review article micrornas as a new mechanism...
TRANSCRIPT

Review ArticlemicroRNAs as a New Mechanism Regulating Adipose TissueInflammation in Obesity and as a Novel Therapeutic Strategy inthe Metabolic Syndrome
Qian Ge1 Sonia Brichard2 Xu Yi3 and QiFu Li1
1 Department of Endocrinology The First Affiliated Hospital of Chongqing Medical University Chongqing 400016 China2 Endocrinology Diabetes and Nutrition Unit Institute of Experimental and Clinical Research Medical SectorUniversity of Louvain 1200 Brussels Belgium
3Neurology Unit Daping Hospital Research Institute of Surgery TheThird Military Medical University Chongqing 400042 China
Correspondence should be addressed to QiFu Li liqifuyeahnet
Received 24 November 2013 Accepted 21 January 2014 Published 24 March 2014
Academic Editor David Kaplan
Copyright copy 2014 Qian Ge et alThis is an open access article distributed under the Creative Commons Attribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Obesity is associated closely with the metabolic syndrome (MS) It is well known that obesity-induced chronic inflammation playsa fundamental role in the pathogenesis of MS White adipose tissue (AT) is the primary site for the initiation and exacerbation ofobesity-associated inflammation Exploring the mechanisms of white AT inflammation and resetting the immunological balancein white AT could be crucial for the management of MS Several prominent molecular mechanisms have been proposed to mediateinflammation in white AT including hypoxia endoplasmic reticulum stress lipotoxicity and metabolic endotoxemia Recently agrowing body of evidence supports the role of miRNAs as a new important inflammatory mediator by regulating both the adaptiveand innate immunity This review will focus on the implication of miRNAs in white AT inflammation in obesity and will alsohighlight the potential of miRNAs as targets for therapeutic intervention in MS as well as the challenges lying in miRNA-targetingtherapeutics
1 Introduction
The incidence of obesity defined as body mass index (BMI) ge30 kgm2 has increased drastically worldwide during recentdecades Obesity is associated with a cluster of metabolicdisorders including increased risk of insulin resistancetype 2 diabetes (T2DM) hypertension dyslipidemia andcardiovascular disease these disorders and obesity per seconstitute a serious threat known as the metabolic syndrome(MS) [1] It is well known that chronic inflammation is a keyfeature of obesity this obesity-induced inflammation buildsthe common soil for the pathogenesis of MS [2] Thereforeresetting the immunological balance in obesity is a crucialapproach for the management of MS
White adipose tissue (AT) has been regarded as theprimary goal of pharmacological intervention since obesity-induced inflammation is mainly initiated and exacerbatedin this organ [3ndash5] Although several prominent molecular
mechanisms have been proposed to trigger inflammationin white AT including hypoxia endoplasmic reticulumstress lipotoxicity and metabolic endotoxemia [5] thesefactors cannot fully explain the origin of inflammationOn the other hand current anti-inflammatory strategiesare not sufficient enough in the treatment of MS Thesefacts suggest that there are lots of unknowns responsiblefor AT inflammation in obesity In recent years a growingbody of evidence has highlighted the modulating rolesof miRNAs in immuneinflammatory system [6 7] andtheir involvement in the obesity-related metabolic disordersincluding T2DM and atherosclerosis [8 9] Specific miRNAshave been implicated in adipogenesis and mature adipocytefunction [10] As yet whether thesemiRNAs could be anothermechanism for mediating AT inflammation and whetherthese tiny molecules could serve as potent therapeutic anti-inflammatory targets forMS are still not fully settledThe aimof the present review is to address these questions
Hindawi Publishing CorporationJournal of Immunology ResearchVolume 2014 Article ID 987285 10 pageshttpdxdoiorg1011552014987285
2 Journal of Immunology Research
2 Obesity Inflammationand Metabolic Syndrome
21 Low-Grade Inflammation as a Key Feature of ObesityDuring the past decade it became clear that inflammationis a key feature of obesity [2] The inflammatory responsetriggered by obesity involves many components of the clas-sical inflammatory response to pathogens and includes theincreases in circulating inflammatory cytokines and acute-phase proteins (eg C-reactive protein) recruitment ofleukocytes to inflamed tissues activation of tissue leuko-cytes and generation of reparative tissue responses (egfibrosis) However the nature of obesity-induced inflam-mation referred to as ldquometainflammationrdquo (metabolicallytriggered inflammation) is unique compared with otherinflammatory paradigms (eg infection and autoimmunedisease) in several key aspects First the chronic nature ofobesity produces a low-grade activation of the innate immunesystem that affects metabolic homeostasis over time Secondchildhood obesity may place individuals at risk for lifelonginflammation since inflammatory markers are elevated inobese children as young as 3 years old Third this chronicinflammation is composed of recurrent acute episodes ofnutrition-related immune activation induced by nutrientavailability (fasting or high-fat meals) This fluctuation maybe associatedwith the induction of pro- or anti-inflammatorymediators Finally obesity-induced chronic inflammation isunique by its multiorgan involvement [11]
22 Obesity-Induced Inflammation Linking Obesity to Meta-bolic Syndrome The metabolic syndrome (MS) refers to theclustering of cardiovascular risk factors that include centralobesity hyperglycemia dyslipidemia and hypertension Theultimate importance of this cluster is to identify individuals athigh risk of both T2DM and cardiovascular disease Growingevidence implicates obesity-induced inflammation as animportant mechanism linking obesity to the MS in metabol-ically active organs Assessment of gene expression networksin obese AT has identified a robust pattern of overexpressedinflammatory genes associated with metabolic disease [1213] Multiple inflammatory mediators abnormally secretedby AT and the crosstalk between immune and metaboliccells can impair insulin signaling and induce oxidativestress and endothelial dysfunction leading to systemicinsulin resistance and cardiovascular disease [5] Deregulatedmacrophage-myocyte and macrophage-hepatocyte signalingcan impair insulin sensitivity as well [14 15] Hypothalamicinflammation which is induced very rapidly by a high-fatdiet [16]may provoke hyperphagia and has been documentedto impair insulin release from 120573 cells peripheral insulinaction and potentiate hypertension [17ndash19] Thus chronicexcess of nutrients such as lipids and glucose may simultane-ously trigger inflammatory responses which further disruptmetabolic function enhancing stress and inflammationSuch a vicious cycle is identified as a mechanism leadingto further metabolic deterioration Therefore breaking thisvicious circle by resetting the immunological balance inobesity is a crucial approach for the management of MS
3 The Fundamental Role of White AdiposeTissue in Obesity-Induced Inflammation
Obesity is characterized by excessive expansion of whiteadipose tissue (AT) which has been thought to be theprimary site for the initiation of obesity-associated inflam-mation Although ATrsquos principal function deals with energystorage it serves as an active secretory organ as well Anumber of bioactive peptides or proteins collectively namedldquoadipokinesrdquo are produced and secreted by fat andor nonfatcells of white AT They act in an autocrineparacrine mannerto regulate local AT function and also act in an endocrinemanner to influence the functions of distant tissues such asliver skeletal muscle and cardiovascular and central nervoussystems [5 20]
In obesity white AT is remodeled dynamically by adi-pocyte hypertrophy (increased size) hyperplasia (increasednumber) immune cell infiltration endothelial cell over-activation and extracellular matrix overproduction [21ndash24] This remodeling may trigger metabolic and hypoxicstress resulting in activation of multiple inflammatorysignaling pathways ultimately leading to dysregulation ofnumerous adipokines including proinflammatory cytokineschemokines growth factors acute-phase proteins andcomplement-like factors Virtually all known adipokines aredysregulated in obesity via multiple mechanisms Such aderegulation is the main feature of AT low-grade inflamma-tion and contributes to the pathogenesis of MS [5]
4 Molecular Mechanisms Mediating WhiteAdipose Tissue Inflammation
Several prominent mechanisms have been highlighted thatcoordinately mediate white AT inflammation
Hypoxia In obesity the growing AT is submitted to hypoxiainduced by hypoperfusion at the earliest stages of expansionObservations have proved that AT is poorly oxygenated inthe obese state in humans and rodents due to a reductionin adipose tissue blood flow [25 26] This hypoxia could bea determinant mediator of obesity-induced inflammation inATby activatingmultiple signaling pathways such asHypoxiaInducible Factor-1120572 (HIF-1120572) and Nuclear Factor kappa B(NF-120581B) in adipocytes andmacrophages thereby altering theexpression of many proinflammatory adipokines [27ndash29] Inaddition some studies reported that hypoxia also inducesAT inflammatory responses indirectly by causing adipocytedeath (eg apoptosis and necrosis) and lipolysis [30ndash32]
Endoplasmic Reticulum Stress The endoplasmic reticulum(ER) is responsible for much of a cellrsquos protein synthesis andfolding but it has also a role in sensing cellular stress [33]Obesity results in conditions that increase the demand on theER such as hypoxia and excess of cytokines lipids or glucoseSubsequently the ER stress induces a complex responseknown as the unfolded protein response (UPR) which altersa cellrsquos transcriptional and translational programs to copewith these stressful conditions and resolve the protein-foldingdefect [33] ER stress and the UPR lead to obesity-induced
Journal of Immunology Research 3
inflammation andmetabolic abnormalities by several distinctmechanisms including the activation of JNK-AP1 (Jun N-terminal Kinase-Activator Protein 1) and IKK (I120581B kinase)-NF120581B pathways the induction of the acute-phase responseand the production of reactive oxygen species (ROS) [34 35]
Lipotoxicity Obesity is characterized by a positive energybalance and the classical response to this nutrient oversupplyis AT hypertrophy However the capacity for lipid storage inhypertrophied subcutaneous AT is limited This limited stor-age capacity coupled with the overstimulation of hormone-sensitive lipase leads to massive increase in free fatty acid(FFA) release and accumulation ectopically FFAs are potentligands that activate toll-like receptor (TLR) signaling [36]TLRs are a family of pattern-recognition receptors that playa critical role in the innate immune system by activatingproinflammatory signaling pathways in response tomicrobialpathogens AT expresses a broad spectrum of TLRs [37ndash39] TLR expression in AT is inducible by inflammatorystimulation and linked to downstream activation of NF-120581B orJNK and subsequent release of proinflammatory adipokines[37 38 40] TLR4-deficient mice and C3HHeJ mice (whichhave a loss-of-function mutation in TLR4) are partially pro-tected from fat-induced inflammation and insulin resistancein their visceral AT [41 42] FFAs also increase the infiltrationand activation of macrophages especially the CD11c+ subsetthereby exacerbating their proinflammatory activity [43]
Metabolic Endotoxemia High-fat diet given to mice chroni-cally increased plasma Lipopolysaccharide (LPS) concentra-tion two to three times a threshold that has been definedas metabolic endotoxemia [44] Obese subjects with type2 diabetes have 76 higher circulating LPS than healthycontrols [39] and this high level of LPS decreased significantlyafter surgical weight loss [45] Elevated plasma LPS levelsresult from increased absorption of LPS across the intestinalbarrier triggered by high-fat diet Recently three underlyingmechanisms have been proposed including the changes ingutmicrobiota environment [44] the increased availability ofchylomicrons [46] and the permeability of the gut epithelium[47ndash49] In murine adipocytes LPS initiates inflammationvia TLR4 and induces the secretion of proinflammatorycytokines via downstream activation of NF-120581B or mitogen-activated protein kinases (MAPK) signaling pathways [50]In human adipocytes stimulation by LPS increases release ofTumor Necrosis Factor-alpha (TNF-120572) Interleukin-6 (IL-6)andMonocyte Chemoattractant Protein-1 (MCP-1) byNF-120581Bactivation andupregulates TLR2 expressionNewly expressedTLR2 further amplifies proinflammatory signals in AT [39]
5 microRNAs as a New Mechanism MediatingWhite Adipose Tissue Inflammation
microRNAs (miRNAs) are endogenous sim22 nt RNAs that canbind to the 31015840-untranslated region (31015840-UTR) of target mRNAsto repress mRNA expression at the posttranscriptional levelAs a group miRNAs may directly regulate expression ofover 30 of human and mouse genes and more than 60
of human protein-coding genes have been under selec-tive pressure to maintain pairing to miRNAs [51] SpecificmiRNAs have been implicated in adipocyte differentiationand mature adipocyte function including lipolysis glucose-uptake and insulin sensitivity [8 10] On the other handmiRNAs have been defined as important immunomodulatorsby regulating the differentiation induction and functionof immune cells and the expression of multiple cytokinesin the immune system These findings could shed lighton the possible links between miRNAs and adipose tissueimmunityinflammation
51 Role of microRNAs in Inflammation
Role of microRNAs in Adaptive Immunity Studies conductedby some groups have demonstrated that miRNA regulationis indispensable for stable immune process miRNAs appearto have a key role in the differentiation of B-cells Ablationof whole miRNAs by deletion of Argonaute 2 (Ago2) orDicer impairs pre-B-cell differentiation and the succeedingperipheral B-cell generation [52] Several individual miRNAshave also been reported to be involved in the B-cell biologyFor example overexpression of miR-17sim92 clusters enhancesB-cell proliferation and survival [53] while miR-150 pro-foundly impairs early B-cell differentiation andmature B-cellresponses [54]
miRNAs have been shown to be key regulators of theT-cell lineage induction pathways Deletion of Dicer at anearly stage of T-cell development compromised the survivalof T-cell lineage [55] The best evidence for miRNAs playinga role in specific developmental stages of T-cell differen-tiation is from miR-181 which reduces the number of T-cells in haematopoietic overexpression systems [56] and alsoincreases the sensitivity of T-cell receptor signaling [57]
In addition miRNAs also play pivotal roles in theinduction function andmaintenance of the regulatory T-cell(Treg) lineage miRNAs can enhance thymic and peripheralinduction of Treg cells [55 58] Dicer-deficient Treg cellsalmost lost their suppressive capacity and anergic profile [58]
Role of microRNAs in Innate Immunity Several miRNAs(includingmiR-155 miR-146a miR-21 andmiR-9) have beenconsistently found to be rapidly induced by innate immuneactivation (eg toll-like receptor) indicating that they mayregulate the innate immune response [59] Target predictionanalyses indicate that up to a half of innate immune genescould be under the direct regulation of miRNAs A study on613 genes which regulate immunity utilizing a computationalapproach identified 285 genes as miRNA targets Major tar-gets include transcription factors cofactors and chromatinmodifiers whereas upstream factors such as ligands andreceptors (cytokines and chemokines) were in general pooror nontargets [60]
The mechanisms by which miRNAs regulate cytokineexpression include direct regulation by binding to seedingsites in mRNA 31015840-UTR and indirect regulation Computa-tional analyses predict thatmost cytokines lackmiRNA targetsites in 31015840-UTRs [60] For example of the interleukin genes(IL1-29) studied 9 had predicted miRNA binding sites Out
4 Journal of Immunology Research
of 20 interleukin receptor genes examined only 2 had highprobability miRNA target sites These findings suggest thatthe regulation of cytokine genes by miRNAs occurs oftenvia indirect mechanism Recent studies indicate indeed thatmiRNAs could indirectly regulate cytokine genes via AU-richelements (ARE) located in the mRNA 31015840-UTR by targetingARE-binding proteins ARE are the cis-acting structural RNAmotifs that are important determinants of cytokine messagestability ARE-mediated mRNA degradation is regulated bya number of trans-acting factors called the ARE-bindingproteins (which include tristetraprolin (TTP) AU rich bind-ing factor 1 (AUF1) and members of the Hu protein R(HUR) family) [61] These ARE-binding proteins are heavilypredicted targets of miRNAs [62] Repression of several AREcomponents by miRNAsmay alter the levels of inflammatorycytokines as well as of other immune genes [60 62]
52 Role of microRNAs in White Adipose Tissue Inflammation
Obesity-Induced Inflammation Is Associated with Dysregu-lated Expression of miRNAs in White Adipose Tissue Low-grade inflammation in AT is a key feature of obesity Anumber of miRNAs have been found dysregulated in whiteAT during obesity and closely associated with obesity-related metabolic disorders A recent study identified 21miRNAs which were differentially expressed in epididymalAT between lean mice fed a standard diet and mice renderedobese by a high-fat diet [63] Ortega et al performedmiRNA array on human subcutaneous AT 50 of the 799miRNAs tested (62) significantly differed between obese(119899 = 9) and lean (119899 = 6) subjects [64] Among these 50miRNAs 17 were highly correlated with BMI and metabolicparameters (fasting glucose andor triglycerides) [64] Thesedata are concordant with those obtained in overweight orobese subjects by Kloting et al They showed significantcorrelations between the expression of selected miRNAsand both AT morphology and key metabolic parametersincluding visceral fat area HbA1c fasting plasma glucoseand circulating leptin adiponectin and IL-6 [65] Anothertwo miRNAs (miR-17ndash5p and miR-132) were significantlydecreased in the omental fat and the circulation of obesesubjects [66] A very recent study defined a set of elevenadipocyte-specific miRNAs downregulated in obese subjectsas all of them were concomitantly altered in both the whiteAT and the isolated adipocytes [67] Moreover inflammationper se could alter white AT miRNA levels Xie et al showedthat TNF-120572 treatment of 3T3-L1 adipocytes mimicked thechanges of miRNA expression observed in AT of obese mice[68] With obesity and inflammation being so intrinsicallyassociated the dysregulated expression ofmiRNAs in inflam-matory adipocytes highly suggests the particular importanceof miRNAs in obesity-induced inflammation
Individual miRNAs Regulate White Adipose Tissue Inflamma-tion Indeed a few individual miRNAs have been reportedas playing a crucial role in the inflammatory state of ATso far (Figure 1) Expression of miR-221 and miR-222 hasbeen positively correlated to TNF-120572 and negatively corre-lated to ApN expression in white AT of mice [69] Yet
the direct effects of both miRNAs on these pro- or anti-inflammatory adipokines are still unraveled miR-132 whichwas downregulated in human obese omental fat [66] hasbeen reported to activate NF-120581B and the transcription of IL-8 and MCP-1 in primary human preadipocytes and in invitro differentiated adipocytes [70] Zhuang et al demon-strated that miR-223 played a crucial role in modulatingmacrophage polarization in a pattern that protects micefrom diet-induced AT inflammation and systemic insulinresistance Consequently miR-223 suppressed the infiltrationof proinflammatory M1 ldquoclassically activatedrdquo macrophagesby targeting Pknox1 in vitro while miR-223 deficient micefed a high-fat diet exhibited increased AT inflammationcharacterized by enhanced proinflammatory activation ofmacrophages [71] The group of Brichard has identifiedseveral miRNAs which were regulated by adiponectin (apromising anti-inflammatory adipokine) in white AT in vivoThe miR883b-5p which was upregulated by adiponectin anddownregulated in human obesity repressed the LPS-bindingprotein (LBP) andTLR4 signaling acting therefore as amajormediator of the anti-inflammatory action of ApN MoreovermiR883b-silencing in the de novo AT formed from in vivodifferentiation of preadipocytes also induced LBP productionand tissue inflammation [72] Very recently Arner et aloverexpressed individual miRNAs (which have been defineddownregulated inAT in human obesity) in human adipocytesdifferentiated in vitro They found that a set of nine miRNAssignificantly reduced the secretion of chemokine (C-Cmotif)ligand 2 (CCL2) which is an initiator of AT inflammationby attracting the migration of inflammatory cells into thetissue Among these miRNAs only miR-126 was predictedby in silico analysis and confirmed by luciferase reporterassay in 3T3-L1 cells to bind directly to the 31015840-UTR of CCL2whereas miR-193b affected indirectly the CCL2 productionthrough downregulating the transcription factors (TFs) ofCCL2 (RELB STAT6 and ETS1) [67] Thus miRNAs maymediate AT inflammation by regulating either the activationof macrophages or the production of adipokines The mech-anism by which miRNAs regulate adipokines in AT seemsvia direct and indirectmechanismsmiRNAsmay act directlyon the target inflammatory adipokines or indirectly by firstregulating the intermediate machinery components like theTFs which in turn control the expression of adipokines
In addition to the innate immunity recent studies havedisclosed the importance of the adaptive immunity in ATinflammationThe subsets of the lymphocyte lineage includ-ing CD8+ and CD4+ T-cells Tregs Natural killer T-cells(NKT) and B-cells have been revealed to infiltrate into ATwhich may orchestrate AT inflammation Lymphocyte infil-tration into AT could even precede macrophage infiltration[73 74] Yet the mechanisms underlying the infiltration oflymphocytes into AT remain largely unknown (Figure 1)Whether miRNAs could play roles in modulating the infil-tration or activation of the lymphocytes in AT may arouseresearch interest and requires to be investigated in the future
miRNAsMediate Intercellular CommunicationMore recentlyadipocytes have been reported to secrete miRNAs in theform of adipocyte-derived microvesicles (ADMs) which
Journal of Immunology Research 5
LymphocytemiRNAs
Pknox1
M1
miRNAMicrovesicle
ApN
miR-884b-5p
LBP
LBP LPS
Adipocyte
miR-132
Macrophage
miR-126
CCL2
miR-193b
miR-223
NF-120581B
IL-8 and MCP-1
RELB STAT6 and ETS1
Figure 1 Scheme for the miRNA-mediated mechanisms in regulating white adipose tissue inflammation The miR-132 activates NF-120581B andthe transcription of downstream adipokines IL-8 and MCP-1 The miR-223 played a crucial role in modulating macrophage polarizationwhich suppresses the infiltration of proinflammatory M1 ldquoclassically activatedrdquo macrophages by targeting Pknox1 The miR883b-5p which isupregulated by ApN represses the expression and secretion of LBP a LPS facilitatorThemiR-126 reduces the production of CCL2 by targetingdirectly the 31015840-UTR of CCL2 while miR-193b inhibited indirectly the CCL2 production through downregulating the transcription factors ofCCL2 (RELB STAT6 and ETS1) The microvesicles released by adipocytes may either go into circulation or transfer into the neighboringadipocytes ormacrophages probably acting as inflammatory communicators between adipocytes macrophages and distant cellsThe arrowsindicate stimulation or activation blunted arrows indicate inhibition ApN adiponectin LBP LPS-binding protein LPS lipopolysaccharideCCL2 chemokine (C-C motif) ligand 2 adapted from [5]
may regulate the function of distant or neighboring cellsOne study found that microvesicles released from cultured3T3-L1 adipocytes harbored 143 miRNAs most of thembeing adipocyte-specific and reflecting the abundance oftheir expression levels in the donor cells Interestinglythe miRNAs-containing microvesicles were transported intocultured macrophages which were incubated with ADMscontaining-medium for 24 h they were also present in ratserum in vivo [75] Yet whether the miRNAs inside thevesicles were functional in their new location remains tobe determined Furthermore another group discovered thatmiRNAs-microvesicle complexes derived from the largerprimary rat adipocytes were transferred into and expressed inthe smaller adipocytes and were involved in the transcriptionof multiple genes for lipid synthesis and cell growth [76]Thus miRNAs could also be important mediators of cellularcrosstalk Since macrophages and hypertrophied adipocytesare the main sources of AT inflammation the miRNA-mediated interactions (adipocytes-macrophages and largeradipocytes-smaller adipocytes)may be novelmechanisms formiRNAs contributing to AT inflammation (Figure 1)
6 Therapeutic Potential of microRNAs inMetabolic Syndrome
The facts that miRNAs are largely dysregulated in obe-sity and that specific miRNAs regulate obesity-associatedinflammation suggests the potential of these molecules astargets for therapeutic intervention in obesity and obesity-related metabolic disorders
61 microRNAs as Diagnostic Tools Recent studies havedemonstrated that miRNAs can be secreted into the circula-tion Chen et al screened serummiRNAs of healthy Chinesesubjects and found over 100 and 91 serum miRNAs in maleand female subjects respectively Yet how these miRNAsmake their way into the circulation is still mysterious [77]New studies have revealed that circulatingmiRNAs can residein microvesicles (exosomes microparticles and apoptoticbodies) and in proteinlipoprotein complexes (high densitylipoprotein (HDL) and Argonaute 2) these formations makethe circulating miRNAs protected and resistant to RNase
6 Journal of Immunology Research
activity and degradation Therefore the levels of miRNAsin serum are remarkable stable reproducible and tissue-specific among individuals This raises the potential of miR-NAs as novel biomarkers formany disordersMultiple reportshave noted the potential use of blood miRNAs as biomarkersfor the obesity cardiovascular diseases atherosclerosis andT2DM Heneghan et al showed that circulating levels ofmiR-17-5p and miR-132 were significantly decreased in obeseindividuals compared to nonobese individuals and reflectedmiRNA expression in omental fat The miRNA expressionin blood and omental fat from obese patients correlatedsignificantly with BMI fasting blood glucose and glycatedhemoglobin [66] Cardiac specific miR-1 miR-208 and miR-499 were induced consistently in blood of patients withmyocardial infarction and strongly correlated to markers ofcardiac damage including troponin T and creatine kinase-MB (CK-MB) activity [78ndash81] CirculatingmiRNAs have alsobeen proposed as highly sensitive biomarkers for T2DM Inthe Bruneck cohort of 822 subjects decreased serum levelsof miR-126 in T2DM emerged as a significant predictorof T2DM and correlated negatively with increasing glucoseintolerance [82]
62 microRNAs as Therapeutic Tools ldquoSilencingrdquo of keymiRNA and ldquoreplacementrdquo of certain tissue-specific miRNAwhose expression is known to be decreased are potentialtherapeutic interventions
Techniques Several techniques have been set up to use thetherapeutic potential of miRNAs miRNAs can be deacti-vated and silenced by anti-miRNA oligonucleotides (AMOs)ldquomiRNA spongesrdquo and ldquomiRNA maskingrdquo [83] AMOsare synthetic antisense oligonucleotides that competitivelyinhibit the interaction between miRNAs and their mRNAtargets The most widely employed types of AMOs are 21015840-O-methyl AMO 21015840-O-methoxyethyl AMOs and LockedNucleic Acids (LNAs) [84] Given that miRNAs have beenobserved to function often in clusters in pathological pro-cesses the ldquomiRNA spongesrdquo have been developed to knock-down multiple miRNAs This technique requires a vectorencoding large quantities of transcripts (anti-miRNAs)whichdisplay numerous and tandem binding sites for a cluster ofmiRNAs of interest The ldquomiRNA maskingrdquo is an alternativemiRNA-knockdown strategy to the AMO approach withthe advantage of targeting miRNAs in a gene-specific man-ner Unlike AMO which binds to the endogenous miRNAdirectly a miRNA mask binds to the miRNArsquos binding sitelocated in the 31015840-UTR of its target mRNA thereby avoidingoff-target effects [83]
Conversely with regard to those miRNAs with decreasedexpression in disease the fundamental principle strategy isto restore their expression This can be achieved throughmiRNA mimicry or plasmidviral vector-encoded miRNAreplacement miRNA mimics are small chemically altereddouble-stranded RNA molecules that imitate endogenousmiRNAs [83] Plasmidviral vectors encoding miRNAs areencouraging strategies to replace miRNA in vivo with goodtransduction efficiency and minimal toxicity [85 86]
microRNAs and Therapeutics Since the obesity-inducedinflammation is considered to build the common soil forthe pathogenesis of MS some potential anti-inflammatorytherapies have been investigated Several drugs in currentclinical practice show the anti-inflammatory properties theperoxisome proliferator-activated receptor (PPAR) familyand nonacetylated salicylates Other strategies have beenexplored by targeting the cytokines and chemokines or theirreceptors (eg the anti-TNF or anti-CCR2 (chemokine (CndashCmotif) receptor 2) therapies) [2] Although there have beensome encouraging results it is likely that the benefits of theseapproaches are limited [87 88] Therefore the developmentof miRNA-based therapeutics has raised interest in this field
To date the greatest efforts have been made in exploringthe potential application of miRNAs therapeutics in cancerand liver disorders Gain or loss of function of individualmiRNAs have been reported in almost every solid andhematological cancer with therapeutic suppressing effectin tumor cell proliferation progression of tumors and themetastatic process [83] The liver specific miR-122 has beenproved to regulate cholesterol biosynthesis and hepatitis Cvirus replication Silencing of miR-122 by intraperitonealadministration of high affinity LNAanti-miR-122 has resultedin dose-dependent lowering of plasma cholesterol in miceand nonhuman primates (monkeys)This was achieved with-out significant adverse reactions or hepatic toxicity [89 90]
Likewise induction or reduction of miRNA expressionthat is known to be underupper-expressed and involved inAT inflammation using viral or liposomal delivery of tissue-specific miRNAs to affected AT could potentially result inrestoration of immune balance to the AT Yet the applicationof miRNAs therapeutics in AT in vivo is poorly investigatedbecause fat cells are insusceptible to selective transfer ofexogenous nucleotide due to the containing lipid dropletsof adipocytes Encouragingly the discovery that miRNAscould be secreted by adipocytes and accepted by neighboringadipocytes or macrophages in the form of adipocyte-derivedmicrovesicles [75 76] raises the hope that it could be possibleto import the targeting miRNAs into AT in vivo by themanner of microvesicles This manner could construct themicrovesicles containing specific miRNAs and inject theconstructed microvesicles into white AT
63 Potential Challenges in microRNA-Based TherapyAlthough significant advances have been made in miRNA-based therapy various challenges remain to be overcomebefore clinical use First one individual miRNA mayhave multiple potential targets which may coordinate orantagonize each otherrsquos functions In addition miRNA-target interactions depend not only on the sequence ofthe target site but also on the cellular context in which theinteractions occurThis complexity explains the difficulties inpredicting the spectrum of side effects and toxicity profileswhich may be associated with miRNA-based therapeuticsSecond replacement of miRNA may potentially lead tooversaturation of endogenous miRNAs pathway High levelsof exogenous miRNA can compete with the endogenousmiRNA biogenesis processing leading to cell toxicity
Journal of Immunology Research 7
[91 92] Another critical issue is the site-specific safeeffective and repeated delivery of miRNA in vivo Thesystemic intravenous delivery of viral or liposomal-mediatedmiRNA oligonucleotides have been well achieved in cancerand liver disease However targeting certain miRNAs inother specific tissues is poorly achieved due to the invasivenature of access for repeat delivery and lack of cell-specificup-take Restoration of tissue-specific miRNA expressioncould be a more rational approach for managing localabnormalities with more efficiency and precision and withless systemic side effects [83 92]
7 Conclusion
Evidence has provided that miRNAs are largely dysregulatedin white AT in human obesity specific miRNAs regulate thepolarization ofmacrophages and the expression and secretionof adipokines in white AT a subset ofmiRNAs could be pack-aged into adipocyte-derived microvesicles and delivered intoblood or neighboring cellsmediating intercellular communi-cation Several reports strongly support the roles of miRNAsas important mediators in AT inflammation Furthermorewith the recent discovery that miRNAs are secreted intocirculation in a stable manner and correlated closely withcrucial metabolic parameters the plasma miRNA profilescould therefore be used as attractive biomarkers for theMS Finally the establishment of miRNAs-based techniquesallows exploring further fascinating roles of miRNAs andtheir applications in the management of MS Despite hugechallenges lying in miRNA-targeting therapeutics especiallyin developing safe and effective delivery of targeting miRNAsin specific tissues in vivo the development of miRNA-basedtherapeutics for MS is warranted because of the severeproblem that metabolic disease poses to society
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This work was supported by grants from the National KeyClinical Department Construction Project
References
[1] Y-H Tseng A M Cypess and C R Kahn ldquoCellular bioen-ergetics as a target for obesity therapyrdquo Nature Reviews DrugDiscovery vol 9 no 6 pp 465ndash482 2010
[2] G S Hotamisligil ldquoInflammation and metabolic disordersrdquoNature vol 444 no 7121 pp 860ndash867 2006
[3] K L Spalding E Arner P O Westermark et al ldquoDynamics offat cell turnover in humansrdquoNature vol 453 no 7196 pp 783ndash787 2008
[4] P Jiao andH Xu ldquoAdipose inflammation cause or consequenceof obesity-related insulin resistancerdquo Diabetes Metabolic Syn-drome and Obesity vol 1 pp 25ndash31 2008
[5] EMaury and SM Brichard ldquoAdipokine dysregulation adiposetissue inflammation and metabolic syndromerdquo Molecular andCellular Endocrinology vol 314 no 1 pp 1ndash16 2010
[6] L-F Lu and A Liston ldquoMicroRNA in the immune systemmicroRNA as an immune systemrdquo Immunology vol 127 no 3pp 291ndash298 2009
[7] D Sayed and M Abdellatif ldquoMicrornas in development anddiseaserdquo Physiological Reviews vol 91 no 3 pp 827ndash887 2011
[8] H M Heneghan N Miller and M J Kerin ldquoRole of microR-NAs in obesity and the metabolic syndromerdquo Obesity Reviewsvol 11 no 5 pp 354ndash361 2010
[9] M Hulsmans D De Keyzer and P Holvoet ldquoMicroRNAsregulating oxidative stress and inflammation in relation toobesity and atherosclerosisrdquo The FASEB Journal vol 25 no 8pp 2515ndash2527 2011
[10] R Alexander H Lodish and L Sun ldquoMicroRNAs in adipoge-nesis and as therapeutic targets for obesityrdquo Expert Opinion onTherapeutic Targets vol 15 no 5 pp 623ndash636 2011
[11] C N Lumeng and A R Saltiel ldquoInflammatory links betweenobesity andmetabolic diseaserdquoThe Journal of Clinical Investiga-tion vol 121 no 6 pp 2111ndash2117 2011
[12] Y Chen J Zhu P Y Lum et al ldquoVariations in DNA elucidatemolecular networks that cause diseaserdquo Nature vol 452 no7186 pp 429ndash435 2008
[13] V Emilsson G Thorleifsson B Zhang et al ldquoGenetics of geneexpression and its effect on diseaserdquo Nature vol 452 no 7186pp 423ndash428 2008
[14] V Varma A Yao-Borengasser N Rasouli et al ldquoMuscle inflam-matory response and insulin resistance synergistic interactionbetween macrophages and fatty acids leads to impaired insulinactionrdquoThe American Journal of Physiology Endocrinology andMetabolism vol 296 no 6 pp E1300ndashE1310 2009
[15] WHuang AMetlakunta N Dedousis et al ldquoDepletion of liverkupffer cells prevents the development of diet-induced hepaticsteatosis and insulin resistancerdquoDiabetes vol 59 no 2 pp 347ndash357 2010
[16] J P Thaler C-X Yi E A Schur et al ldquoObesity is associatedwith hypothalamic injury in rodents and humansrdquoThe Journalof Clinical Investigation vol 122 no 1 pp 153ndash162 2012
[17] Y-M Kang Y Ma J-P Zheng et al ldquoBrain nuclear factor-kappa B activation contributes to neurohumoral excitation inangiotensin II-induced hypertensionrdquoCardiovascular Researchvol 82 no 3 pp 503ndash512 2009
[18] V C Calegari A S Torsoni E C Vanzela et al ldquoInflammationof the hypothalamus leads to defective pancreatic islet functionrdquoThe Journal of Biological Chemistry vol 286 no 15 pp 12870ndash12880 2011
[19] S Purkayastha H Zhang G Zhang Z Ahmed Y Wangand D Cai ldquoNeural dysregulation of peripheral insulin actionand blood pressure by brain endoplasmic reticulum stressrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 108 no 7 pp 2939ndash2944 2011
[20] M E Trujillo and P E Scherer ldquoAdipose tissue-derived factorsimpact on health and diseaserdquo Endocrine Reviews vol 27 no 7pp 762ndash778 2006
[21] E Maury K Ehala-Aleksejev Y Guiot R Detry A Vanden-hooft and S M Brichard ldquoAdipokines oversecreted by omentaladipose tissue in human obesityrdquo The American Journal ofPhysiology Endocrinology and Metabolism vol 293 no 3 ppE656ndashE665 2007
8 Journal of Immunology Research
[22] V Bourlier A Zakaroff-Girard A Miranville et alldquoRemodeling phenotype of human subcutaneous adiposetissue macrophagesrdquo Circulation vol 117 no 6 pp 806ndash8152008
[23] I M Faust P R Johnson J S Stern and J Hirsch ldquoDiet-induced adipocyte number increase in adult rats a new modelof obesityrdquo The American Journal of Physiology vol 235 no 3pp E279ndashE286 1978
[24] C Henegar J Tordjman V Achard et al ldquoAdipose tissuetranscriptomic signature highlights the pathological relevanceof extracellular matrix in human obesityrdquo Genome Biology vol9 article R14 2008
[25] B Kabon A Nagele D Reddy et al ldquoObesity decreasesperioperative tissue oxygenationrdquo Anesthesiology vol 100 no2 pp 274ndash280 2004
[26] K A Virtanen P Lonnroth R Parkkola et al ldquoGlucose uptakeand perfusion in subcutaneous and visceral adipose tissueduring insulin stimulation in nonobese and obese humansrdquoJournal of Clinical Endocrinology and Metabolism vol 87 no8 pp 3902ndash3910 2002
[27] N Hosogai A Fukuhara K Oshima et al ldquoAdipose tissuehypoxia in obesity and its impact on adipocytokine dysregula-tionrdquo Diabetes vol 56 no 4 pp 901ndash911 2007
[28] J Ye Z Gao J Yin andQHe ldquoHypoxia is a potential risk factorfor chronic inflammation and adiponectin reduction in adiposetissue of obob and dietary obese micerdquo The American Journalof Physiology Endocrinology andMetabolism vol 293 no 4 ppE1118ndashE1128 2007
[29] B Wang I S Wood and P Trayhurn ldquoDysregulation of theexpression and secretion of inflammation-related adipokines byhypoxia in human adipocytesrdquoPflugers Archiv European Journalof Physiology vol 455 no 3 pp 479ndash492 2007
[30] J Yin Z GaoQHeD Zhou ZGuo and J Ye ldquoRole of hypoxiain obesity-induced disorders of glucose and lipidmetabolism inadipose tissuerdquoTheAmerican Journal of Physiology Endocrinol-ogy and Metabolism vol 296 no 2 pp E333ndashE342 2009
[31] Z Yun H L Maecker R S Johnson and A J GiaccialdquoInhibition of PPAR1205742 gene expression by the HIF-1-regulatedgene DEC1Stra13 a mechanism for regulation of adipogenesisby hypoxiardquo Developmental Cell vol 2 no 3 pp 331ndash341 2002
[32] L Liu T P Cash R G Jones B Keith C B Thompson andM C Simon ldquoHypoxia-induced energy stress regulates mRNAtranslation and cell growthrdquo Molecular Cell vol 21 no 4 pp521ndash531 2006
[33] K Zhang and R J Kaufman ldquoFrom endoplasmic-reticulumstress to the inflammatory responserdquoNature vol 454 no 7203pp 455ndash462 2008
[34] B P Tu and J S Weissman ldquoOxidative protein folding ineukaryotes mechanisms and consequencesrdquo Journal of CellBiology vol 164 no 3 pp 341ndash346 2004
[35] C M Haynes E A Titus and A A Cooper ldquoDegradation ofmisfolded proteins prevents ER-derived oxidative stress and celldeathrdquoMolecular Cell vol 15 no 5 pp 767ndash776 2004
[36] A Schaeffler P Gross R Buettner et al ldquoFatty acid-inducedinduction of Toll-like receptor-4nuclear factor-120581B pathway inadipocytes links nutritional signalling with innate immunityrdquoImmunology vol 126 no 2 pp 233ndash245 2009
[37] A Kopp C BuechlerMNeumeier et al ldquoInnate immunity andadipocyte function ligand-specific activation of multiple toll-like receptors modulates cytokine adipokine and chemokinesecretion in adipocytesrdquo Obesity vol 17 no 4 pp 648ndash6562009
[38] O I Vitseva K Tanriverdi T T Tchkonia et al ldquoInducibletoll-like receptor and NF-120581B regulatory pathway expression inhuman adipose tissuerdquoObesity vol 16 no 5 pp 932ndash937 2008
[39] S J Creely P G McTernan C M Kusminski et alldquoLipopolysaccharide activates an innate immune systemresponse in human adipose tissue in obesity and type 2diabetesrdquo The American Journal of Physiology Endocrinologyand Metabolism vol 292 no 3 pp E740ndashE747 2007
[40] A Schaffler J Scholmerich and B Salzberger ldquoAdipose tissueas an immunological organ toll-like receptors C1qTNFs andCTRPsrdquoTrends in Immunology vol 28 no 9 pp 393ndash399 2007
[41] J E Davis N K Gabler J Walker-Daniels and M E SpurlockldquoTlr-4 deficiency selectively protects against obesity induced bydiets high in saturated fatrdquoObesity vol 16 no 6 pp 1248ndash12552008
[42] T Suganami T Mieda M Itoh Y Shimoda Y Kamei and YOgawa ldquoAttenuation of obesity-induced adipose tissue inflam-mation in C3HHeJ mice carrying a toll-like receptor 4 muta-tionrdquo Biochemical and Biophysical Research Communicationsvol 354 no 1 pp 45ndash49 2007
[43] X Prieur T Roszer and M Ricote ldquoLipotoxicity inmacrophages evidence from diseases associated with themetabolic syndromerdquo Biochimica et Biophysica Acta Molecularand Cell Biology of Lipids vol 1801 no 3 pp 327ndash337 2010
[44] P D Cani J AmarM A Iglesias et al ldquoMetabolic endotoxemiainitiates obesity and insulin resistancerdquo Diabetes vol 56 no 7pp 1761ndash1772 2007
[45] S V Monte J A Caruana H Ghanim et al ldquoReduction inendotoxemia oxidative and inflammatory stress and insulinresistance after Roux-en-Y gastric bypass surgery in patientswith morbid obesity and type 2 diabetes mellitusrdquo Surgery vol151 no 4 pp 587ndash593 2012
[46] S Ghoshal J Witta J Zhong W de Villiers and E EckhardtldquoChylomicrons promote intestinal absorption of lipopolysac-charidesrdquo Journal of Lipid Research vol 50 no 1 pp 90ndash972009
[47] P D Cani R Bibiloni C Knauf et al ldquoChanges in gut micro-biota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in micerdquo Diabetesvol 57 no 6 pp 1470ndash1481 2008
[48] G GMuccioli D Naslain F Backhed et al ldquoThe endocannabi-noid system links gut microbiota to adipogenesisrdquo MolecularSystems Biology vol 6 article 392 2010
[49] P D Cani S Possemiers T Van De Wiele et al ldquoChanges ingut microbiota control inflammation in obese mice througha mechanism involving GLP-2-driven improvement of gutpermeabilityrdquo Gut vol 58 no 8 pp 1091ndash1103 2009
[50] A Schaffler and J Scholmerich ldquoInnate immunity and adiposetissue biologyrdquoTrends in Immunology vol 31 no 6 pp 228ndash2352010
[51] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[52] D OrsquoCarroll I Mecklenbrauker P P Das et al ldquoA Slicer-independent role for Argonaute 2 in hematopoiesis and themicroRNA pathwayrdquoGenes and Development vol 21 no 16 pp1999ndash2004 2007
[53] C Xiao L Srinivasan D P Calado et al ldquoLymphoproliferativedisease and autoimmunity in mice with increased miR-17-92expression in lymphocytesrdquo Nature Immunology vol 9 no 4pp 405ndash414 2008
Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Journal of Immunology Research
2 Obesity Inflammationand Metabolic Syndrome
21 Low-Grade Inflammation as a Key Feature of ObesityDuring the past decade it became clear that inflammationis a key feature of obesity [2] The inflammatory responsetriggered by obesity involves many components of the clas-sical inflammatory response to pathogens and includes theincreases in circulating inflammatory cytokines and acute-phase proteins (eg C-reactive protein) recruitment ofleukocytes to inflamed tissues activation of tissue leuko-cytes and generation of reparative tissue responses (egfibrosis) However the nature of obesity-induced inflam-mation referred to as ldquometainflammationrdquo (metabolicallytriggered inflammation) is unique compared with otherinflammatory paradigms (eg infection and autoimmunedisease) in several key aspects First the chronic nature ofobesity produces a low-grade activation of the innate immunesystem that affects metabolic homeostasis over time Secondchildhood obesity may place individuals at risk for lifelonginflammation since inflammatory markers are elevated inobese children as young as 3 years old Third this chronicinflammation is composed of recurrent acute episodes ofnutrition-related immune activation induced by nutrientavailability (fasting or high-fat meals) This fluctuation maybe associatedwith the induction of pro- or anti-inflammatorymediators Finally obesity-induced chronic inflammation isunique by its multiorgan involvement [11]
22 Obesity-Induced Inflammation Linking Obesity to Meta-bolic Syndrome The metabolic syndrome (MS) refers to theclustering of cardiovascular risk factors that include centralobesity hyperglycemia dyslipidemia and hypertension Theultimate importance of this cluster is to identify individuals athigh risk of both T2DM and cardiovascular disease Growingevidence implicates obesity-induced inflammation as animportant mechanism linking obesity to the MS in metabol-ically active organs Assessment of gene expression networksin obese AT has identified a robust pattern of overexpressedinflammatory genes associated with metabolic disease [1213] Multiple inflammatory mediators abnormally secretedby AT and the crosstalk between immune and metaboliccells can impair insulin signaling and induce oxidativestress and endothelial dysfunction leading to systemicinsulin resistance and cardiovascular disease [5] Deregulatedmacrophage-myocyte and macrophage-hepatocyte signalingcan impair insulin sensitivity as well [14 15] Hypothalamicinflammation which is induced very rapidly by a high-fatdiet [16]may provoke hyperphagia and has been documentedto impair insulin release from 120573 cells peripheral insulinaction and potentiate hypertension [17ndash19] Thus chronicexcess of nutrients such as lipids and glucose may simultane-ously trigger inflammatory responses which further disruptmetabolic function enhancing stress and inflammationSuch a vicious cycle is identified as a mechanism leadingto further metabolic deterioration Therefore breaking thisvicious circle by resetting the immunological balance inobesity is a crucial approach for the management of MS
3 The Fundamental Role of White AdiposeTissue in Obesity-Induced Inflammation
Obesity is characterized by excessive expansion of whiteadipose tissue (AT) which has been thought to be theprimary site for the initiation of obesity-associated inflam-mation Although ATrsquos principal function deals with energystorage it serves as an active secretory organ as well Anumber of bioactive peptides or proteins collectively namedldquoadipokinesrdquo are produced and secreted by fat andor nonfatcells of white AT They act in an autocrineparacrine mannerto regulate local AT function and also act in an endocrinemanner to influence the functions of distant tissues such asliver skeletal muscle and cardiovascular and central nervoussystems [5 20]
In obesity white AT is remodeled dynamically by adi-pocyte hypertrophy (increased size) hyperplasia (increasednumber) immune cell infiltration endothelial cell over-activation and extracellular matrix overproduction [21ndash24] This remodeling may trigger metabolic and hypoxicstress resulting in activation of multiple inflammatorysignaling pathways ultimately leading to dysregulation ofnumerous adipokines including proinflammatory cytokineschemokines growth factors acute-phase proteins andcomplement-like factors Virtually all known adipokines aredysregulated in obesity via multiple mechanisms Such aderegulation is the main feature of AT low-grade inflamma-tion and contributes to the pathogenesis of MS [5]
4 Molecular Mechanisms Mediating WhiteAdipose Tissue Inflammation
Several prominent mechanisms have been highlighted thatcoordinately mediate white AT inflammation
Hypoxia In obesity the growing AT is submitted to hypoxiainduced by hypoperfusion at the earliest stages of expansionObservations have proved that AT is poorly oxygenated inthe obese state in humans and rodents due to a reductionin adipose tissue blood flow [25 26] This hypoxia could bea determinant mediator of obesity-induced inflammation inATby activatingmultiple signaling pathways such asHypoxiaInducible Factor-1120572 (HIF-1120572) and Nuclear Factor kappa B(NF-120581B) in adipocytes andmacrophages thereby altering theexpression of many proinflammatory adipokines [27ndash29] Inaddition some studies reported that hypoxia also inducesAT inflammatory responses indirectly by causing adipocytedeath (eg apoptosis and necrosis) and lipolysis [30ndash32]
Endoplasmic Reticulum Stress The endoplasmic reticulum(ER) is responsible for much of a cellrsquos protein synthesis andfolding but it has also a role in sensing cellular stress [33]Obesity results in conditions that increase the demand on theER such as hypoxia and excess of cytokines lipids or glucoseSubsequently the ER stress induces a complex responseknown as the unfolded protein response (UPR) which altersa cellrsquos transcriptional and translational programs to copewith these stressful conditions and resolve the protein-foldingdefect [33] ER stress and the UPR lead to obesity-induced
Journal of Immunology Research 3
inflammation andmetabolic abnormalities by several distinctmechanisms including the activation of JNK-AP1 (Jun N-terminal Kinase-Activator Protein 1) and IKK (I120581B kinase)-NF120581B pathways the induction of the acute-phase responseand the production of reactive oxygen species (ROS) [34 35]
Lipotoxicity Obesity is characterized by a positive energybalance and the classical response to this nutrient oversupplyis AT hypertrophy However the capacity for lipid storage inhypertrophied subcutaneous AT is limited This limited stor-age capacity coupled with the overstimulation of hormone-sensitive lipase leads to massive increase in free fatty acid(FFA) release and accumulation ectopically FFAs are potentligands that activate toll-like receptor (TLR) signaling [36]TLRs are a family of pattern-recognition receptors that playa critical role in the innate immune system by activatingproinflammatory signaling pathways in response tomicrobialpathogens AT expresses a broad spectrum of TLRs [37ndash39] TLR expression in AT is inducible by inflammatorystimulation and linked to downstream activation of NF-120581B orJNK and subsequent release of proinflammatory adipokines[37 38 40] TLR4-deficient mice and C3HHeJ mice (whichhave a loss-of-function mutation in TLR4) are partially pro-tected from fat-induced inflammation and insulin resistancein their visceral AT [41 42] FFAs also increase the infiltrationand activation of macrophages especially the CD11c+ subsetthereby exacerbating their proinflammatory activity [43]
Metabolic Endotoxemia High-fat diet given to mice chroni-cally increased plasma Lipopolysaccharide (LPS) concentra-tion two to three times a threshold that has been definedas metabolic endotoxemia [44] Obese subjects with type2 diabetes have 76 higher circulating LPS than healthycontrols [39] and this high level of LPS decreased significantlyafter surgical weight loss [45] Elevated plasma LPS levelsresult from increased absorption of LPS across the intestinalbarrier triggered by high-fat diet Recently three underlyingmechanisms have been proposed including the changes ingutmicrobiota environment [44] the increased availability ofchylomicrons [46] and the permeability of the gut epithelium[47ndash49] In murine adipocytes LPS initiates inflammationvia TLR4 and induces the secretion of proinflammatorycytokines via downstream activation of NF-120581B or mitogen-activated protein kinases (MAPK) signaling pathways [50]In human adipocytes stimulation by LPS increases release ofTumor Necrosis Factor-alpha (TNF-120572) Interleukin-6 (IL-6)andMonocyte Chemoattractant Protein-1 (MCP-1) byNF-120581Bactivation andupregulates TLR2 expressionNewly expressedTLR2 further amplifies proinflammatory signals in AT [39]
5 microRNAs as a New Mechanism MediatingWhite Adipose Tissue Inflammation
microRNAs (miRNAs) are endogenous sim22 nt RNAs that canbind to the 31015840-untranslated region (31015840-UTR) of target mRNAsto repress mRNA expression at the posttranscriptional levelAs a group miRNAs may directly regulate expression ofover 30 of human and mouse genes and more than 60
of human protein-coding genes have been under selec-tive pressure to maintain pairing to miRNAs [51] SpecificmiRNAs have been implicated in adipocyte differentiationand mature adipocyte function including lipolysis glucose-uptake and insulin sensitivity [8 10] On the other handmiRNAs have been defined as important immunomodulatorsby regulating the differentiation induction and functionof immune cells and the expression of multiple cytokinesin the immune system These findings could shed lighton the possible links between miRNAs and adipose tissueimmunityinflammation
51 Role of microRNAs in Inflammation
Role of microRNAs in Adaptive Immunity Studies conductedby some groups have demonstrated that miRNA regulationis indispensable for stable immune process miRNAs appearto have a key role in the differentiation of B-cells Ablationof whole miRNAs by deletion of Argonaute 2 (Ago2) orDicer impairs pre-B-cell differentiation and the succeedingperipheral B-cell generation [52] Several individual miRNAshave also been reported to be involved in the B-cell biologyFor example overexpression of miR-17sim92 clusters enhancesB-cell proliferation and survival [53] while miR-150 pro-foundly impairs early B-cell differentiation andmature B-cellresponses [54]
miRNAs have been shown to be key regulators of theT-cell lineage induction pathways Deletion of Dicer at anearly stage of T-cell development compromised the survivalof T-cell lineage [55] The best evidence for miRNAs playinga role in specific developmental stages of T-cell differen-tiation is from miR-181 which reduces the number of T-cells in haematopoietic overexpression systems [56] and alsoincreases the sensitivity of T-cell receptor signaling [57]
In addition miRNAs also play pivotal roles in theinduction function andmaintenance of the regulatory T-cell(Treg) lineage miRNAs can enhance thymic and peripheralinduction of Treg cells [55 58] Dicer-deficient Treg cellsalmost lost their suppressive capacity and anergic profile [58]
Role of microRNAs in Innate Immunity Several miRNAs(includingmiR-155 miR-146a miR-21 andmiR-9) have beenconsistently found to be rapidly induced by innate immuneactivation (eg toll-like receptor) indicating that they mayregulate the innate immune response [59] Target predictionanalyses indicate that up to a half of innate immune genescould be under the direct regulation of miRNAs A study on613 genes which regulate immunity utilizing a computationalapproach identified 285 genes as miRNA targets Major tar-gets include transcription factors cofactors and chromatinmodifiers whereas upstream factors such as ligands andreceptors (cytokines and chemokines) were in general pooror nontargets [60]
The mechanisms by which miRNAs regulate cytokineexpression include direct regulation by binding to seedingsites in mRNA 31015840-UTR and indirect regulation Computa-tional analyses predict thatmost cytokines lackmiRNA targetsites in 31015840-UTRs [60] For example of the interleukin genes(IL1-29) studied 9 had predicted miRNA binding sites Out
4 Journal of Immunology Research
of 20 interleukin receptor genes examined only 2 had highprobability miRNA target sites These findings suggest thatthe regulation of cytokine genes by miRNAs occurs oftenvia indirect mechanism Recent studies indicate indeed thatmiRNAs could indirectly regulate cytokine genes via AU-richelements (ARE) located in the mRNA 31015840-UTR by targetingARE-binding proteins ARE are the cis-acting structural RNAmotifs that are important determinants of cytokine messagestability ARE-mediated mRNA degradation is regulated bya number of trans-acting factors called the ARE-bindingproteins (which include tristetraprolin (TTP) AU rich bind-ing factor 1 (AUF1) and members of the Hu protein R(HUR) family) [61] These ARE-binding proteins are heavilypredicted targets of miRNAs [62] Repression of several AREcomponents by miRNAsmay alter the levels of inflammatorycytokines as well as of other immune genes [60 62]
52 Role of microRNAs in White Adipose Tissue Inflammation
Obesity-Induced Inflammation Is Associated with Dysregu-lated Expression of miRNAs in White Adipose Tissue Low-grade inflammation in AT is a key feature of obesity Anumber of miRNAs have been found dysregulated in whiteAT during obesity and closely associated with obesity-related metabolic disorders A recent study identified 21miRNAs which were differentially expressed in epididymalAT between lean mice fed a standard diet and mice renderedobese by a high-fat diet [63] Ortega et al performedmiRNA array on human subcutaneous AT 50 of the 799miRNAs tested (62) significantly differed between obese(119899 = 9) and lean (119899 = 6) subjects [64] Among these 50miRNAs 17 were highly correlated with BMI and metabolicparameters (fasting glucose andor triglycerides) [64] Thesedata are concordant with those obtained in overweight orobese subjects by Kloting et al They showed significantcorrelations between the expression of selected miRNAsand both AT morphology and key metabolic parametersincluding visceral fat area HbA1c fasting plasma glucoseand circulating leptin adiponectin and IL-6 [65] Anothertwo miRNAs (miR-17ndash5p and miR-132) were significantlydecreased in the omental fat and the circulation of obesesubjects [66] A very recent study defined a set of elevenadipocyte-specific miRNAs downregulated in obese subjectsas all of them were concomitantly altered in both the whiteAT and the isolated adipocytes [67] Moreover inflammationper se could alter white AT miRNA levels Xie et al showedthat TNF-120572 treatment of 3T3-L1 adipocytes mimicked thechanges of miRNA expression observed in AT of obese mice[68] With obesity and inflammation being so intrinsicallyassociated the dysregulated expression ofmiRNAs in inflam-matory adipocytes highly suggests the particular importanceof miRNAs in obesity-induced inflammation
Individual miRNAs Regulate White Adipose Tissue Inflamma-tion Indeed a few individual miRNAs have been reportedas playing a crucial role in the inflammatory state of ATso far (Figure 1) Expression of miR-221 and miR-222 hasbeen positively correlated to TNF-120572 and negatively corre-lated to ApN expression in white AT of mice [69] Yet
the direct effects of both miRNAs on these pro- or anti-inflammatory adipokines are still unraveled miR-132 whichwas downregulated in human obese omental fat [66] hasbeen reported to activate NF-120581B and the transcription of IL-8 and MCP-1 in primary human preadipocytes and in invitro differentiated adipocytes [70] Zhuang et al demon-strated that miR-223 played a crucial role in modulatingmacrophage polarization in a pattern that protects micefrom diet-induced AT inflammation and systemic insulinresistance Consequently miR-223 suppressed the infiltrationof proinflammatory M1 ldquoclassically activatedrdquo macrophagesby targeting Pknox1 in vitro while miR-223 deficient micefed a high-fat diet exhibited increased AT inflammationcharacterized by enhanced proinflammatory activation ofmacrophages [71] The group of Brichard has identifiedseveral miRNAs which were regulated by adiponectin (apromising anti-inflammatory adipokine) in white AT in vivoThe miR883b-5p which was upregulated by adiponectin anddownregulated in human obesity repressed the LPS-bindingprotein (LBP) andTLR4 signaling acting therefore as amajormediator of the anti-inflammatory action of ApN MoreovermiR883b-silencing in the de novo AT formed from in vivodifferentiation of preadipocytes also induced LBP productionand tissue inflammation [72] Very recently Arner et aloverexpressed individual miRNAs (which have been defineddownregulated inAT in human obesity) in human adipocytesdifferentiated in vitro They found that a set of nine miRNAssignificantly reduced the secretion of chemokine (C-Cmotif)ligand 2 (CCL2) which is an initiator of AT inflammationby attracting the migration of inflammatory cells into thetissue Among these miRNAs only miR-126 was predictedby in silico analysis and confirmed by luciferase reporterassay in 3T3-L1 cells to bind directly to the 31015840-UTR of CCL2whereas miR-193b affected indirectly the CCL2 productionthrough downregulating the transcription factors (TFs) ofCCL2 (RELB STAT6 and ETS1) [67] Thus miRNAs maymediate AT inflammation by regulating either the activationof macrophages or the production of adipokines The mech-anism by which miRNAs regulate adipokines in AT seemsvia direct and indirectmechanismsmiRNAsmay act directlyon the target inflammatory adipokines or indirectly by firstregulating the intermediate machinery components like theTFs which in turn control the expression of adipokines
In addition to the innate immunity recent studies havedisclosed the importance of the adaptive immunity in ATinflammationThe subsets of the lymphocyte lineage includ-ing CD8+ and CD4+ T-cells Tregs Natural killer T-cells(NKT) and B-cells have been revealed to infiltrate into ATwhich may orchestrate AT inflammation Lymphocyte infil-tration into AT could even precede macrophage infiltration[73 74] Yet the mechanisms underlying the infiltration oflymphocytes into AT remain largely unknown (Figure 1)Whether miRNAs could play roles in modulating the infil-tration or activation of the lymphocytes in AT may arouseresearch interest and requires to be investigated in the future
miRNAsMediate Intercellular CommunicationMore recentlyadipocytes have been reported to secrete miRNAs in theform of adipocyte-derived microvesicles (ADMs) which
Journal of Immunology Research 5
LymphocytemiRNAs
Pknox1
M1
miRNAMicrovesicle
ApN
miR-884b-5p
LBP
LBP LPS
Adipocyte
miR-132
Macrophage
miR-126
CCL2
miR-193b
miR-223
NF-120581B
IL-8 and MCP-1
RELB STAT6 and ETS1
Figure 1 Scheme for the miRNA-mediated mechanisms in regulating white adipose tissue inflammation The miR-132 activates NF-120581B andthe transcription of downstream adipokines IL-8 and MCP-1 The miR-223 played a crucial role in modulating macrophage polarizationwhich suppresses the infiltration of proinflammatory M1 ldquoclassically activatedrdquo macrophages by targeting Pknox1 The miR883b-5p which isupregulated by ApN represses the expression and secretion of LBP a LPS facilitatorThemiR-126 reduces the production of CCL2 by targetingdirectly the 31015840-UTR of CCL2 while miR-193b inhibited indirectly the CCL2 production through downregulating the transcription factors ofCCL2 (RELB STAT6 and ETS1) The microvesicles released by adipocytes may either go into circulation or transfer into the neighboringadipocytes ormacrophages probably acting as inflammatory communicators between adipocytes macrophages and distant cellsThe arrowsindicate stimulation or activation blunted arrows indicate inhibition ApN adiponectin LBP LPS-binding protein LPS lipopolysaccharideCCL2 chemokine (C-C motif) ligand 2 adapted from [5]
may regulate the function of distant or neighboring cellsOne study found that microvesicles released from cultured3T3-L1 adipocytes harbored 143 miRNAs most of thembeing adipocyte-specific and reflecting the abundance oftheir expression levels in the donor cells Interestinglythe miRNAs-containing microvesicles were transported intocultured macrophages which were incubated with ADMscontaining-medium for 24 h they were also present in ratserum in vivo [75] Yet whether the miRNAs inside thevesicles were functional in their new location remains tobe determined Furthermore another group discovered thatmiRNAs-microvesicle complexes derived from the largerprimary rat adipocytes were transferred into and expressed inthe smaller adipocytes and were involved in the transcriptionof multiple genes for lipid synthesis and cell growth [76]Thus miRNAs could also be important mediators of cellularcrosstalk Since macrophages and hypertrophied adipocytesare the main sources of AT inflammation the miRNA-mediated interactions (adipocytes-macrophages and largeradipocytes-smaller adipocytes)may be novelmechanisms formiRNAs contributing to AT inflammation (Figure 1)
6 Therapeutic Potential of microRNAs inMetabolic Syndrome
The facts that miRNAs are largely dysregulated in obe-sity and that specific miRNAs regulate obesity-associatedinflammation suggests the potential of these molecules astargets for therapeutic intervention in obesity and obesity-related metabolic disorders
61 microRNAs as Diagnostic Tools Recent studies havedemonstrated that miRNAs can be secreted into the circula-tion Chen et al screened serummiRNAs of healthy Chinesesubjects and found over 100 and 91 serum miRNAs in maleand female subjects respectively Yet how these miRNAsmake their way into the circulation is still mysterious [77]New studies have revealed that circulatingmiRNAs can residein microvesicles (exosomes microparticles and apoptoticbodies) and in proteinlipoprotein complexes (high densitylipoprotein (HDL) and Argonaute 2) these formations makethe circulating miRNAs protected and resistant to RNase
6 Journal of Immunology Research
activity and degradation Therefore the levels of miRNAsin serum are remarkable stable reproducible and tissue-specific among individuals This raises the potential of miR-NAs as novel biomarkers formany disordersMultiple reportshave noted the potential use of blood miRNAs as biomarkersfor the obesity cardiovascular diseases atherosclerosis andT2DM Heneghan et al showed that circulating levels ofmiR-17-5p and miR-132 were significantly decreased in obeseindividuals compared to nonobese individuals and reflectedmiRNA expression in omental fat The miRNA expressionin blood and omental fat from obese patients correlatedsignificantly with BMI fasting blood glucose and glycatedhemoglobin [66] Cardiac specific miR-1 miR-208 and miR-499 were induced consistently in blood of patients withmyocardial infarction and strongly correlated to markers ofcardiac damage including troponin T and creatine kinase-MB (CK-MB) activity [78ndash81] CirculatingmiRNAs have alsobeen proposed as highly sensitive biomarkers for T2DM Inthe Bruneck cohort of 822 subjects decreased serum levelsof miR-126 in T2DM emerged as a significant predictorof T2DM and correlated negatively with increasing glucoseintolerance [82]
62 microRNAs as Therapeutic Tools ldquoSilencingrdquo of keymiRNA and ldquoreplacementrdquo of certain tissue-specific miRNAwhose expression is known to be decreased are potentialtherapeutic interventions
Techniques Several techniques have been set up to use thetherapeutic potential of miRNAs miRNAs can be deacti-vated and silenced by anti-miRNA oligonucleotides (AMOs)ldquomiRNA spongesrdquo and ldquomiRNA maskingrdquo [83] AMOsare synthetic antisense oligonucleotides that competitivelyinhibit the interaction between miRNAs and their mRNAtargets The most widely employed types of AMOs are 21015840-O-methyl AMO 21015840-O-methoxyethyl AMOs and LockedNucleic Acids (LNAs) [84] Given that miRNAs have beenobserved to function often in clusters in pathological pro-cesses the ldquomiRNA spongesrdquo have been developed to knock-down multiple miRNAs This technique requires a vectorencoding large quantities of transcripts (anti-miRNAs)whichdisplay numerous and tandem binding sites for a cluster ofmiRNAs of interest The ldquomiRNA maskingrdquo is an alternativemiRNA-knockdown strategy to the AMO approach withthe advantage of targeting miRNAs in a gene-specific man-ner Unlike AMO which binds to the endogenous miRNAdirectly a miRNA mask binds to the miRNArsquos binding sitelocated in the 31015840-UTR of its target mRNA thereby avoidingoff-target effects [83]
Conversely with regard to those miRNAs with decreasedexpression in disease the fundamental principle strategy isto restore their expression This can be achieved throughmiRNA mimicry or plasmidviral vector-encoded miRNAreplacement miRNA mimics are small chemically altereddouble-stranded RNA molecules that imitate endogenousmiRNAs [83] Plasmidviral vectors encoding miRNAs areencouraging strategies to replace miRNA in vivo with goodtransduction efficiency and minimal toxicity [85 86]
microRNAs and Therapeutics Since the obesity-inducedinflammation is considered to build the common soil forthe pathogenesis of MS some potential anti-inflammatorytherapies have been investigated Several drugs in currentclinical practice show the anti-inflammatory properties theperoxisome proliferator-activated receptor (PPAR) familyand nonacetylated salicylates Other strategies have beenexplored by targeting the cytokines and chemokines or theirreceptors (eg the anti-TNF or anti-CCR2 (chemokine (CndashCmotif) receptor 2) therapies) [2] Although there have beensome encouraging results it is likely that the benefits of theseapproaches are limited [87 88] Therefore the developmentof miRNA-based therapeutics has raised interest in this field
To date the greatest efforts have been made in exploringthe potential application of miRNAs therapeutics in cancerand liver disorders Gain or loss of function of individualmiRNAs have been reported in almost every solid andhematological cancer with therapeutic suppressing effectin tumor cell proliferation progression of tumors and themetastatic process [83] The liver specific miR-122 has beenproved to regulate cholesterol biosynthesis and hepatitis Cvirus replication Silencing of miR-122 by intraperitonealadministration of high affinity LNAanti-miR-122 has resultedin dose-dependent lowering of plasma cholesterol in miceand nonhuman primates (monkeys)This was achieved with-out significant adverse reactions or hepatic toxicity [89 90]
Likewise induction or reduction of miRNA expressionthat is known to be underupper-expressed and involved inAT inflammation using viral or liposomal delivery of tissue-specific miRNAs to affected AT could potentially result inrestoration of immune balance to the AT Yet the applicationof miRNAs therapeutics in AT in vivo is poorly investigatedbecause fat cells are insusceptible to selective transfer ofexogenous nucleotide due to the containing lipid dropletsof adipocytes Encouragingly the discovery that miRNAscould be secreted by adipocytes and accepted by neighboringadipocytes or macrophages in the form of adipocyte-derivedmicrovesicles [75 76] raises the hope that it could be possibleto import the targeting miRNAs into AT in vivo by themanner of microvesicles This manner could construct themicrovesicles containing specific miRNAs and inject theconstructed microvesicles into white AT
63 Potential Challenges in microRNA-Based TherapyAlthough significant advances have been made in miRNA-based therapy various challenges remain to be overcomebefore clinical use First one individual miRNA mayhave multiple potential targets which may coordinate orantagonize each otherrsquos functions In addition miRNA-target interactions depend not only on the sequence ofthe target site but also on the cellular context in which theinteractions occurThis complexity explains the difficulties inpredicting the spectrum of side effects and toxicity profileswhich may be associated with miRNA-based therapeuticsSecond replacement of miRNA may potentially lead tooversaturation of endogenous miRNAs pathway High levelsof exogenous miRNA can compete with the endogenousmiRNA biogenesis processing leading to cell toxicity
Journal of Immunology Research 7
[91 92] Another critical issue is the site-specific safeeffective and repeated delivery of miRNA in vivo Thesystemic intravenous delivery of viral or liposomal-mediatedmiRNA oligonucleotides have been well achieved in cancerand liver disease However targeting certain miRNAs inother specific tissues is poorly achieved due to the invasivenature of access for repeat delivery and lack of cell-specificup-take Restoration of tissue-specific miRNA expressioncould be a more rational approach for managing localabnormalities with more efficiency and precision and withless systemic side effects [83 92]
7 Conclusion
Evidence has provided that miRNAs are largely dysregulatedin white AT in human obesity specific miRNAs regulate thepolarization ofmacrophages and the expression and secretionof adipokines in white AT a subset ofmiRNAs could be pack-aged into adipocyte-derived microvesicles and delivered intoblood or neighboring cellsmediating intercellular communi-cation Several reports strongly support the roles of miRNAsas important mediators in AT inflammation Furthermorewith the recent discovery that miRNAs are secreted intocirculation in a stable manner and correlated closely withcrucial metabolic parameters the plasma miRNA profilescould therefore be used as attractive biomarkers for theMS Finally the establishment of miRNAs-based techniquesallows exploring further fascinating roles of miRNAs andtheir applications in the management of MS Despite hugechallenges lying in miRNA-targeting therapeutics especiallyin developing safe and effective delivery of targeting miRNAsin specific tissues in vivo the development of miRNA-basedtherapeutics for MS is warranted because of the severeproblem that metabolic disease poses to society
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This work was supported by grants from the National KeyClinical Department Construction Project
References
[1] Y-H Tseng A M Cypess and C R Kahn ldquoCellular bioen-ergetics as a target for obesity therapyrdquo Nature Reviews DrugDiscovery vol 9 no 6 pp 465ndash482 2010
[2] G S Hotamisligil ldquoInflammation and metabolic disordersrdquoNature vol 444 no 7121 pp 860ndash867 2006
[3] K L Spalding E Arner P O Westermark et al ldquoDynamics offat cell turnover in humansrdquoNature vol 453 no 7196 pp 783ndash787 2008
[4] P Jiao andH Xu ldquoAdipose inflammation cause or consequenceof obesity-related insulin resistancerdquo Diabetes Metabolic Syn-drome and Obesity vol 1 pp 25ndash31 2008
[5] EMaury and SM Brichard ldquoAdipokine dysregulation adiposetissue inflammation and metabolic syndromerdquo Molecular andCellular Endocrinology vol 314 no 1 pp 1ndash16 2010
[6] L-F Lu and A Liston ldquoMicroRNA in the immune systemmicroRNA as an immune systemrdquo Immunology vol 127 no 3pp 291ndash298 2009
[7] D Sayed and M Abdellatif ldquoMicrornas in development anddiseaserdquo Physiological Reviews vol 91 no 3 pp 827ndash887 2011
[8] H M Heneghan N Miller and M J Kerin ldquoRole of microR-NAs in obesity and the metabolic syndromerdquo Obesity Reviewsvol 11 no 5 pp 354ndash361 2010
[9] M Hulsmans D De Keyzer and P Holvoet ldquoMicroRNAsregulating oxidative stress and inflammation in relation toobesity and atherosclerosisrdquo The FASEB Journal vol 25 no 8pp 2515ndash2527 2011
[10] R Alexander H Lodish and L Sun ldquoMicroRNAs in adipoge-nesis and as therapeutic targets for obesityrdquo Expert Opinion onTherapeutic Targets vol 15 no 5 pp 623ndash636 2011
[11] C N Lumeng and A R Saltiel ldquoInflammatory links betweenobesity andmetabolic diseaserdquoThe Journal of Clinical Investiga-tion vol 121 no 6 pp 2111ndash2117 2011
[12] Y Chen J Zhu P Y Lum et al ldquoVariations in DNA elucidatemolecular networks that cause diseaserdquo Nature vol 452 no7186 pp 429ndash435 2008
[13] V Emilsson G Thorleifsson B Zhang et al ldquoGenetics of geneexpression and its effect on diseaserdquo Nature vol 452 no 7186pp 423ndash428 2008
[14] V Varma A Yao-Borengasser N Rasouli et al ldquoMuscle inflam-matory response and insulin resistance synergistic interactionbetween macrophages and fatty acids leads to impaired insulinactionrdquoThe American Journal of Physiology Endocrinology andMetabolism vol 296 no 6 pp E1300ndashE1310 2009
[15] WHuang AMetlakunta N Dedousis et al ldquoDepletion of liverkupffer cells prevents the development of diet-induced hepaticsteatosis and insulin resistancerdquoDiabetes vol 59 no 2 pp 347ndash357 2010
[16] J P Thaler C-X Yi E A Schur et al ldquoObesity is associatedwith hypothalamic injury in rodents and humansrdquoThe Journalof Clinical Investigation vol 122 no 1 pp 153ndash162 2012
[17] Y-M Kang Y Ma J-P Zheng et al ldquoBrain nuclear factor-kappa B activation contributes to neurohumoral excitation inangiotensin II-induced hypertensionrdquoCardiovascular Researchvol 82 no 3 pp 503ndash512 2009
[18] V C Calegari A S Torsoni E C Vanzela et al ldquoInflammationof the hypothalamus leads to defective pancreatic islet functionrdquoThe Journal of Biological Chemistry vol 286 no 15 pp 12870ndash12880 2011
[19] S Purkayastha H Zhang G Zhang Z Ahmed Y Wangand D Cai ldquoNeural dysregulation of peripheral insulin actionand blood pressure by brain endoplasmic reticulum stressrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 108 no 7 pp 2939ndash2944 2011
[20] M E Trujillo and P E Scherer ldquoAdipose tissue-derived factorsimpact on health and diseaserdquo Endocrine Reviews vol 27 no 7pp 762ndash778 2006
[21] E Maury K Ehala-Aleksejev Y Guiot R Detry A Vanden-hooft and S M Brichard ldquoAdipokines oversecreted by omentaladipose tissue in human obesityrdquo The American Journal ofPhysiology Endocrinology and Metabolism vol 293 no 3 ppE656ndashE665 2007
8 Journal of Immunology Research
[22] V Bourlier A Zakaroff-Girard A Miranville et alldquoRemodeling phenotype of human subcutaneous adiposetissue macrophagesrdquo Circulation vol 117 no 6 pp 806ndash8152008
[23] I M Faust P R Johnson J S Stern and J Hirsch ldquoDiet-induced adipocyte number increase in adult rats a new modelof obesityrdquo The American Journal of Physiology vol 235 no 3pp E279ndashE286 1978
[24] C Henegar J Tordjman V Achard et al ldquoAdipose tissuetranscriptomic signature highlights the pathological relevanceof extracellular matrix in human obesityrdquo Genome Biology vol9 article R14 2008
[25] B Kabon A Nagele D Reddy et al ldquoObesity decreasesperioperative tissue oxygenationrdquo Anesthesiology vol 100 no2 pp 274ndash280 2004
[26] K A Virtanen P Lonnroth R Parkkola et al ldquoGlucose uptakeand perfusion in subcutaneous and visceral adipose tissueduring insulin stimulation in nonobese and obese humansrdquoJournal of Clinical Endocrinology and Metabolism vol 87 no8 pp 3902ndash3910 2002
[27] N Hosogai A Fukuhara K Oshima et al ldquoAdipose tissuehypoxia in obesity and its impact on adipocytokine dysregula-tionrdquo Diabetes vol 56 no 4 pp 901ndash911 2007
[28] J Ye Z Gao J Yin andQHe ldquoHypoxia is a potential risk factorfor chronic inflammation and adiponectin reduction in adiposetissue of obob and dietary obese micerdquo The American Journalof Physiology Endocrinology andMetabolism vol 293 no 4 ppE1118ndashE1128 2007
[29] B Wang I S Wood and P Trayhurn ldquoDysregulation of theexpression and secretion of inflammation-related adipokines byhypoxia in human adipocytesrdquoPflugers Archiv European Journalof Physiology vol 455 no 3 pp 479ndash492 2007
[30] J Yin Z GaoQHeD Zhou ZGuo and J Ye ldquoRole of hypoxiain obesity-induced disorders of glucose and lipidmetabolism inadipose tissuerdquoTheAmerican Journal of Physiology Endocrinol-ogy and Metabolism vol 296 no 2 pp E333ndashE342 2009
[31] Z Yun H L Maecker R S Johnson and A J GiaccialdquoInhibition of PPAR1205742 gene expression by the HIF-1-regulatedgene DEC1Stra13 a mechanism for regulation of adipogenesisby hypoxiardquo Developmental Cell vol 2 no 3 pp 331ndash341 2002
[32] L Liu T P Cash R G Jones B Keith C B Thompson andM C Simon ldquoHypoxia-induced energy stress regulates mRNAtranslation and cell growthrdquo Molecular Cell vol 21 no 4 pp521ndash531 2006
[33] K Zhang and R J Kaufman ldquoFrom endoplasmic-reticulumstress to the inflammatory responserdquoNature vol 454 no 7203pp 455ndash462 2008
[34] B P Tu and J S Weissman ldquoOxidative protein folding ineukaryotes mechanisms and consequencesrdquo Journal of CellBiology vol 164 no 3 pp 341ndash346 2004
[35] C M Haynes E A Titus and A A Cooper ldquoDegradation ofmisfolded proteins prevents ER-derived oxidative stress and celldeathrdquoMolecular Cell vol 15 no 5 pp 767ndash776 2004
[36] A Schaeffler P Gross R Buettner et al ldquoFatty acid-inducedinduction of Toll-like receptor-4nuclear factor-120581B pathway inadipocytes links nutritional signalling with innate immunityrdquoImmunology vol 126 no 2 pp 233ndash245 2009
[37] A Kopp C BuechlerMNeumeier et al ldquoInnate immunity andadipocyte function ligand-specific activation of multiple toll-like receptors modulates cytokine adipokine and chemokinesecretion in adipocytesrdquo Obesity vol 17 no 4 pp 648ndash6562009
[38] O I Vitseva K Tanriverdi T T Tchkonia et al ldquoInducibletoll-like receptor and NF-120581B regulatory pathway expression inhuman adipose tissuerdquoObesity vol 16 no 5 pp 932ndash937 2008
[39] S J Creely P G McTernan C M Kusminski et alldquoLipopolysaccharide activates an innate immune systemresponse in human adipose tissue in obesity and type 2diabetesrdquo The American Journal of Physiology Endocrinologyand Metabolism vol 292 no 3 pp E740ndashE747 2007
[40] A Schaffler J Scholmerich and B Salzberger ldquoAdipose tissueas an immunological organ toll-like receptors C1qTNFs andCTRPsrdquoTrends in Immunology vol 28 no 9 pp 393ndash399 2007
[41] J E Davis N K Gabler J Walker-Daniels and M E SpurlockldquoTlr-4 deficiency selectively protects against obesity induced bydiets high in saturated fatrdquoObesity vol 16 no 6 pp 1248ndash12552008
[42] T Suganami T Mieda M Itoh Y Shimoda Y Kamei and YOgawa ldquoAttenuation of obesity-induced adipose tissue inflam-mation in C3HHeJ mice carrying a toll-like receptor 4 muta-tionrdquo Biochemical and Biophysical Research Communicationsvol 354 no 1 pp 45ndash49 2007
[43] X Prieur T Roszer and M Ricote ldquoLipotoxicity inmacrophages evidence from diseases associated with themetabolic syndromerdquo Biochimica et Biophysica Acta Molecularand Cell Biology of Lipids vol 1801 no 3 pp 327ndash337 2010
[44] P D Cani J AmarM A Iglesias et al ldquoMetabolic endotoxemiainitiates obesity and insulin resistancerdquo Diabetes vol 56 no 7pp 1761ndash1772 2007
[45] S V Monte J A Caruana H Ghanim et al ldquoReduction inendotoxemia oxidative and inflammatory stress and insulinresistance after Roux-en-Y gastric bypass surgery in patientswith morbid obesity and type 2 diabetes mellitusrdquo Surgery vol151 no 4 pp 587ndash593 2012
[46] S Ghoshal J Witta J Zhong W de Villiers and E EckhardtldquoChylomicrons promote intestinal absorption of lipopolysac-charidesrdquo Journal of Lipid Research vol 50 no 1 pp 90ndash972009
[47] P D Cani R Bibiloni C Knauf et al ldquoChanges in gut micro-biota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in micerdquo Diabetesvol 57 no 6 pp 1470ndash1481 2008
[48] G GMuccioli D Naslain F Backhed et al ldquoThe endocannabi-noid system links gut microbiota to adipogenesisrdquo MolecularSystems Biology vol 6 article 392 2010
[49] P D Cani S Possemiers T Van De Wiele et al ldquoChanges ingut microbiota control inflammation in obese mice througha mechanism involving GLP-2-driven improvement of gutpermeabilityrdquo Gut vol 58 no 8 pp 1091ndash1103 2009
[50] A Schaffler and J Scholmerich ldquoInnate immunity and adiposetissue biologyrdquoTrends in Immunology vol 31 no 6 pp 228ndash2352010
[51] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[52] D OrsquoCarroll I Mecklenbrauker P P Das et al ldquoA Slicer-independent role for Argonaute 2 in hematopoiesis and themicroRNA pathwayrdquoGenes and Development vol 21 no 16 pp1999ndash2004 2007
[53] C Xiao L Srinivasan D P Calado et al ldquoLymphoproliferativedisease and autoimmunity in mice with increased miR-17-92expression in lymphocytesrdquo Nature Immunology vol 9 no 4pp 405ndash414 2008
Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 3
inflammation andmetabolic abnormalities by several distinctmechanisms including the activation of JNK-AP1 (Jun N-terminal Kinase-Activator Protein 1) and IKK (I120581B kinase)-NF120581B pathways the induction of the acute-phase responseand the production of reactive oxygen species (ROS) [34 35]
Lipotoxicity Obesity is characterized by a positive energybalance and the classical response to this nutrient oversupplyis AT hypertrophy However the capacity for lipid storage inhypertrophied subcutaneous AT is limited This limited stor-age capacity coupled with the overstimulation of hormone-sensitive lipase leads to massive increase in free fatty acid(FFA) release and accumulation ectopically FFAs are potentligands that activate toll-like receptor (TLR) signaling [36]TLRs are a family of pattern-recognition receptors that playa critical role in the innate immune system by activatingproinflammatory signaling pathways in response tomicrobialpathogens AT expresses a broad spectrum of TLRs [37ndash39] TLR expression in AT is inducible by inflammatorystimulation and linked to downstream activation of NF-120581B orJNK and subsequent release of proinflammatory adipokines[37 38 40] TLR4-deficient mice and C3HHeJ mice (whichhave a loss-of-function mutation in TLR4) are partially pro-tected from fat-induced inflammation and insulin resistancein their visceral AT [41 42] FFAs also increase the infiltrationand activation of macrophages especially the CD11c+ subsetthereby exacerbating their proinflammatory activity [43]
Metabolic Endotoxemia High-fat diet given to mice chroni-cally increased plasma Lipopolysaccharide (LPS) concentra-tion two to three times a threshold that has been definedas metabolic endotoxemia [44] Obese subjects with type2 diabetes have 76 higher circulating LPS than healthycontrols [39] and this high level of LPS decreased significantlyafter surgical weight loss [45] Elevated plasma LPS levelsresult from increased absorption of LPS across the intestinalbarrier triggered by high-fat diet Recently three underlyingmechanisms have been proposed including the changes ingutmicrobiota environment [44] the increased availability ofchylomicrons [46] and the permeability of the gut epithelium[47ndash49] In murine adipocytes LPS initiates inflammationvia TLR4 and induces the secretion of proinflammatorycytokines via downstream activation of NF-120581B or mitogen-activated protein kinases (MAPK) signaling pathways [50]In human adipocytes stimulation by LPS increases release ofTumor Necrosis Factor-alpha (TNF-120572) Interleukin-6 (IL-6)andMonocyte Chemoattractant Protein-1 (MCP-1) byNF-120581Bactivation andupregulates TLR2 expressionNewly expressedTLR2 further amplifies proinflammatory signals in AT [39]
5 microRNAs as a New Mechanism MediatingWhite Adipose Tissue Inflammation
microRNAs (miRNAs) are endogenous sim22 nt RNAs that canbind to the 31015840-untranslated region (31015840-UTR) of target mRNAsto repress mRNA expression at the posttranscriptional levelAs a group miRNAs may directly regulate expression ofover 30 of human and mouse genes and more than 60
of human protein-coding genes have been under selec-tive pressure to maintain pairing to miRNAs [51] SpecificmiRNAs have been implicated in adipocyte differentiationand mature adipocyte function including lipolysis glucose-uptake and insulin sensitivity [8 10] On the other handmiRNAs have been defined as important immunomodulatorsby regulating the differentiation induction and functionof immune cells and the expression of multiple cytokinesin the immune system These findings could shed lighton the possible links between miRNAs and adipose tissueimmunityinflammation
51 Role of microRNAs in Inflammation
Role of microRNAs in Adaptive Immunity Studies conductedby some groups have demonstrated that miRNA regulationis indispensable for stable immune process miRNAs appearto have a key role in the differentiation of B-cells Ablationof whole miRNAs by deletion of Argonaute 2 (Ago2) orDicer impairs pre-B-cell differentiation and the succeedingperipheral B-cell generation [52] Several individual miRNAshave also been reported to be involved in the B-cell biologyFor example overexpression of miR-17sim92 clusters enhancesB-cell proliferation and survival [53] while miR-150 pro-foundly impairs early B-cell differentiation andmature B-cellresponses [54]
miRNAs have been shown to be key regulators of theT-cell lineage induction pathways Deletion of Dicer at anearly stage of T-cell development compromised the survivalof T-cell lineage [55] The best evidence for miRNAs playinga role in specific developmental stages of T-cell differen-tiation is from miR-181 which reduces the number of T-cells in haematopoietic overexpression systems [56] and alsoincreases the sensitivity of T-cell receptor signaling [57]
In addition miRNAs also play pivotal roles in theinduction function andmaintenance of the regulatory T-cell(Treg) lineage miRNAs can enhance thymic and peripheralinduction of Treg cells [55 58] Dicer-deficient Treg cellsalmost lost their suppressive capacity and anergic profile [58]
Role of microRNAs in Innate Immunity Several miRNAs(includingmiR-155 miR-146a miR-21 andmiR-9) have beenconsistently found to be rapidly induced by innate immuneactivation (eg toll-like receptor) indicating that they mayregulate the innate immune response [59] Target predictionanalyses indicate that up to a half of innate immune genescould be under the direct regulation of miRNAs A study on613 genes which regulate immunity utilizing a computationalapproach identified 285 genes as miRNA targets Major tar-gets include transcription factors cofactors and chromatinmodifiers whereas upstream factors such as ligands andreceptors (cytokines and chemokines) were in general pooror nontargets [60]
The mechanisms by which miRNAs regulate cytokineexpression include direct regulation by binding to seedingsites in mRNA 31015840-UTR and indirect regulation Computa-tional analyses predict thatmost cytokines lackmiRNA targetsites in 31015840-UTRs [60] For example of the interleukin genes(IL1-29) studied 9 had predicted miRNA binding sites Out
4 Journal of Immunology Research
of 20 interleukin receptor genes examined only 2 had highprobability miRNA target sites These findings suggest thatthe regulation of cytokine genes by miRNAs occurs oftenvia indirect mechanism Recent studies indicate indeed thatmiRNAs could indirectly regulate cytokine genes via AU-richelements (ARE) located in the mRNA 31015840-UTR by targetingARE-binding proteins ARE are the cis-acting structural RNAmotifs that are important determinants of cytokine messagestability ARE-mediated mRNA degradation is regulated bya number of trans-acting factors called the ARE-bindingproteins (which include tristetraprolin (TTP) AU rich bind-ing factor 1 (AUF1) and members of the Hu protein R(HUR) family) [61] These ARE-binding proteins are heavilypredicted targets of miRNAs [62] Repression of several AREcomponents by miRNAsmay alter the levels of inflammatorycytokines as well as of other immune genes [60 62]
52 Role of microRNAs in White Adipose Tissue Inflammation
Obesity-Induced Inflammation Is Associated with Dysregu-lated Expression of miRNAs in White Adipose Tissue Low-grade inflammation in AT is a key feature of obesity Anumber of miRNAs have been found dysregulated in whiteAT during obesity and closely associated with obesity-related metabolic disorders A recent study identified 21miRNAs which were differentially expressed in epididymalAT between lean mice fed a standard diet and mice renderedobese by a high-fat diet [63] Ortega et al performedmiRNA array on human subcutaneous AT 50 of the 799miRNAs tested (62) significantly differed between obese(119899 = 9) and lean (119899 = 6) subjects [64] Among these 50miRNAs 17 were highly correlated with BMI and metabolicparameters (fasting glucose andor triglycerides) [64] Thesedata are concordant with those obtained in overweight orobese subjects by Kloting et al They showed significantcorrelations between the expression of selected miRNAsand both AT morphology and key metabolic parametersincluding visceral fat area HbA1c fasting plasma glucoseand circulating leptin adiponectin and IL-6 [65] Anothertwo miRNAs (miR-17ndash5p and miR-132) were significantlydecreased in the omental fat and the circulation of obesesubjects [66] A very recent study defined a set of elevenadipocyte-specific miRNAs downregulated in obese subjectsas all of them were concomitantly altered in both the whiteAT and the isolated adipocytes [67] Moreover inflammationper se could alter white AT miRNA levels Xie et al showedthat TNF-120572 treatment of 3T3-L1 adipocytes mimicked thechanges of miRNA expression observed in AT of obese mice[68] With obesity and inflammation being so intrinsicallyassociated the dysregulated expression ofmiRNAs in inflam-matory adipocytes highly suggests the particular importanceof miRNAs in obesity-induced inflammation
Individual miRNAs Regulate White Adipose Tissue Inflamma-tion Indeed a few individual miRNAs have been reportedas playing a crucial role in the inflammatory state of ATso far (Figure 1) Expression of miR-221 and miR-222 hasbeen positively correlated to TNF-120572 and negatively corre-lated to ApN expression in white AT of mice [69] Yet
the direct effects of both miRNAs on these pro- or anti-inflammatory adipokines are still unraveled miR-132 whichwas downregulated in human obese omental fat [66] hasbeen reported to activate NF-120581B and the transcription of IL-8 and MCP-1 in primary human preadipocytes and in invitro differentiated adipocytes [70] Zhuang et al demon-strated that miR-223 played a crucial role in modulatingmacrophage polarization in a pattern that protects micefrom diet-induced AT inflammation and systemic insulinresistance Consequently miR-223 suppressed the infiltrationof proinflammatory M1 ldquoclassically activatedrdquo macrophagesby targeting Pknox1 in vitro while miR-223 deficient micefed a high-fat diet exhibited increased AT inflammationcharacterized by enhanced proinflammatory activation ofmacrophages [71] The group of Brichard has identifiedseveral miRNAs which were regulated by adiponectin (apromising anti-inflammatory adipokine) in white AT in vivoThe miR883b-5p which was upregulated by adiponectin anddownregulated in human obesity repressed the LPS-bindingprotein (LBP) andTLR4 signaling acting therefore as amajormediator of the anti-inflammatory action of ApN MoreovermiR883b-silencing in the de novo AT formed from in vivodifferentiation of preadipocytes also induced LBP productionand tissue inflammation [72] Very recently Arner et aloverexpressed individual miRNAs (which have been defineddownregulated inAT in human obesity) in human adipocytesdifferentiated in vitro They found that a set of nine miRNAssignificantly reduced the secretion of chemokine (C-Cmotif)ligand 2 (CCL2) which is an initiator of AT inflammationby attracting the migration of inflammatory cells into thetissue Among these miRNAs only miR-126 was predictedby in silico analysis and confirmed by luciferase reporterassay in 3T3-L1 cells to bind directly to the 31015840-UTR of CCL2whereas miR-193b affected indirectly the CCL2 productionthrough downregulating the transcription factors (TFs) ofCCL2 (RELB STAT6 and ETS1) [67] Thus miRNAs maymediate AT inflammation by regulating either the activationof macrophages or the production of adipokines The mech-anism by which miRNAs regulate adipokines in AT seemsvia direct and indirectmechanismsmiRNAsmay act directlyon the target inflammatory adipokines or indirectly by firstregulating the intermediate machinery components like theTFs which in turn control the expression of adipokines
In addition to the innate immunity recent studies havedisclosed the importance of the adaptive immunity in ATinflammationThe subsets of the lymphocyte lineage includ-ing CD8+ and CD4+ T-cells Tregs Natural killer T-cells(NKT) and B-cells have been revealed to infiltrate into ATwhich may orchestrate AT inflammation Lymphocyte infil-tration into AT could even precede macrophage infiltration[73 74] Yet the mechanisms underlying the infiltration oflymphocytes into AT remain largely unknown (Figure 1)Whether miRNAs could play roles in modulating the infil-tration or activation of the lymphocytes in AT may arouseresearch interest and requires to be investigated in the future
miRNAsMediate Intercellular CommunicationMore recentlyadipocytes have been reported to secrete miRNAs in theform of adipocyte-derived microvesicles (ADMs) which
Journal of Immunology Research 5
LymphocytemiRNAs
Pknox1
M1
miRNAMicrovesicle
ApN
miR-884b-5p
LBP
LBP LPS
Adipocyte
miR-132
Macrophage
miR-126
CCL2
miR-193b
miR-223
NF-120581B
IL-8 and MCP-1
RELB STAT6 and ETS1
Figure 1 Scheme for the miRNA-mediated mechanisms in regulating white adipose tissue inflammation The miR-132 activates NF-120581B andthe transcription of downstream adipokines IL-8 and MCP-1 The miR-223 played a crucial role in modulating macrophage polarizationwhich suppresses the infiltration of proinflammatory M1 ldquoclassically activatedrdquo macrophages by targeting Pknox1 The miR883b-5p which isupregulated by ApN represses the expression and secretion of LBP a LPS facilitatorThemiR-126 reduces the production of CCL2 by targetingdirectly the 31015840-UTR of CCL2 while miR-193b inhibited indirectly the CCL2 production through downregulating the transcription factors ofCCL2 (RELB STAT6 and ETS1) The microvesicles released by adipocytes may either go into circulation or transfer into the neighboringadipocytes ormacrophages probably acting as inflammatory communicators between adipocytes macrophages and distant cellsThe arrowsindicate stimulation or activation blunted arrows indicate inhibition ApN adiponectin LBP LPS-binding protein LPS lipopolysaccharideCCL2 chemokine (C-C motif) ligand 2 adapted from [5]
may regulate the function of distant or neighboring cellsOne study found that microvesicles released from cultured3T3-L1 adipocytes harbored 143 miRNAs most of thembeing adipocyte-specific and reflecting the abundance oftheir expression levels in the donor cells Interestinglythe miRNAs-containing microvesicles were transported intocultured macrophages which were incubated with ADMscontaining-medium for 24 h they were also present in ratserum in vivo [75] Yet whether the miRNAs inside thevesicles were functional in their new location remains tobe determined Furthermore another group discovered thatmiRNAs-microvesicle complexes derived from the largerprimary rat adipocytes were transferred into and expressed inthe smaller adipocytes and were involved in the transcriptionof multiple genes for lipid synthesis and cell growth [76]Thus miRNAs could also be important mediators of cellularcrosstalk Since macrophages and hypertrophied adipocytesare the main sources of AT inflammation the miRNA-mediated interactions (adipocytes-macrophages and largeradipocytes-smaller adipocytes)may be novelmechanisms formiRNAs contributing to AT inflammation (Figure 1)
6 Therapeutic Potential of microRNAs inMetabolic Syndrome
The facts that miRNAs are largely dysregulated in obe-sity and that specific miRNAs regulate obesity-associatedinflammation suggests the potential of these molecules astargets for therapeutic intervention in obesity and obesity-related metabolic disorders
61 microRNAs as Diagnostic Tools Recent studies havedemonstrated that miRNAs can be secreted into the circula-tion Chen et al screened serummiRNAs of healthy Chinesesubjects and found over 100 and 91 serum miRNAs in maleand female subjects respectively Yet how these miRNAsmake their way into the circulation is still mysterious [77]New studies have revealed that circulatingmiRNAs can residein microvesicles (exosomes microparticles and apoptoticbodies) and in proteinlipoprotein complexes (high densitylipoprotein (HDL) and Argonaute 2) these formations makethe circulating miRNAs protected and resistant to RNase
6 Journal of Immunology Research
activity and degradation Therefore the levels of miRNAsin serum are remarkable stable reproducible and tissue-specific among individuals This raises the potential of miR-NAs as novel biomarkers formany disordersMultiple reportshave noted the potential use of blood miRNAs as biomarkersfor the obesity cardiovascular diseases atherosclerosis andT2DM Heneghan et al showed that circulating levels ofmiR-17-5p and miR-132 were significantly decreased in obeseindividuals compared to nonobese individuals and reflectedmiRNA expression in omental fat The miRNA expressionin blood and omental fat from obese patients correlatedsignificantly with BMI fasting blood glucose and glycatedhemoglobin [66] Cardiac specific miR-1 miR-208 and miR-499 were induced consistently in blood of patients withmyocardial infarction and strongly correlated to markers ofcardiac damage including troponin T and creatine kinase-MB (CK-MB) activity [78ndash81] CirculatingmiRNAs have alsobeen proposed as highly sensitive biomarkers for T2DM Inthe Bruneck cohort of 822 subjects decreased serum levelsof miR-126 in T2DM emerged as a significant predictorof T2DM and correlated negatively with increasing glucoseintolerance [82]
62 microRNAs as Therapeutic Tools ldquoSilencingrdquo of keymiRNA and ldquoreplacementrdquo of certain tissue-specific miRNAwhose expression is known to be decreased are potentialtherapeutic interventions
Techniques Several techniques have been set up to use thetherapeutic potential of miRNAs miRNAs can be deacti-vated and silenced by anti-miRNA oligonucleotides (AMOs)ldquomiRNA spongesrdquo and ldquomiRNA maskingrdquo [83] AMOsare synthetic antisense oligonucleotides that competitivelyinhibit the interaction between miRNAs and their mRNAtargets The most widely employed types of AMOs are 21015840-O-methyl AMO 21015840-O-methoxyethyl AMOs and LockedNucleic Acids (LNAs) [84] Given that miRNAs have beenobserved to function often in clusters in pathological pro-cesses the ldquomiRNA spongesrdquo have been developed to knock-down multiple miRNAs This technique requires a vectorencoding large quantities of transcripts (anti-miRNAs)whichdisplay numerous and tandem binding sites for a cluster ofmiRNAs of interest The ldquomiRNA maskingrdquo is an alternativemiRNA-knockdown strategy to the AMO approach withthe advantage of targeting miRNAs in a gene-specific man-ner Unlike AMO which binds to the endogenous miRNAdirectly a miRNA mask binds to the miRNArsquos binding sitelocated in the 31015840-UTR of its target mRNA thereby avoidingoff-target effects [83]
Conversely with regard to those miRNAs with decreasedexpression in disease the fundamental principle strategy isto restore their expression This can be achieved throughmiRNA mimicry or plasmidviral vector-encoded miRNAreplacement miRNA mimics are small chemically altereddouble-stranded RNA molecules that imitate endogenousmiRNAs [83] Plasmidviral vectors encoding miRNAs areencouraging strategies to replace miRNA in vivo with goodtransduction efficiency and minimal toxicity [85 86]
microRNAs and Therapeutics Since the obesity-inducedinflammation is considered to build the common soil forthe pathogenesis of MS some potential anti-inflammatorytherapies have been investigated Several drugs in currentclinical practice show the anti-inflammatory properties theperoxisome proliferator-activated receptor (PPAR) familyand nonacetylated salicylates Other strategies have beenexplored by targeting the cytokines and chemokines or theirreceptors (eg the anti-TNF or anti-CCR2 (chemokine (CndashCmotif) receptor 2) therapies) [2] Although there have beensome encouraging results it is likely that the benefits of theseapproaches are limited [87 88] Therefore the developmentof miRNA-based therapeutics has raised interest in this field
To date the greatest efforts have been made in exploringthe potential application of miRNAs therapeutics in cancerand liver disorders Gain or loss of function of individualmiRNAs have been reported in almost every solid andhematological cancer with therapeutic suppressing effectin tumor cell proliferation progression of tumors and themetastatic process [83] The liver specific miR-122 has beenproved to regulate cholesterol biosynthesis and hepatitis Cvirus replication Silencing of miR-122 by intraperitonealadministration of high affinity LNAanti-miR-122 has resultedin dose-dependent lowering of plasma cholesterol in miceand nonhuman primates (monkeys)This was achieved with-out significant adverse reactions or hepatic toxicity [89 90]
Likewise induction or reduction of miRNA expressionthat is known to be underupper-expressed and involved inAT inflammation using viral or liposomal delivery of tissue-specific miRNAs to affected AT could potentially result inrestoration of immune balance to the AT Yet the applicationof miRNAs therapeutics in AT in vivo is poorly investigatedbecause fat cells are insusceptible to selective transfer ofexogenous nucleotide due to the containing lipid dropletsof adipocytes Encouragingly the discovery that miRNAscould be secreted by adipocytes and accepted by neighboringadipocytes or macrophages in the form of adipocyte-derivedmicrovesicles [75 76] raises the hope that it could be possibleto import the targeting miRNAs into AT in vivo by themanner of microvesicles This manner could construct themicrovesicles containing specific miRNAs and inject theconstructed microvesicles into white AT
63 Potential Challenges in microRNA-Based TherapyAlthough significant advances have been made in miRNA-based therapy various challenges remain to be overcomebefore clinical use First one individual miRNA mayhave multiple potential targets which may coordinate orantagonize each otherrsquos functions In addition miRNA-target interactions depend not only on the sequence ofthe target site but also on the cellular context in which theinteractions occurThis complexity explains the difficulties inpredicting the spectrum of side effects and toxicity profileswhich may be associated with miRNA-based therapeuticsSecond replacement of miRNA may potentially lead tooversaturation of endogenous miRNAs pathway High levelsof exogenous miRNA can compete with the endogenousmiRNA biogenesis processing leading to cell toxicity
Journal of Immunology Research 7
[91 92] Another critical issue is the site-specific safeeffective and repeated delivery of miRNA in vivo Thesystemic intravenous delivery of viral or liposomal-mediatedmiRNA oligonucleotides have been well achieved in cancerand liver disease However targeting certain miRNAs inother specific tissues is poorly achieved due to the invasivenature of access for repeat delivery and lack of cell-specificup-take Restoration of tissue-specific miRNA expressioncould be a more rational approach for managing localabnormalities with more efficiency and precision and withless systemic side effects [83 92]
7 Conclusion
Evidence has provided that miRNAs are largely dysregulatedin white AT in human obesity specific miRNAs regulate thepolarization ofmacrophages and the expression and secretionof adipokines in white AT a subset ofmiRNAs could be pack-aged into adipocyte-derived microvesicles and delivered intoblood or neighboring cellsmediating intercellular communi-cation Several reports strongly support the roles of miRNAsas important mediators in AT inflammation Furthermorewith the recent discovery that miRNAs are secreted intocirculation in a stable manner and correlated closely withcrucial metabolic parameters the plasma miRNA profilescould therefore be used as attractive biomarkers for theMS Finally the establishment of miRNAs-based techniquesallows exploring further fascinating roles of miRNAs andtheir applications in the management of MS Despite hugechallenges lying in miRNA-targeting therapeutics especiallyin developing safe and effective delivery of targeting miRNAsin specific tissues in vivo the development of miRNA-basedtherapeutics for MS is warranted because of the severeproblem that metabolic disease poses to society
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This work was supported by grants from the National KeyClinical Department Construction Project
References
[1] Y-H Tseng A M Cypess and C R Kahn ldquoCellular bioen-ergetics as a target for obesity therapyrdquo Nature Reviews DrugDiscovery vol 9 no 6 pp 465ndash482 2010
[2] G S Hotamisligil ldquoInflammation and metabolic disordersrdquoNature vol 444 no 7121 pp 860ndash867 2006
[3] K L Spalding E Arner P O Westermark et al ldquoDynamics offat cell turnover in humansrdquoNature vol 453 no 7196 pp 783ndash787 2008
[4] P Jiao andH Xu ldquoAdipose inflammation cause or consequenceof obesity-related insulin resistancerdquo Diabetes Metabolic Syn-drome and Obesity vol 1 pp 25ndash31 2008
[5] EMaury and SM Brichard ldquoAdipokine dysregulation adiposetissue inflammation and metabolic syndromerdquo Molecular andCellular Endocrinology vol 314 no 1 pp 1ndash16 2010
[6] L-F Lu and A Liston ldquoMicroRNA in the immune systemmicroRNA as an immune systemrdquo Immunology vol 127 no 3pp 291ndash298 2009
[7] D Sayed and M Abdellatif ldquoMicrornas in development anddiseaserdquo Physiological Reviews vol 91 no 3 pp 827ndash887 2011
[8] H M Heneghan N Miller and M J Kerin ldquoRole of microR-NAs in obesity and the metabolic syndromerdquo Obesity Reviewsvol 11 no 5 pp 354ndash361 2010
[9] M Hulsmans D De Keyzer and P Holvoet ldquoMicroRNAsregulating oxidative stress and inflammation in relation toobesity and atherosclerosisrdquo The FASEB Journal vol 25 no 8pp 2515ndash2527 2011
[10] R Alexander H Lodish and L Sun ldquoMicroRNAs in adipoge-nesis and as therapeutic targets for obesityrdquo Expert Opinion onTherapeutic Targets vol 15 no 5 pp 623ndash636 2011
[11] C N Lumeng and A R Saltiel ldquoInflammatory links betweenobesity andmetabolic diseaserdquoThe Journal of Clinical Investiga-tion vol 121 no 6 pp 2111ndash2117 2011
[12] Y Chen J Zhu P Y Lum et al ldquoVariations in DNA elucidatemolecular networks that cause diseaserdquo Nature vol 452 no7186 pp 429ndash435 2008
[13] V Emilsson G Thorleifsson B Zhang et al ldquoGenetics of geneexpression and its effect on diseaserdquo Nature vol 452 no 7186pp 423ndash428 2008
[14] V Varma A Yao-Borengasser N Rasouli et al ldquoMuscle inflam-matory response and insulin resistance synergistic interactionbetween macrophages and fatty acids leads to impaired insulinactionrdquoThe American Journal of Physiology Endocrinology andMetabolism vol 296 no 6 pp E1300ndashE1310 2009
[15] WHuang AMetlakunta N Dedousis et al ldquoDepletion of liverkupffer cells prevents the development of diet-induced hepaticsteatosis and insulin resistancerdquoDiabetes vol 59 no 2 pp 347ndash357 2010
[16] J P Thaler C-X Yi E A Schur et al ldquoObesity is associatedwith hypothalamic injury in rodents and humansrdquoThe Journalof Clinical Investigation vol 122 no 1 pp 153ndash162 2012
[17] Y-M Kang Y Ma J-P Zheng et al ldquoBrain nuclear factor-kappa B activation contributes to neurohumoral excitation inangiotensin II-induced hypertensionrdquoCardiovascular Researchvol 82 no 3 pp 503ndash512 2009
[18] V C Calegari A S Torsoni E C Vanzela et al ldquoInflammationof the hypothalamus leads to defective pancreatic islet functionrdquoThe Journal of Biological Chemistry vol 286 no 15 pp 12870ndash12880 2011
[19] S Purkayastha H Zhang G Zhang Z Ahmed Y Wangand D Cai ldquoNeural dysregulation of peripheral insulin actionand blood pressure by brain endoplasmic reticulum stressrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 108 no 7 pp 2939ndash2944 2011
[20] M E Trujillo and P E Scherer ldquoAdipose tissue-derived factorsimpact on health and diseaserdquo Endocrine Reviews vol 27 no 7pp 762ndash778 2006
[21] E Maury K Ehala-Aleksejev Y Guiot R Detry A Vanden-hooft and S M Brichard ldquoAdipokines oversecreted by omentaladipose tissue in human obesityrdquo The American Journal ofPhysiology Endocrinology and Metabolism vol 293 no 3 ppE656ndashE665 2007
8 Journal of Immunology Research
[22] V Bourlier A Zakaroff-Girard A Miranville et alldquoRemodeling phenotype of human subcutaneous adiposetissue macrophagesrdquo Circulation vol 117 no 6 pp 806ndash8152008
[23] I M Faust P R Johnson J S Stern and J Hirsch ldquoDiet-induced adipocyte number increase in adult rats a new modelof obesityrdquo The American Journal of Physiology vol 235 no 3pp E279ndashE286 1978
[24] C Henegar J Tordjman V Achard et al ldquoAdipose tissuetranscriptomic signature highlights the pathological relevanceof extracellular matrix in human obesityrdquo Genome Biology vol9 article R14 2008
[25] B Kabon A Nagele D Reddy et al ldquoObesity decreasesperioperative tissue oxygenationrdquo Anesthesiology vol 100 no2 pp 274ndash280 2004
[26] K A Virtanen P Lonnroth R Parkkola et al ldquoGlucose uptakeand perfusion in subcutaneous and visceral adipose tissueduring insulin stimulation in nonobese and obese humansrdquoJournal of Clinical Endocrinology and Metabolism vol 87 no8 pp 3902ndash3910 2002
[27] N Hosogai A Fukuhara K Oshima et al ldquoAdipose tissuehypoxia in obesity and its impact on adipocytokine dysregula-tionrdquo Diabetes vol 56 no 4 pp 901ndash911 2007
[28] J Ye Z Gao J Yin andQHe ldquoHypoxia is a potential risk factorfor chronic inflammation and adiponectin reduction in adiposetissue of obob and dietary obese micerdquo The American Journalof Physiology Endocrinology andMetabolism vol 293 no 4 ppE1118ndashE1128 2007
[29] B Wang I S Wood and P Trayhurn ldquoDysregulation of theexpression and secretion of inflammation-related adipokines byhypoxia in human adipocytesrdquoPflugers Archiv European Journalof Physiology vol 455 no 3 pp 479ndash492 2007
[30] J Yin Z GaoQHeD Zhou ZGuo and J Ye ldquoRole of hypoxiain obesity-induced disorders of glucose and lipidmetabolism inadipose tissuerdquoTheAmerican Journal of Physiology Endocrinol-ogy and Metabolism vol 296 no 2 pp E333ndashE342 2009
[31] Z Yun H L Maecker R S Johnson and A J GiaccialdquoInhibition of PPAR1205742 gene expression by the HIF-1-regulatedgene DEC1Stra13 a mechanism for regulation of adipogenesisby hypoxiardquo Developmental Cell vol 2 no 3 pp 331ndash341 2002
[32] L Liu T P Cash R G Jones B Keith C B Thompson andM C Simon ldquoHypoxia-induced energy stress regulates mRNAtranslation and cell growthrdquo Molecular Cell vol 21 no 4 pp521ndash531 2006
[33] K Zhang and R J Kaufman ldquoFrom endoplasmic-reticulumstress to the inflammatory responserdquoNature vol 454 no 7203pp 455ndash462 2008
[34] B P Tu and J S Weissman ldquoOxidative protein folding ineukaryotes mechanisms and consequencesrdquo Journal of CellBiology vol 164 no 3 pp 341ndash346 2004
[35] C M Haynes E A Titus and A A Cooper ldquoDegradation ofmisfolded proteins prevents ER-derived oxidative stress and celldeathrdquoMolecular Cell vol 15 no 5 pp 767ndash776 2004
[36] A Schaeffler P Gross R Buettner et al ldquoFatty acid-inducedinduction of Toll-like receptor-4nuclear factor-120581B pathway inadipocytes links nutritional signalling with innate immunityrdquoImmunology vol 126 no 2 pp 233ndash245 2009
[37] A Kopp C BuechlerMNeumeier et al ldquoInnate immunity andadipocyte function ligand-specific activation of multiple toll-like receptors modulates cytokine adipokine and chemokinesecretion in adipocytesrdquo Obesity vol 17 no 4 pp 648ndash6562009
[38] O I Vitseva K Tanriverdi T T Tchkonia et al ldquoInducibletoll-like receptor and NF-120581B regulatory pathway expression inhuman adipose tissuerdquoObesity vol 16 no 5 pp 932ndash937 2008
[39] S J Creely P G McTernan C M Kusminski et alldquoLipopolysaccharide activates an innate immune systemresponse in human adipose tissue in obesity and type 2diabetesrdquo The American Journal of Physiology Endocrinologyand Metabolism vol 292 no 3 pp E740ndashE747 2007
[40] A Schaffler J Scholmerich and B Salzberger ldquoAdipose tissueas an immunological organ toll-like receptors C1qTNFs andCTRPsrdquoTrends in Immunology vol 28 no 9 pp 393ndash399 2007
[41] J E Davis N K Gabler J Walker-Daniels and M E SpurlockldquoTlr-4 deficiency selectively protects against obesity induced bydiets high in saturated fatrdquoObesity vol 16 no 6 pp 1248ndash12552008
[42] T Suganami T Mieda M Itoh Y Shimoda Y Kamei and YOgawa ldquoAttenuation of obesity-induced adipose tissue inflam-mation in C3HHeJ mice carrying a toll-like receptor 4 muta-tionrdquo Biochemical and Biophysical Research Communicationsvol 354 no 1 pp 45ndash49 2007
[43] X Prieur T Roszer and M Ricote ldquoLipotoxicity inmacrophages evidence from diseases associated with themetabolic syndromerdquo Biochimica et Biophysica Acta Molecularand Cell Biology of Lipids vol 1801 no 3 pp 327ndash337 2010
[44] P D Cani J AmarM A Iglesias et al ldquoMetabolic endotoxemiainitiates obesity and insulin resistancerdquo Diabetes vol 56 no 7pp 1761ndash1772 2007
[45] S V Monte J A Caruana H Ghanim et al ldquoReduction inendotoxemia oxidative and inflammatory stress and insulinresistance after Roux-en-Y gastric bypass surgery in patientswith morbid obesity and type 2 diabetes mellitusrdquo Surgery vol151 no 4 pp 587ndash593 2012
[46] S Ghoshal J Witta J Zhong W de Villiers and E EckhardtldquoChylomicrons promote intestinal absorption of lipopolysac-charidesrdquo Journal of Lipid Research vol 50 no 1 pp 90ndash972009
[47] P D Cani R Bibiloni C Knauf et al ldquoChanges in gut micro-biota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in micerdquo Diabetesvol 57 no 6 pp 1470ndash1481 2008
[48] G GMuccioli D Naslain F Backhed et al ldquoThe endocannabi-noid system links gut microbiota to adipogenesisrdquo MolecularSystems Biology vol 6 article 392 2010
[49] P D Cani S Possemiers T Van De Wiele et al ldquoChanges ingut microbiota control inflammation in obese mice througha mechanism involving GLP-2-driven improvement of gutpermeabilityrdquo Gut vol 58 no 8 pp 1091ndash1103 2009
[50] A Schaffler and J Scholmerich ldquoInnate immunity and adiposetissue biologyrdquoTrends in Immunology vol 31 no 6 pp 228ndash2352010
[51] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[52] D OrsquoCarroll I Mecklenbrauker P P Das et al ldquoA Slicer-independent role for Argonaute 2 in hematopoiesis and themicroRNA pathwayrdquoGenes and Development vol 21 no 16 pp1999ndash2004 2007
[53] C Xiao L Srinivasan D P Calado et al ldquoLymphoproliferativedisease and autoimmunity in mice with increased miR-17-92expression in lymphocytesrdquo Nature Immunology vol 9 no 4pp 405ndash414 2008
Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Journal of Immunology Research
of 20 interleukin receptor genes examined only 2 had highprobability miRNA target sites These findings suggest thatthe regulation of cytokine genes by miRNAs occurs oftenvia indirect mechanism Recent studies indicate indeed thatmiRNAs could indirectly regulate cytokine genes via AU-richelements (ARE) located in the mRNA 31015840-UTR by targetingARE-binding proteins ARE are the cis-acting structural RNAmotifs that are important determinants of cytokine messagestability ARE-mediated mRNA degradation is regulated bya number of trans-acting factors called the ARE-bindingproteins (which include tristetraprolin (TTP) AU rich bind-ing factor 1 (AUF1) and members of the Hu protein R(HUR) family) [61] These ARE-binding proteins are heavilypredicted targets of miRNAs [62] Repression of several AREcomponents by miRNAsmay alter the levels of inflammatorycytokines as well as of other immune genes [60 62]
52 Role of microRNAs in White Adipose Tissue Inflammation
Obesity-Induced Inflammation Is Associated with Dysregu-lated Expression of miRNAs in White Adipose Tissue Low-grade inflammation in AT is a key feature of obesity Anumber of miRNAs have been found dysregulated in whiteAT during obesity and closely associated with obesity-related metabolic disorders A recent study identified 21miRNAs which were differentially expressed in epididymalAT between lean mice fed a standard diet and mice renderedobese by a high-fat diet [63] Ortega et al performedmiRNA array on human subcutaneous AT 50 of the 799miRNAs tested (62) significantly differed between obese(119899 = 9) and lean (119899 = 6) subjects [64] Among these 50miRNAs 17 were highly correlated with BMI and metabolicparameters (fasting glucose andor triglycerides) [64] Thesedata are concordant with those obtained in overweight orobese subjects by Kloting et al They showed significantcorrelations between the expression of selected miRNAsand both AT morphology and key metabolic parametersincluding visceral fat area HbA1c fasting plasma glucoseand circulating leptin adiponectin and IL-6 [65] Anothertwo miRNAs (miR-17ndash5p and miR-132) were significantlydecreased in the omental fat and the circulation of obesesubjects [66] A very recent study defined a set of elevenadipocyte-specific miRNAs downregulated in obese subjectsas all of them were concomitantly altered in both the whiteAT and the isolated adipocytes [67] Moreover inflammationper se could alter white AT miRNA levels Xie et al showedthat TNF-120572 treatment of 3T3-L1 adipocytes mimicked thechanges of miRNA expression observed in AT of obese mice[68] With obesity and inflammation being so intrinsicallyassociated the dysregulated expression ofmiRNAs in inflam-matory adipocytes highly suggests the particular importanceof miRNAs in obesity-induced inflammation
Individual miRNAs Regulate White Adipose Tissue Inflamma-tion Indeed a few individual miRNAs have been reportedas playing a crucial role in the inflammatory state of ATso far (Figure 1) Expression of miR-221 and miR-222 hasbeen positively correlated to TNF-120572 and negatively corre-lated to ApN expression in white AT of mice [69] Yet
the direct effects of both miRNAs on these pro- or anti-inflammatory adipokines are still unraveled miR-132 whichwas downregulated in human obese omental fat [66] hasbeen reported to activate NF-120581B and the transcription of IL-8 and MCP-1 in primary human preadipocytes and in invitro differentiated adipocytes [70] Zhuang et al demon-strated that miR-223 played a crucial role in modulatingmacrophage polarization in a pattern that protects micefrom diet-induced AT inflammation and systemic insulinresistance Consequently miR-223 suppressed the infiltrationof proinflammatory M1 ldquoclassically activatedrdquo macrophagesby targeting Pknox1 in vitro while miR-223 deficient micefed a high-fat diet exhibited increased AT inflammationcharacterized by enhanced proinflammatory activation ofmacrophages [71] The group of Brichard has identifiedseveral miRNAs which were regulated by adiponectin (apromising anti-inflammatory adipokine) in white AT in vivoThe miR883b-5p which was upregulated by adiponectin anddownregulated in human obesity repressed the LPS-bindingprotein (LBP) andTLR4 signaling acting therefore as amajormediator of the anti-inflammatory action of ApN MoreovermiR883b-silencing in the de novo AT formed from in vivodifferentiation of preadipocytes also induced LBP productionand tissue inflammation [72] Very recently Arner et aloverexpressed individual miRNAs (which have been defineddownregulated inAT in human obesity) in human adipocytesdifferentiated in vitro They found that a set of nine miRNAssignificantly reduced the secretion of chemokine (C-Cmotif)ligand 2 (CCL2) which is an initiator of AT inflammationby attracting the migration of inflammatory cells into thetissue Among these miRNAs only miR-126 was predictedby in silico analysis and confirmed by luciferase reporterassay in 3T3-L1 cells to bind directly to the 31015840-UTR of CCL2whereas miR-193b affected indirectly the CCL2 productionthrough downregulating the transcription factors (TFs) ofCCL2 (RELB STAT6 and ETS1) [67] Thus miRNAs maymediate AT inflammation by regulating either the activationof macrophages or the production of adipokines The mech-anism by which miRNAs regulate adipokines in AT seemsvia direct and indirectmechanismsmiRNAsmay act directlyon the target inflammatory adipokines or indirectly by firstregulating the intermediate machinery components like theTFs which in turn control the expression of adipokines
In addition to the innate immunity recent studies havedisclosed the importance of the adaptive immunity in ATinflammationThe subsets of the lymphocyte lineage includ-ing CD8+ and CD4+ T-cells Tregs Natural killer T-cells(NKT) and B-cells have been revealed to infiltrate into ATwhich may orchestrate AT inflammation Lymphocyte infil-tration into AT could even precede macrophage infiltration[73 74] Yet the mechanisms underlying the infiltration oflymphocytes into AT remain largely unknown (Figure 1)Whether miRNAs could play roles in modulating the infil-tration or activation of the lymphocytes in AT may arouseresearch interest and requires to be investigated in the future
miRNAsMediate Intercellular CommunicationMore recentlyadipocytes have been reported to secrete miRNAs in theform of adipocyte-derived microvesicles (ADMs) which
Journal of Immunology Research 5
LymphocytemiRNAs
Pknox1
M1
miRNAMicrovesicle
ApN
miR-884b-5p
LBP
LBP LPS
Adipocyte
miR-132
Macrophage
miR-126
CCL2
miR-193b
miR-223
NF-120581B
IL-8 and MCP-1
RELB STAT6 and ETS1
Figure 1 Scheme for the miRNA-mediated mechanisms in regulating white adipose tissue inflammation The miR-132 activates NF-120581B andthe transcription of downstream adipokines IL-8 and MCP-1 The miR-223 played a crucial role in modulating macrophage polarizationwhich suppresses the infiltration of proinflammatory M1 ldquoclassically activatedrdquo macrophages by targeting Pknox1 The miR883b-5p which isupregulated by ApN represses the expression and secretion of LBP a LPS facilitatorThemiR-126 reduces the production of CCL2 by targetingdirectly the 31015840-UTR of CCL2 while miR-193b inhibited indirectly the CCL2 production through downregulating the transcription factors ofCCL2 (RELB STAT6 and ETS1) The microvesicles released by adipocytes may either go into circulation or transfer into the neighboringadipocytes ormacrophages probably acting as inflammatory communicators between adipocytes macrophages and distant cellsThe arrowsindicate stimulation or activation blunted arrows indicate inhibition ApN adiponectin LBP LPS-binding protein LPS lipopolysaccharideCCL2 chemokine (C-C motif) ligand 2 adapted from [5]
may regulate the function of distant or neighboring cellsOne study found that microvesicles released from cultured3T3-L1 adipocytes harbored 143 miRNAs most of thembeing adipocyte-specific and reflecting the abundance oftheir expression levels in the donor cells Interestinglythe miRNAs-containing microvesicles were transported intocultured macrophages which were incubated with ADMscontaining-medium for 24 h they were also present in ratserum in vivo [75] Yet whether the miRNAs inside thevesicles were functional in their new location remains tobe determined Furthermore another group discovered thatmiRNAs-microvesicle complexes derived from the largerprimary rat adipocytes were transferred into and expressed inthe smaller adipocytes and were involved in the transcriptionof multiple genes for lipid synthesis and cell growth [76]Thus miRNAs could also be important mediators of cellularcrosstalk Since macrophages and hypertrophied adipocytesare the main sources of AT inflammation the miRNA-mediated interactions (adipocytes-macrophages and largeradipocytes-smaller adipocytes)may be novelmechanisms formiRNAs contributing to AT inflammation (Figure 1)
6 Therapeutic Potential of microRNAs inMetabolic Syndrome
The facts that miRNAs are largely dysregulated in obe-sity and that specific miRNAs regulate obesity-associatedinflammation suggests the potential of these molecules astargets for therapeutic intervention in obesity and obesity-related metabolic disorders
61 microRNAs as Diagnostic Tools Recent studies havedemonstrated that miRNAs can be secreted into the circula-tion Chen et al screened serummiRNAs of healthy Chinesesubjects and found over 100 and 91 serum miRNAs in maleand female subjects respectively Yet how these miRNAsmake their way into the circulation is still mysterious [77]New studies have revealed that circulatingmiRNAs can residein microvesicles (exosomes microparticles and apoptoticbodies) and in proteinlipoprotein complexes (high densitylipoprotein (HDL) and Argonaute 2) these formations makethe circulating miRNAs protected and resistant to RNase
6 Journal of Immunology Research
activity and degradation Therefore the levels of miRNAsin serum are remarkable stable reproducible and tissue-specific among individuals This raises the potential of miR-NAs as novel biomarkers formany disordersMultiple reportshave noted the potential use of blood miRNAs as biomarkersfor the obesity cardiovascular diseases atherosclerosis andT2DM Heneghan et al showed that circulating levels ofmiR-17-5p and miR-132 were significantly decreased in obeseindividuals compared to nonobese individuals and reflectedmiRNA expression in omental fat The miRNA expressionin blood and omental fat from obese patients correlatedsignificantly with BMI fasting blood glucose and glycatedhemoglobin [66] Cardiac specific miR-1 miR-208 and miR-499 were induced consistently in blood of patients withmyocardial infarction and strongly correlated to markers ofcardiac damage including troponin T and creatine kinase-MB (CK-MB) activity [78ndash81] CirculatingmiRNAs have alsobeen proposed as highly sensitive biomarkers for T2DM Inthe Bruneck cohort of 822 subjects decreased serum levelsof miR-126 in T2DM emerged as a significant predictorof T2DM and correlated negatively with increasing glucoseintolerance [82]
62 microRNAs as Therapeutic Tools ldquoSilencingrdquo of keymiRNA and ldquoreplacementrdquo of certain tissue-specific miRNAwhose expression is known to be decreased are potentialtherapeutic interventions
Techniques Several techniques have been set up to use thetherapeutic potential of miRNAs miRNAs can be deacti-vated and silenced by anti-miRNA oligonucleotides (AMOs)ldquomiRNA spongesrdquo and ldquomiRNA maskingrdquo [83] AMOsare synthetic antisense oligonucleotides that competitivelyinhibit the interaction between miRNAs and their mRNAtargets The most widely employed types of AMOs are 21015840-O-methyl AMO 21015840-O-methoxyethyl AMOs and LockedNucleic Acids (LNAs) [84] Given that miRNAs have beenobserved to function often in clusters in pathological pro-cesses the ldquomiRNA spongesrdquo have been developed to knock-down multiple miRNAs This technique requires a vectorencoding large quantities of transcripts (anti-miRNAs)whichdisplay numerous and tandem binding sites for a cluster ofmiRNAs of interest The ldquomiRNA maskingrdquo is an alternativemiRNA-knockdown strategy to the AMO approach withthe advantage of targeting miRNAs in a gene-specific man-ner Unlike AMO which binds to the endogenous miRNAdirectly a miRNA mask binds to the miRNArsquos binding sitelocated in the 31015840-UTR of its target mRNA thereby avoidingoff-target effects [83]
Conversely with regard to those miRNAs with decreasedexpression in disease the fundamental principle strategy isto restore their expression This can be achieved throughmiRNA mimicry or plasmidviral vector-encoded miRNAreplacement miRNA mimics are small chemically altereddouble-stranded RNA molecules that imitate endogenousmiRNAs [83] Plasmidviral vectors encoding miRNAs areencouraging strategies to replace miRNA in vivo with goodtransduction efficiency and minimal toxicity [85 86]
microRNAs and Therapeutics Since the obesity-inducedinflammation is considered to build the common soil forthe pathogenesis of MS some potential anti-inflammatorytherapies have been investigated Several drugs in currentclinical practice show the anti-inflammatory properties theperoxisome proliferator-activated receptor (PPAR) familyand nonacetylated salicylates Other strategies have beenexplored by targeting the cytokines and chemokines or theirreceptors (eg the anti-TNF or anti-CCR2 (chemokine (CndashCmotif) receptor 2) therapies) [2] Although there have beensome encouraging results it is likely that the benefits of theseapproaches are limited [87 88] Therefore the developmentof miRNA-based therapeutics has raised interest in this field
To date the greatest efforts have been made in exploringthe potential application of miRNAs therapeutics in cancerand liver disorders Gain or loss of function of individualmiRNAs have been reported in almost every solid andhematological cancer with therapeutic suppressing effectin tumor cell proliferation progression of tumors and themetastatic process [83] The liver specific miR-122 has beenproved to regulate cholesterol biosynthesis and hepatitis Cvirus replication Silencing of miR-122 by intraperitonealadministration of high affinity LNAanti-miR-122 has resultedin dose-dependent lowering of plasma cholesterol in miceand nonhuman primates (monkeys)This was achieved with-out significant adverse reactions or hepatic toxicity [89 90]
Likewise induction or reduction of miRNA expressionthat is known to be underupper-expressed and involved inAT inflammation using viral or liposomal delivery of tissue-specific miRNAs to affected AT could potentially result inrestoration of immune balance to the AT Yet the applicationof miRNAs therapeutics in AT in vivo is poorly investigatedbecause fat cells are insusceptible to selective transfer ofexogenous nucleotide due to the containing lipid dropletsof adipocytes Encouragingly the discovery that miRNAscould be secreted by adipocytes and accepted by neighboringadipocytes or macrophages in the form of adipocyte-derivedmicrovesicles [75 76] raises the hope that it could be possibleto import the targeting miRNAs into AT in vivo by themanner of microvesicles This manner could construct themicrovesicles containing specific miRNAs and inject theconstructed microvesicles into white AT
63 Potential Challenges in microRNA-Based TherapyAlthough significant advances have been made in miRNA-based therapy various challenges remain to be overcomebefore clinical use First one individual miRNA mayhave multiple potential targets which may coordinate orantagonize each otherrsquos functions In addition miRNA-target interactions depend not only on the sequence ofthe target site but also on the cellular context in which theinteractions occurThis complexity explains the difficulties inpredicting the spectrum of side effects and toxicity profileswhich may be associated with miRNA-based therapeuticsSecond replacement of miRNA may potentially lead tooversaturation of endogenous miRNAs pathway High levelsof exogenous miRNA can compete with the endogenousmiRNA biogenesis processing leading to cell toxicity
Journal of Immunology Research 7
[91 92] Another critical issue is the site-specific safeeffective and repeated delivery of miRNA in vivo Thesystemic intravenous delivery of viral or liposomal-mediatedmiRNA oligonucleotides have been well achieved in cancerand liver disease However targeting certain miRNAs inother specific tissues is poorly achieved due to the invasivenature of access for repeat delivery and lack of cell-specificup-take Restoration of tissue-specific miRNA expressioncould be a more rational approach for managing localabnormalities with more efficiency and precision and withless systemic side effects [83 92]
7 Conclusion
Evidence has provided that miRNAs are largely dysregulatedin white AT in human obesity specific miRNAs regulate thepolarization ofmacrophages and the expression and secretionof adipokines in white AT a subset ofmiRNAs could be pack-aged into adipocyte-derived microvesicles and delivered intoblood or neighboring cellsmediating intercellular communi-cation Several reports strongly support the roles of miRNAsas important mediators in AT inflammation Furthermorewith the recent discovery that miRNAs are secreted intocirculation in a stable manner and correlated closely withcrucial metabolic parameters the plasma miRNA profilescould therefore be used as attractive biomarkers for theMS Finally the establishment of miRNAs-based techniquesallows exploring further fascinating roles of miRNAs andtheir applications in the management of MS Despite hugechallenges lying in miRNA-targeting therapeutics especiallyin developing safe and effective delivery of targeting miRNAsin specific tissues in vivo the development of miRNA-basedtherapeutics for MS is warranted because of the severeproblem that metabolic disease poses to society
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This work was supported by grants from the National KeyClinical Department Construction Project
References
[1] Y-H Tseng A M Cypess and C R Kahn ldquoCellular bioen-ergetics as a target for obesity therapyrdquo Nature Reviews DrugDiscovery vol 9 no 6 pp 465ndash482 2010
[2] G S Hotamisligil ldquoInflammation and metabolic disordersrdquoNature vol 444 no 7121 pp 860ndash867 2006
[3] K L Spalding E Arner P O Westermark et al ldquoDynamics offat cell turnover in humansrdquoNature vol 453 no 7196 pp 783ndash787 2008
[4] P Jiao andH Xu ldquoAdipose inflammation cause or consequenceof obesity-related insulin resistancerdquo Diabetes Metabolic Syn-drome and Obesity vol 1 pp 25ndash31 2008
[5] EMaury and SM Brichard ldquoAdipokine dysregulation adiposetissue inflammation and metabolic syndromerdquo Molecular andCellular Endocrinology vol 314 no 1 pp 1ndash16 2010
[6] L-F Lu and A Liston ldquoMicroRNA in the immune systemmicroRNA as an immune systemrdquo Immunology vol 127 no 3pp 291ndash298 2009
[7] D Sayed and M Abdellatif ldquoMicrornas in development anddiseaserdquo Physiological Reviews vol 91 no 3 pp 827ndash887 2011
[8] H M Heneghan N Miller and M J Kerin ldquoRole of microR-NAs in obesity and the metabolic syndromerdquo Obesity Reviewsvol 11 no 5 pp 354ndash361 2010
[9] M Hulsmans D De Keyzer and P Holvoet ldquoMicroRNAsregulating oxidative stress and inflammation in relation toobesity and atherosclerosisrdquo The FASEB Journal vol 25 no 8pp 2515ndash2527 2011
[10] R Alexander H Lodish and L Sun ldquoMicroRNAs in adipoge-nesis and as therapeutic targets for obesityrdquo Expert Opinion onTherapeutic Targets vol 15 no 5 pp 623ndash636 2011
[11] C N Lumeng and A R Saltiel ldquoInflammatory links betweenobesity andmetabolic diseaserdquoThe Journal of Clinical Investiga-tion vol 121 no 6 pp 2111ndash2117 2011
[12] Y Chen J Zhu P Y Lum et al ldquoVariations in DNA elucidatemolecular networks that cause diseaserdquo Nature vol 452 no7186 pp 429ndash435 2008
[13] V Emilsson G Thorleifsson B Zhang et al ldquoGenetics of geneexpression and its effect on diseaserdquo Nature vol 452 no 7186pp 423ndash428 2008
[14] V Varma A Yao-Borengasser N Rasouli et al ldquoMuscle inflam-matory response and insulin resistance synergistic interactionbetween macrophages and fatty acids leads to impaired insulinactionrdquoThe American Journal of Physiology Endocrinology andMetabolism vol 296 no 6 pp E1300ndashE1310 2009
[15] WHuang AMetlakunta N Dedousis et al ldquoDepletion of liverkupffer cells prevents the development of diet-induced hepaticsteatosis and insulin resistancerdquoDiabetes vol 59 no 2 pp 347ndash357 2010
[16] J P Thaler C-X Yi E A Schur et al ldquoObesity is associatedwith hypothalamic injury in rodents and humansrdquoThe Journalof Clinical Investigation vol 122 no 1 pp 153ndash162 2012
[17] Y-M Kang Y Ma J-P Zheng et al ldquoBrain nuclear factor-kappa B activation contributes to neurohumoral excitation inangiotensin II-induced hypertensionrdquoCardiovascular Researchvol 82 no 3 pp 503ndash512 2009
[18] V C Calegari A S Torsoni E C Vanzela et al ldquoInflammationof the hypothalamus leads to defective pancreatic islet functionrdquoThe Journal of Biological Chemistry vol 286 no 15 pp 12870ndash12880 2011
[19] S Purkayastha H Zhang G Zhang Z Ahmed Y Wangand D Cai ldquoNeural dysregulation of peripheral insulin actionand blood pressure by brain endoplasmic reticulum stressrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 108 no 7 pp 2939ndash2944 2011
[20] M E Trujillo and P E Scherer ldquoAdipose tissue-derived factorsimpact on health and diseaserdquo Endocrine Reviews vol 27 no 7pp 762ndash778 2006
[21] E Maury K Ehala-Aleksejev Y Guiot R Detry A Vanden-hooft and S M Brichard ldquoAdipokines oversecreted by omentaladipose tissue in human obesityrdquo The American Journal ofPhysiology Endocrinology and Metabolism vol 293 no 3 ppE656ndashE665 2007
8 Journal of Immunology Research
[22] V Bourlier A Zakaroff-Girard A Miranville et alldquoRemodeling phenotype of human subcutaneous adiposetissue macrophagesrdquo Circulation vol 117 no 6 pp 806ndash8152008
[23] I M Faust P R Johnson J S Stern and J Hirsch ldquoDiet-induced adipocyte number increase in adult rats a new modelof obesityrdquo The American Journal of Physiology vol 235 no 3pp E279ndashE286 1978
[24] C Henegar J Tordjman V Achard et al ldquoAdipose tissuetranscriptomic signature highlights the pathological relevanceof extracellular matrix in human obesityrdquo Genome Biology vol9 article R14 2008
[25] B Kabon A Nagele D Reddy et al ldquoObesity decreasesperioperative tissue oxygenationrdquo Anesthesiology vol 100 no2 pp 274ndash280 2004
[26] K A Virtanen P Lonnroth R Parkkola et al ldquoGlucose uptakeand perfusion in subcutaneous and visceral adipose tissueduring insulin stimulation in nonobese and obese humansrdquoJournal of Clinical Endocrinology and Metabolism vol 87 no8 pp 3902ndash3910 2002
[27] N Hosogai A Fukuhara K Oshima et al ldquoAdipose tissuehypoxia in obesity and its impact on adipocytokine dysregula-tionrdquo Diabetes vol 56 no 4 pp 901ndash911 2007
[28] J Ye Z Gao J Yin andQHe ldquoHypoxia is a potential risk factorfor chronic inflammation and adiponectin reduction in adiposetissue of obob and dietary obese micerdquo The American Journalof Physiology Endocrinology andMetabolism vol 293 no 4 ppE1118ndashE1128 2007
[29] B Wang I S Wood and P Trayhurn ldquoDysregulation of theexpression and secretion of inflammation-related adipokines byhypoxia in human adipocytesrdquoPflugers Archiv European Journalof Physiology vol 455 no 3 pp 479ndash492 2007
[30] J Yin Z GaoQHeD Zhou ZGuo and J Ye ldquoRole of hypoxiain obesity-induced disorders of glucose and lipidmetabolism inadipose tissuerdquoTheAmerican Journal of Physiology Endocrinol-ogy and Metabolism vol 296 no 2 pp E333ndashE342 2009
[31] Z Yun H L Maecker R S Johnson and A J GiaccialdquoInhibition of PPAR1205742 gene expression by the HIF-1-regulatedgene DEC1Stra13 a mechanism for regulation of adipogenesisby hypoxiardquo Developmental Cell vol 2 no 3 pp 331ndash341 2002
[32] L Liu T P Cash R G Jones B Keith C B Thompson andM C Simon ldquoHypoxia-induced energy stress regulates mRNAtranslation and cell growthrdquo Molecular Cell vol 21 no 4 pp521ndash531 2006
[33] K Zhang and R J Kaufman ldquoFrom endoplasmic-reticulumstress to the inflammatory responserdquoNature vol 454 no 7203pp 455ndash462 2008
[34] B P Tu and J S Weissman ldquoOxidative protein folding ineukaryotes mechanisms and consequencesrdquo Journal of CellBiology vol 164 no 3 pp 341ndash346 2004
[35] C M Haynes E A Titus and A A Cooper ldquoDegradation ofmisfolded proteins prevents ER-derived oxidative stress and celldeathrdquoMolecular Cell vol 15 no 5 pp 767ndash776 2004
[36] A Schaeffler P Gross R Buettner et al ldquoFatty acid-inducedinduction of Toll-like receptor-4nuclear factor-120581B pathway inadipocytes links nutritional signalling with innate immunityrdquoImmunology vol 126 no 2 pp 233ndash245 2009
[37] A Kopp C BuechlerMNeumeier et al ldquoInnate immunity andadipocyte function ligand-specific activation of multiple toll-like receptors modulates cytokine adipokine and chemokinesecretion in adipocytesrdquo Obesity vol 17 no 4 pp 648ndash6562009
[38] O I Vitseva K Tanriverdi T T Tchkonia et al ldquoInducibletoll-like receptor and NF-120581B regulatory pathway expression inhuman adipose tissuerdquoObesity vol 16 no 5 pp 932ndash937 2008
[39] S J Creely P G McTernan C M Kusminski et alldquoLipopolysaccharide activates an innate immune systemresponse in human adipose tissue in obesity and type 2diabetesrdquo The American Journal of Physiology Endocrinologyand Metabolism vol 292 no 3 pp E740ndashE747 2007
[40] A Schaffler J Scholmerich and B Salzberger ldquoAdipose tissueas an immunological organ toll-like receptors C1qTNFs andCTRPsrdquoTrends in Immunology vol 28 no 9 pp 393ndash399 2007
[41] J E Davis N K Gabler J Walker-Daniels and M E SpurlockldquoTlr-4 deficiency selectively protects against obesity induced bydiets high in saturated fatrdquoObesity vol 16 no 6 pp 1248ndash12552008
[42] T Suganami T Mieda M Itoh Y Shimoda Y Kamei and YOgawa ldquoAttenuation of obesity-induced adipose tissue inflam-mation in C3HHeJ mice carrying a toll-like receptor 4 muta-tionrdquo Biochemical and Biophysical Research Communicationsvol 354 no 1 pp 45ndash49 2007
[43] X Prieur T Roszer and M Ricote ldquoLipotoxicity inmacrophages evidence from diseases associated with themetabolic syndromerdquo Biochimica et Biophysica Acta Molecularand Cell Biology of Lipids vol 1801 no 3 pp 327ndash337 2010
[44] P D Cani J AmarM A Iglesias et al ldquoMetabolic endotoxemiainitiates obesity and insulin resistancerdquo Diabetes vol 56 no 7pp 1761ndash1772 2007
[45] S V Monte J A Caruana H Ghanim et al ldquoReduction inendotoxemia oxidative and inflammatory stress and insulinresistance after Roux-en-Y gastric bypass surgery in patientswith morbid obesity and type 2 diabetes mellitusrdquo Surgery vol151 no 4 pp 587ndash593 2012
[46] S Ghoshal J Witta J Zhong W de Villiers and E EckhardtldquoChylomicrons promote intestinal absorption of lipopolysac-charidesrdquo Journal of Lipid Research vol 50 no 1 pp 90ndash972009
[47] P D Cani R Bibiloni C Knauf et al ldquoChanges in gut micro-biota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in micerdquo Diabetesvol 57 no 6 pp 1470ndash1481 2008
[48] G GMuccioli D Naslain F Backhed et al ldquoThe endocannabi-noid system links gut microbiota to adipogenesisrdquo MolecularSystems Biology vol 6 article 392 2010
[49] P D Cani S Possemiers T Van De Wiele et al ldquoChanges ingut microbiota control inflammation in obese mice througha mechanism involving GLP-2-driven improvement of gutpermeabilityrdquo Gut vol 58 no 8 pp 1091ndash1103 2009
[50] A Schaffler and J Scholmerich ldquoInnate immunity and adiposetissue biologyrdquoTrends in Immunology vol 31 no 6 pp 228ndash2352010
[51] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[52] D OrsquoCarroll I Mecklenbrauker P P Das et al ldquoA Slicer-independent role for Argonaute 2 in hematopoiesis and themicroRNA pathwayrdquoGenes and Development vol 21 no 16 pp1999ndash2004 2007
[53] C Xiao L Srinivasan D P Calado et al ldquoLymphoproliferativedisease and autoimmunity in mice with increased miR-17-92expression in lymphocytesrdquo Nature Immunology vol 9 no 4pp 405ndash414 2008
Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
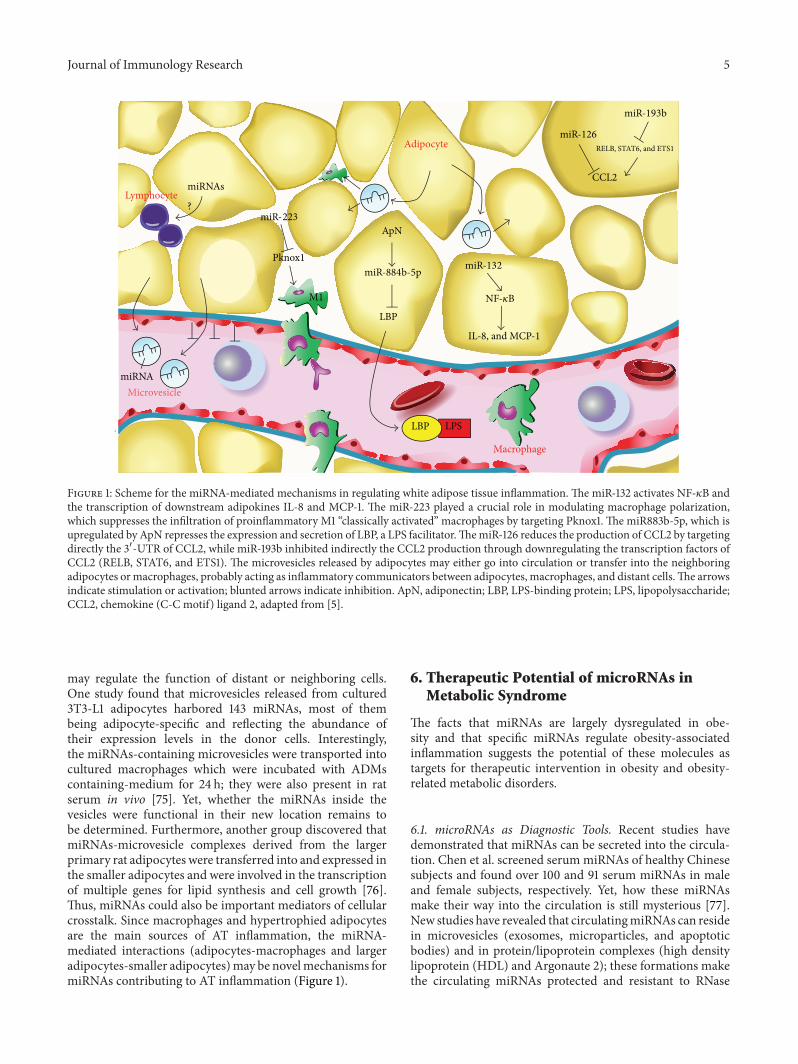
Journal of Immunology Research 5
LymphocytemiRNAs
Pknox1
M1
miRNAMicrovesicle
ApN
miR-884b-5p
LBP
LBP LPS
Adipocyte
miR-132
Macrophage
miR-126
CCL2
miR-193b
miR-223
NF-120581B
IL-8 and MCP-1
RELB STAT6 and ETS1
Figure 1 Scheme for the miRNA-mediated mechanisms in regulating white adipose tissue inflammation The miR-132 activates NF-120581B andthe transcription of downstream adipokines IL-8 and MCP-1 The miR-223 played a crucial role in modulating macrophage polarizationwhich suppresses the infiltration of proinflammatory M1 ldquoclassically activatedrdquo macrophages by targeting Pknox1 The miR883b-5p which isupregulated by ApN represses the expression and secretion of LBP a LPS facilitatorThemiR-126 reduces the production of CCL2 by targetingdirectly the 31015840-UTR of CCL2 while miR-193b inhibited indirectly the CCL2 production through downregulating the transcription factors ofCCL2 (RELB STAT6 and ETS1) The microvesicles released by adipocytes may either go into circulation or transfer into the neighboringadipocytes ormacrophages probably acting as inflammatory communicators between adipocytes macrophages and distant cellsThe arrowsindicate stimulation or activation blunted arrows indicate inhibition ApN adiponectin LBP LPS-binding protein LPS lipopolysaccharideCCL2 chemokine (C-C motif) ligand 2 adapted from [5]
may regulate the function of distant or neighboring cellsOne study found that microvesicles released from cultured3T3-L1 adipocytes harbored 143 miRNAs most of thembeing adipocyte-specific and reflecting the abundance oftheir expression levels in the donor cells Interestinglythe miRNAs-containing microvesicles were transported intocultured macrophages which were incubated with ADMscontaining-medium for 24 h they were also present in ratserum in vivo [75] Yet whether the miRNAs inside thevesicles were functional in their new location remains tobe determined Furthermore another group discovered thatmiRNAs-microvesicle complexes derived from the largerprimary rat adipocytes were transferred into and expressed inthe smaller adipocytes and were involved in the transcriptionof multiple genes for lipid synthesis and cell growth [76]Thus miRNAs could also be important mediators of cellularcrosstalk Since macrophages and hypertrophied adipocytesare the main sources of AT inflammation the miRNA-mediated interactions (adipocytes-macrophages and largeradipocytes-smaller adipocytes)may be novelmechanisms formiRNAs contributing to AT inflammation (Figure 1)
6 Therapeutic Potential of microRNAs inMetabolic Syndrome
The facts that miRNAs are largely dysregulated in obe-sity and that specific miRNAs regulate obesity-associatedinflammation suggests the potential of these molecules astargets for therapeutic intervention in obesity and obesity-related metabolic disorders
61 microRNAs as Diagnostic Tools Recent studies havedemonstrated that miRNAs can be secreted into the circula-tion Chen et al screened serummiRNAs of healthy Chinesesubjects and found over 100 and 91 serum miRNAs in maleand female subjects respectively Yet how these miRNAsmake their way into the circulation is still mysterious [77]New studies have revealed that circulatingmiRNAs can residein microvesicles (exosomes microparticles and apoptoticbodies) and in proteinlipoprotein complexes (high densitylipoprotein (HDL) and Argonaute 2) these formations makethe circulating miRNAs protected and resistant to RNase
6 Journal of Immunology Research
activity and degradation Therefore the levels of miRNAsin serum are remarkable stable reproducible and tissue-specific among individuals This raises the potential of miR-NAs as novel biomarkers formany disordersMultiple reportshave noted the potential use of blood miRNAs as biomarkersfor the obesity cardiovascular diseases atherosclerosis andT2DM Heneghan et al showed that circulating levels ofmiR-17-5p and miR-132 were significantly decreased in obeseindividuals compared to nonobese individuals and reflectedmiRNA expression in omental fat The miRNA expressionin blood and omental fat from obese patients correlatedsignificantly with BMI fasting blood glucose and glycatedhemoglobin [66] Cardiac specific miR-1 miR-208 and miR-499 were induced consistently in blood of patients withmyocardial infarction and strongly correlated to markers ofcardiac damage including troponin T and creatine kinase-MB (CK-MB) activity [78ndash81] CirculatingmiRNAs have alsobeen proposed as highly sensitive biomarkers for T2DM Inthe Bruneck cohort of 822 subjects decreased serum levelsof miR-126 in T2DM emerged as a significant predictorof T2DM and correlated negatively with increasing glucoseintolerance [82]
62 microRNAs as Therapeutic Tools ldquoSilencingrdquo of keymiRNA and ldquoreplacementrdquo of certain tissue-specific miRNAwhose expression is known to be decreased are potentialtherapeutic interventions
Techniques Several techniques have been set up to use thetherapeutic potential of miRNAs miRNAs can be deacti-vated and silenced by anti-miRNA oligonucleotides (AMOs)ldquomiRNA spongesrdquo and ldquomiRNA maskingrdquo [83] AMOsare synthetic antisense oligonucleotides that competitivelyinhibit the interaction between miRNAs and their mRNAtargets The most widely employed types of AMOs are 21015840-O-methyl AMO 21015840-O-methoxyethyl AMOs and LockedNucleic Acids (LNAs) [84] Given that miRNAs have beenobserved to function often in clusters in pathological pro-cesses the ldquomiRNA spongesrdquo have been developed to knock-down multiple miRNAs This technique requires a vectorencoding large quantities of transcripts (anti-miRNAs)whichdisplay numerous and tandem binding sites for a cluster ofmiRNAs of interest The ldquomiRNA maskingrdquo is an alternativemiRNA-knockdown strategy to the AMO approach withthe advantage of targeting miRNAs in a gene-specific man-ner Unlike AMO which binds to the endogenous miRNAdirectly a miRNA mask binds to the miRNArsquos binding sitelocated in the 31015840-UTR of its target mRNA thereby avoidingoff-target effects [83]
Conversely with regard to those miRNAs with decreasedexpression in disease the fundamental principle strategy isto restore their expression This can be achieved throughmiRNA mimicry or plasmidviral vector-encoded miRNAreplacement miRNA mimics are small chemically altereddouble-stranded RNA molecules that imitate endogenousmiRNAs [83] Plasmidviral vectors encoding miRNAs areencouraging strategies to replace miRNA in vivo with goodtransduction efficiency and minimal toxicity [85 86]
microRNAs and Therapeutics Since the obesity-inducedinflammation is considered to build the common soil forthe pathogenesis of MS some potential anti-inflammatorytherapies have been investigated Several drugs in currentclinical practice show the anti-inflammatory properties theperoxisome proliferator-activated receptor (PPAR) familyand nonacetylated salicylates Other strategies have beenexplored by targeting the cytokines and chemokines or theirreceptors (eg the anti-TNF or anti-CCR2 (chemokine (CndashCmotif) receptor 2) therapies) [2] Although there have beensome encouraging results it is likely that the benefits of theseapproaches are limited [87 88] Therefore the developmentof miRNA-based therapeutics has raised interest in this field
To date the greatest efforts have been made in exploringthe potential application of miRNAs therapeutics in cancerand liver disorders Gain or loss of function of individualmiRNAs have been reported in almost every solid andhematological cancer with therapeutic suppressing effectin tumor cell proliferation progression of tumors and themetastatic process [83] The liver specific miR-122 has beenproved to regulate cholesterol biosynthesis and hepatitis Cvirus replication Silencing of miR-122 by intraperitonealadministration of high affinity LNAanti-miR-122 has resultedin dose-dependent lowering of plasma cholesterol in miceand nonhuman primates (monkeys)This was achieved with-out significant adverse reactions or hepatic toxicity [89 90]
Likewise induction or reduction of miRNA expressionthat is known to be underupper-expressed and involved inAT inflammation using viral or liposomal delivery of tissue-specific miRNAs to affected AT could potentially result inrestoration of immune balance to the AT Yet the applicationof miRNAs therapeutics in AT in vivo is poorly investigatedbecause fat cells are insusceptible to selective transfer ofexogenous nucleotide due to the containing lipid dropletsof adipocytes Encouragingly the discovery that miRNAscould be secreted by adipocytes and accepted by neighboringadipocytes or macrophages in the form of adipocyte-derivedmicrovesicles [75 76] raises the hope that it could be possibleto import the targeting miRNAs into AT in vivo by themanner of microvesicles This manner could construct themicrovesicles containing specific miRNAs and inject theconstructed microvesicles into white AT
63 Potential Challenges in microRNA-Based TherapyAlthough significant advances have been made in miRNA-based therapy various challenges remain to be overcomebefore clinical use First one individual miRNA mayhave multiple potential targets which may coordinate orantagonize each otherrsquos functions In addition miRNA-target interactions depend not only on the sequence ofthe target site but also on the cellular context in which theinteractions occurThis complexity explains the difficulties inpredicting the spectrum of side effects and toxicity profileswhich may be associated with miRNA-based therapeuticsSecond replacement of miRNA may potentially lead tooversaturation of endogenous miRNAs pathway High levelsof exogenous miRNA can compete with the endogenousmiRNA biogenesis processing leading to cell toxicity
Journal of Immunology Research 7
[91 92] Another critical issue is the site-specific safeeffective and repeated delivery of miRNA in vivo Thesystemic intravenous delivery of viral or liposomal-mediatedmiRNA oligonucleotides have been well achieved in cancerand liver disease However targeting certain miRNAs inother specific tissues is poorly achieved due to the invasivenature of access for repeat delivery and lack of cell-specificup-take Restoration of tissue-specific miRNA expressioncould be a more rational approach for managing localabnormalities with more efficiency and precision and withless systemic side effects [83 92]
7 Conclusion
Evidence has provided that miRNAs are largely dysregulatedin white AT in human obesity specific miRNAs regulate thepolarization ofmacrophages and the expression and secretionof adipokines in white AT a subset ofmiRNAs could be pack-aged into adipocyte-derived microvesicles and delivered intoblood or neighboring cellsmediating intercellular communi-cation Several reports strongly support the roles of miRNAsas important mediators in AT inflammation Furthermorewith the recent discovery that miRNAs are secreted intocirculation in a stable manner and correlated closely withcrucial metabolic parameters the plasma miRNA profilescould therefore be used as attractive biomarkers for theMS Finally the establishment of miRNAs-based techniquesallows exploring further fascinating roles of miRNAs andtheir applications in the management of MS Despite hugechallenges lying in miRNA-targeting therapeutics especiallyin developing safe and effective delivery of targeting miRNAsin specific tissues in vivo the development of miRNA-basedtherapeutics for MS is warranted because of the severeproblem that metabolic disease poses to society
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This work was supported by grants from the National KeyClinical Department Construction Project
References
[1] Y-H Tseng A M Cypess and C R Kahn ldquoCellular bioen-ergetics as a target for obesity therapyrdquo Nature Reviews DrugDiscovery vol 9 no 6 pp 465ndash482 2010
[2] G S Hotamisligil ldquoInflammation and metabolic disordersrdquoNature vol 444 no 7121 pp 860ndash867 2006
[3] K L Spalding E Arner P O Westermark et al ldquoDynamics offat cell turnover in humansrdquoNature vol 453 no 7196 pp 783ndash787 2008
[4] P Jiao andH Xu ldquoAdipose inflammation cause or consequenceof obesity-related insulin resistancerdquo Diabetes Metabolic Syn-drome and Obesity vol 1 pp 25ndash31 2008
[5] EMaury and SM Brichard ldquoAdipokine dysregulation adiposetissue inflammation and metabolic syndromerdquo Molecular andCellular Endocrinology vol 314 no 1 pp 1ndash16 2010
[6] L-F Lu and A Liston ldquoMicroRNA in the immune systemmicroRNA as an immune systemrdquo Immunology vol 127 no 3pp 291ndash298 2009
[7] D Sayed and M Abdellatif ldquoMicrornas in development anddiseaserdquo Physiological Reviews vol 91 no 3 pp 827ndash887 2011
[8] H M Heneghan N Miller and M J Kerin ldquoRole of microR-NAs in obesity and the metabolic syndromerdquo Obesity Reviewsvol 11 no 5 pp 354ndash361 2010
[9] M Hulsmans D De Keyzer and P Holvoet ldquoMicroRNAsregulating oxidative stress and inflammation in relation toobesity and atherosclerosisrdquo The FASEB Journal vol 25 no 8pp 2515ndash2527 2011
[10] R Alexander H Lodish and L Sun ldquoMicroRNAs in adipoge-nesis and as therapeutic targets for obesityrdquo Expert Opinion onTherapeutic Targets vol 15 no 5 pp 623ndash636 2011
[11] C N Lumeng and A R Saltiel ldquoInflammatory links betweenobesity andmetabolic diseaserdquoThe Journal of Clinical Investiga-tion vol 121 no 6 pp 2111ndash2117 2011
[12] Y Chen J Zhu P Y Lum et al ldquoVariations in DNA elucidatemolecular networks that cause diseaserdquo Nature vol 452 no7186 pp 429ndash435 2008
[13] V Emilsson G Thorleifsson B Zhang et al ldquoGenetics of geneexpression and its effect on diseaserdquo Nature vol 452 no 7186pp 423ndash428 2008
[14] V Varma A Yao-Borengasser N Rasouli et al ldquoMuscle inflam-matory response and insulin resistance synergistic interactionbetween macrophages and fatty acids leads to impaired insulinactionrdquoThe American Journal of Physiology Endocrinology andMetabolism vol 296 no 6 pp E1300ndashE1310 2009
[15] WHuang AMetlakunta N Dedousis et al ldquoDepletion of liverkupffer cells prevents the development of diet-induced hepaticsteatosis and insulin resistancerdquoDiabetes vol 59 no 2 pp 347ndash357 2010
[16] J P Thaler C-X Yi E A Schur et al ldquoObesity is associatedwith hypothalamic injury in rodents and humansrdquoThe Journalof Clinical Investigation vol 122 no 1 pp 153ndash162 2012
[17] Y-M Kang Y Ma J-P Zheng et al ldquoBrain nuclear factor-kappa B activation contributes to neurohumoral excitation inangiotensin II-induced hypertensionrdquoCardiovascular Researchvol 82 no 3 pp 503ndash512 2009
[18] V C Calegari A S Torsoni E C Vanzela et al ldquoInflammationof the hypothalamus leads to defective pancreatic islet functionrdquoThe Journal of Biological Chemistry vol 286 no 15 pp 12870ndash12880 2011
[19] S Purkayastha H Zhang G Zhang Z Ahmed Y Wangand D Cai ldquoNeural dysregulation of peripheral insulin actionand blood pressure by brain endoplasmic reticulum stressrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 108 no 7 pp 2939ndash2944 2011
[20] M E Trujillo and P E Scherer ldquoAdipose tissue-derived factorsimpact on health and diseaserdquo Endocrine Reviews vol 27 no 7pp 762ndash778 2006
[21] E Maury K Ehala-Aleksejev Y Guiot R Detry A Vanden-hooft and S M Brichard ldquoAdipokines oversecreted by omentaladipose tissue in human obesityrdquo The American Journal ofPhysiology Endocrinology and Metabolism vol 293 no 3 ppE656ndashE665 2007
8 Journal of Immunology Research
[22] V Bourlier A Zakaroff-Girard A Miranville et alldquoRemodeling phenotype of human subcutaneous adiposetissue macrophagesrdquo Circulation vol 117 no 6 pp 806ndash8152008
[23] I M Faust P R Johnson J S Stern and J Hirsch ldquoDiet-induced adipocyte number increase in adult rats a new modelof obesityrdquo The American Journal of Physiology vol 235 no 3pp E279ndashE286 1978
[24] C Henegar J Tordjman V Achard et al ldquoAdipose tissuetranscriptomic signature highlights the pathological relevanceof extracellular matrix in human obesityrdquo Genome Biology vol9 article R14 2008
[25] B Kabon A Nagele D Reddy et al ldquoObesity decreasesperioperative tissue oxygenationrdquo Anesthesiology vol 100 no2 pp 274ndash280 2004
[26] K A Virtanen P Lonnroth R Parkkola et al ldquoGlucose uptakeand perfusion in subcutaneous and visceral adipose tissueduring insulin stimulation in nonobese and obese humansrdquoJournal of Clinical Endocrinology and Metabolism vol 87 no8 pp 3902ndash3910 2002
[27] N Hosogai A Fukuhara K Oshima et al ldquoAdipose tissuehypoxia in obesity and its impact on adipocytokine dysregula-tionrdquo Diabetes vol 56 no 4 pp 901ndash911 2007
[28] J Ye Z Gao J Yin andQHe ldquoHypoxia is a potential risk factorfor chronic inflammation and adiponectin reduction in adiposetissue of obob and dietary obese micerdquo The American Journalof Physiology Endocrinology andMetabolism vol 293 no 4 ppE1118ndashE1128 2007
[29] B Wang I S Wood and P Trayhurn ldquoDysregulation of theexpression and secretion of inflammation-related adipokines byhypoxia in human adipocytesrdquoPflugers Archiv European Journalof Physiology vol 455 no 3 pp 479ndash492 2007
[30] J Yin Z GaoQHeD Zhou ZGuo and J Ye ldquoRole of hypoxiain obesity-induced disorders of glucose and lipidmetabolism inadipose tissuerdquoTheAmerican Journal of Physiology Endocrinol-ogy and Metabolism vol 296 no 2 pp E333ndashE342 2009
[31] Z Yun H L Maecker R S Johnson and A J GiaccialdquoInhibition of PPAR1205742 gene expression by the HIF-1-regulatedgene DEC1Stra13 a mechanism for regulation of adipogenesisby hypoxiardquo Developmental Cell vol 2 no 3 pp 331ndash341 2002
[32] L Liu T P Cash R G Jones B Keith C B Thompson andM C Simon ldquoHypoxia-induced energy stress regulates mRNAtranslation and cell growthrdquo Molecular Cell vol 21 no 4 pp521ndash531 2006
[33] K Zhang and R J Kaufman ldquoFrom endoplasmic-reticulumstress to the inflammatory responserdquoNature vol 454 no 7203pp 455ndash462 2008
[34] B P Tu and J S Weissman ldquoOxidative protein folding ineukaryotes mechanisms and consequencesrdquo Journal of CellBiology vol 164 no 3 pp 341ndash346 2004
[35] C M Haynes E A Titus and A A Cooper ldquoDegradation ofmisfolded proteins prevents ER-derived oxidative stress and celldeathrdquoMolecular Cell vol 15 no 5 pp 767ndash776 2004
[36] A Schaeffler P Gross R Buettner et al ldquoFatty acid-inducedinduction of Toll-like receptor-4nuclear factor-120581B pathway inadipocytes links nutritional signalling with innate immunityrdquoImmunology vol 126 no 2 pp 233ndash245 2009
[37] A Kopp C BuechlerMNeumeier et al ldquoInnate immunity andadipocyte function ligand-specific activation of multiple toll-like receptors modulates cytokine adipokine and chemokinesecretion in adipocytesrdquo Obesity vol 17 no 4 pp 648ndash6562009
[38] O I Vitseva K Tanriverdi T T Tchkonia et al ldquoInducibletoll-like receptor and NF-120581B regulatory pathway expression inhuman adipose tissuerdquoObesity vol 16 no 5 pp 932ndash937 2008
[39] S J Creely P G McTernan C M Kusminski et alldquoLipopolysaccharide activates an innate immune systemresponse in human adipose tissue in obesity and type 2diabetesrdquo The American Journal of Physiology Endocrinologyand Metabolism vol 292 no 3 pp E740ndashE747 2007
[40] A Schaffler J Scholmerich and B Salzberger ldquoAdipose tissueas an immunological organ toll-like receptors C1qTNFs andCTRPsrdquoTrends in Immunology vol 28 no 9 pp 393ndash399 2007
[41] J E Davis N K Gabler J Walker-Daniels and M E SpurlockldquoTlr-4 deficiency selectively protects against obesity induced bydiets high in saturated fatrdquoObesity vol 16 no 6 pp 1248ndash12552008
[42] T Suganami T Mieda M Itoh Y Shimoda Y Kamei and YOgawa ldquoAttenuation of obesity-induced adipose tissue inflam-mation in C3HHeJ mice carrying a toll-like receptor 4 muta-tionrdquo Biochemical and Biophysical Research Communicationsvol 354 no 1 pp 45ndash49 2007
[43] X Prieur T Roszer and M Ricote ldquoLipotoxicity inmacrophages evidence from diseases associated with themetabolic syndromerdquo Biochimica et Biophysica Acta Molecularand Cell Biology of Lipids vol 1801 no 3 pp 327ndash337 2010
[44] P D Cani J AmarM A Iglesias et al ldquoMetabolic endotoxemiainitiates obesity and insulin resistancerdquo Diabetes vol 56 no 7pp 1761ndash1772 2007
[45] S V Monte J A Caruana H Ghanim et al ldquoReduction inendotoxemia oxidative and inflammatory stress and insulinresistance after Roux-en-Y gastric bypass surgery in patientswith morbid obesity and type 2 diabetes mellitusrdquo Surgery vol151 no 4 pp 587ndash593 2012
[46] S Ghoshal J Witta J Zhong W de Villiers and E EckhardtldquoChylomicrons promote intestinal absorption of lipopolysac-charidesrdquo Journal of Lipid Research vol 50 no 1 pp 90ndash972009
[47] P D Cani R Bibiloni C Knauf et al ldquoChanges in gut micro-biota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in micerdquo Diabetesvol 57 no 6 pp 1470ndash1481 2008
[48] G GMuccioli D Naslain F Backhed et al ldquoThe endocannabi-noid system links gut microbiota to adipogenesisrdquo MolecularSystems Biology vol 6 article 392 2010
[49] P D Cani S Possemiers T Van De Wiele et al ldquoChanges ingut microbiota control inflammation in obese mice througha mechanism involving GLP-2-driven improvement of gutpermeabilityrdquo Gut vol 58 no 8 pp 1091ndash1103 2009
[50] A Schaffler and J Scholmerich ldquoInnate immunity and adiposetissue biologyrdquoTrends in Immunology vol 31 no 6 pp 228ndash2352010
[51] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[52] D OrsquoCarroll I Mecklenbrauker P P Das et al ldquoA Slicer-independent role for Argonaute 2 in hematopoiesis and themicroRNA pathwayrdquoGenes and Development vol 21 no 16 pp1999ndash2004 2007
[53] C Xiao L Srinivasan D P Calado et al ldquoLymphoproliferativedisease and autoimmunity in mice with increased miR-17-92expression in lymphocytesrdquo Nature Immunology vol 9 no 4pp 405ndash414 2008
Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Journal of Immunology Research
activity and degradation Therefore the levels of miRNAsin serum are remarkable stable reproducible and tissue-specific among individuals This raises the potential of miR-NAs as novel biomarkers formany disordersMultiple reportshave noted the potential use of blood miRNAs as biomarkersfor the obesity cardiovascular diseases atherosclerosis andT2DM Heneghan et al showed that circulating levels ofmiR-17-5p and miR-132 were significantly decreased in obeseindividuals compared to nonobese individuals and reflectedmiRNA expression in omental fat The miRNA expressionin blood and omental fat from obese patients correlatedsignificantly with BMI fasting blood glucose and glycatedhemoglobin [66] Cardiac specific miR-1 miR-208 and miR-499 were induced consistently in blood of patients withmyocardial infarction and strongly correlated to markers ofcardiac damage including troponin T and creatine kinase-MB (CK-MB) activity [78ndash81] CirculatingmiRNAs have alsobeen proposed as highly sensitive biomarkers for T2DM Inthe Bruneck cohort of 822 subjects decreased serum levelsof miR-126 in T2DM emerged as a significant predictorof T2DM and correlated negatively with increasing glucoseintolerance [82]
62 microRNAs as Therapeutic Tools ldquoSilencingrdquo of keymiRNA and ldquoreplacementrdquo of certain tissue-specific miRNAwhose expression is known to be decreased are potentialtherapeutic interventions
Techniques Several techniques have been set up to use thetherapeutic potential of miRNAs miRNAs can be deacti-vated and silenced by anti-miRNA oligonucleotides (AMOs)ldquomiRNA spongesrdquo and ldquomiRNA maskingrdquo [83] AMOsare synthetic antisense oligonucleotides that competitivelyinhibit the interaction between miRNAs and their mRNAtargets The most widely employed types of AMOs are 21015840-O-methyl AMO 21015840-O-methoxyethyl AMOs and LockedNucleic Acids (LNAs) [84] Given that miRNAs have beenobserved to function often in clusters in pathological pro-cesses the ldquomiRNA spongesrdquo have been developed to knock-down multiple miRNAs This technique requires a vectorencoding large quantities of transcripts (anti-miRNAs)whichdisplay numerous and tandem binding sites for a cluster ofmiRNAs of interest The ldquomiRNA maskingrdquo is an alternativemiRNA-knockdown strategy to the AMO approach withthe advantage of targeting miRNAs in a gene-specific man-ner Unlike AMO which binds to the endogenous miRNAdirectly a miRNA mask binds to the miRNArsquos binding sitelocated in the 31015840-UTR of its target mRNA thereby avoidingoff-target effects [83]
Conversely with regard to those miRNAs with decreasedexpression in disease the fundamental principle strategy isto restore their expression This can be achieved throughmiRNA mimicry or plasmidviral vector-encoded miRNAreplacement miRNA mimics are small chemically altereddouble-stranded RNA molecules that imitate endogenousmiRNAs [83] Plasmidviral vectors encoding miRNAs areencouraging strategies to replace miRNA in vivo with goodtransduction efficiency and minimal toxicity [85 86]
microRNAs and Therapeutics Since the obesity-inducedinflammation is considered to build the common soil forthe pathogenesis of MS some potential anti-inflammatorytherapies have been investigated Several drugs in currentclinical practice show the anti-inflammatory properties theperoxisome proliferator-activated receptor (PPAR) familyand nonacetylated salicylates Other strategies have beenexplored by targeting the cytokines and chemokines or theirreceptors (eg the anti-TNF or anti-CCR2 (chemokine (CndashCmotif) receptor 2) therapies) [2] Although there have beensome encouraging results it is likely that the benefits of theseapproaches are limited [87 88] Therefore the developmentof miRNA-based therapeutics has raised interest in this field
To date the greatest efforts have been made in exploringthe potential application of miRNAs therapeutics in cancerand liver disorders Gain or loss of function of individualmiRNAs have been reported in almost every solid andhematological cancer with therapeutic suppressing effectin tumor cell proliferation progression of tumors and themetastatic process [83] The liver specific miR-122 has beenproved to regulate cholesterol biosynthesis and hepatitis Cvirus replication Silencing of miR-122 by intraperitonealadministration of high affinity LNAanti-miR-122 has resultedin dose-dependent lowering of plasma cholesterol in miceand nonhuman primates (monkeys)This was achieved with-out significant adverse reactions or hepatic toxicity [89 90]
Likewise induction or reduction of miRNA expressionthat is known to be underupper-expressed and involved inAT inflammation using viral or liposomal delivery of tissue-specific miRNAs to affected AT could potentially result inrestoration of immune balance to the AT Yet the applicationof miRNAs therapeutics in AT in vivo is poorly investigatedbecause fat cells are insusceptible to selective transfer ofexogenous nucleotide due to the containing lipid dropletsof adipocytes Encouragingly the discovery that miRNAscould be secreted by adipocytes and accepted by neighboringadipocytes or macrophages in the form of adipocyte-derivedmicrovesicles [75 76] raises the hope that it could be possibleto import the targeting miRNAs into AT in vivo by themanner of microvesicles This manner could construct themicrovesicles containing specific miRNAs and inject theconstructed microvesicles into white AT
63 Potential Challenges in microRNA-Based TherapyAlthough significant advances have been made in miRNA-based therapy various challenges remain to be overcomebefore clinical use First one individual miRNA mayhave multiple potential targets which may coordinate orantagonize each otherrsquos functions In addition miRNA-target interactions depend not only on the sequence ofthe target site but also on the cellular context in which theinteractions occurThis complexity explains the difficulties inpredicting the spectrum of side effects and toxicity profileswhich may be associated with miRNA-based therapeuticsSecond replacement of miRNA may potentially lead tooversaturation of endogenous miRNAs pathway High levelsof exogenous miRNA can compete with the endogenousmiRNA biogenesis processing leading to cell toxicity
Journal of Immunology Research 7
[91 92] Another critical issue is the site-specific safeeffective and repeated delivery of miRNA in vivo Thesystemic intravenous delivery of viral or liposomal-mediatedmiRNA oligonucleotides have been well achieved in cancerand liver disease However targeting certain miRNAs inother specific tissues is poorly achieved due to the invasivenature of access for repeat delivery and lack of cell-specificup-take Restoration of tissue-specific miRNA expressioncould be a more rational approach for managing localabnormalities with more efficiency and precision and withless systemic side effects [83 92]
7 Conclusion
Evidence has provided that miRNAs are largely dysregulatedin white AT in human obesity specific miRNAs regulate thepolarization ofmacrophages and the expression and secretionof adipokines in white AT a subset ofmiRNAs could be pack-aged into adipocyte-derived microvesicles and delivered intoblood or neighboring cellsmediating intercellular communi-cation Several reports strongly support the roles of miRNAsas important mediators in AT inflammation Furthermorewith the recent discovery that miRNAs are secreted intocirculation in a stable manner and correlated closely withcrucial metabolic parameters the plasma miRNA profilescould therefore be used as attractive biomarkers for theMS Finally the establishment of miRNAs-based techniquesallows exploring further fascinating roles of miRNAs andtheir applications in the management of MS Despite hugechallenges lying in miRNA-targeting therapeutics especiallyin developing safe and effective delivery of targeting miRNAsin specific tissues in vivo the development of miRNA-basedtherapeutics for MS is warranted because of the severeproblem that metabolic disease poses to society
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This work was supported by grants from the National KeyClinical Department Construction Project
References
[1] Y-H Tseng A M Cypess and C R Kahn ldquoCellular bioen-ergetics as a target for obesity therapyrdquo Nature Reviews DrugDiscovery vol 9 no 6 pp 465ndash482 2010
[2] G S Hotamisligil ldquoInflammation and metabolic disordersrdquoNature vol 444 no 7121 pp 860ndash867 2006
[3] K L Spalding E Arner P O Westermark et al ldquoDynamics offat cell turnover in humansrdquoNature vol 453 no 7196 pp 783ndash787 2008
[4] P Jiao andH Xu ldquoAdipose inflammation cause or consequenceof obesity-related insulin resistancerdquo Diabetes Metabolic Syn-drome and Obesity vol 1 pp 25ndash31 2008
[5] EMaury and SM Brichard ldquoAdipokine dysregulation adiposetissue inflammation and metabolic syndromerdquo Molecular andCellular Endocrinology vol 314 no 1 pp 1ndash16 2010
[6] L-F Lu and A Liston ldquoMicroRNA in the immune systemmicroRNA as an immune systemrdquo Immunology vol 127 no 3pp 291ndash298 2009
[7] D Sayed and M Abdellatif ldquoMicrornas in development anddiseaserdquo Physiological Reviews vol 91 no 3 pp 827ndash887 2011
[8] H M Heneghan N Miller and M J Kerin ldquoRole of microR-NAs in obesity and the metabolic syndromerdquo Obesity Reviewsvol 11 no 5 pp 354ndash361 2010
[9] M Hulsmans D De Keyzer and P Holvoet ldquoMicroRNAsregulating oxidative stress and inflammation in relation toobesity and atherosclerosisrdquo The FASEB Journal vol 25 no 8pp 2515ndash2527 2011
[10] R Alexander H Lodish and L Sun ldquoMicroRNAs in adipoge-nesis and as therapeutic targets for obesityrdquo Expert Opinion onTherapeutic Targets vol 15 no 5 pp 623ndash636 2011
[11] C N Lumeng and A R Saltiel ldquoInflammatory links betweenobesity andmetabolic diseaserdquoThe Journal of Clinical Investiga-tion vol 121 no 6 pp 2111ndash2117 2011
[12] Y Chen J Zhu P Y Lum et al ldquoVariations in DNA elucidatemolecular networks that cause diseaserdquo Nature vol 452 no7186 pp 429ndash435 2008
[13] V Emilsson G Thorleifsson B Zhang et al ldquoGenetics of geneexpression and its effect on diseaserdquo Nature vol 452 no 7186pp 423ndash428 2008
[14] V Varma A Yao-Borengasser N Rasouli et al ldquoMuscle inflam-matory response and insulin resistance synergistic interactionbetween macrophages and fatty acids leads to impaired insulinactionrdquoThe American Journal of Physiology Endocrinology andMetabolism vol 296 no 6 pp E1300ndashE1310 2009
[15] WHuang AMetlakunta N Dedousis et al ldquoDepletion of liverkupffer cells prevents the development of diet-induced hepaticsteatosis and insulin resistancerdquoDiabetes vol 59 no 2 pp 347ndash357 2010
[16] J P Thaler C-X Yi E A Schur et al ldquoObesity is associatedwith hypothalamic injury in rodents and humansrdquoThe Journalof Clinical Investigation vol 122 no 1 pp 153ndash162 2012
[17] Y-M Kang Y Ma J-P Zheng et al ldquoBrain nuclear factor-kappa B activation contributes to neurohumoral excitation inangiotensin II-induced hypertensionrdquoCardiovascular Researchvol 82 no 3 pp 503ndash512 2009
[18] V C Calegari A S Torsoni E C Vanzela et al ldquoInflammationof the hypothalamus leads to defective pancreatic islet functionrdquoThe Journal of Biological Chemistry vol 286 no 15 pp 12870ndash12880 2011
[19] S Purkayastha H Zhang G Zhang Z Ahmed Y Wangand D Cai ldquoNeural dysregulation of peripheral insulin actionand blood pressure by brain endoplasmic reticulum stressrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 108 no 7 pp 2939ndash2944 2011
[20] M E Trujillo and P E Scherer ldquoAdipose tissue-derived factorsimpact on health and diseaserdquo Endocrine Reviews vol 27 no 7pp 762ndash778 2006
[21] E Maury K Ehala-Aleksejev Y Guiot R Detry A Vanden-hooft and S M Brichard ldquoAdipokines oversecreted by omentaladipose tissue in human obesityrdquo The American Journal ofPhysiology Endocrinology and Metabolism vol 293 no 3 ppE656ndashE665 2007
8 Journal of Immunology Research
[22] V Bourlier A Zakaroff-Girard A Miranville et alldquoRemodeling phenotype of human subcutaneous adiposetissue macrophagesrdquo Circulation vol 117 no 6 pp 806ndash8152008
[23] I M Faust P R Johnson J S Stern and J Hirsch ldquoDiet-induced adipocyte number increase in adult rats a new modelof obesityrdquo The American Journal of Physiology vol 235 no 3pp E279ndashE286 1978
[24] C Henegar J Tordjman V Achard et al ldquoAdipose tissuetranscriptomic signature highlights the pathological relevanceof extracellular matrix in human obesityrdquo Genome Biology vol9 article R14 2008
[25] B Kabon A Nagele D Reddy et al ldquoObesity decreasesperioperative tissue oxygenationrdquo Anesthesiology vol 100 no2 pp 274ndash280 2004
[26] K A Virtanen P Lonnroth R Parkkola et al ldquoGlucose uptakeand perfusion in subcutaneous and visceral adipose tissueduring insulin stimulation in nonobese and obese humansrdquoJournal of Clinical Endocrinology and Metabolism vol 87 no8 pp 3902ndash3910 2002
[27] N Hosogai A Fukuhara K Oshima et al ldquoAdipose tissuehypoxia in obesity and its impact on adipocytokine dysregula-tionrdquo Diabetes vol 56 no 4 pp 901ndash911 2007
[28] J Ye Z Gao J Yin andQHe ldquoHypoxia is a potential risk factorfor chronic inflammation and adiponectin reduction in adiposetissue of obob and dietary obese micerdquo The American Journalof Physiology Endocrinology andMetabolism vol 293 no 4 ppE1118ndashE1128 2007
[29] B Wang I S Wood and P Trayhurn ldquoDysregulation of theexpression and secretion of inflammation-related adipokines byhypoxia in human adipocytesrdquoPflugers Archiv European Journalof Physiology vol 455 no 3 pp 479ndash492 2007
[30] J Yin Z GaoQHeD Zhou ZGuo and J Ye ldquoRole of hypoxiain obesity-induced disorders of glucose and lipidmetabolism inadipose tissuerdquoTheAmerican Journal of Physiology Endocrinol-ogy and Metabolism vol 296 no 2 pp E333ndashE342 2009
[31] Z Yun H L Maecker R S Johnson and A J GiaccialdquoInhibition of PPAR1205742 gene expression by the HIF-1-regulatedgene DEC1Stra13 a mechanism for regulation of adipogenesisby hypoxiardquo Developmental Cell vol 2 no 3 pp 331ndash341 2002
[32] L Liu T P Cash R G Jones B Keith C B Thompson andM C Simon ldquoHypoxia-induced energy stress regulates mRNAtranslation and cell growthrdquo Molecular Cell vol 21 no 4 pp521ndash531 2006
[33] K Zhang and R J Kaufman ldquoFrom endoplasmic-reticulumstress to the inflammatory responserdquoNature vol 454 no 7203pp 455ndash462 2008
[34] B P Tu and J S Weissman ldquoOxidative protein folding ineukaryotes mechanisms and consequencesrdquo Journal of CellBiology vol 164 no 3 pp 341ndash346 2004
[35] C M Haynes E A Titus and A A Cooper ldquoDegradation ofmisfolded proteins prevents ER-derived oxidative stress and celldeathrdquoMolecular Cell vol 15 no 5 pp 767ndash776 2004
[36] A Schaeffler P Gross R Buettner et al ldquoFatty acid-inducedinduction of Toll-like receptor-4nuclear factor-120581B pathway inadipocytes links nutritional signalling with innate immunityrdquoImmunology vol 126 no 2 pp 233ndash245 2009
[37] A Kopp C BuechlerMNeumeier et al ldquoInnate immunity andadipocyte function ligand-specific activation of multiple toll-like receptors modulates cytokine adipokine and chemokinesecretion in adipocytesrdquo Obesity vol 17 no 4 pp 648ndash6562009
[38] O I Vitseva K Tanriverdi T T Tchkonia et al ldquoInducibletoll-like receptor and NF-120581B regulatory pathway expression inhuman adipose tissuerdquoObesity vol 16 no 5 pp 932ndash937 2008
[39] S J Creely P G McTernan C M Kusminski et alldquoLipopolysaccharide activates an innate immune systemresponse in human adipose tissue in obesity and type 2diabetesrdquo The American Journal of Physiology Endocrinologyand Metabolism vol 292 no 3 pp E740ndashE747 2007
[40] A Schaffler J Scholmerich and B Salzberger ldquoAdipose tissueas an immunological organ toll-like receptors C1qTNFs andCTRPsrdquoTrends in Immunology vol 28 no 9 pp 393ndash399 2007
[41] J E Davis N K Gabler J Walker-Daniels and M E SpurlockldquoTlr-4 deficiency selectively protects against obesity induced bydiets high in saturated fatrdquoObesity vol 16 no 6 pp 1248ndash12552008
[42] T Suganami T Mieda M Itoh Y Shimoda Y Kamei and YOgawa ldquoAttenuation of obesity-induced adipose tissue inflam-mation in C3HHeJ mice carrying a toll-like receptor 4 muta-tionrdquo Biochemical and Biophysical Research Communicationsvol 354 no 1 pp 45ndash49 2007
[43] X Prieur T Roszer and M Ricote ldquoLipotoxicity inmacrophages evidence from diseases associated with themetabolic syndromerdquo Biochimica et Biophysica Acta Molecularand Cell Biology of Lipids vol 1801 no 3 pp 327ndash337 2010
[44] P D Cani J AmarM A Iglesias et al ldquoMetabolic endotoxemiainitiates obesity and insulin resistancerdquo Diabetes vol 56 no 7pp 1761ndash1772 2007
[45] S V Monte J A Caruana H Ghanim et al ldquoReduction inendotoxemia oxidative and inflammatory stress and insulinresistance after Roux-en-Y gastric bypass surgery in patientswith morbid obesity and type 2 diabetes mellitusrdquo Surgery vol151 no 4 pp 587ndash593 2012
[46] S Ghoshal J Witta J Zhong W de Villiers and E EckhardtldquoChylomicrons promote intestinal absorption of lipopolysac-charidesrdquo Journal of Lipid Research vol 50 no 1 pp 90ndash972009
[47] P D Cani R Bibiloni C Knauf et al ldquoChanges in gut micro-biota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in micerdquo Diabetesvol 57 no 6 pp 1470ndash1481 2008
[48] G GMuccioli D Naslain F Backhed et al ldquoThe endocannabi-noid system links gut microbiota to adipogenesisrdquo MolecularSystems Biology vol 6 article 392 2010
[49] P D Cani S Possemiers T Van De Wiele et al ldquoChanges ingut microbiota control inflammation in obese mice througha mechanism involving GLP-2-driven improvement of gutpermeabilityrdquo Gut vol 58 no 8 pp 1091ndash1103 2009
[50] A Schaffler and J Scholmerich ldquoInnate immunity and adiposetissue biologyrdquoTrends in Immunology vol 31 no 6 pp 228ndash2352010
[51] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[52] D OrsquoCarroll I Mecklenbrauker P P Das et al ldquoA Slicer-independent role for Argonaute 2 in hematopoiesis and themicroRNA pathwayrdquoGenes and Development vol 21 no 16 pp1999ndash2004 2007
[53] C Xiao L Srinivasan D P Calado et al ldquoLymphoproliferativedisease and autoimmunity in mice with increased miR-17-92expression in lymphocytesrdquo Nature Immunology vol 9 no 4pp 405ndash414 2008
Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 7
[91 92] Another critical issue is the site-specific safeeffective and repeated delivery of miRNA in vivo Thesystemic intravenous delivery of viral or liposomal-mediatedmiRNA oligonucleotides have been well achieved in cancerand liver disease However targeting certain miRNAs inother specific tissues is poorly achieved due to the invasivenature of access for repeat delivery and lack of cell-specificup-take Restoration of tissue-specific miRNA expressioncould be a more rational approach for managing localabnormalities with more efficiency and precision and withless systemic side effects [83 92]
7 Conclusion
Evidence has provided that miRNAs are largely dysregulatedin white AT in human obesity specific miRNAs regulate thepolarization ofmacrophages and the expression and secretionof adipokines in white AT a subset ofmiRNAs could be pack-aged into adipocyte-derived microvesicles and delivered intoblood or neighboring cellsmediating intercellular communi-cation Several reports strongly support the roles of miRNAsas important mediators in AT inflammation Furthermorewith the recent discovery that miRNAs are secreted intocirculation in a stable manner and correlated closely withcrucial metabolic parameters the plasma miRNA profilescould therefore be used as attractive biomarkers for theMS Finally the establishment of miRNAs-based techniquesallows exploring further fascinating roles of miRNAs andtheir applications in the management of MS Despite hugechallenges lying in miRNA-targeting therapeutics especiallyin developing safe and effective delivery of targeting miRNAsin specific tissues in vivo the development of miRNA-basedtherapeutics for MS is warranted because of the severeproblem that metabolic disease poses to society
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This work was supported by grants from the National KeyClinical Department Construction Project
References
[1] Y-H Tseng A M Cypess and C R Kahn ldquoCellular bioen-ergetics as a target for obesity therapyrdquo Nature Reviews DrugDiscovery vol 9 no 6 pp 465ndash482 2010
[2] G S Hotamisligil ldquoInflammation and metabolic disordersrdquoNature vol 444 no 7121 pp 860ndash867 2006
[3] K L Spalding E Arner P O Westermark et al ldquoDynamics offat cell turnover in humansrdquoNature vol 453 no 7196 pp 783ndash787 2008
[4] P Jiao andH Xu ldquoAdipose inflammation cause or consequenceof obesity-related insulin resistancerdquo Diabetes Metabolic Syn-drome and Obesity vol 1 pp 25ndash31 2008
[5] EMaury and SM Brichard ldquoAdipokine dysregulation adiposetissue inflammation and metabolic syndromerdquo Molecular andCellular Endocrinology vol 314 no 1 pp 1ndash16 2010
[6] L-F Lu and A Liston ldquoMicroRNA in the immune systemmicroRNA as an immune systemrdquo Immunology vol 127 no 3pp 291ndash298 2009
[7] D Sayed and M Abdellatif ldquoMicrornas in development anddiseaserdquo Physiological Reviews vol 91 no 3 pp 827ndash887 2011
[8] H M Heneghan N Miller and M J Kerin ldquoRole of microR-NAs in obesity and the metabolic syndromerdquo Obesity Reviewsvol 11 no 5 pp 354ndash361 2010
[9] M Hulsmans D De Keyzer and P Holvoet ldquoMicroRNAsregulating oxidative stress and inflammation in relation toobesity and atherosclerosisrdquo The FASEB Journal vol 25 no 8pp 2515ndash2527 2011
[10] R Alexander H Lodish and L Sun ldquoMicroRNAs in adipoge-nesis and as therapeutic targets for obesityrdquo Expert Opinion onTherapeutic Targets vol 15 no 5 pp 623ndash636 2011
[11] C N Lumeng and A R Saltiel ldquoInflammatory links betweenobesity andmetabolic diseaserdquoThe Journal of Clinical Investiga-tion vol 121 no 6 pp 2111ndash2117 2011
[12] Y Chen J Zhu P Y Lum et al ldquoVariations in DNA elucidatemolecular networks that cause diseaserdquo Nature vol 452 no7186 pp 429ndash435 2008
[13] V Emilsson G Thorleifsson B Zhang et al ldquoGenetics of geneexpression and its effect on diseaserdquo Nature vol 452 no 7186pp 423ndash428 2008
[14] V Varma A Yao-Borengasser N Rasouli et al ldquoMuscle inflam-matory response and insulin resistance synergistic interactionbetween macrophages and fatty acids leads to impaired insulinactionrdquoThe American Journal of Physiology Endocrinology andMetabolism vol 296 no 6 pp E1300ndashE1310 2009
[15] WHuang AMetlakunta N Dedousis et al ldquoDepletion of liverkupffer cells prevents the development of diet-induced hepaticsteatosis and insulin resistancerdquoDiabetes vol 59 no 2 pp 347ndash357 2010
[16] J P Thaler C-X Yi E A Schur et al ldquoObesity is associatedwith hypothalamic injury in rodents and humansrdquoThe Journalof Clinical Investigation vol 122 no 1 pp 153ndash162 2012
[17] Y-M Kang Y Ma J-P Zheng et al ldquoBrain nuclear factor-kappa B activation contributes to neurohumoral excitation inangiotensin II-induced hypertensionrdquoCardiovascular Researchvol 82 no 3 pp 503ndash512 2009
[18] V C Calegari A S Torsoni E C Vanzela et al ldquoInflammationof the hypothalamus leads to defective pancreatic islet functionrdquoThe Journal of Biological Chemistry vol 286 no 15 pp 12870ndash12880 2011
[19] S Purkayastha H Zhang G Zhang Z Ahmed Y Wangand D Cai ldquoNeural dysregulation of peripheral insulin actionand blood pressure by brain endoplasmic reticulum stressrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 108 no 7 pp 2939ndash2944 2011
[20] M E Trujillo and P E Scherer ldquoAdipose tissue-derived factorsimpact on health and diseaserdquo Endocrine Reviews vol 27 no 7pp 762ndash778 2006
[21] E Maury K Ehala-Aleksejev Y Guiot R Detry A Vanden-hooft and S M Brichard ldquoAdipokines oversecreted by omentaladipose tissue in human obesityrdquo The American Journal ofPhysiology Endocrinology and Metabolism vol 293 no 3 ppE656ndashE665 2007
8 Journal of Immunology Research
[22] V Bourlier A Zakaroff-Girard A Miranville et alldquoRemodeling phenotype of human subcutaneous adiposetissue macrophagesrdquo Circulation vol 117 no 6 pp 806ndash8152008
[23] I M Faust P R Johnson J S Stern and J Hirsch ldquoDiet-induced adipocyte number increase in adult rats a new modelof obesityrdquo The American Journal of Physiology vol 235 no 3pp E279ndashE286 1978
[24] C Henegar J Tordjman V Achard et al ldquoAdipose tissuetranscriptomic signature highlights the pathological relevanceof extracellular matrix in human obesityrdquo Genome Biology vol9 article R14 2008
[25] B Kabon A Nagele D Reddy et al ldquoObesity decreasesperioperative tissue oxygenationrdquo Anesthesiology vol 100 no2 pp 274ndash280 2004
[26] K A Virtanen P Lonnroth R Parkkola et al ldquoGlucose uptakeand perfusion in subcutaneous and visceral adipose tissueduring insulin stimulation in nonobese and obese humansrdquoJournal of Clinical Endocrinology and Metabolism vol 87 no8 pp 3902ndash3910 2002
[27] N Hosogai A Fukuhara K Oshima et al ldquoAdipose tissuehypoxia in obesity and its impact on adipocytokine dysregula-tionrdquo Diabetes vol 56 no 4 pp 901ndash911 2007
[28] J Ye Z Gao J Yin andQHe ldquoHypoxia is a potential risk factorfor chronic inflammation and adiponectin reduction in adiposetissue of obob and dietary obese micerdquo The American Journalof Physiology Endocrinology andMetabolism vol 293 no 4 ppE1118ndashE1128 2007
[29] B Wang I S Wood and P Trayhurn ldquoDysregulation of theexpression and secretion of inflammation-related adipokines byhypoxia in human adipocytesrdquoPflugers Archiv European Journalof Physiology vol 455 no 3 pp 479ndash492 2007
[30] J Yin Z GaoQHeD Zhou ZGuo and J Ye ldquoRole of hypoxiain obesity-induced disorders of glucose and lipidmetabolism inadipose tissuerdquoTheAmerican Journal of Physiology Endocrinol-ogy and Metabolism vol 296 no 2 pp E333ndashE342 2009
[31] Z Yun H L Maecker R S Johnson and A J GiaccialdquoInhibition of PPAR1205742 gene expression by the HIF-1-regulatedgene DEC1Stra13 a mechanism for regulation of adipogenesisby hypoxiardquo Developmental Cell vol 2 no 3 pp 331ndash341 2002
[32] L Liu T P Cash R G Jones B Keith C B Thompson andM C Simon ldquoHypoxia-induced energy stress regulates mRNAtranslation and cell growthrdquo Molecular Cell vol 21 no 4 pp521ndash531 2006
[33] K Zhang and R J Kaufman ldquoFrom endoplasmic-reticulumstress to the inflammatory responserdquoNature vol 454 no 7203pp 455ndash462 2008
[34] B P Tu and J S Weissman ldquoOxidative protein folding ineukaryotes mechanisms and consequencesrdquo Journal of CellBiology vol 164 no 3 pp 341ndash346 2004
[35] C M Haynes E A Titus and A A Cooper ldquoDegradation ofmisfolded proteins prevents ER-derived oxidative stress and celldeathrdquoMolecular Cell vol 15 no 5 pp 767ndash776 2004
[36] A Schaeffler P Gross R Buettner et al ldquoFatty acid-inducedinduction of Toll-like receptor-4nuclear factor-120581B pathway inadipocytes links nutritional signalling with innate immunityrdquoImmunology vol 126 no 2 pp 233ndash245 2009
[37] A Kopp C BuechlerMNeumeier et al ldquoInnate immunity andadipocyte function ligand-specific activation of multiple toll-like receptors modulates cytokine adipokine and chemokinesecretion in adipocytesrdquo Obesity vol 17 no 4 pp 648ndash6562009
[38] O I Vitseva K Tanriverdi T T Tchkonia et al ldquoInducibletoll-like receptor and NF-120581B regulatory pathway expression inhuman adipose tissuerdquoObesity vol 16 no 5 pp 932ndash937 2008
[39] S J Creely P G McTernan C M Kusminski et alldquoLipopolysaccharide activates an innate immune systemresponse in human adipose tissue in obesity and type 2diabetesrdquo The American Journal of Physiology Endocrinologyand Metabolism vol 292 no 3 pp E740ndashE747 2007
[40] A Schaffler J Scholmerich and B Salzberger ldquoAdipose tissueas an immunological organ toll-like receptors C1qTNFs andCTRPsrdquoTrends in Immunology vol 28 no 9 pp 393ndash399 2007
[41] J E Davis N K Gabler J Walker-Daniels and M E SpurlockldquoTlr-4 deficiency selectively protects against obesity induced bydiets high in saturated fatrdquoObesity vol 16 no 6 pp 1248ndash12552008
[42] T Suganami T Mieda M Itoh Y Shimoda Y Kamei and YOgawa ldquoAttenuation of obesity-induced adipose tissue inflam-mation in C3HHeJ mice carrying a toll-like receptor 4 muta-tionrdquo Biochemical and Biophysical Research Communicationsvol 354 no 1 pp 45ndash49 2007
[43] X Prieur T Roszer and M Ricote ldquoLipotoxicity inmacrophages evidence from diseases associated with themetabolic syndromerdquo Biochimica et Biophysica Acta Molecularand Cell Biology of Lipids vol 1801 no 3 pp 327ndash337 2010
[44] P D Cani J AmarM A Iglesias et al ldquoMetabolic endotoxemiainitiates obesity and insulin resistancerdquo Diabetes vol 56 no 7pp 1761ndash1772 2007
[45] S V Monte J A Caruana H Ghanim et al ldquoReduction inendotoxemia oxidative and inflammatory stress and insulinresistance after Roux-en-Y gastric bypass surgery in patientswith morbid obesity and type 2 diabetes mellitusrdquo Surgery vol151 no 4 pp 587ndash593 2012
[46] S Ghoshal J Witta J Zhong W de Villiers and E EckhardtldquoChylomicrons promote intestinal absorption of lipopolysac-charidesrdquo Journal of Lipid Research vol 50 no 1 pp 90ndash972009
[47] P D Cani R Bibiloni C Knauf et al ldquoChanges in gut micro-biota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in micerdquo Diabetesvol 57 no 6 pp 1470ndash1481 2008
[48] G GMuccioli D Naslain F Backhed et al ldquoThe endocannabi-noid system links gut microbiota to adipogenesisrdquo MolecularSystems Biology vol 6 article 392 2010
[49] P D Cani S Possemiers T Van De Wiele et al ldquoChanges ingut microbiota control inflammation in obese mice througha mechanism involving GLP-2-driven improvement of gutpermeabilityrdquo Gut vol 58 no 8 pp 1091ndash1103 2009
[50] A Schaffler and J Scholmerich ldquoInnate immunity and adiposetissue biologyrdquoTrends in Immunology vol 31 no 6 pp 228ndash2352010
[51] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[52] D OrsquoCarroll I Mecklenbrauker P P Das et al ldquoA Slicer-independent role for Argonaute 2 in hematopoiesis and themicroRNA pathwayrdquoGenes and Development vol 21 no 16 pp1999ndash2004 2007
[53] C Xiao L Srinivasan D P Calado et al ldquoLymphoproliferativedisease and autoimmunity in mice with increased miR-17-92expression in lymphocytesrdquo Nature Immunology vol 9 no 4pp 405ndash414 2008
Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Journal of Immunology Research
[22] V Bourlier A Zakaroff-Girard A Miranville et alldquoRemodeling phenotype of human subcutaneous adiposetissue macrophagesrdquo Circulation vol 117 no 6 pp 806ndash8152008
[23] I M Faust P R Johnson J S Stern and J Hirsch ldquoDiet-induced adipocyte number increase in adult rats a new modelof obesityrdquo The American Journal of Physiology vol 235 no 3pp E279ndashE286 1978
[24] C Henegar J Tordjman V Achard et al ldquoAdipose tissuetranscriptomic signature highlights the pathological relevanceof extracellular matrix in human obesityrdquo Genome Biology vol9 article R14 2008
[25] B Kabon A Nagele D Reddy et al ldquoObesity decreasesperioperative tissue oxygenationrdquo Anesthesiology vol 100 no2 pp 274ndash280 2004
[26] K A Virtanen P Lonnroth R Parkkola et al ldquoGlucose uptakeand perfusion in subcutaneous and visceral adipose tissueduring insulin stimulation in nonobese and obese humansrdquoJournal of Clinical Endocrinology and Metabolism vol 87 no8 pp 3902ndash3910 2002
[27] N Hosogai A Fukuhara K Oshima et al ldquoAdipose tissuehypoxia in obesity and its impact on adipocytokine dysregula-tionrdquo Diabetes vol 56 no 4 pp 901ndash911 2007
[28] J Ye Z Gao J Yin andQHe ldquoHypoxia is a potential risk factorfor chronic inflammation and adiponectin reduction in adiposetissue of obob and dietary obese micerdquo The American Journalof Physiology Endocrinology andMetabolism vol 293 no 4 ppE1118ndashE1128 2007
[29] B Wang I S Wood and P Trayhurn ldquoDysregulation of theexpression and secretion of inflammation-related adipokines byhypoxia in human adipocytesrdquoPflugers Archiv European Journalof Physiology vol 455 no 3 pp 479ndash492 2007
[30] J Yin Z GaoQHeD Zhou ZGuo and J Ye ldquoRole of hypoxiain obesity-induced disorders of glucose and lipidmetabolism inadipose tissuerdquoTheAmerican Journal of Physiology Endocrinol-ogy and Metabolism vol 296 no 2 pp E333ndashE342 2009
[31] Z Yun H L Maecker R S Johnson and A J GiaccialdquoInhibition of PPAR1205742 gene expression by the HIF-1-regulatedgene DEC1Stra13 a mechanism for regulation of adipogenesisby hypoxiardquo Developmental Cell vol 2 no 3 pp 331ndash341 2002
[32] L Liu T P Cash R G Jones B Keith C B Thompson andM C Simon ldquoHypoxia-induced energy stress regulates mRNAtranslation and cell growthrdquo Molecular Cell vol 21 no 4 pp521ndash531 2006
[33] K Zhang and R J Kaufman ldquoFrom endoplasmic-reticulumstress to the inflammatory responserdquoNature vol 454 no 7203pp 455ndash462 2008
[34] B P Tu and J S Weissman ldquoOxidative protein folding ineukaryotes mechanisms and consequencesrdquo Journal of CellBiology vol 164 no 3 pp 341ndash346 2004
[35] C M Haynes E A Titus and A A Cooper ldquoDegradation ofmisfolded proteins prevents ER-derived oxidative stress and celldeathrdquoMolecular Cell vol 15 no 5 pp 767ndash776 2004
[36] A Schaeffler P Gross R Buettner et al ldquoFatty acid-inducedinduction of Toll-like receptor-4nuclear factor-120581B pathway inadipocytes links nutritional signalling with innate immunityrdquoImmunology vol 126 no 2 pp 233ndash245 2009
[37] A Kopp C BuechlerMNeumeier et al ldquoInnate immunity andadipocyte function ligand-specific activation of multiple toll-like receptors modulates cytokine adipokine and chemokinesecretion in adipocytesrdquo Obesity vol 17 no 4 pp 648ndash6562009
[38] O I Vitseva K Tanriverdi T T Tchkonia et al ldquoInducibletoll-like receptor and NF-120581B regulatory pathway expression inhuman adipose tissuerdquoObesity vol 16 no 5 pp 932ndash937 2008
[39] S J Creely P G McTernan C M Kusminski et alldquoLipopolysaccharide activates an innate immune systemresponse in human adipose tissue in obesity and type 2diabetesrdquo The American Journal of Physiology Endocrinologyand Metabolism vol 292 no 3 pp E740ndashE747 2007
[40] A Schaffler J Scholmerich and B Salzberger ldquoAdipose tissueas an immunological organ toll-like receptors C1qTNFs andCTRPsrdquoTrends in Immunology vol 28 no 9 pp 393ndash399 2007
[41] J E Davis N K Gabler J Walker-Daniels and M E SpurlockldquoTlr-4 deficiency selectively protects against obesity induced bydiets high in saturated fatrdquoObesity vol 16 no 6 pp 1248ndash12552008
[42] T Suganami T Mieda M Itoh Y Shimoda Y Kamei and YOgawa ldquoAttenuation of obesity-induced adipose tissue inflam-mation in C3HHeJ mice carrying a toll-like receptor 4 muta-tionrdquo Biochemical and Biophysical Research Communicationsvol 354 no 1 pp 45ndash49 2007
[43] X Prieur T Roszer and M Ricote ldquoLipotoxicity inmacrophages evidence from diseases associated with themetabolic syndromerdquo Biochimica et Biophysica Acta Molecularand Cell Biology of Lipids vol 1801 no 3 pp 327ndash337 2010
[44] P D Cani J AmarM A Iglesias et al ldquoMetabolic endotoxemiainitiates obesity and insulin resistancerdquo Diabetes vol 56 no 7pp 1761ndash1772 2007
[45] S V Monte J A Caruana H Ghanim et al ldquoReduction inendotoxemia oxidative and inflammatory stress and insulinresistance after Roux-en-Y gastric bypass surgery in patientswith morbid obesity and type 2 diabetes mellitusrdquo Surgery vol151 no 4 pp 587ndash593 2012
[46] S Ghoshal J Witta J Zhong W de Villiers and E EckhardtldquoChylomicrons promote intestinal absorption of lipopolysac-charidesrdquo Journal of Lipid Research vol 50 no 1 pp 90ndash972009
[47] P D Cani R Bibiloni C Knauf et al ldquoChanges in gut micro-biota control metabolic endotoxemia-induced inflammation inhigh-fat diet-induced obesity and diabetes in micerdquo Diabetesvol 57 no 6 pp 1470ndash1481 2008
[48] G GMuccioli D Naslain F Backhed et al ldquoThe endocannabi-noid system links gut microbiota to adipogenesisrdquo MolecularSystems Biology vol 6 article 392 2010
[49] P D Cani S Possemiers T Van De Wiele et al ldquoChanges ingut microbiota control inflammation in obese mice througha mechanism involving GLP-2-driven improvement of gutpermeabilityrdquo Gut vol 58 no 8 pp 1091ndash1103 2009
[50] A Schaffler and J Scholmerich ldquoInnate immunity and adiposetissue biologyrdquoTrends in Immunology vol 31 no 6 pp 228ndash2352010
[51] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[52] D OrsquoCarroll I Mecklenbrauker P P Das et al ldquoA Slicer-independent role for Argonaute 2 in hematopoiesis and themicroRNA pathwayrdquoGenes and Development vol 21 no 16 pp1999ndash2004 2007
[53] C Xiao L Srinivasan D P Calado et al ldquoLymphoproliferativedisease and autoimmunity in mice with increased miR-17-92expression in lymphocytesrdquo Nature Immunology vol 9 no 4pp 405ndash414 2008
Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Journal of Immunology Research 9
[54] B Zhou S Wang C Mayr D P Bartel and H F Lodish ldquomiR-150 amicroRNA expressed inmature B and T cells blocks earlyB cell development when expressed prematurelyrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 104 no 17 pp 7080ndash7085 2007
[55] B S Cobb T B Nesterova E Thompson et al ldquoT cell lineagechoice and differentiation in the absence of the RNase IIIenzyme Dicerrdquo Journal of Experimental Medicine vol 201 no9 pp 1367ndash1373 2005
[56] C-Z Chen L Li H F Lodish and D P Bartel ldquoMicroRNAsmodulate hematopoietic lineage differentiationrdquo Science vol303 no 5654 pp 83ndash86 2004
[57] Q-J Li J Chau P J R Ebert et al ldquomiR-181a is an intrinsicmodulator of T cell sensitivity and selectionrdquo Cell vol 129 no1 pp 147ndash161 2007
[58] M M W Chong J P Rasmussen A Y Rudensky and DR Littman ldquoThe RNAseIII enzyme Drosha is critical in Tcells for preventing lethal inflammatory diseaserdquo Journal ofExperimental Medicine vol 205 no 9 pp 2005ndash2017 2008
[59] L A OrsquoNeill F J Sheedy and C E McCoy ldquoMicroRNAsthe fine-tuners of toll-like receptor signallingrdquo Nature ReviewsImmunology vol 11 no 3 pp 163ndash175 2011
[60] A J Asirvatham C J Gregorie Z Hu W J Magner andT B Tomasi ldquoMicroRNA targets in immune genes and theDicerArgonaute and ARE machinery componentsrdquoMolecularImmunology vol 45 no 7 pp 1995ndash2006 2008
[61] T Bakheet M Frevel B R G Williams W Greer and K SA Khabar ldquoARED humanAU-rich element-containingmRNAdatabase reveals an unexpectedly diverse functional repertoireof encoded proteinsrdquo Nucleic Acids Research vol 29 no 1 pp246ndash254 2001
[62] A J Asirvatham W J Magner and T B Tomasi ldquomiRNAregulation of cytokine genesrdquo Cytokine vol 45 no 2 pp 58ndash69 2009
[63] D V Chartoumpekis A Zaravinos P G Ziros et al ldquoDifferen-tial expression of microRNAs in adipose tissue after long-termhigh-fat diet-induced obesity in micerdquo PLoS ONE vol 7 no 4Article ID e34872 2012
[64] F J Ortega J M Moreno-Navarrete G Pardo et al ldquoMiRNAexpression profile of human subcutaneous adipose and duringadipocyte differentiationrdquo PLoS ONE vol 5 no 2 Article IDe9022 2010
[65] N Kloting S Berthold P Kovacs et al ldquoMicroRNA expressionin human omental and subcutaneous adipose tissuerdquo PLoSONE vol 4 no 3 Article ID e4699 2009
[66] H M Heneghan N Miller O J McAnena T OrsquoBrien and MJ Kerin ldquoDifferential miRNA expression in omental adiposetissue and in the circulation of obese patients identifies novelmetabolic biomarkersrdquo Journal of Clinical Endocrinology andMetabolism vol 96 no 5 pp E846ndashE850 2011
[67] E Arner N Mejhert A Kulyte et al ldquoAdipose tissue microR-NAs as regulators of CCL2 production in human obesityrdquoDiabetes vol 61 pp 1986ndash1993 2012
[68] H Xie B Lim and H F Lodish ldquoMicroRNAs induced duringadipogenesis that accelerate fat cell development are downreg-ulated in obesityrdquo Diabetes vol 58 no 5 pp 1050ndash1057 2009
[69] P Parra F Serra and A Palou ldquoExpression of adipose MicroR-NAs is sensitive to dietary conjugated linoleic acid treatment inmicerdquo PLoS ONE vol 5 no 9 Article ID e13005 2010
[70] J C Strum J H Johnson J Ward et al ldquoMicroRNA 132 regu-lates nutritional stress-induced chemokine production through
repression of SirT1rdquoMolecular Endocrinology vol 23 no 11 pp1876ndash1884 2009
[71] G Zhuang C Meng X Guo et al ldquoA novel regulator ofmacrophage activation miR-223 in obesity associated adiposetissue inflammationrdquo Circulation vol 125 no 23 pp 2892ndash2903 2012
[72] Q Ge J Gerard L Noel I Scroyen and S M BrichardldquoMicroRNAs regulated by adiponectin as novel targets forcontrolling adipose tissue inflammationrdquo Endocrinology vol153 no 11 pp 5285ndash5296 2012
[73] A Chatzigeorgiou K P Karalis S R Bornstein and TChavakis ldquoLymphocytes in obesity-related adipose tissueinflammationrdquoDiabetologia vol 55 no 10 pp 2583ndash2592 2012
[74] M Poggi J JagerO Paulmyer-Lacroix et al ldquoThe inflammatoryreceptor CD40 is expressed on human adipocytes contributionto crosstalk between lymphocytes and adipocytesrdquo Diabetolo-gia vol 52 no 6 pp 1152ndash1163 2009
[75] R Ogawa C Tanaka M Sato et al ldquoAdipocyte-derivedmicrovesicles contain RNA that is transported intomacrophages and might be secreted into blood circulationrdquoBiochemical and Biophysical Research Communications vol398 no 4 pp 723ndash729 2010
[76] G Muller M Schneider G Biemer-Daub and S WiedldquoMicrovesicles released from rat adipocytes and harboringglycosylphosphatidylinositol-anchored proteins transfer RNAstimulating lipid synthesisrdquo Cellular Signalling vol 23 no 7 pp1207ndash1223 2011
[77] X Chen Y Ba L Ma et al ldquoCharacterization of microRNAs inserum a novel class of biomarkers for diagnosis of cancer andother diseasesrdquoCell Research vol 18 no 10 pp 997ndash1006 2008
[78] Y DrsquoAlessandra P Devanna F Limana et al ldquoCirculatingmicroRNAs are new and sensitive biomarkers of myocardialinfarctionrdquo European Heart Journal vol 31 no 22 pp 2765ndash2773 2010
[79] Y Cheng N Tan J Yang et al ldquoA translational study of cir-culating cell-free microRNA-1 in acute myocardial infarctionrdquoClinical Science vol 119 no 2 pp 87ndash95 2010
[80] M F Corsten R Dennert S Jochems et al ldquoCirculat-ing MicroRNA-208b and MicroRNA-499 reflect myocardialdamage in cardiovascular diseaserdquo Circulation CardiovascularGenetics vol 3 no 6 pp 499ndash506 2010
[81] X Ji R Takahashi Y Hiura G Hirokawa Y Fukushima andN Iwai ldquoPlasma miR-208 as a biomarker of myocardial injuryrdquoClinical Chemistry vol 55 no 11 pp 1944ndash1949 2009
[82] A Zampetaki S Kiechl I Drozdov et al ldquoPlasma MicroRNAprofiling reveals loss of endothelial MiR-126 and other MicroR-NAs in type 2 diabetesrdquo Circulation Research vol 107 no 6 pp810ndash817 2010
[83] A M McDermott H M Heneghan N Miller and M J KerinldquoThe therapeutic potential of microRNAs disease modulatorsand drug targetsrdquo Pharmaceutical Research vol 28 no 12 pp3016ndash3029 2011
[84] JWeiler J Hunziker and JHall ldquoAnti-miRNAoligonucleotides(AMOs) ammunition to target miRNAs implicated in humandiseaserdquo Gene Therapy vol 13 no 6 pp 496ndash502 2006
[85] A Colin M Faideau N Dufour et al ldquoEngineered lentiviralvector targeting astrocytes in vivordquo Glia vol 57 no 6 pp 667ndash679 2009
[86] B D Brown M A Venneri A Zingale L S Sergi and L Nal-dini ldquoEndogenous microRNA regulation suppresses transgeneexpression in hematopoietic lineages and enables stable genetransferrdquo Nature Medicine vol 12 no 5 pp 585ndash591 2006
10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Journal of Immunology Research
[87] L E Bernstein J Berry S Kim B Canavan and S KGrinspoon ldquoEffects of etanercept in patients with themetabolicsyndromerdquo Archives of Internal Medicine vol 166 no 8 pp902ndash908 2006
[88] H Dominguez H Storgaard C Rask-Madsen et al ldquoMetabolicand vascular effects of tumor necrosis factor-120572 blockade withetanercept in obese patients with type 2 diabetesrdquo Journal ofVascular Research vol 42 no 6 pp 517ndash525 2005
[89] J Krutzfeldt N Rajewsky R Braich et al ldquoSilencing ofmicroRNAs in vivowith rsquoantagomirsrsquordquoNature vol 438 no 7068pp 685ndash689 2005
[90] J Elmen M Lindow S Schutz et al ldquoLNA-mediatedmicroRNA silencing in non-human primatesrdquo Nature vol 452no 7189 pp 896ndash899 2008
[91] D Grimm K L Streetz C L Jopling et al ldquoFatality in micedue to oversaturation of cellular microRNAshort hairpin RNApathwaysrdquo Nature vol 441 no 7092 pp 537ndash541 2006
[92] C R Sibley Y Seow and M J Wood ldquoNovel RNA-basedstrategies for therapeutic gene silencingrdquo Molecular Therapyvol 18 no 3 pp 466ndash476 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom