review of membrane potentials, action potentials, …. ion channels and resting membrane potential a...
TRANSCRIPT

Review of membrane potentials, action potentials, synaptic transmission and paroxysmal depolarization © L Stan Leung, Department of Physiology and Pharmacology, University of Western Ontario Objectives: To review topics on: 1. Ionic permeability and membrane potential – Nernst & Goldman equations 2. Action potential – voltage dependent Na+ channel, activation and inactivation
and propagation 3. Synaptic transmission – pre- and postsynaptic components, EPSP and IPSP,
GABAergic and glutamate receptors 4. Paroxysmal depolarization and ictal discharges References 1. Kandel ER et al. Principles of Neural Science, 4th edition, 2000. 2. Alberts et al. “Molecular Biology of the Cell, 4th Edition, 2002.
1

2

1. Ion Channels and Resting Membrane Potential A resting membrane potential is found for all cells of the body. It is of particular importance in neurons. Depolarization or decrease of the magnitude of the resting potential may give rise to the action potential. Ionic channels are responsible for different potentials (resting potentials, action potentials or postsynaptic potentials). 1.1 Recording of electrical activities Intracellular versus extracellular recordings. Intracellular electrodes: Sharp electrode – conventional intracellular recordings, typically used for recording
voltage inside a cell, a glass tubing is pulled to a sharp tip of < 1 μm diameter Patch electrodes – low resistance electrode, tightly sealed to membrane, to record
membrane currents, single ion channels Extracellular electrode to record electroencephalogram (EEG), local field potential, can
be small (microelectrode) to large electrode, e.g., EEG discs; EMG (electromyogram, of muscles) and EOG (electro-oculogram) for eye muscles are also extracellular recordings
Also to record multiple or single unit (neuronal) activity extracellularly, unit recordings typically meant recording of action potentials (spikes)
1.1.2 Recording Membrane Potentials of a cell (Kandel p. 127) Figure shows a cell is impaled with a sharp electrode made by pulling a heated
glass capillary tube (tip size < 1 μm diameter). The microelectrode is typically filled with a concentrated K+ solution (e.g., 2 M KCl) for intracellular recording (since K+ is more concentrated inside the cell). Concentrated Na+ solutions may be used to fill extracellularly recording micropipettes. The concentrated solution acts as a conductor. A reference electrode is placed outside the membrane of the cell and the appropriate connections are made to an amplifier (or an oscilloscope). On the oscilloscope, the measurement is voltage (y-axis) versus time (x-axis).
3

1.1.3. Ionic concentration across a mammalian neuron (Kandel p.128-130). There
are 4 main ions in the central nervous system, sodium, potassium, chloride and calcium. Unfortunately, Kandel et al. only listed the ion concentrations in the squid, which are significantly different from those in mammals. Approximate values of the concentrations are:
Ion inside conc. outside conc. Nernst potential (37oC) (mM) (mM) (round off) K+ 135 3.1 -96 Na+ 31 145 +40 Cl- 7 120 -75 Ca++ 0.01 2 +70
4

1.1.4 Ion Channels and patch recordings. Kandel, Chapter 6, p.105-123. Ion channels are protein channels or pores in the membrane made up of
proteins. The protein channels are embedded in the impermeable lipid bilayer. The pore (of the order of nm in diameter) may be specific for a particular ion (i.e. selective for charge and size), and it may be activated or inactivated by voltage as in the action potential sodium channel. Ion channel may have a chemical sensor that detects binding with a chemical (neurotransmitter, e.g.), voltage (electrical field) or mechanical stretch.
5

There are several main types of ion channels, categorized according to the
gating/signalling (Kandel p.229-230): 1. Non-gated leak channels, like those participating in the resting membrane,
which are not gated by voltage or by neurotransmitter. 2. Voltage-gated channels 3. Ligand-gated or Neurotransmitter-gated channels, which can have direct or
indirect types. In a directly gated channel, also called an ionotropic receptor, the neurotransmitter binds to the receptor, which is the same as the ion channel, and opens the channel. Nicotinic acetylcholine-gated channel is an example, in which Ach binds to the receptor which is the same molecule as the channel. In an indirectly gated channel, also called a metabotropic receptor, the neurotransmitter binds to a receptor that through second messengers, signals to ion channels to make them open or close. The indirect type has separate receptor (that receives the signal) and ion channel (that mediates ionic changes), with intermediate signalling molecules like G-proteins, followed by cAMP, phosphoinositol, etc.
6

1.1.5 Aquaporins (AQPs) are proteins that form an aqueous pore, or water channel,
on the membrane. Aquaporins are found in kidney tubules, epithelia and glial cells in the brain. Aquaporin (AQ1) on the chloroid plexus may facilitate the secretion of cerebrospinal fluid. Aquaporins (AQ4 and others) on glial cells facilitate the uptake of water from the extracellular space in the brain; water uptake is positively correlated with K+ uptake. Water flow at the blood-brain barrier is partly controlled by aquaporins on the astrocyte end feet membrane, and thus aquaporins regulate brain edema. AQP4 has been implicated as a marker of the inflammatory demyelinating disease neuromyelitis optica, a form of multiple sclerosis in which inflammatory lesions are restricted to the optic nerve and spinal cord (Verkman AS. 2009. Aquaporins: translating bench research to human disease. J Exp Biol. 212:1707-15).
1.1.6 The equilibrium potential is the quasi-stable potential where chemical
(concentration) force is equal to the electrical force. One way to estimate the polarity (positive or negative sign) of the equilibrium potential for each ion is: (1) know where is the ion more abundant, intra- or extracellular, e.g. K+ is more abundant intracellularly, (2) the chemical (concentration) force points from the high to low concentration and moves ions in this direction, e.g., K+ chemical gradient points from in to outside of the cell, and will move K+ ions out of the cell,
7

8

9

1.1.7. Nernst equation. Kandel p. 125-134. The magnitude of the equilibrium potential can be estimated by the Nernst equation if the actual concentrations in and outside the cell are known, in the case of K+, EK = (RT/ Fz) ln [Ko/Ki] = 62 log [Ko/Ki] in mV at 37oC where EK = equilibrium potential for K+, R= gas constant, T= absolute temperature, F= Faraday's constant, z= valence =1 for K+, (note it will be -1 for Cl-1) Ko= outside K+ concentration, Ki = inside K+ concentration. If the resting membrane potential (RMP) is determined solely by K+, then RMP = EK = 62 log Ko - 62 log Ki in mV at 37oC This is the equation of a linear relation between RMP and (log Ko)
10

Kandel p. 132-134. Goldman-Hodgkin-Katz (constant field) equation emphasizes the participation of all ions according to their permeabilities; it reduces to the Nernst equation for a single ion if the other ions are not permeable (P=0), e.g., if at rest, the permeability to K+ is much larger than that to Na+ or Cl-, the resting membrane potential is the Nernst equation EK. The Goldman equation is
Eresting = RT/F ln{ [PNaNao + PKKo + PClCli]/[PNaNai + PKKi + PClClo]} Where PNa = permeability to Na+
PK = permeability to K+
PCl = permeability to Cl-
11

12

2. Action Potential 2.1 Time course and permeability of an action potential. Depolarization results in a subthreshold local potential that triggers a regenerative, voltage-dependent event, the action potential. Action potentials (also called spikes) are central to signal processing in the brain, and changes in the pattern and frequency of action potentials indicate changes in sensory, motor, or internal signals. An action potential in vivo describes a change of membrane voltage with time. The action potential is known to be mediated by a fast transient increase in Na+ conductance (gNa) followed by a delayed and more prolonged increase in K+ conductance (gK). Conductance is an electrical term that is equivalent to permeability.
2.2 Voltage Clamp study of action potential. In experimental studies, it is important to study ionic currents during the action potential, with the voltage experimentally fixed (clamped) at a particular value. In late 1940s and early 1950s, British physiologists used voltage clamp, a state-of-the-art technology at the time, to record membrane currents from the squid axon. Effectively, the voltage of a membrane is maintained constant, at the value determined by the researcher, by an electronic device that quickly injects current through the membrane. Membrane currents are measured during voltage clamp, which is a valuable tool to study voltage-dependent events.
13

Voltage Clamp by two electrodescan be done in giant squid axon
Measure current
Squid axonlongitudinal section
Commandvoltage
600 um
Figure The large diameter of the squid axon allows penetration of two electrodes, one for injecting current and one for recording voltage.
Figure Currents of an action potential induced by a step in the command voltage during voltage clamp. To evoke action currents, the axon membrane is stepped from a hyperpolarized potential (-60 mV) to a depolarized potential (0 mV). Action currents are the currents of the action potential. The action current is first inward (down) for several msec, associated with Na+, and then followed by an outward K+ current. The convention is that outward current is positive.
14

Figure. A delayed outward K+ current was induced by a voltage step in the squid axon when Na+ was replaced with choline+, a cation that could not pass through the Na+ channel (Na+ inward current was abolished). This was done by Hodgkin and Huxley (H & H) in the 1940s. Today, we can also block the voltage-sensitive sodium current with a specific Na+ channel blocker tetrodotoxin (TTX) derived from the Puffer fish. TTX was discovered in late 1950s. The isolated the K+ current, called the delayed rectifier current Ik , is outward (up) and lasts as long as the voltage is depolarized (i.e., no inactivation is found for this current).
15

Figure. Subtracting the delayed outward current from the total action current gives the inward current in the action current. H and H derived the pure Na+ current by subtracting the K+ current from the total current. Today, sodium current in the squid axon could also be isolated by adding tetraethylammonium (TEA), a drug that blocks the delayed rectifier K+ channel (Ik ). By stepping to different voltages, Hodgkin and Huxley also showed that the sodium and potassium currents during an action potential were voltage-dependent. They constructed a theoretical model of action potential generation that is still valid today. Recording shows that the Na+ channel inactivates. Once the sodium channels are turned off (inactivated), they cannot be activated again by further depolarization. To be activated by a depolarizing command pulse, the inactivation must be relieved. This is done, as shown by the following figure, by returning to a more hyperpolarized level. These results suggest that the Na+ channel has two gates, an activation and an inactivation gates, and that it can exist in three major states: 1) normally closed state near resting membrane potential, when activation gate is closed but inactivation gate is open: 2) open (i.e., both gates are open); and 3) inactivated state with inactivation gate closed but activation gate open (i.e., nonconducting and not capable of being opened by a further depolarization). Schematically, this relationship can be shown as follows.
16

2.3. K+ channels (Alberts p. 652-653; Kandel p. 158-159), in addition to Na+ channel inactivation, help to repolarize the action potential. Three common types of K+ currents that help to repolarize an action potential are: (1) The classical K+ channels present on squid axons called the delayed rectifier K+ channels Ik , and these channels are blocked by TEA. In voltage clamp recordings, Ik is slow to start and has no or little inactivation (Figure below). (2) In mammalian axons, repolarization of the action potential is also mediated by transient and voltage-dependent A-type K+ channels (IA) that are blocked by 4-aminopyridine (4-AP). IA is an example of early K+ channels (Alberts p. 653) that activate and inactivate quickly with depolarization (Figure below). The early activation of IA has a suppressive effect on firing the action potential (i.e., increases the action potential threshold). (3) Ca2+ dependent K+ channels.
17

2.4 Propagation of an action potential
Figure. Local circuit (or electrotonic) currents are responsible for the initial depolarization that leads to propagation of action potential. A) Resting membrane with negative charges inside and positive charges outside the membrane. B) Action potential is assumed at membrane near point 1 with inward Na+ current flowing from point 1 to intracellular point 2. C) A local circuit current consists of a longitudinal intracellular current from point 2 to 3, outward passive membrane current (from 3 to 4), and extracellular current from point 4 to point 1. Outward passive current depolarizes the membrane at point 4. D) Depolarization activates voltage-gated inward Na+ currents at point 4, causing more depolarization and starting an action potential now at point 4 (instead of point 1 in B). Meanwhile, membrane at point 1 repolarizes and is refractory to another action potential.
18

Figure. In myelinated nerve, the local circuit involves two nodes and not internodes. Local circuit (electrotonic) currents depolarize the next node and activate voltage-dependent Na+ currents (and the action potential) at the next node. Since the action potential jumps (salteres) from one node to the other, it is known as saltatory conduction. Myelinated axons are covered with a myelin sheath formed by Schwann cells wrapping around the axon, except at the nodes (Alberts p 641; figure above). Myelin serves to increase resistance and decrease capacitance of the axon membrane. Less charge will leak through the myelinated portion of the membrane, and an increase in membrane resistance will increase the space constant, or equivalently, relatively more currents will flow internally along the axon from one node to the next node.
19

3 Synaptic Transmission For neurons to communicate with each other, signals have to transmit across connections between neurons called synapses. As demonstrated by Sherrington in the early 1900, before the structure of the synapse is known, synaptic transmission is use-dependent and susceptible to drugs. Synapses are sites for signal processing. Temporal and spatial summation occurs in the soma and dendrites of neurons. Changes in synaptic strength form the cellular basis for learning and memory. Many drugs such as general anesthetics, sedatives, antidepressants, etc., act on synaptic transmission rather than action potential propagation. 3.1 Synaptic transmission. A synapse is a connection between two nerve cells (Alberts p. 645-646). There are two main types of synapses – electrical and chemical synapses. In electrical synapses, electrical continuity is maintained between pre- and postsynaptic neurons, and action potential spreads quickly from pre- to post-synaptic membrane, providing fast and reliable transmission. Gap junctions (connexons) between pre- and postsynaptic cells may allow passage of small molecules as well as electrical currents. In chemical synapses, there is a physical gap (cleft) between pre- and postsynaptic membranes and communication between cells depends on a chemical substance (neurotransmitter) that travels across the gap. Chemical synaptic transmission is slower (delay of >0.3 msec) and can be modulated (i.e. changed) more readily than electrical transmission.
20

For a neuron to signal to another, its action potential has to transmit cross a synapse. Electrical synapse will allow the action potential to be transmitted directly to the postsynaptic cell. At chemical synapses, there is a physical cleft that the action potential cannot jump across. Instead, the electrical signal is converted to a chemical signal that diffuses across this synaptic cleft. The sequence of chemical synaptic transmission is given below (Kandel Fig. 10.7; p. 182-185; Alberts p. 649-651): (1) presynaptic depolarization by action potential opens voltage-dependent
Ca2+ channels on the presynaptic terminals (2) Ca2+ influx to nerve terminal cause exocytosis of vesicles containing
neurotransmitters and release the neurotransmitter into synaptic cleft (3) neurotransmitter acts on postsynaptic receptor after diffusion across
synaptic cleft (4) postsynaptic receptor is a transmitter-gated ion channel that opens with
transmitter binding; some channels close with transmitter binding. (5) postsynaptic membrane potential changes, increasing or decreasing the
probability of action potential discharges
3.2 Excitatory postsynaptic potential (EPSP) depolarizes a cell and tends to generate an action potential. Inhibitory postsynaptic potential (IPSP) hyperpolarizes a cell or keeps it near the resting potential, thus suppressing generation of action potentials. Thus, whether an action potential is fired depends on the sum of EPSPs and IPSPs converging on a neuron. Thousands of excitatory and inhibitory synapses converge on a neuron in the brain.
Figure of an excitatory postsynaptic potential. The duration of the EPSP or is typically 15-30 ms in duration.
21

EPSP in neurons is typically mediated by an increase of permeability to both Na+
and K+. The equilibrium potential of an EPSP, like an end plate potential (EPP), is typically about 0 mV (somewhat closer to the Na+ than the K+ equilibrium potential). Nicotinic Ach receptor is an excitatory neurotransmitter receptor. Glutamate receptors or glutamate-gated ion channels are common excitatory postsynaptic receptors in the brain.
Figure of inhibitory postsynaptic potentials (IPSPs). IPSPs in the brain are typically longer than EPSPs, and of duration > 30 ms. Gamma-aminobutyric acid (GABA) is a common inhibitory neurotransmitter in the brain. IPSP is typically mediated by either an increase in Cl- or K+ permeability. One
type of GABA receptor, the GABA-A receptors, open Cl- channels with equilibrium potential of about –70 mV, while another type, the GABA-B receptors, open K+ channels with equilibrium potential of -90 mV.
IPSPs are such named because they suppress excitation or the generation of
action potentials. GABA-A receptor mediated IPSPs typically have large conductances such that their opening will move the neuron towards the equilibrium potential of the IPSP, i.e., near –70 mV.
22

3.3. Glutamate receptors (Kandel, p.212-217; Fig.12-5). Glutamate is the most
common excitatory neurotransmitter in the brain, with 4 major types of receptors: i) AMPA (AMPA= alpha-amino-3-hydroxyl-5-methyl-isoxazole-4-
proprionate), ii) kainate, iii) N-methyl-D-aspartate (NMDA) and iv) metabotropic The AMPA, kainate and NMDA receptors are direct ligand-gated channels, or iontropic glutamate channels. The action of glutamate on iontropic receptors is always excitatory, i.e., an EPSP is induced with reversal potential of about 0 mV, and the underlying permeability change is mediated by both Na+ and K+. NMDA receptor channel is also permeable to Ca++. Metabotropic receptors (mGluRs) are indirectly gated channels that affect metabolic processes; they act through G-proteins, and signal through phospholipase C, diacyl glycerol and inositol triphosphate. Either excitation or inhibition of a neuron may be the result of mGluR activation.
The NMDA receptor is unique in being both ligand and voltage gated, i.e., it will not open even with a large amount of glutamate unless the postsynaptic cell is depolarised. Depolarization removes a Mg2+ ion that blocks the channel. NMDA receptor channel is permeable to Na+, K+ and Ca++. Ca++ influx is important since it is the signal for synaptic plasticity, which is necessary during development or for learning and memory. Excessive Ca++ influx may, however, be toxic for the cell. Glycine is a co-agonist at the NMDA receptor. Psychomimetic drugs such as phencyclidine and ketamine are partial blockers of NMDA receptors. NMDA receptors are also implicated in neurotoxicity associated with ischemia, hypoglycemia, and neurodegenerative diseases.
23

24

4. Paroxysmal depolarization, interictal and ictal discharges Seizure activity is characterized by abnormal, synchronous activity among a
population of neurons. The paroxysmal depolarization shift was first reported as the abnormal transient event induced by penicillin on the cortex. It corresponds to an interictal spike, or an abnormal event between seizures, rather than the seizure (ictal discharge).
25
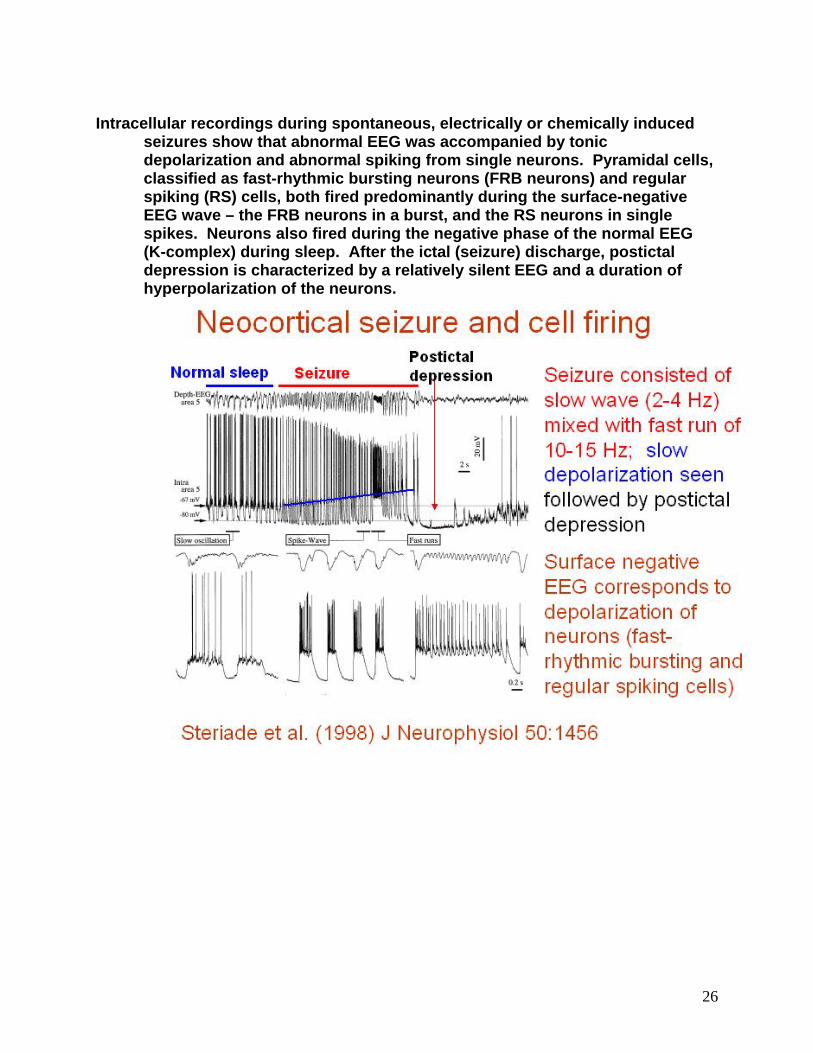
Intracellular recordings during spontaneous, electrically or chemically induced
seizures show that abnormal EEG was accompanied by tonic depolarization and abnormal spiking from single neurons. Pyramidal cells, classified as fast-rhythmic bursting neurons (FRB neurons) and regular spiking (RS) cells, both fired predominantly during the surface-negative EEG wave – the FRB neurons in a burst, and the RS neurons in single spikes. Neurons also fired during the negative phase of the normal EEG (K-complex) during sleep. After the ictal (seizure) discharge, postictal depression is characterized by a relatively silent EEG and a duration of hyperpolarization of the neurons.
26