181: 512-521 757
758 Kochian LV, Pineros MA, Liu J, Magalhaes JV (2015) Plant adaptation to Acid soils: the 759
molecular basis for crop aluminum resistance. Annu Rev Plant Biol 66: 571-598 760 761 Konishi M, Yanagisawa S (2013) Arabidopsis NIN-like transcription factors have a central role 762
in nitrate signalling. Nat Commun 4: 1617 763 764 Koyama H, Toda T, Hara T (2001) Brief exposure to low-pH stress causes irreversible damage 765
to the growing root in Arabidopsis thaliana: pectin-Ca interaction may play an important 766 role in proton rhizotoxicity. J Exp Bot 52: 361-368 767
768 Kumpf RP, Nowack MK (2015) The root cap: a short story of life and death. J Exp Bot. 769
doi:10.1093/jxb/erv295 770 Levesque MP, Vernoux T, Busch W, Cui H, Wang JY, Blilou I, Hassan H, Nakajima N, 771 Matsumoto N, Lohmann JU, et al (2006) Whole-genome analysis of the SHORT-772 ROOT developmental pathway in Arabidopsis. PLoS Biol 4(5): e143. DOI: 773 10.1371/journal.pbio.0040143 774 Lewis JA, Papavizas GC (1991) Biocontrol of plant disease: the approach for tomorrow, Crop 775 Protection, 10: 95 776 Lumsden RD, Lewis JA, Fravel DR (1995) Formulation and delivery of biocontrol agents for 777 use against soilborne plant pathogens. In F Hall, J Barry eds. Biorational Pest Control 778 Agents, Formation and Delivery. American Chemical Society, Washington DC, pp 166-779 182 780 Li X, Ma J, Hiradate S, Matsumoto H (2000) Mucilage strongly binds aluminum but does not 781
prevent roots from aluminum injury in Zea mays. Physiol Plantarum 108: 152-160 782 783 Lin H, Goodenough UW (2007) Gametogenesis in the Chlamydomonas reinhardtii minus 784
mating type is controlled by two genes, MID and MTD1. Genetics 176: 913-925 785 786
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

25
Marchive C, Roudier F, Castaings L, Brehaut V, Blondet E, Colot V, Meyer C, Krapp A 787 (2013) Nuclear retention of the transcription factor NLP7 orchestrates the early response 788 to nitrate in plants. Nat Commun 4: 1713 789
790 Marsh JF, Rakocevic A, Mitra RM, Brocard L, Sun J, Eschstruth A, Long SR, Schultze M, 791
Ratet P, Oldroyd GE (2007) Medicago truncatula NIN is essential for rhizobial-792 independent nodule organogenesis induced by autoactive calcium/calmodulin-dependent 793 protein kinase. Plant Physiol 144: 324-335 794
795 Mouille G, Ralet MC, Cavelier C, Eland C, Effroy D, Hematy K, McCartney L, Truong 796
HN, Gaudon V, et al (2007) Homogalacturonan synthesis in Arabidopsis thaliana 797 requires a Golgi-localized protein with a putative methyltransferase domain. Plant 798 Journal 50: 605-14 799
800 Nagahashi G, Douds DD, Jr. (2004) Isolated root caps, border cells, and mucilage from host 801
roots stimulate hyphal branching of the arbuscular mycorrhizal fungus, Gigaspora 802 gigantea. Mycol Res 108: 1079-1088 803
804 Plancot B, Santaella C, Jaber R, Kiefer-Meyer MC, Follet-Gueye ML, Leprince J, Gattin I, 805
Souc C, Driouich A, Vicre-Gibouin M (2013) Deciphering the responses of root border-806 like cells of Arabidopsis and flax to pathogen-derived elicitors. Plant Physiol 163: 1584-807 1597 808
809 Provart NJ, Gil P, Chen W, Han B, CHang H-S, Wang X, Zhu T (2003) Gene Expression 810
Phenotypes of Arabidopsis Associated with Sensitivity to Low Temperatures. Plant 811 Physiol 132 (893-906) 812
813 Sabba RP, Lulai EC (2002) Histological analysis of the maturation of native and wound 814
periderm in potato (Solanum tuberosum L.) Tuber. Ann Bot 90: 1-10 815 816 Schauser L, Roussis A, Stiller J, Stougaard J (1999) A plant regulator controlling 817
development of symbiotic root nodules. Nature 402: 191-195 818 819 Schauser L, Wieloch W, Stougaard J (2005) Evolution of NIN-like proteins in Arabidopsis, 820
rice, and Lotus japonicus. J Mol Evol 60: 229-237 821 822 Sims DA, Gamon JA (2002) Relationships between leaf pigment content and spectral 823
reflectance across a wide range of species, leaf structures and developmental stages. 824 Remote Sensing of Environment 81: 337-354 825
826 Sterling C (1970) Crystal-structure of ruthenium red and stereochemistry of its pectic stain. 827
American Journal of Botany 57: 172-175 828 829 Tjamos EC, Beckman CH (1989) Vascular Wilt Diseases of Plants. Berlin: Springer-Verlag. 830 831
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

26
Truog E (1946) Soil Reaction Influence on Availability of Plant Nutrients. Soil Science Society 832 Proceedings: 305 - 308 833
834 Vicre M, Santaella C, Blanchet S, Gateau A, Driouich A (2005) Root border-like cells of 835
Arabidopsis. Microscopical characterization and role in the interaction with rhizobacteria. 836 Plant Physiol 138: 998-1008 837
838 Watson BS, Bedair MF, Urbanczyk-Wochniak E, Huhman DV, Yang DS, Allen SN, Li W, 839
Tang Y, Sumner LW (2015) Integrated metabolomics and transcriptomics reveal 840 enhanced specialized metabolism in Medicago truncatula root border cells. Plant Physiol 841 167: 1699-1716 842
843 Wen F, Zhu Y, Hawes MC (1999) Effect of pectin methylesterase gene expression on pea root 844
development. Plant Cell 11: 1129-1140 845 846 Willats WG, Limberg G, Buchholt HC, van Alebeek GJ, Benen J, Christensen TM, Visser 847
J, Voragen A, Mikkelsen JD, Knox JP (2000) Analysis of pectic epitopes recognised 848 by hybridoma and phage display monoclonal antibodies using defined oligosaccharides, 849 polysaccharides, and enzymatic degradation. Carbohyd Res 327: 309-320 850
851 Willemsen V, Bauch M, Bennett T, Campilho A, Wolkenfelt H, Xu J, Haseloff J, Scheres B 852
(2008) The NAC domain transcription factors FEZ and SOMBRERO control the 853 orientation of cell division plane in Arabidopsis root stem cells. Dev Cell 15: 913-922 854
855 Wolf S, Mouille G, Pelloux J (2009) Homogalacturonan methyl-esterification and plant 856
development. Mol Plant 2: 851-860 857 858 Xie F, Murray JD, Kim J, Heckmann AB, Edwards A, Oldroyd GE, Downie JA (2012) 859
Legume pectate lyase required for root infection by rhizobia. Proc Natl Acad Sci USA 860 109: 633-638 861
862
www.plantphysiol.orgon July 2, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Columella root cap: border-like cells
Columella root cap
Columella stem cells
Lateral Root Cap
Lateral Root Cap: border-like cells *
* **
Figure 1: The Arabidopsis root cap. Border-like cells are the last layer at the tip of the root (arrow) and contain cells derived from both columella and lateral root caps. Stars show the living columella border-like cells. White dashed line illustrates where border-like cell separation occurs to release an intact layer of cells.

Figure 2
nlp7-1
pNLP7: NLP7: GFP/ nlp7-1
Standard pH (5.7) Low pH (4.0)
C
E
A
WT
B
F
D
Figure 2: Low pH causes single BLC release in Arabidopsis. A and B: WT root grown at standard (A) and low pH (B). C and D: nlp7-1 root grown at standard (C) and low pH (D). E and F: Roots of the complemented line (pNLP7:NLP7:GFP in nlp7-1) grown at standard (E) and low pH (F). No GFP is seen in E and F because the GFP filter was not used for these images. Blue arrows point to BLCs released as single cells (B, C, D) or loose BLC adhesion (F). G: Graph showing the percentage of roots with BLCs released as single cells in WT, nlp7-1 and the NLP7 complemented line. Different letters indicate significant differences by analysis of variance (ANOVA) and Tukey’s HSD test. Scale bar indicates 50 µm and is the same for all panels. n > 50 for each genotype and each condition.
G
0
10
20
30
40
50
60
70
80
90
100 pH 4.0
pH 5.7
% o
f roo
ts s
how
ing
sing
le B
LC re
leas
e
B
A
B
B
C C

Figure 3
Figure 3: NLP7 expression in root. A) Transgenic plants expressing pNLP7:GUS. NLP7 is expressed in the columella root cap as well as the elongation and maturation zone of the root. Scale bar = 100 µm. 20X magnification. B) and C) nlp7-1 mutant complemented with pNLP7:NLP7:GFP showing that NLP7 is expressed in B) cytoplasm and C) nucleus of the BLCs. D) NLP7 expression in the nucleus of cells in the differentiation zone of root. White arrow indicates the nucleus and the scale bar indicates 50 µm. B, C, and D are 40X magnification.
A B
DC D

Figure 4
Std pH
Low pH
Figure 4: Mutations in NLP7 lead to a decrease in pectin in roots at both standard and low pH. Ruthenium red staining of WT, nlp7-1 and complementation line (pNLP7:NLP7:GFP/nlp7-1). All roots are 7 dai. Scale bar indicates 100 µm. Magnification 20X. The experiment was repeated twice with n > 10 for each genotype and treatment in each replicate.
nlp7-1 WTpNLP7:NLP7:GFP/
nlp7-1

Figure 5
Aver
age
CTF
(arb
itrar
y un
its)
E
Figure 5: Mutations in NLP7 lead to decreased levels of HG in roots. A) and C) WT, B) and D) nlp7-1. Whole-mount immunolabeling of WT and nlp7-1 roots grown at pH 5.7. JIM5 (top panel) and JIM7 (middle panel). E) and F) Average corrected total fluorescence (CTF) for labeling intensity of JIM5 (left) and JIM7 (right). nlp7-1 roots show significantly (P < 0.005) decreased labeling for each. Scale bar indicates 50 µM. Similar numbers of slices per z-stack were used for WT and nlp7-1 for each antibody, with each slice 5 µm (see materials and methods for details). The area extending from the root tip to 200 µM was selected for fluorescence quantification. The experiment was repeated twice and the average of at least 9 seedlings for each genotype and each antibody is shown. Magnification 40X.
WT nlp7-1
JIM5
JIM7
**
A
C D
B
F
0
10000
20000
30000
40000
50000
3 X 104
4 X 104
5 X 104
2 X 104
1 X 104
**
0
5000
10000
15000
20000
25000
.5 X 104
1 X 104
1.5 X 104
2.5 X 104
2.0 X 104
WT nlp7-1

0
500
1000
1500
2000
2500
3000
3500
4000
nmol
s/m
g of
dry
wt
WT nlp7-1
Figure 6: Mutations in NLP7 decrease cellulose content in roots. Acid hydrolysis assay showing decreased cellulose content in 5 dai nlp7-1 roots grown at standard pH. ** = P < 0.001 (two-tailed t-test). Bars indicate standard deviation for two biological replicates.
**
Figure 6

C
Figure 7
C D
Figure 7: NLP7 regulates border-like cell release by altering expression of cell wall modifying enzymes. A) Graph showing relative expression levels of cell wall modifying enzymes in nlp7-1 mutant by qRT-PCR. B)Total number of root cap layers in various mutants involved in cell wall modification. C) and D) BLCs adhere more tightly to the root cap of the cel5 mutant at pH 4.0. C) 7 dai WT root growing at pH 4.0 showing release of single border-like cells (white arrow). D) 7 dai cel5 mutant growing at pH 4.0 showing cells adhering to the root cap (white arrow) and several released root cap layers adjacent to the root cap (white arrowheads). Scale bar indicates 50 µm. C and D are 40X magnification. * represents P < 0.05 and ** represents P < 0.001.
A
Rel
ativ
e Fo
ld C
hang
e
0
1
2
3
4
5
6
7
CEL5 XTH5 PL QUA1 QUA2
WT nlp7-1
* **
**
B
WT nlp7-1 cel5 xth5 pl
Tota
l num
ber o
f roo
t cap
laye
rs
0
2
4
6
8
10
12**
**
**
*

C
0.0
2.0
4.0
6.0
8.0
10.0
12.0
Aver
age
num
ber o
f roo
t cap
la
yers
WT nlp7-1 cel5 cel5 nlp7-1 A
B
Figure 8: nlp7-1 mutant phenotype depends on expression of CEL5. A) Arrows point to border-like cells or cell layers (in cel5). Border-like cells of the nlp7-1 mutant are released as single cells, while those of the cel5 mutant stick to the root cap. The double mutant phenocopies WT. B) Average number of root cap layers in each genotype in A. Scale bar indicates 50 µm. n > 13 for each genotype. 40X magnification. Letters indicate significant differences by ANOVA and Tukey’s HSD test.
Figure 8
B B
A
C
0
0.5
1
1.5
2
2.5
3
3.5
4
BRN1 BRN2 SMB1
Rel
ativ
e Fo
ld D
iffer
ence
Figure 9: Quantitative RT-PCR analysis of BRN1, BRN2, and SMB expression reveals that these genes are activated in nlp7-1. Stars indicate significance. * = P < 0.05; ** = P < 0.01 (two-tailed paired t-test). Error bars show standard deviation.
Figure 9
** **
* WT nlp7-1
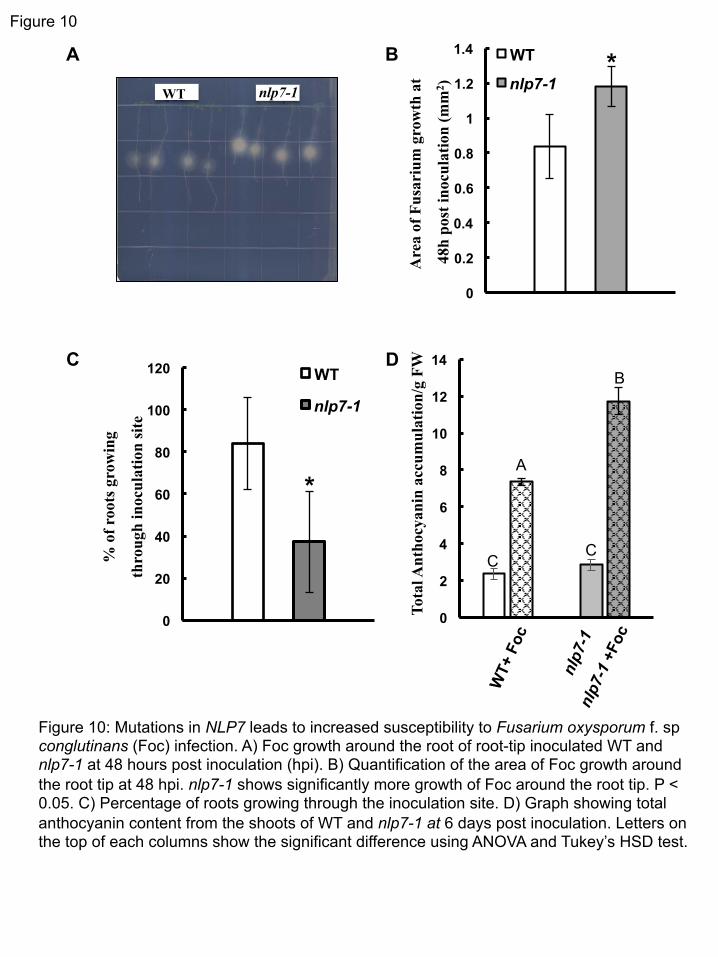
Figure 10
Figure 10: Mutations in NLP7 leads to increased susceptibility to Fusarium oxysporum f. sp conglutinans (Foc) infection. A) Foc growth around the root of root-tip inoculated WT and nlp7-1 at 48 hours post inoculation (hpi). B) Quantification of the area of Foc growth around the root tip at 48 hpi. nlp7-1 shows significantly more growth of Foc around the root tip. P < 0.05. C) Percentage of roots growing through the inoculation site. D) Graph showing total anthocyanin content from the shoots of WT and nlp7-1 at 6 days post inoculation. Letters on the top of each columns show the significant difference using ANOVA and Tukey’s HSD test.
A
C
0
20
40
60
80
100
120 WT
nlp7-1
*
% o
f roo
ts g
row
ing
thro
ugh
inoc
ulat
ion
site
0
0.2
0.4
0.6
0.8
1
1.2
1.4 WT
nlp7-1 *
Are
a of
Fus
ariu
m g
row
th a
t
48h
pos
t ino
cula
tion
(mm
2 )
C
D
C
0
2
4
6
8
10
12
14
Tota
l Ant
hocy
anin
acc
umul
atio
n/g
FW
C
B
A
B
nlp7-1 WT

C
BRN1/2 NLP7 SMB
Low pH
Pectin Lyase XTH5
Border-like cell loosening
Border-like cell release
CEL5
Figure 11: Model for NLP7 control of border-like cell release. See text for details.
Figure 11

Parsed CitationsBarlow P (2003) The Root Cap: Cell Dynamics, Cell Differentiation and Cap Function. J Plant Growth Regul 21: 261-186
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Benfey PN, Scheres B (2000) Root development. Curr Biol 10: R813-815Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bennett T, van den Toorn A, Sanchez-Perez GF, Campilho A, Willemsen V, Snel B, Scheres B (2010) SOMBRERO, BEARSKIN1,and BEARSKIN2 regulate root cap maturation in Arabidopsis. Plant Cell 22: 640-654
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bennett T, van den Toorn A, Willemsen V, Scheres B (2014) Precise control of plant stem cell activity through parallel regulatoryinputs. Development 141: 4055-4064
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Blancaflor EB, Fasano JM, Gilroy S (1998) Mapping the functional roles of cap cells in the response of Arabidopsis primary roots togravity. Plant Physiol 116: 213-222
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Blancaflor EB, Fasano JM, Gilroy S (1999) Laser ablation of root cap cells: implications for models of graviperception. Adv SpaceRes 24: 731-738
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Borisov AY, Madsen LH, Tsyganov VE, Umehara Y, Voroshilova VA, Batagov AO, Sandal N, Mortensen A, Schauser L, Ellis N, et al(2003) The Sym35 gene required for root nodule development in pea is an ortholog of Nin from Lotus japonicus. Plant Physiol 131:1009-1017
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bouton S, Leboeuf E, Mouille G, Leydecker MT, Talbotec J, Granier F, Lahaye M, Hofte H, Truong HN (2002) QUASIMODO1encodes a putative membrane-bound glycosyltransferase required for normal pectin synthesis and cell adhesion in Arabidopsis.Plant Cell 14: 2577-2590
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Brady SM, Orlando DA, Lee JY, Wang JY, Koch J, Dinneny JR, Mace D, Ohler U, Benfey PN (2007) A high-resolution rootspatiotemporal map reveals dominant expression patterns. Science 318: 801-806
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Camargo A, Llamas A, Schnell RA, Higuera JJ, Gonzalez-Ballester D, Lefebvre PA, Fernandez E, Galvan A (2007) Nitrate signalingby the regulatory gene NIT2 in Chlamydomonas. Plant Cell 19: 3491-3503
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Carpita NC, Gibeaut DM (1993) Structural models of primary cell walls in flowering plants: consistency of molecular structure withthe physical properties of the walls during growth. Plant J 3: 1-30
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Castaings L, Camargo A, Pocholle D, Gaudon V, Texier Y, Boutet-Mercey S, Taconnat L, Renou JP, Daniel-Vedele F, Fernandez E,et al (2009) The nodule inception-like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J 57: 426-435
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chardin C, Girin T, Roudier F, Meyer C, Krapp A (2014) The plant RWP-RK transcription factors: key regulators of nitrogenresponses and of gametophyte development. J Exp Bot 65: 5577-5587
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Clausen MH, Willats WG, Knox JP (2003) Synthetic methyl hexagalacturonate hapten inhibitors of anti-homogalacturonanmonoclonal antibodies LM7, JIM5 and JIM7. Carbohydr Res 338: 1797-1800
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. PlantJ 16: 735-743
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Czechowski T, Bari RP, Stitt M, Scheible WR, Udvardi MK (2004) Real-time RT-PCR profiling of over 1400 Arabidopsistranscription factors: unprecedented sensitivity reveals novel root- and shoot-specific genes. Plant J 38: 366-379
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
del Campillo E, Abdel-Aziz A, Crawford D, Patterson SE (2004) Root cap specific expression of an endo-beta-1,4-D-glucanase(cellulase): a new marker to study root development in Arabidopsis. Plant Mol Biol 56: 309-323
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Driouich A, Follet-Gueye ML, Vicre-Gibouin M, Hawes M (2013) Root border cells and secretions as critical elements in plant hostdefense. Curr Opin Plant Biol 16: 489-495
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dubois M, Gilles K, Hamilton J, Rebers P, Smith F (1956) Colorimetric Method for Determination of Sugars and RelatedSubstances. Anal. Chem 28: 350-356
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Durand C, Vicre-Gibouin M, Follet-Gueye ML, Duponchel L, Moreau M, Lerouge P, Driouich A (2009) The organization pattern ofroot border-like cells of Arabidopsis is dependent on cell wall homogalacturonan. Plant Physiol 150: 1411-1421
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Eapen D, Barroso ML, Ponce G, Campos ME, Cassab GI (2005) Hydrotropism: root growth responses to water. Trends Plant Sci 10:44-50
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fendrych M, Van Hautegem T, Van Durme M, Olvera-Carrillo Y, Huysmans M, Karimi M, Lippens S, Guerin CJ, Krebs M,Schumacher K, et al (2014) Programmed cell death controlled by ANAC033/SOMBRERO determines root cap organ size inArabidopsis. Curr Biol 24: 931-940
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ferris PJ, Goodenough UW (1997) Mating type in Chlamydomonas is specified by mid, the minus-dominance gene. Genetics 146:859-869
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gunawardena U, Hawes MC (2002) Tissue specific localization of root infection by fungal pathogens: role of root border cells. MolPlant Microbe Interact 15: 1128-1136
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hawes MC, Brigham LA, Wen F, Woo HH, Zhu Y (1998) Function of root border cells in plant health: pioneers in the rhizosphere.Annu Rev Phytopathol 36: 311-327
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hawes MC, Gunawardena U, Miyasaka S, Zhao X (2000) The role of root border cells in plant defense. Trends Plant Sci 5: 128-133Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hawes MC, Lin HJ (1990) Correlation of Pectolytic Enzyme Activity with the Programmed Release of Cells from Root Caps of Pea(Pisum sativum). Plant Physiol 94: 1855-1859

Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Iuchi S, Koyama H, Iuchi A, Kobayashi Y, Kitabayashi S, Kobayashi Y, Ikka T, Hirayama T, Shinozaki K, Kobayashi M (2007) Zincfinger protein STOP1 is critical for proton tolerance in Arabidopsis and coregulates a key gene in aluminum tolerance. Proc NatlAcad Sci USA 104: 9900-9905
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Iyer-Pascuzzi AS, Jackson T, Cui H, Petricka JJ, Busch W, Tsukagoshi H, Benfey PN (2011) Cell identity regulators linkdevelopment and stress responses in the Arabidopsis root. Dev Cell 21: 770-782
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Knox JP, Linstead PJ, King J, Cooper C, Roberts K (1990) Pectin esterification is spatially regulated both within cell walls andbetween developing tissues of root apices. Planta 181: 512-521
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kochian LV, Pineros MA, Liu J, Magalhaes JV (2015) Plant adaptation to Acid soils: the molecular basis for crop aluminumresistance. Annu Rev Plant Biol 66: 571-598
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Konishi M, Yanagisawa S (2013) Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nat Commun 4:1617
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Koyama H, Toda T, Hara T (2001) Brief exposure to low-pH stress causes irreversible damage to the growing root in Arabidopsisthaliana: pectin-Ca interaction may play an important role in proton rhizotoxicity. J Exp Bot 52: 361-368
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kumpf RP, Nowack MK (2015) The root cap: a short story of life and death. J Exp Bot. doi:10.1093/jxb/erv295Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Levesque MP, Vernoux T, Busch W, Cui H, Wang JY, Blilou I, Hassan H, Nakajima N, Matsumoto N, Lohmann JU, et al (2006) Whole-genome analysis of the SHORT-ROOT developmental pathway in Arabidopsis. PLoS Biol 4(5): e143. DOI:10.1371/journal.pbio.0040143
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lewis JA, Papavizas GC (1991) Biocontrol of plant disease: the approach for tomorrow, Crop Protection, 10: 95
Lumsden RD, Lewis JA, Fravel DR (1995) Formulation and delivery of biocontrol agents for use against soilborne plant pathogens.In F Hall, J Barry eds. Biorational Pest Control Agents, Formation and Delivery. American Chemical Society, Washington DC, pp166- 182
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li X, Ma J, Hiradate S, Matsumoto H (2000) Mucilage strongly binds aluminum but does not prevent roots from aluminum injury inZea mays. Physiol Plantarum 108: 152-160
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lin H, Goodenough UW (2007) Gametogenesis in the Chlamydomonas reinhardtii minus mating type is controlled by two genes,MID and MTD1. Genetics 176: 913-925
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Marchive C, Roudier F, Castaings L, Brehaut V, Blondet E, Colot V, Meyer C, Krapp A (2013) Nuclear retention of the transcriptionfactor NLP7 orchestrates the early response to nitrate in plants. Nat Commun 4: 1713
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Marsh JF, Rakocevic A, Mitra RM, Brocard L, Sun J, Eschstruth A, Long SR, Schultze M, Ratet P, Oldroyd GE (2007) Medicago

truncatula NIN is essential for rhizobial-independent nodule organogenesis induced by autoactive calcium/calmodulin-dependentprotein kinase. Plant Physiol 144: 324-335
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mouille G, Ralet MC, Cavelier C, Eland C, Effroy D, Hematy K, McCartney L, Truong HN, Gaudon V, et al (2007) Homogalacturonansynthesis in Arabidopsis thaliana requires a Golgi-localized protein with a putative methyltransferase domain. Plant Journal 50:605-14
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nagahashi G, Douds DD, Jr. (2004) Isolated root caps, border cells, and mucilage from host roots stimulate hyphal branching of thearbuscular mycorrhizal fungus, Gigaspora gigantea. Mycol Res 108: 1079-1088
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Plancot B, Santaella C, Jaber R, Kiefer-Meyer MC, Follet-Gueye ML, Leprince J, Gattin I, Souc C, Driouich A, Vicre-Gibouin M(2013) Deciphering the responses of root border-like cells of Arabidopsis and flax to pathogen-derived elicitors. Plant Physiol 163:1584-1597
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Provart NJ, Gil P, Chen W, Han B, CHang H-S, Wang X, Zhu T (2003) Gene Expression Phenotypes of Arabidopsis Associated withSensitivity to Low Temperatures. Plant Physiol 132 (893-906)
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sabba RP, Lulai EC (2002) Histological analysis of the maturation of native and wound periderm in potato (Solanum tuberosum L.)Tuber. Ann Bot 90: 1-10
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schauser L, Roussis A, Stiller J, Stougaard J (1999) A plant regulator controlling development of symbiotic root nodules. Nature402: 191-195
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schauser L, Wieloch W, Stougaard J (2005) Evolution of NIN-like proteins in Arabidopsis, rice, and Lotus japonicus. J Mol Evol 60:229-237
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sims DA, Gamon JA (2002) Relationships between leaf pigment content and spectral reflectance across a wide range of species,leaf structures and developmental stages. Remote Sensing of Environment 81: 337-354
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sterling C (1970) Crystal-structure of ruthenium red and stereochemistry of its pectic stain. American Journal of Botany 57: 172-175Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tjamos EC, Beckman CH (1989) Vascular Wilt Diseases of Plants. Berlin: Springer-Verlag.Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Truog E (1946) Soil Reaction Influence on Availability of Plant Nutrients. Soil Science Society Proceedings: 305 - 308Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vicre M, Santaella C, Blanchet S, Gateau A, Driouich A (2005) Root border-like cells of Arabidopsis. Microscopical characterizationand role in the interaction with rhizobacteria. Plant Physiol 138: 998-1008
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Watson BS, Bedair MF, Urbanczyk-Wochniak E, Huhman DV, Yang DS, Allen SN, Li W, Tang Y, Sumner LW (2015) Integratedmetabolomics and transcriptomics reveal enhanced specialized metabolism in Medicago truncatula root border cells. Plant Physiol167: 1699-1716

Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wen F, Zhu Y, Hawes MC (1999) Effect of pectin methylesterase gene expression on pea root development. Plant Cell 11: 1129-1140
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Willats WG, Limberg G, Buchholt HC, van Alebeek GJ, Benen J, Christensen TM, Visser J, Voragen A, Mikkelsen JD, Knox JP(2000) Analysis of pectic epitopes recognised by hybridoma and phage display monoclonal antibodies using definedoligosaccharides, polysaccharides, and enzymatic degradation. Carbohyd Res 327: 309-320
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Willemsen V, Bauch M, Bennett T, Campilho A, Wolkenfelt H, Xu J, Haseloff J, Scheres B (2008) The NAC domain transcriptionfactors FEZ and SOMBRERO control the orientation of cell division plane in Arabidopsis root stem cells. Dev Cell 15: 913-922
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wolf S, Mouille G, Pelloux J (2009) Homogalacturonan methyl-esterification and plant development. Mol Plant 2: 851-860Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Xie F, Murray JD, Kim J, Heckmann AB, Edwards A, Oldroyd GE, Downie JA (2012) Legume pectate lyase required for root infectionby rhizobia. Proc Natl Acad Sci USA 109: 633-638
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title