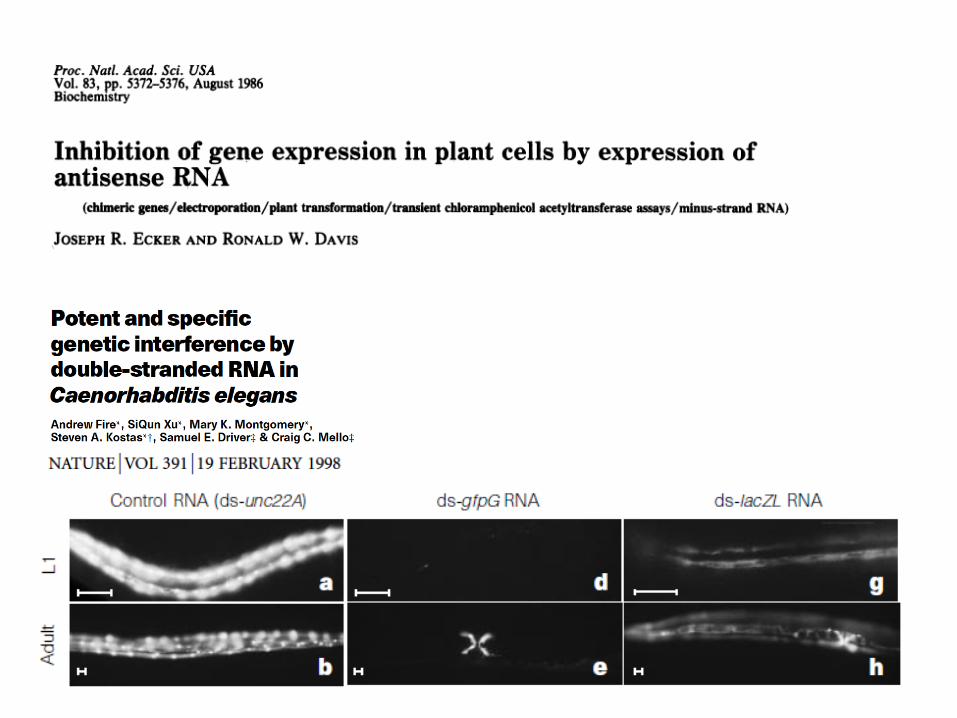
rna interferencia prize in physiology and medicine 2006 for their discovery of rna interference –...
TRANSCRIPT

RNAinterferencia
dsRNA~10xúčinnejšiaakosamotnésensealeboan=sense
RNA

Nobel prize in Physiology and Medicine 2006 for their discovery of RNA interference – gene silencing
by double-stranded RNA
AndrewFireStanfordUniversity
CraigMelloUniversityofMassachuseIs

RNA interferencia
RNA interferencia - post-transkripčné sekvenčne-špecifické
utlmovanie génov (gene silencing) dvojvláknovou RNA Dvojkrokový mechanistický model:
1. Iniciačný krok (Dicer) – Tvorba miRNA/siRNA
2. Efektorový krok (RISC/RITS) – Degradácia cieľovej mRNA – Inhibícia translácie – Modifikácia chromatínu
https://www.youtube.com/watch?v=cK-OGB1_ELE

Dicer • postrach každej dsRNA
• člen rodiny RNáz III s dsRNA-špecifickou endoribonukleázovou aktivitou, štiepi dsRNA na 19-25 nukleotidové fragmenty a odštepuje vlásenku z pre-miRNA
• Výsledná dsRNA má 2 nt
prečnievanie na 3´koncoch • 2-O-metylácia pri 3´konci rastliných
miRNA/siRNA - stabilita
!"#$%&'(&)*&+,'!'-./&0$/"%'1&//'2*./.34
"#$%&#'
()*+,%-'.
5(26
7*0&%
89:;
/012--
" 345 678 3797:; :<
=7778- =7778>3458?@</AB&%C$-'&4D32>CEFCEG
D=H32>CEFCEG,I&J.C="42%++,2>CEFCEG
,I&J.C="4*&I.CE#'2>CEFCEG
(-*-%)'C'
F'="4>CEFCEG
8C$&I2>CEFCEG A!2&EF>CEFCEG
20!2&EF>CEFCEG
K%C$&I2-$*CLC*)
F'=H8
F'=H8
345
=7778-
=7778>
B&%C$-'& 4D32>CEFCEG7*0&% 89:;! "
#
MN,+I*
?EOCEFCEG
4PQ2%+-FCEG
8C$CEG2-EF,I+F#$*I&%&-'&
$''(&3$/"#*.<'.='7*0&%'>%.0&,,*<3%
(&3$/"#*.<'.='5(26'"<?'89:'>%.#&*<,
(&3$/"#*.<'.='@*(!8',#"A*/*#4
8C$&IR8C$&IJ;!"#$!%&'
4PQ;ST
-"#$%&(BC1
6%&D(BC1
4PQ;ST
BK(UVSBK31V
D=H3
:&*JUW2.C=J<;W2.C=J;X
.C=J;T0W
.C=J<A>
.C=J;TX-W
.C=;A0>
.C=J;UW
.C=AT-W
.C=J1XW
.C=J;<0>
.C=J;0;
:&*JU
D?DT-EF2D?DU
87KA:<
YK=3
8C$&I
:7"</
6(373;7=M;"
H(87"A8
?????? ???
6&
Z=";-EF2Z="<R!"#$%&'"()*+*,-#.#/"$,'
3"3-'&2+%FJA0
D=H3
4PQ
4PQ
3
QB
M=Y
3 643Y43Y<W24YDA
T3B
K*->C%C[-*C+E
K*->C%C[-*C+EW23J>+F)2%+$-%C[-*C+E
3J>+F)2%+$-%C[-*C+EW2*I-E'%-*C+E-%2I&,I&''C+E
4PQ3 MP@=
=&F#$&F28C$&I2>CEFCEG\
4PQ34=3
()*+,-.(/0!1)&%'
=&%C&L&F2*-IG&*2I&,I&''C+E
0! A!
;W1<<2--
AXX2--
484=;
7E+'CE&
0!2&EF2+]G#CF&2="4
A!2&EF2+]G#CF&2="4
4$*CL&2'C*&22DI,
345
"
3797
678:<
:;
the region between the PAZ and RNase III domains functions as a ‘molecular ruler’ (REFS!46,104) (FIG.!2b). The structure of the Dicer helicase domain remains unknown, because Dicer of G.!intestinalis lacks the domains upstream of the PAZ domain. However, on the basis of several electron microscopy studies, it has been suggested that human Dicer is an L-shaped mol-ecule107–109 (FIG.!3d). It will be important to solve the high-resolution structure of mammalian Dicer in complex with its natural substrate.
Dicer binds to pre-miRNA with a preference for a two-nucleotide-long 3! overhang that was initially gener-ated by Drosha46 (FIG.!2b). In general, Dicer cleavage sites are located at a fixed distance from the 3! end of the ter-minus of dsRNAs (the 3!-counting rule)46,104,110–112. This distance is typically 21–25 nucleotides in length and depends on the species and the type of Dicer. In"mammals and flies, there is an additional mechanism to deter-mine the cleavage site of pre-miRNA: Dicer binds to the 5!"phosphorylated end of the pre-miRNA and cleaves it 22 nucleotides away from the 5! end (the 5!-countin g rule)105. The 5! end binding occurs when the end is thermodynamically unstable, but not when the end is"strongly paired (such as through G·C base pairs). The structure of the PAZ domain of human Dicer shows two basic pockets that bind to the 5!"end and 3!"end of the pre-miRNA105,106 (FIG.!2b). The pockets are spatially arranged in a way that they can be occupied simultane-ously by the 5! end and 3! end of the pre-miRNA when the RNA has a two-nucleotide-long 3! overhang. This structural arrangement may explain at least in part the preference of human Dicer for a two-nucleotide-long 3!"overhang structure.
Dicer regulation by its cofactors and products. RNase"III proteins often interact with dsRBD proteins (FIG.!3a). Drosophila spp. Dcr-1 binds to one of the two isoforms of Loquacious (Loqs), Loqs-PA and Loqs-PB, each"of which contains three dsRBDs113–115. Both isoforms are necessary for the production of most mi RNAs, whereas a subset of mi RNAs requires Loqs-PB specifi-cally for their generation116–118. Loqs-PB can alter the Dcr-1-processing site to yield specific miRNA iso-forms116. Similarly, human Dicer interacts with TAR RNA-binding protein (TRBP; encoded by TARBP2), a homologue of Loqs-PB with three dsRBDs119–121 (FIG.!3c). The dsRBD cofactor PACT (also known as PRKRA) also associates with mammalian Dicer121,122, but its exact role in the miRNA pathway remains to be determined. Like Loqs, TRBP modulates the processing efficiency of some pre-mi RNAs and tunes the length of mature mi RNAs116,123,124. Unlike Loqs, the mammalian dsRBD cofactors (TRBP or PACT) do not seem to be essential for Dicer-mediated pre-miRNA processing, as purified mammalian Dicer is comparable to the Dicer–dsRBD cofactor complex in the processing of pre-mi RNAs121. Of note, TRBP and PACT were previously described as negative and positive regulators of dsRNA-dependent protein kinase"R (PKR), respectively125. The relation-ship between the miRNA pathway and PKR signalling is unknown.
!"#$"%&
NATURE REVIEWS | !"#$%&#'()%$##)*+"#",-) VOLUME 15 | AUGUST 2014 | !"!
© 2014 Macmillan Publishers Limited. All rights reserved

Tkanivovo-špecifický knock-out Dicera ovplyvňuje vývin končatín u myší
WT Dicerknock‐out
Harfeakol.,2005

RISC: RNA-induced silencing complex
• Argonaut (Ago) proteín rozpoznáva a viaže dsRNA (napomáha Dicer), vzniká pre-RISC
• Dochádza k rozpleteniu dsRNA, jedno vlákno (guide strain) zostáva viazané v komplexe, kým druhé vlákno (passenger strain) je degradované, vzniká RISC
• Ako rozpozná RISC, ktoré vlákno je vodiace (guide)?
Na základe relatívnej termodynamickej stability koncov duplexu (asymetrické pravidlo) – vlákno s menej stabilným 5‘koncom slúži ako vodiace (guide)
!"#$%&'(&)*&+,'!'-./&0$/"%'1&//'2*./.34
"#$%&#'
()*+,%-'.
5(26
7*0&%
89:;
/012--
" 345 678 3797:; :<
=7778- =7778>3458?@</AB&%C$-'&4D32>CEFCEG
D=H32>CEFCEG,I&J.C="42%++,2>CEFCEG
,I&J.C="4*&I.CE#'2>CEFCEG
(-*-%)'C'
F'="4>CEFCEG
8C$&I2>CEFCEG A!2&EF>CEFCEG
20!2&EF>CEFCEG
K%C$&I2-$*CLC*)
F'=H8
F'=H8
345
=7778-
=7778>
B&%C$-'& 4D32>CEFCEG7*0&% 89:;! "
#
MN,+I*
?EOCEFCEG
4PQ2%+-FCEG
8C$CEG2-EF,I+F#$*I&%&-'&
$''(&3$/"#*.<'.='7*0&%'>%.0&,,*<3%
(&3$/"#*.<'.='5(26'"<?'89:'>%.#&*<,
(&3$/"#*.<'.='@*(!8',#"A*/*#4
8C$&IR8C$&IJ;!"#$!%&'
4PQ;ST
-"#$%&(BC1
6%&D(BC1
4PQ;ST
BK(UVSBK31V
D=H3
:&*JUW2.C=J<;W2.C=J;X
.C=J;T0W
.C=J<A>
.C=J;TX-W
.C=;A0>
.C=J;UW
.C=AT-W
.C=J1XW
.C=J;<0>
.C=J;0;
:&*JU
D?DT-EF2D?DU
87KA:<
YK=3
8C$&I
:7"</
6(373;7=M;"
H(87"A8
?????? ???
6&
Z=";-EF2Z="<R!"#$%&'"()*+*,-#.#/"$,'
3"3-'&2+%FJA0
D=H3
4PQ
4PQ
3
QB
M=Y
3 643Y43Y<W24YDA
T3B
K*->C%C[-*C+E
K*->C%C[-*C+EW23J>+F)2%+$-%C[-*C+E
3J>+F)2%+$-%C[-*C+EW2*I-E'%-*C+E-%2I&,I&''C+E
4PQ3 MP@=
=&F#$&F28C$&I2>CEFCEG\
4PQ34=3
()*+,-.(/0!1)&%'
=&%C&L&F2*-IG&*2I&,I&''C+E
0! A!
;W1<<2--
AXX2--
484=;
7E+'CE&
0!2&EF2+]G#CF&2="4
A!2&EF2+]G#CF&2="4
4$*CL&2'C*&22DI,
345
"
3797
678:<
:;
the region between the PAZ and RNase III domains functions as a ‘molecular ruler’ (REFS!46,104) (FIG.!2b). The structure of the Dicer helicase domain remains unknown, because Dicer of G.!intestinalis lacks the domains upstream of the PAZ domain. However, on the basis of several electron microscopy studies, it has been suggested that human Dicer is an L-shaped mol-ecule107–109 (FIG.!3d). It will be important to solve the high-resolution structure of mammalian Dicer in complex with its natural substrate.
Dicer binds to pre-miRNA with a preference for a two-nucleotide-long 3! overhang that was initially gener-ated by Drosha46 (FIG.!2b). In general, Dicer cleavage sites are located at a fixed distance from the 3! end of the ter-minus of dsRNAs (the 3!-counting rule)46,104,110–112. This distance is typically 21–25 nucleotides in length and depends on the species and the type of Dicer. In"mammals and flies, there is an additional mechanism to deter-mine the cleavage site of pre-miRNA: Dicer binds to the 5!"phosphorylated end of the pre-miRNA and cleaves it 22 nucleotides away from the 5! end (the 5!-countin g rule)105. The 5! end binding occurs when the end is thermodynamically unstable, but not when the end is"strongly paired (such as through G·C base pairs). The structure of the PAZ domain of human Dicer shows two basic pockets that bind to the 5!"end and 3!"end of the pre-miRNA105,106 (FIG.!2b). The pockets are spatially arranged in a way that they can be occupied simultane-ously by the 5! end and 3! end of the pre-miRNA when the RNA has a two-nucleotide-long 3! overhang. This structural arrangement may explain at least in part the preference of human Dicer for a two-nucleotide-long 3!"overhang structure.
Dicer regulation by its cofactors and products. RNase"III proteins often interact with dsRBD proteins (FIG.!3a). Drosophila spp. Dcr-1 binds to one of the two isoforms of Loquacious (Loqs), Loqs-PA and Loqs-PB, each"of which contains three dsRBDs113–115. Both isoforms are necessary for the production of most mi RNAs, whereas a subset of mi RNAs requires Loqs-PB specifi-cally for their generation116–118. Loqs-PB can alter the Dcr-1-processing site to yield specific miRNA iso-forms116. Similarly, human Dicer interacts with TAR RNA-binding protein (TRBP; encoded by TARBP2), a homologue of Loqs-PB with three dsRBDs119–121 (FIG.!3c). The dsRBD cofactor PACT (also known as PRKRA) also associates with mammalian Dicer121,122, but its exact role in the miRNA pathway remains to be determined. Like Loqs, TRBP modulates the processing efficiency of some pre-mi RNAs and tunes the length of mature mi RNAs116,123,124. Unlike Loqs, the mammalian dsRBD cofactors (TRBP or PACT) do not seem to be essential for Dicer-mediated pre-miRNA processing, as purified mammalian Dicer is comparable to the Dicer–dsRBD cofactor complex in the processing of pre-mi RNAs121. Of note, TRBP and PACT were previously described as negative and positive regulators of dsRNA-dependent protein kinase"R (PKR), respectively125. The relation-ship between the miRNA pathway and PKR signalling is unknown.
!"#$"%&
NATURE REVIEWS | !"#$%&#'()%$##)*+"#",-) VOLUME 15 | AUGUST 2014 | !"!
© 2014 Macmillan Publishers Limited. All rights reserved
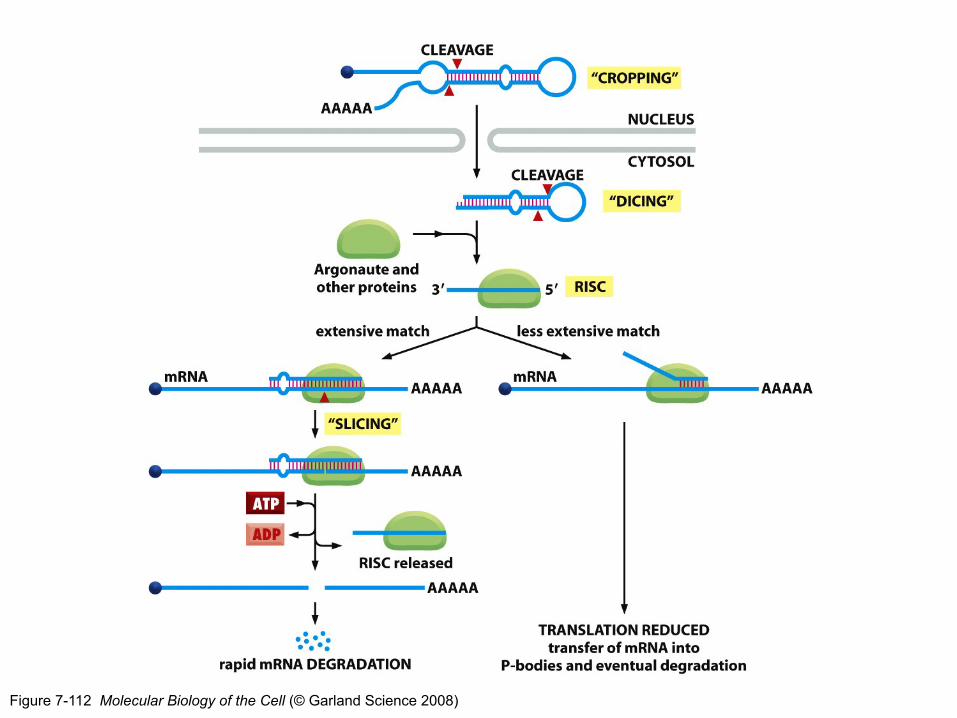
Figure 7-112 Molecular Biology of the Cell (© Garland Science 2008)

Figure 7-115 Molecular Biology of the Cell (© Garland Science 2008)

MaléutlmujúceRNAuživočíchov
PachyteneThe stage of meiotic prophase that immediately follows the zygotene. It is characterized by paired chromosomes that are condensed and visibly divided into chromatids, and by the occurrence of crossing-over.
Let-7 familyThe let-7 gene was initially discovered as an essential developmental gene in Caenorhabditis!elegans and, later, as one of the first two microRNAs (mi RNAs). The let-7 miRNA family is highly conserved throughout bilaterian animals, and it suppresses cell proliferation and promotes cell!differentiation. It is also a tumour suppressor.
but it is generally considered that mi RNAs with identi-cal sequences at nucleotides 2–8 of the mature miRNA belong to the same ‘miRNA family’ (REF.!4). For instance, the human genome contains 14 paralogous loci (encoding ‘miRNA sisters’) that belong to the let-7 family. Thirty-four miRNA families are phylogenetically conserved from C.!elegans to humans, and 196 miRNA families are conserved among mammals16,20. miRNA sisters gener-ally act redundantly on target mRNAs, but distinct roles have also been suggested21. Some mi RNAs share a com-mon evolutionary origin but diverge in the miRNA seed. For instance, miR-141 and miR-200c belong to a deeply
conserved miR-200 superfamily and differ by one nucleo-tide in their miRNA seeds. Deletion of each sister locus showed that the targets of miR-141 and miR-200c barely overlap, which illustrates the importance of the miRNA seed sequence in miRNA function and evolution22.
The nomenclature of miRNA genes is somewhat inconsistent. The genes found in early genetic studies were named after their phenotypes (for example, lin-4, let-7 and lsy-6), whereas most mi RNAs found from clon-ing or sequencing received numerical names (for example, the lin-4 homologues in other species are called mir-125). Genes encoding miRNA sisters are indicated with lettered suffixes (for example, mir-125a and mir-125b). If the same mature miRNA is generated from multiple separate loci, numeric suffixes are added at the end of the names of the miRNA loci (for example, mir-125b-1 and mir-125b-2). Each locus produces two mature mi RNAs: one from the 5! strand and one from the 3! strand of the precursor (for example, miR-125a-5p and miR-125a-3p). However, one arm (called the ‘guide’ strand) is usually much more prevalent (96–99% of the sum on average) and more bio-logically active than the other arm (the ‘passenger’ strand, which is known as miRNA*).
Transcriptional regulation. miRNA sequences are located within various genomic contexts. In humans, the major-ity of canonical mi RNAs are encoded by introns of non-codin g or coding transcripts, but some mi RNAs are encoded by exonic regions. Often, several miRNA"loci are in close proximity to each other, constituting a poly-cistronic transcription unit23. The mi RNAs in the same cluster are generally co-transcribed, but the individual mi RNAs can be additionally regulated at the post-transcriptional level. One of the most deeply conserved clusters is the mir-100~let-7~mir-125 cluster, which has an important role in the development of bilaterian ani-mals24. The let-7 miRNA, but not the other mi RNAs, is suppressed post-transcriptionally in embryonic stem cells and in certain cancer cells in mammals.
Precise locations of the miRNA promoters have not yet been mapped for most miRNA genes but can be inferred from collective analysis of CpG islands, RNA sequenc-ing data and ChIP–seq (chromatin immuno precipitation followed by sequencing) data25. Some miRNA genes reside in the introns of protein-coding genes and, thus, share the promoter of the host gene. However, it has
!"#$%$&$!"#$$%&%'#(%)*+)&+$,#""+$%"-*'%*.+/01$+%*+#*%,#"$
!"#$#%&$#%'"$##%(&)*$%'+,#-%*.%-(&//%-0/#12013%456-%01%&10(&/-7%(02$*456-%8(0%456-9:%
-0456-%&1;%<=>=?01'#$&2'013%456-%8,0456-9@:AB%(0%456-%&$#%3#1#$&'#;%.$*(%-"*$'%"&0$,01%
!"#$%&'%()*%$*+,*-(./0%/1(.2-%23%(42%!"/$*%5556('7*%782(*.-$%9:82$)/;/-<%:.1*8=>%$.!"#$%
8CD@%1E2/#*'0;#-%/*139%&$#%;#$0F#;%.$*(%/*13%;*EG/#?-'$&1;#;%456-%*$%/*13%-'#(H/**,%
-'$E2'E$#-%'"$*E3"%I02#$%,$*2#--013@B%!"#+%(#;0&'#%'"#%,*-'?'$&1-2$0,'0*1&/%-E,,$#--0*1%
*.%'$&1-2$0,'-%&1;%'$&1-,*-*1-:%&1;%2*1'$0GE'#%'*%&1'0F0$&/%;#.#12#@B%,0456-%8DJHKL%
1E2/#*'0;#-%/*139%&$#%1*'%;#,#1;#1'%*1%45&-#%===?'+,#%,$*'#01-%&1;%&$#%,$*;E2#;%.$*(%
-013/#?-'$&1;#;%,$#2E$-*$-%G+%&1%#1;*1E2/#&-#%2&//#;%ME22"010%8&/-*%N1*O1%&-%
(0'*2"*1;$0&/%2&$;0*/0,01%"+;$*/&-#%01%"E(&1-9%&1;%&-%+#'%E10;#1'0.0#;%'$0((013%
#1P+(#-D:DQRHDQSB%!"#%(&01%.E12'0*1%*.%,0456-%0-%'*%-0/#12#%'$&1-,*-&G/#%#/#(#1'-%01%
3#$(/01#%2#//-:%&/'"*E3"%'"#%$*/#-%*.%-*(#%,0456-%8.*$%#T&(,/#:%pachytene%,0456-%&1;%'"*-#%,$*;E2#;%*E'-0;#%*.%'"#%3#$(%/01#9%&$#%-'0//%#103(&'02B%U.%'"#%'O*%-EG2/&;#-%*.%
6$3*1&E'#%,$*'#01-%86VU%&1;%<=>=9:%(0%456-%&1;%-0456-%&$#%&--*20&'#;%O0'"%'"#%
#?@;782(*.-$A%4)*8*/$%7.!"#$%&.-<%(2%B5C5%782(*.-$>
!"#$%&"' ()*+, ')*+, -)*+,
'()*+, -..$)/01("+23(4 -.%$)/01("+23(4 .5678$)/01("+23(4
9:"0(442)*$();<=(4
>:"4,?$?)3$>20(:
>20(: @/00,2)2$?)3$/)A)"B)$+:2==2)*$();<=(4
CDE$4/F01?3(4
CDE CDE 9GHG
I(0,?)24=$"J$?0+2")
D%K:?)41?+2")?1$:(L:(442")
D%=MNC$3(*:?3?+2")
MNC$01(?O?*( D%K:?)40:2L+2")?1$":$L"4+P+:?)40:2L+2")?$1$:(L:(442")$"J$+:?)4L"4")4
D%I/1+2*()(:?+2")?1$(L2*()(+20$L,()"=()?$2)$B":=4
Q/)0+2") M(*/1?+2")$"J$L:"+(2)P0"32)*$*()(4
D%M(*/1?+2")$"J$L:"+(2)P0"32)*$*()(4$?)3$+:?)4L"4")4
D%C)+2O2:?1$3(J()0(
D%9:(PL?0,<+()($L2MNCR$+:?)4L"4")$421()02)*
D%9?0,<+()($L2MNCR$/)A)"B)D%L2MNCP12A($4=?11$MNC$2)$4"=?R$/)A)"B)
!"#$.$&$,%/01+2%).-*-$%$+3#(45#6+%*+3"#*($
W0N#%'"#0$%&10(&/%2*E1'#$,&$'-:%,/&1'%,$0(&$+%(02$*456-%8,$0?(0%456-9%&$#%(*-'/+%'$&1-2$0G#;%G+%456%,*/+(#$&-#%==%&1;%
'"#0$%/#13'"%0-%"03"/+%"#'#$*3#1#*E-@@:@K:@JB%X*O#F#$:%E1/0N#%&10(&/%(0%456-:%,/&1'%(0456%,$*2#--013%0-%2*(,/#'#;%01%'"#%
1E2/#E-:%&1;%"*(*/*3E#-%*.%I$*-"&%&1;%IVY4A%&$#%1*'%.*E1;%01%,/&1'-B%I=YZ4?W=[Z%@%8IYW@9%,$*2#--#-%(*-'%,$0?(0%456-%
G+%-#\E#1'0&/%2/#&F&3#B%!"#%456?G01;013%,$*'#01%I6>IWZ%8IIW9%01'#$&2'-%O0'"%IYW@%&1;%-'&G0/0P#-%,$0?(0%456-%01%
1E2/#&$%.*20%2&//#;%;02013%G*;0#-%8I?G*;0#-9B%!"#%P012?.013#$%,$*'#01%]Z446!Z%8]Z9:%'"#%;*EG/#?-'$&1;#;%456?G01;013%
,$*'#01%X^<U56]!=Y%WZ6_Z]%@%8X^W@9:%IYW@%&1;%'"#%1E2/#&$%2&,?G01;013%2*(,/#T%.*$(%&%2*(,/#T%&1;%,$*2#--%
,$0?(0%456-B%̀ *//*O013%,$*2#--013:%'"#%(0456Ha,&--#13#$%-'$&1;b%8(0456c9%;E,/#T%0-%D!?!?(#'"+/&'#;%&'%'"#%K!;*-<%&'%Xd6%Z5X65YZ4%@%8XZ5@9:%O"02"%G/*2N-%E$0;+/&'0*1%8G+%XZ5@%]d<<4Z]]U4%@%8XZ]U@99%&1;%;#2&+%*.%(0%456-%8G+%K!He!%#T*$0G*1E2/#&-#-%]f6WW?456?IZV46I=5V%5dYWZ6]Z%@%8]I5@9:%]I5D%&1;%]I5K9B%,$#?(0%456-%*$%(&'E$#%(0%456-%&$#%
'"#1%#T,*$'#;%'*%'"#%2+'*,/&-(%G+%X6]!^%8X]!9:%'"#%,/&1'%"*(*/*3E#%*.%#T,*$'01%e%8Zg<e9B%61%&;;0'0*1&/%#T,*$'%,&'"O&+%
-##(-%'*%G#%01F*/F#;%GE'%$#(&01-%E1N1*O1B%=1%'"#%2+'*,/&-(:%(0%456-%&$#%/*&;#;%*1'*%2+'*,/&-(02%64VU56d!Z%86VU9%
,$*'#01-:%*.%O"02"%6VU@%0-%'"#%(&)*$%,/&+#$%.*$%'"#%(0456%,&'"O&+B
!"#$"%&
789+| AUGUST 2014 | VOLUME 15 .///01#$%&"023(4&"5)"/'4(362"667)3
© 2014 Macmillan Publishers Limited. All rights reserved
HaaKim,2014,NatRevMolCellBiol

miRNAasiRNAsalíšiapárovanímscieľovouRNA
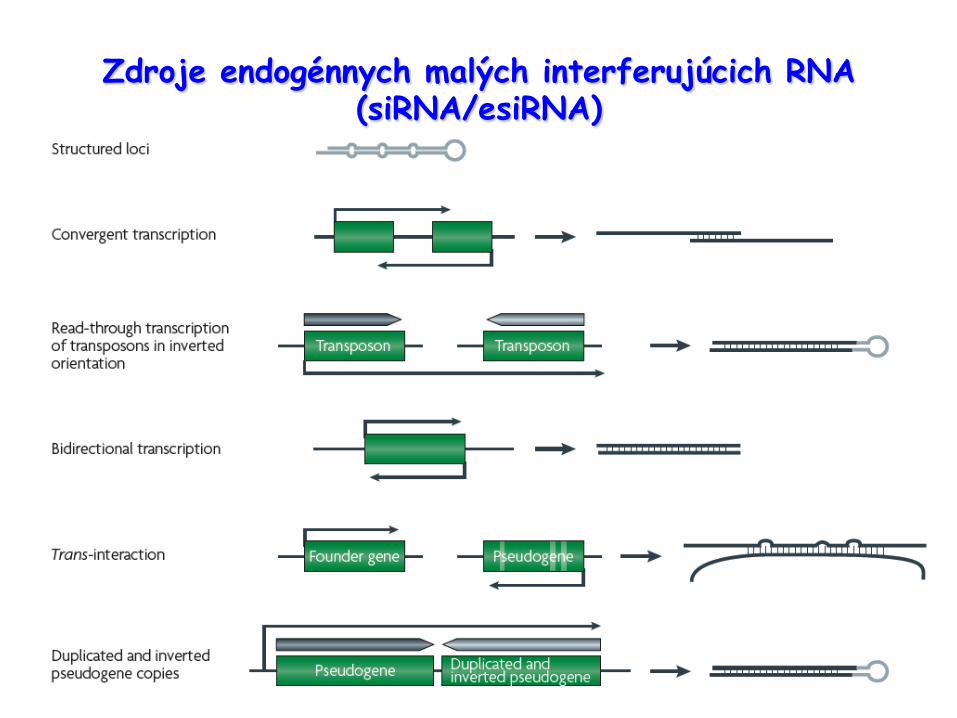
Zdroje endogénnych malých interferujúcich RNA (siRNA/esiRNA)

Účinky malých interferujúcich RNA u rastlín a živočíchov

ReguláciamiRNA/siRNA• iniciáciatranskripciePolIIaPolIII• lncRNAšpongie• circRNA
ciRS‐7sequestersandinac=vatesmiR‐7butisresistanttomiR‐7regula=onduetoitscircularnatureprovidesapla_ormfortrappingthemiR‐7/Argonautecomplex,its‘‘supersponging’’capacityantagonizesmiR‐7ac=vity
Kjems, Mao (2013) Cell
m

Chalkónsyntáza
RdRPRNA‐závisláRNApolymeráza(rastliny,červy,niektorékvasinky)

Muljoakol.(2010)WIREsSystemsBiologyandMedicine
EvolúciaRNAinterferencie

PoruchyRNAinterferencie
uviaceroľudskýchochorení–dereguláciamiRNAalebonarušeniefunkciekomponentovRNAi:‐DiGeorgovsyndróm(1:3000del):srdcovédefekty,imunodeficiencia,schizofrénia,pos=hujeasi30génov,Droshaproteín,defektvbiogenézemiRNA‐ niektorémiRNAnajúfunkciunádorovýchsupresorovaleboprotoonkogénov(veľaanalyzovanýchmiRNAležívoblas=achčastýchchromozómovýchprestávbpozorovanýchvnádorochalebonafragilnýchmiestach)

NiektorénenádorovéchorobyviazanésmiRNA

RNA‐seq

ReverznátranskripciamiRNA
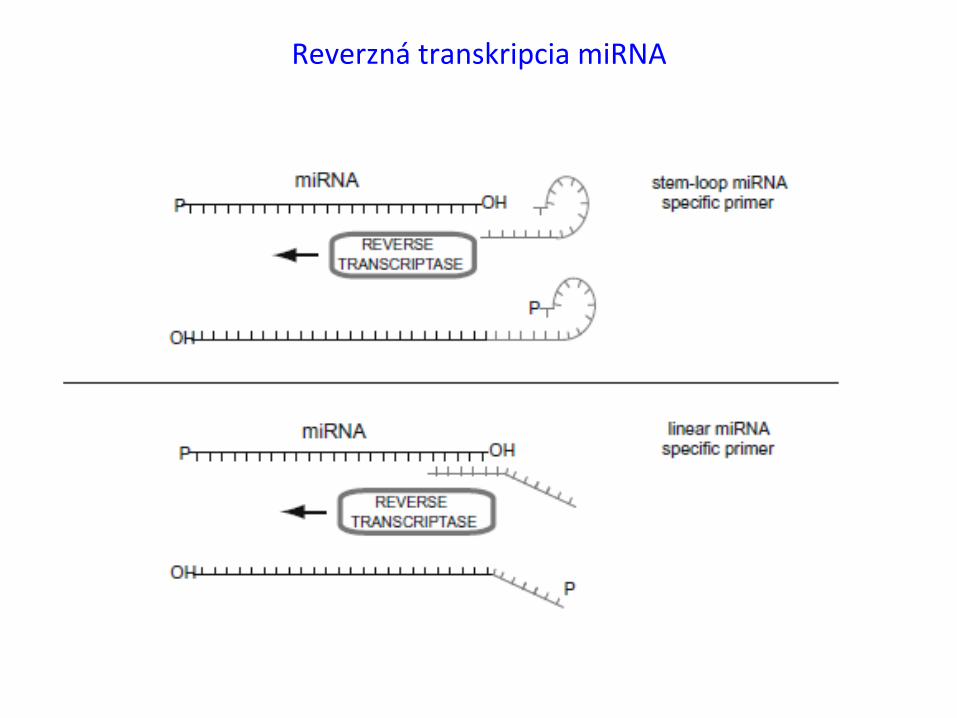
ReverznátranskripciamiRNA

Experimentálne využitie
• Rastliny - injekcia dsRNA alebo transfekcia je postačujúca
pre RNAi (RNA-závislá RNA polymeráza) • Cicavčie bunky
- nepostačujúce (stabilné konštrukty) - injekcia dlhých dsRNA aktivuje dsRNA-závislú proteín kinázu – aktivuje sa interferónová odpoveď typu 1 a nešpecifický globálny shutdown translácie

• dsRNA syntetizované chemicky
• Vektory na báze plazmidov - krátkodobý charakter - nízka účinnosť transfekcie - plazmidy na báze promótora RNA polymerázy
Pol III: aktívna vo všetkých bunkách Pol II: tkanivovo špecifický génový silencing
• Vektory na báze vírusov
- adenovírusy, adeno-asociované vírusy alebo lentivírusy - vysoká účinnosť ale môžu mať vedľajšie účinky
ProdukciasiRNA
U6
H1
siRNA
U6
Hairpin loop
Sense
Antisense

TitráciamiRNApomocoumiRNAšpongie
EbertaSharp,2010
miRNA

Dodanie (delivery) siRNA
In vitro - Chemická transfekcia - Elektroporácia do parazitov
In vivo
- intramuskulárna injekcia - endocytóza nanočastíc
Dodanie (delivery) siRNA

Použi=esiRNA
VýskumTerapia• cytotoxickáodpoveďpovnesenícudzorodejdsDNA
• dostatočneefeklvnevneseniesiRNA/miRNAdo cieľovýchbuniek
• trvalosťúčinku
• schopnosťvektoroviden=fikovať/infikovaťcieľovébunky
• špecifickéutlmenieibamutantnejverziegénu

Pareledonesp.‐1,8oC(Antarklda)
Octopusvulgaris30oC(Portoriko)
Rýchlosť poklesu akčného potenciálu na oddychovú hodnotu je závislá na teplote okolia; Kinetika otvárania draslíkového kanála v axónoch chobotníc žijúcich v arktických a tropických vodách je podobná, pričom sekvencia génov kódujúcich tento kanál sa nelíši

Funkčné zmeny kódujúcich sekvencií v dôsledku editovania RNA A I
AdaptácianeurónovchobotnicenachladnejšiepodnebiejesprostredkovanáeditovanímRNAdraslíkovéhokanála(voltage‐dependentK+channel)
GareIaRosenthal(2012)Science)

Editovanie RNA (RNA editing)
• mechanizmus post-transkripčnej úpravy RNA, pri ktorom dochádza k zmene sekvencie:
- substitúcia/modifikácia - inzercia/delécia
• A, G, U, C, I
• editovanie RNA môže meniť význam informácie v mRNA (zmena kódovania)

Editovanie RNA nie je:
Maturácia RNA - delécia, prípadne ligácia dlhších úsekov RNA Modifikácia RNA – biochemická zmena vnášajúca neštandardné nukleotidy do RNA (neplatí pre inozín)

Editovanie RNA „C-to-U“ A novel form of tissue-specific RNA processing produces apolipoprotein-B48 in intestine. Powell LM, Wallis SC, Pease RJ, Edwards YH, Knott TJ, Scott J. Cell. 1987 Sep 11;50(6):831-40.
Human proapolipoprotein A-I: development of an antibody to the propeptide as a probe of apolipoprotein A-I biosynthesis and processing. Hospattankar AV, Fairwell T, Appella E, Meng M, Brewer HB Jr. Biochem Biophys Res Commun. 1987 Nov 30;149(1):289-96
Apolipoprotein B-48 is the product of a messenger RNA with an organ-specific in-frame stop codon. Chen SH, Habib G, Yang CY, Gu ZW, Lee BR, Weng SA, Silberman SR, Cai SJ, Deslypere JP, Rosseneu M, et al. Science. 1987 Oct 16;238(4825):363-6

Editovanie RNA v cicavčích bunkách Editovaniepre‐mRNAapolipoproteínu‐B
pečeňLDL,transportcholesteroluvkrvi,C‐koncovádoménasaviaženaLDLreceptor,ktorýje
prítomnývplazma=ckejmembráne
tenkéčrevoChylomikróny,absorpcialipidovvtenkom
čreve,transportTG,zmenavpozícii6666zCnaUvnesieposIranskripčne„stop“kodón
(ci=díndeamináza)