regiospecific nitrosation of n-(terminal) blocked … · - 3 - n 2o 3-induced tryptophan...
TRANSCRIPT

Regiospecific nitrosation of N-(terminal) blocked tryptophan derivatives by N2O3 at
physiological pH
Michael Kirsch*§, Anke Fuchs§, and Herbert de Groot
¥Institut für Physiologische Chemie, Universitätsklinikum, Hufelandstrasse 55, D-45122 Essen
*To whom correspondence should be addressed:
Dr. M. Kirsch
Institut für Physiologische Chemie
Universitätsklinikum Essen
Hufelandstr. 55
D-45122 Essen
Germany
Tel.: 49 - 201 / 723 - 4107
Fax: 49 - 201 / 723 - 5943
E-Mail: [email protected]
§ M. Kirsch and A. Fuchs contributed equally to the work presented here.
RUNNING TITLE: N2O3-induced tryptophan nitrosation
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on January 27, 2003 as Manuscript M300237200 by guest on O
ctober 13, 2018http://w
ww
.jbc.org/D
ownloaded from

- 2 -
N2O3-induced tryptophan nitrosation
SUMMARY
N2O3 formed from nitric oxide in the presence of oxygen attacks thiols in proteins to yield S-
nitrosothiols which are believed to play a central role in NO-signaling. In the present study we
examined the N-nitrosation of N-terminal blocked tryptophan derivatives in the presence of N2O3-
generating systems, such as preformed nitric oxide and nitric oxide-donor compounds in the presence
of oxygen at pH 7.4. Under these conditions N-nitrosation of N-acetyl-tryptophan and lysine-
tryptophan-lysine, respectively, was unequivocally proven by UV-Vis spectroscopy as well as by 15N-
NMR spectrometry. Competition experiments performed with the known N2O3-scavenger morpholine
demonstrated that the selected tryptophan-derivatives were nitrosated by N2O3 with similar rate
constants. It is further shown that addition of ascorbate (vitamin C) induced release of nitric oxide from
N-acetyl-N-nitrosotryptophan as polarographically monitored with a NO-electrode. Theoretical
considerations strongly suggested that the reactivity of protein-bound tryptophan would be high
enough to effectively compete with protein-bound cysteine for N2O3. Conclusively, our data clearly
demonstrate that N2O3 nitrosates the secondary amine function (NIndole) at the indole ring of N-blocked
tryptophan with high reactivity at physiological pH values.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 3 -
N2O3-induced tryptophan nitrosation
INTRODUCTION
Nitric oxide (•NO) is involved in a great variety of physiological and pathophysiological processes,
including the regulation of vascular tonus, immune response and neurotransmission (1). In mammals
the basal level of •NO (in the aqueous phase) is in the nanomolar range (2). However, the local •NO
concentrations can substantially be increased in areas with high NO synthase activity and/or in
hydrophobic regions (3,4), thereby facilitating the formation of dinitrogen trioxide (N2O3) (equations 1
(5) and 2 (6)).
2 •NO + O2
→ 2 •NO2 k1 = 2.9 x 106 M-2 s-1 (Eq. 1)
•NO + •NO2 N2O3 k2 = 1.1 x 109 M-1 s-1 (Eq. 2)
k-2 = 8.03 x 104 s-1
Since N2O3 is highly effective in nitrosating sulfhydryl groups (7) (equations 3 (8) and 4 (2)),
RSH + N2O3 → RSNO + (H+) + NO2- k3 = 6.4 x 106 M-1 s-1 (Eq. 3)
RS- + N2O3 → RSNO + NO2- k4 = 1.8 x 108 M-1 s-1 (Eq. 4)
the putative formation of S-nitrosothiols is now generally believed to be of high physiological
importance in vivo ((9) and references therein). For instance, S-nitrosothiols can operate as nitric
oxide donating compounds in vitro (equation 5, (10)), and they are believed to induce •NO-like
biological activities in vivo, probably by releasing •NO (11,12).
RSNO + ascorbate / Cu2+ → RSNO + products + Cu1+ → •NO + RS— + Cu2+ (Eq. 5)
Further, S-nitrosation of thiols is postulated to induce several functional protein modifications in vivo
(13-18). Beside thiols, secondary amines also react rapidly with N2O3 (7). Consequently, the question
arose whether (protein-bound) tryptophan can be nitrosated at the nitrogen atom of the indole ring at
physiological pH 7.4. It has recently been demonstrated that melatonin (N-acetyl-5-
methoxytryptamine) is N-nitrosated by NaNO2/HCl as well as by •NO/O2 at pH 7.4. However, the
reported rate constant for the latter reaction is rather low (k(melatonin + •NO) = 0.5 M-1s-1 (19)).
Keeping the low physiological concentrations of melatonin in mind (e.g. the maximal concentration in
human serum is ~75 pg/ml (20)), this reaction therefore should not proceed to a significant extent in
vivo. A similar reaction has not been established for tryptophan at pH 7.4 so far. However, there are
indications that the secondary amine of tryptophan (NIndole) can be nitrosated when the primary amine
function (NAlanine) is N-blocked. Consequently, albumin (3,21-23) or, more simply, N-acetyltryptophan
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 4 -
N2O3-induced tryptophan nitrosation
(24) were N-nitrosated by N2O3 generated in situ by HCl/NaNO2 (Scheme 1).
Scheme 1
Nitrosamines in general are well-known carcinogens (25). Similarly, Venitt et al. (26) observed
mutagenic effects of N-acetyl-N-nitrosotryptophan in bacteria. On the other side, as protein-bound N-
nitrosotryptophan was able to both stimulate vasorelaxation and inhibit platelet aggregation (21) there
might be a putative physiological significance of N-nitrosotryptophan-derivatives. This idea, however,
has not been further developed probably because thiols are now believed to be the main physiological
targets for N2O3 in proteins.
In contrast to this opinion we here unequivocally demonstrate the N2O3-mediated nitrosation of
N-blocked tryptophan (N-acetyltryptophan and the tripeptide lysine-tryptophan-lysine) by N2O3 at
physiological pH 7.4. In addition, competition experiments performed with the common N2O3
scavenger morpholine demonstrate that N2O3 reacts fast with N-(terminal)-blocked tryptophan. Finally,
as liberation of •NO is evident after reaction of N -acetyl-N-nitrosotryptophan with ascorbate, N-
nitrosotryptophan is expected to exhibit similar •NO donating capabilities as S-nitrosocysteine.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 5 -
N2O3-induced tryptophan nitrosation
EXPERIMENTAL PROCEDURES
L-tryptophan, N-acetyl-tryptophan, melatonin, lysine-tryptophan-lysine, L-cysteine, morpholine,
piperazine and 15N-labeled sodium nitrite were obtained from Aldrich-Sigma (Deisenhofen, Germany).
MAMA NONOate and spermine NONOate were purchased from Situs (Düsseldorf, Germany).
Experimental conditions - Since nitrosation reactions are sensitive to the presence of metal ions
solutions were exposed to chelating resin (Chelex 100) as described previously (27).
Nitrosation by preformed •NO-gas - Nitric oxide (14•NO) or 15N-labeled nitric oxide (15•NO) were daily
prepared by the addition of glacial acetic acid to an aqueous solution of K4[Fe(CN)6] (0.7 M) containing
either NaNO2 or Na15NO2 (0.72 M) under oxygen-free conditions. The overall stoichiometry is
presented by the equation:
K4 [Fe(CN)6] + Na15NO2 + 2 CH3COOH →
K3[Fe(CN)6] + CH3COOK + CH3COONa + H2O + 15•NO (Eq. 6)
The reliability of this procedure was confirmed by detecting the release of nitric oxide polarographically
with a graphite nitric oxide sensing electrode (World Precision Instruments, Berlin, Germany).
N-blocked derivatives of tryptophan were nitrosated in oxygen-saturated phosphate/triglycine buffer
(250/25 mM, pH 8.0) with the above preformed •NO-gas by bubbling it gently into the solution through
a needle until the pH value had dropped to 7.4.
Nitrosation by NO-donor compounds - MAMA NONOate and spermine NONOate were prepared as
100-fold stock solutions in 10 mM NaOH at 4°C and used immediately. A stock solution of
nitrosocysteine (100 mM) was prepared by S-nitrosation of 100 mM cysteine with equimolar amounts
of NaNO2 in acidic solution (pH 2) at 0°C (24). Nitrosation reactions were performed in 1 ml potassium
phosphate/NaHCO3/CO2 buffer (50 mM/25 mM/5 %, pH 7.4, 37°C) in 35 mm dishes.
Nitrosation by NaNO2 under acidic conditions - Substances were exposed to equimolar concentrations
of sodium nitrite in H2O equilibrated with glacial acetic acid to pH 3.5.
15N-NMR identification of nitrosated products - Immediately after nitrosation sample aliquots were
supplemented with 10 % D2O and analyzed by 15N-NMR spectrometry. The adducts were identified by
50.67 MHz 15N-NMR spectrometry on a Bruker AVANCE DRX 500 instrument. Chemical shifts (δ) are
given in ppm relative to neat nitromethane (δ=0) as external standard.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 6 -
N2O3-induced tryptophan nitrosation
Reactivity of N-acetyl-tryptophan towards N2O3 - Morpholine (0 – 100 mM) or piperazine (0 – 20 mM)
and L-tryptophan (2 mM) or its derivatives were incubated with 0.5 mM MAMA NONOate in potassium
phosphate buffer (50 mM, pH 7.5, 37°C) for 30 min. The reaction products were spectrophotometrically
analyzed at 335 nm (or 346 nm for nitrosomelatonin) with a SPECORD S 100 spectrophotometer from
Analytik Jena (Jena, Germany). A similar experiment was carried out with piperazine as competitor.
Control experiments demonstrated that transnitrosation reactions did not proceed at this pH between
either nitrosomorpholine + N-acetyl-tryptophan, nitrosopiperazine + N-acetyl-tryptophan, morpholine +
N-acetyl-N-nitrosotryptophan or piperazine + N-acetyl-N-nitrosotryptophan, in line with observations
reported by Meyer et al. (28).
Decay kinetic experiments – The decay kinetics of N-acetyl-N-nitrosotryptophan (100 µM) at various
temperatures (15 – 45°C) were determined in air-tight quartz cuvettes by rapid scan monitoring the
UV-Vis absorption at λMax= 335 nm (ε335 = 6100 M-1cm-1, (24)) at 12 – 30 time steps. In between the
recordings the samples were protected from light. The temperature control was maintained at ± 0.1°C.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 7 -
N2O3-induced tryptophan nitrosation
RESULTS
Formation of N-nitrosotryptophan in phosphate buffer pH 7.4 - Evidence exists that at acidic conditions
similar to that in the stomach the NaNO2-dependent nitrosation of either HSA, BSA or the dipeptide
glycine-tryptophan yields N-nitrosotryptophan (21,23). The main nitrosating species in this reaction is
postulated to be dinitrogen trioxide (N2O3) formed from HNO2 dehydration (7,29). Provided that this is
indeed the case, N2O3 formed from •NO in the presence of oxygen at pH 7.4 has strongly to be
expected to nitrosate N-terminal-blocked tryptophan-derivatives, like N-acetyl-tryptophan and peptide-
associated tryptophan (lysine-tryptophan-lysine), too.
In order to verify the nitrosation of N-blocked tryptophan derivatives by N2O3 at pH 7.4 we selected
various N2O3-generating systems. We employed preformed nitric oxide as well as in situ generation of
•NO from S-nitrosocysteine and spermine NONOate, respectively. In these systems, N2O3 is formed
from oxidation of nitric oxide (•NO autoxidation) according to equations 1,2 (see Introduction). Since
the formation of N-acetyl-N-nitrosotryptophan from the reaction of N-acetyl-tryptophan with NaNO2 at
pH 3.5 has been described in detail (24,30) we used this reaction as a reference system. In analogy to
data of Bonnet et al. (24) we recorded the UV-Vis spectrum of N-acetyl-N-nitrosotryptophan with an
absorption maximum at 335 nm (Fig.1A, trace b). Interestingly, an almost identical UV-Vis spectrum
was observed when N-acetyl-tryptophan was allowed to react with preformed nitric oxide in the
presence of oxygen at physiological pH 7.4 (Fig. 1A, trace e). This very strongly indicated that N2O3
can indeed nitrosate the secondary amine of the indole system in tryptophan. Likewise, the UV-Vis
spectrum of N-acetyl-N-nitrosotryptophan was also detected during reaction of N-acetyl-tryptophan
with the above mentioned nitric oxide donating compounds in the presence of air (Fig. 1A, traces c
and d).
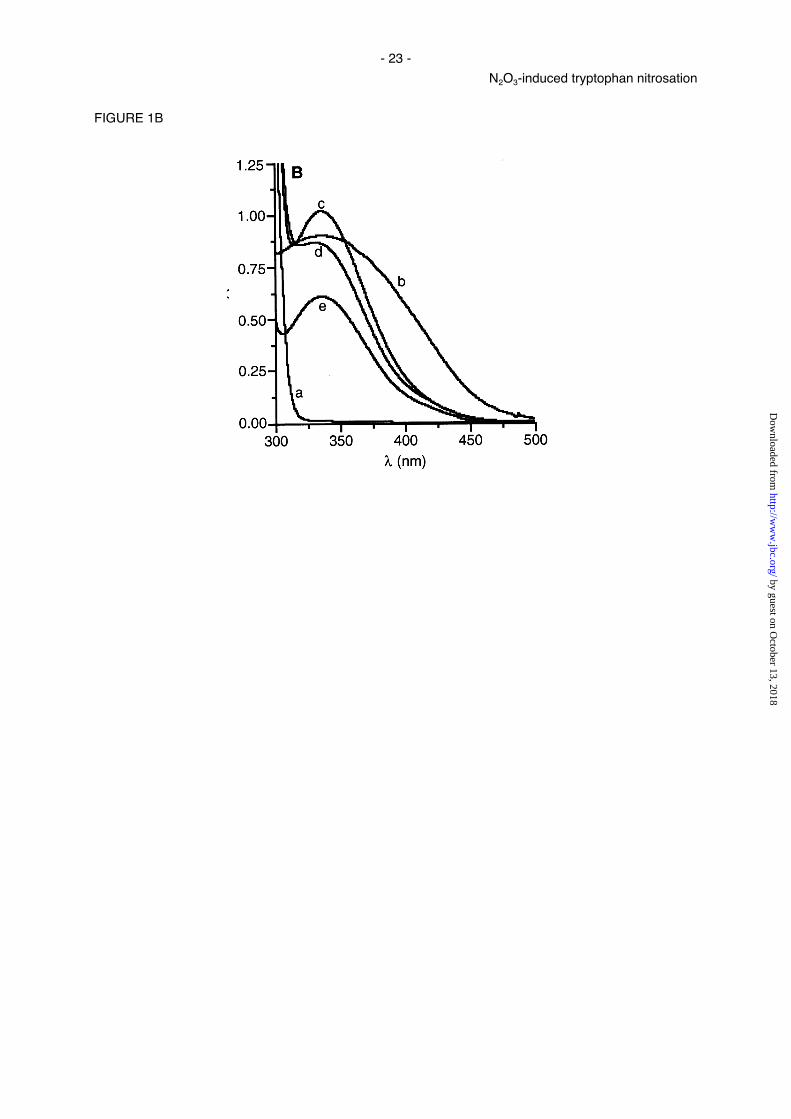
Since in vivo N-terminal-blocked tryptophan is primarily present in proteins, the N2O3-mediated
nitrosation of the nitrogen atom of the indole ring was also investigated for peptide-bound tryptophan
by employing lysine-tryptophan-lysine. In full agreement with the experiments performed with N-acetyl-
tryptophan (see above) all applied N2O3 generating systems were able to effectively nitrosate peptide-
bound tryptophan at the selected pH-values (Fig. 1B). In contrast, the nitrosation of N-unprotected L-
tryptophan by the N2O3-generating systems was much less effective (data not shown) in line with data
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 8 -
N2O3-induced tryptophan nitrosation
from the literature (28,30). Conclusively, only N-terminal blocked tryptophan but not L-tryptophan is a
relevant target for N2O3 at physiological pH.
Detection of N-nitroso-tryptophan by 15N-NMR spectrometry - Since UV-Vis absorption spectra cannot
provide unambiguous evidence that the secondary amine function of the indole ring was truly
nitrosated, we used 15N-NMR spectrometry as a more reliable analytical tool. In 1986, Dorie et al.
recorded the 15N-NMR spectrum of 15N-labeled N-acetyl-15N-nitrosotryptophan from reaction of N-
acetyl-tryptophan with Na15NO2 at pH 4 (31). As commonly observed for N-nitroso compounds (32) the
15N-NMR spectrum exhibited two resonances, at 184.6 ppm and 169.6 ppm relative to neat
nitromethane, of the Z- and E-conformer of N-acetyl-N-nitrosotryptophan, respectively. Figure 2A
shows the 15N-NMR spectrum of preformed (authentic) N-acetyl-15N-nitrosotryptophan at pH 7.4,
exhibiting two resonance lines at 180.8 ppm and 166.3 ppm, respectively. The small shift difference of
approximately 3 ppm compared to the data of Dorie et al. may be explained by differences in the
recording conditions, e.g. different pH values as well as improvements of the 15N-NMR spectrometric
techniques during the past years. Nevertheless, nitrosation of the nitrogen atom at the indole ring is
proven by this characteristic 15N-NMR spectrum. When N-acetyl-tryptophan was reacted with
authentic 15NO in the presence of oxygen, only the two new 15N-NMR resonances at 180.1 ppm and
165.6 ppm were detected, consistent with the formation of N-acetyl-N-nitrosotryptophan (Fig. 2B). To
the best of our knowledge, this is the first direct proof that a tryptophan derivative can be nitrosated by
N2O3 at the nitrogen atom of the indole system at physiological pH.
Analogous to N-acetyl-tryptophan the tripeptide lysine-tryptophan-lysine was nitrosated at pH 7.4
by preformed 15NO as proven by the 1 5N-NMR resonance lines at 181.1 ppm and 166.7 ppm,
respectively (Fig. 2C).
Reactivity of N-blocked tryptophan-derivatives towards N2O3 – For a discussion of the putative
significance of tryptophan nitrosation in vivo it is necessary to determine the rate constants of the
reaction of N-acetyl-tryptophan or its derivatives with N2O3 at pH 7.5. As mentioned above, Blanchard
et al. (19) reported that melatonin reacts with •NO in the presence of oxygen at pH 7.4 with a second-
order rate constant of only 0.5 M-1 s-1. However, these experiments were performed in the presence of
200 mM Hepes buffer which is known to react with reactive nitrogen/oxygen species to yield both H2O2
(33,34) and a •NO-donating compound (35). Therefore we evaluated the rate constants of N-
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 9 -
N2O3-induced tryptophan nitrosation
nitrosation of N-blocked tryptophan derivatives with N2O3 by a competition method, employing
morpholine as N2O3 scavenger (2,36,37). The reaction of 2 mM N-acetyl-tryptophan with 0.5 mM
MAMA NONOate yielded about 180 µM N-acetyl-N-nitrosotryptophan. Morpholine competitively
inhibited the MAMA NONOate-induced nitrosation of N-acetyl-tryptophan in a non-linear manner with
an IC50 value of 13.4 mM (Fig. 3). The protonated form of morpholine, that is morpholinium, with a pKa
value of 8.23 at 37°C (38) is not expected to react with N2O3 (39). Thus, the fraction of unprotonated
morpholine is about 15.7% at pH 7.5. Taken this fraction into account, the true IC50 value is given by
13.4 mM x 0.157 = 2.1 mM. Conclusively, as 2 mM N-acetyl-tryptophan was used (see above), N2O3
reacts at 37°C with virtually identical rate constant with both N-acetyl-tryptophan and morpholine. In
order to verify the presumption that only the unprotonated fraction of the secondary amine effectively
reacts with N2O3, a control experiment was performed with piperazine as a competitive N2O3
scavenger. At pH 7.5 about 99% of piperazine (pKa = 5.55 (7)) exists in the unprotonated form, thus,
as expected, piperazine was more effective than morpholine in inhibiting the MAMA NONOate-
induced nitrosation of N-acetyl-tryptophan (Fig. 3). From inspection of Figure 3 it can be deduced that
MAMA NONOate-induced nitrosation of N-acetyl-tryptophan was half-maximally inhibited at a
piperazine concentration of 2.7 mM. The observation that the corrected IC50 value of morpholine is
lower than the experimental IC50 value of piperazine (Table I), is in agreement with the report that N2O3
reacts faster with morpholine than with piperazine (7). Not unexpected, the reactivity of other N-
terminal blocked tryptophan derivatives towards N2O3 were found to be similar (Table I). In order to
compare the rate constant of these nitrosation reactions with other nitrosation reactions reported in the
literature, one experiment with N-acetyl-tryptophan was performed at 25°C and a rate constant of 4.4 x
107 M-1s-1 was obtained. Thus, N-blocked tryptophan reacts rather fast with N2O3 (see Discussion).
Decomposition of N-acetyl-N-nitrosotryptophan - In contrast to common, stable nitrosamines, protein-
bound N-nitrosotryptophan has been reported to undergo slow decay at pH 2 (23). In order to quantify
this capability at pH 7.4, we analyzed the decomposition of N-acetyl-N-nitrosotryptophan by monitoring
its absorption at 335 nm. At 37°C N -acetyl-N-nitrosotryptophan decayed in a first-order manner
(Figure 4A). Similarly, first-order decay kinetics were also observed at other temperatures (15°C -
45°C, data not shown). From the excellent Arrhenius plot of the rate data (Figure 4B) an activation
barrier of Ea = 13.2 ± 0.1 kcal mol-1 and an A-factor of 1.7 ± 0.3 x 105 s-1 was extracted. This A-factor
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 10 -
N2O3-induced tryptophan nitrosation
appears to be extremely low for a simple first-order (homolysis) decomposition. From the Arrhenius
parameters the half-life of N-acetyl-N-nitrosotryptophan at physiological conditions (T = 37°C, pH 7.4)
can be calculated to 140 min. Thus, the N-nitrosamines of N-blocked tryptophan derivatives are rather
long-lived intermediates.
Since the half-life of peptide-bound N-nitrosotryptophan was found to be similar to that of N-
acetyl-N-nitrosotryptophan (data not shown), one may ask whether it would make any sense for
physiological functions to nitrosate tryptophan in vivo. Recently, Harohalli et al. (23) reported that N-
nitrosotryptophan very slowly releases •NO at pH 2 and Blanchard-Fillion et al. (40) observed that
nitrosomelatonin spontaneously decays at pH 7.4 with release of nitric oxide at a yield of 71%. In our
hands, however, authentic nitrosomelatonin released only negligible amounts of nitric oxide on
decomposition in Hepes-free buffer solution (data not shown). Noticeably neither Harohalli et al. (23)
nor Blanchard-Fillion et al. (40) directly detected •NO e.g., with a NO-electrode.
As, nitrosothiols release nitric oxide in the presence of vitamin C (10), we compared the
potential of ascorbate to induce •NO-release from S -nitrosoglutathione versus N-acetyl-N-
nitrosotryptophan (Fig. 5). In the absence of copper ions, the release of •NO from N-acetyl-N-
nitrosotryptophan after addition of ascorbate was about 5-fold higher than from S-nitrosoglutathione. In
contrast, a “simple” N-nitrosamine like N-nitrosomorpholine did not liberate nitric oxide under similar
conditions (data not shown). The low •NO-releasing efficiency of S-nitrosoglutathione is explained by
the rigorous depletion of copper ions in the applied buffer solution. In the presence of 10 µM Cu2+,
however, S-nitrosoglutathione as well as N-acetyl-N-nitrosotryptophan did release nitric oxide with
nearly the same yield in the presence of ascorbate (data not shown). In the absence of vitamin C only
negligible amounts of nitric oxide were released from either N-acetyl-N-nitrosotryptophan or S-
nitrosoglutathione on decomposition. These results unequivocally demonstrate that N-acetyl-N-
nitrosotryptophan has nitric oxide-releasing capabilities similar to S-nitrosocysteine.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 11 -
N2O3-induced tryptophan nitrosation
DISCUSSION
The results presented above clearly show that N2O3 nitrosates the secondary amine function at the
indole ring of N-blocked tryptophan with high reactivity at physiological pH values. It has been known
before that indole and indole-derivatives are easily attacked by N2O3 under acidic conditions, however,
with exclusive C-nitrosation thus yielding 3–nitroso products (30,41). Such products were not
observed for tryptophan because here the C-3 position is blocked by the alanine residue, rendering
nitrosation at the position N-1 (NIndole) more feasible. Nitrosation at C-2 is only possible for derivatives
carrying powerful electron donors at the C-3 position (30). Our data indicate that N-acetyl-tryptophan,
melatonin, and the tripeptide lysine-tryptophan-lysine are directly nitrosated by N2O3 because these
nitrosation reactions could effectively be inhibited by the N2O3 scavengers morpholine and piperazine,
respectively. Contrary, Turjanski et al. (42) reported that melatonin is mainly nitrosated by a combined
attack of the radicals •NO2 and •NO. It should be noted that such a mechanism has never been proven
to proceed for ordinary amines and that a variety of side products, i.e. N-nitro-, C-nitro, and C-nitroso-
compounds, had to be produced by an operating radical mechanism. Noticeably, the 15N-NMR data
demonstrated that reaction of 15N2O3 with N -blocked tryptophan exclusively yields 1 5N-
nitrosotryptophan.
Recently, the reaction between albumin and N2O3 generated from NaNO2 at low pH has been
monitored by UV-spectroscopy. On the basis of these measurements it was assumed that tryptophan
would be nitrosated too (21,23). However, this conclusion has not been generally accepted (22,43). As
there is presently no specific test for N-nitrosotryptophan available and because N-nitrosotryptophan-
derivatives and S-nitrosocysteine have almost identical UV-Vis absorptions (λmax ca. 335 nm (24) and
ca. 340 nm (30), respectively), the efficiency of nitrosation of both tryptophan and cysteine in proteins
is experimentally hard to verify by UV-Vis spectroscopy. On the other hand, the effectiveness of these
reactions can be reasonably estimated on the basis of the experimental rate constants and
concentrations of both amino acids in proteins. At physiological pH-values tryptophan exists practically
exclusively in the reactive non-protonated form. Hence, the rate constant of N2O3 with protein-bound
tryptophan can be assumed to be k = 4.4 x 107 M-1s-1 (see Results). Contrary, thiols are only
marginally deprotonated at pH 7.4. From the average pKa values of ca. 8.2 of thiolates in proteins (22)
it can be deduced that only a fraction of 13.5% of the cysteine-residues are viable targets for N2O3.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 12 -
N2O3-induced tryptophan nitrosation
Thus, as the rate constants for reaction of thiolates (RS-) with N2O3 is about k = 2 x 108 M-1s-1 (2),
protein-bound cysteine should react at pH 7.4 with N2O3 with a rate constant of k (cysteine + N2O3) = 2
x 108 M-1s-1 x 0.135 = 2.7 x 107 M-1s-1. Thus, the rate constant of reaction of protein-bound tryptophan
with N2O3 is, somewhat unexpected, estimated to be about 63% higher than the rate constant of
protein-bound cysteine with N2O3. In order to verify that this conclusion can be extended to the
reaction rate, the amounts of both tryptophan and cysteine residues in proteins were verified by using
the RCSB Protein Data Bank (44) for those proteins which are believed to be S-nitrosated (Table II).
From the data of Table II it can be deduced that in the selected proteins the total amount of cysteine is
higher than the total amount of tryptophan. With regard to the differences in the rate constants (see
above), one can now estimate that N2O3 reacts primarily (40 – 90%) with tryptophan. Thus, we
hypothesize that protein-bound tryptophan should be preferentially nitrosated under physiological
conditions. In order to verify this prediction, we are currently developing highly sensitive protocols for
the detection of both N-nitrosotryptophan and S-nitrosothiols in proteins.
In this paper we additionally demonstrate that N-nitrosotryptophan has the capability of releasing
nitric oxide in the presence of ascorbate. This implies that N-nitrosotryptophan might operate as a
nitric oxide carrier, a property which to date has been more or less exclusively attributed to S-
nitrosothiols (7,11,45,46). However, as the chemistry of N -nitrosotryptophan derivatives in
physiological environment is not very well developed, it is yet too early to consider N-nitrosotryptophan
derivatives as “harmless” compounds in biological systems. It might be assumed that potentially
harmful reactive nitrogen species like peroxynitrite (40) or N2O3 are deactivated by their reaction with
tryptophan residues and this may them give some antioxidative functions. However, the observation of
Venitt et al. (26) that N-acetyl-N-nitrosotryptophan at 5 – 15 mM induces mutagenicity in bacteria is in
our view easily explained by its capability to release nitric oxide which is known to induce such effects
at unphysiological high concentrations (47). In order not to be misunderstood: harmful effects of N-
nitrosotryptophan-derivatives cannot be ruled out. It should be remembered that harmful reactions are
also known for S-nitrosothiols. For example, it has been reported that S-nitrosothiols react with thiols
to yield nitroxyl (48), which generates hydroxyl radicals and/or peroxynitrite in the absence and in the
presence of oxygen, respectively (49).
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 13 -
N2O3-induced tryptophan nitrosation
Because of the fact that N-nitrosotryptophan-derivatives are rather long-lived yet not “indefinitely”
stable compounds in physiological environment (the hydrolysis of N-acetyl-N-nitrosotryptophan is
expected to yield the harmless products tryptophan and nitrite (28)), one may speculate that this could
further decrease the putative mutagenic potential of N-nitrosotryptophan. On the other hand, the
observed half-life (t 1/2 > 2 h) is so long that N-nitrosotryptophan-derivatives (NIndoleNO) may participate
in physiological processes, e.g. in the transport and release of nitric oxide. In fact, Zhang et al. (21)
observed that peptide-bound N-nitrosotryptophan induces vasorelaxation of rabbit aortic rings, a
function which is typical for freely diffusing nitric oxide (45). Conclusively we hypothesize that a
putative physiological potential of N-nitrosotryptophan should be more important than its
pathophysiological one. This feature has to be clarified in the near future. In any case, we identified,
beside cysteine, a second major target for N2O3 in proteins. This fact will strongly influence the
understanding of protein nitrosation in general.
Acknowledgments - We thank Dr. H.-G. Korth for a series of clarifying discussions pertaining to the
nature of the underlying chemistry and for useful comments on this manuscript. We would also like to
thank Dipl. Ing. H. Bandmann for advice on the NMR technique. The present investigation would have
been impossible without the technical assistance of Ms. Heimeshoff, Ms. Holzhauser, and Ms.
Wensing.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 14 -
N2O3-induced tryptophan nitrosation
References
1. Moncada, S., Palmer, R. M. J., and Higgs, E. A. (1991) Pharmacol. Rev. 43, 109-142
2. Goldstein, S., and Czapski, G. (1996) J. Am. Chem. Soc. 118, 3419 - 3425
3. Nedospasov, A., Rafikov, R., Beda, N., and Nudler, E. (2000) Proc. Natl. Acad. Sci. USA 97,
13543 - 13548
4. Liu, X., Miller, M. J. S., Joshi, M. S., Thomas, D. D., and Lancaster, J. R. (1998) Proc. Natl.
Acad. Sci. USA 95, 2175 - 2179
5. Goldstein, S., and Czapski, G. (1995) J. Am. Chem. Soc. 117, 12078 - 12084
6. Grätzel, M., Taniguchi, S., and Henglein, A. (1970) Ber. Bunsen-Ges. Phys. Chem. 74, 488 -
492
7. Williams, D. L. H. (1986) Nitrosation, Cambridge University Press, Cambridge
8. Keshive, M., Singh, S., Wishnok, J., Tannenbaum, S., and Deen, W. (1996) Chem. Res.
Toxicol. 9, 988-993
9. Stamler, J. S., Lamas, S., and Fang, F. C. (2001) Cell 106, 675-683
10. Holmes, A. J., and Williams, D. L. H. (2000) J. Chem. Soc. Perkin Trans. 2 , 1639-1644
11. Stamler, J. S., Simon, D. I., Osborne, J. A., Mullins, M. E., Jaraki, O., Michel, T., Singel, D. J.,
and Loscalzo, J. (1992) Proc. Natl. Acad. Sci. USA 89, 444-448
12. Ignarro, L. J., Lipton, H., Edwards, J. C., Baricos, W. H., Hyman, H. L., Kadowitz, P. J., and
Gruetter, C. A. (1981) J. Pharmacol. Exp. Ther. 218, 739-749
13. Xu, L., Eu, J. P., Meissner, G., and Stamler, J. S. (1998) Science 279, 234-237
14. Xu, K. Y., Huso, D. L., Dawson, T. M., Bredt, D. S., and Becker, L. C. (1999) Proc. Natl. Acad.
Sci. USA 96, 657-662
15. Park, H.-S., Ho, S.-H., Kim, M.-S., Lee, S. H., and Choi, E.-J. (2000) Proc. Natl. Acad. Sci.
USA 97, 14382-14387
16. DeMaster, E., Redfern, B., Quast, B., Dahlseid, T., and Nagasawa, H. (1997) Alcohol 14, 181-
189
17. Borgs, M., Bollen, M., Keppens, S., Yap, S. H., Stalmans, W., and Vanstapel, F. (1996)
Hepatology 23, 1564-1571
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 15 -
N2O3-induced tryptophan nitrosation
18. Duhe, R., Nielsen, M., Dittman, A., Villacres, E., Choi, E., and Storm, D. (1994) J. Biol. Chem.
269, 7290-7296
19. Blanchard, B., Pompon, D., and Ducrocq, C. (2000) J. Pineal Res. 29, 184-192
20. Arendt, J., Wirz, J. A., and Bradtke, J. (1979) Ann. Clin. Biochem. 16, 307-312
21. Zhang, Y. Y., Xu, A.-M., Nomen, M., Walsh, M., Keaney, J. F., Jr., and Loscalzo, J. (1996) J.
Biol. Chem. 271, 14271-14279
22. Noble, D. R., and Williams, D. L. H. (2001) J. Chem. Soc. Perkin Trans. 2 , 13-17
23. Harohalli, K., Petersen, C. E., Ha, C.-E., Feix, J. B., and Bhagavan, N. V. (2001) J. Biomed.
Sci. 9, 47-58
24. Bonnett, R., and Holleyhead, R. (1974) J Chem Soc Perkin Trans 1, 962-964
25. Lijinski, W., and Epstein, S. S. (1970) Nature 255, 21-23
26. Venitt, S., Crofton-Sleigh, C., Ooi, S. L., and Bonnett, R. (1980) Carcinogenesis 1, 523-532
27. Kirsch, M., and de Groot, H. (2000) J. Biol. Chem. 275, 16702 - 16708
28. Meyer, T., Williams, D., Bonnett, R., and Ooi, S. (1982) J. Chem. Soc. Perkin Trans. 2 , 1383-
1387
29. Garcia-Santos, M. D., Gonzales-Mancebo, S., Hernandez-Benito, J., Calle, E., and Casado, J.
(2002) J Am Chem Soc 124, 2177-2182
30. Bonnett, R., and Nicolaidou, P. (1979) J. Chem. Soc. Perkin Trans. 1 (1969-1974), 1969
31. Dorie, J., Gouesnard, J. P., Mechin, B., and Mellet, P. (1986) J. Mol. Struct. 144, 185 - 189
32. Witanowski, M., Stefaniak, L., and Webb, G. A. (1993) Annual Reports on NMR Spectroscopy.
Nitrogen NMR Spectroscopy (Webb, G. A., Ed.), 25, ACADEMIC PRESS, London
33. Kirsch, M., Lomonosova, E. E., Korth, H.-G., Sustmann, R., and de Groot, H. (1998) J. Biol.
Chem. 273, 12716 - 12724
34. Lomonosova, L. L., Kirsch, M., Rauen, U., and de Groot, H. (1998) Free Radical Biol. Med. 24,
522 - 528
35. Schmidt, K., Pfeiffer, S., and Mayer, B. (1998) Free Radical Biol. Med. 24, 859-862
36. Lewis, R. S., Tannenbaum, S. R., and Deen, W. D. (1995) J. Am. Chem. Soc. 117, 3933 -
3939
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 16 -
N2O3-induced tryptophan nitrosation
37. Kirsch, M., Korth, H.-G., Sustmann, R., and de Groot, H. (2000) Chem. Res. Toxicol. 13, 451 -
461
38. Hetzer, H. B., Bates, R. G., and Robinson, R. A. (1966) J. Phys. Chem. 70, 2869 - 2872
39. Lewis, R. S., Tamir, S., Tannenbaum, S. R., and Deen, W. M. (1995) J. Biol. Chem. 270,
29350 - 29355
40. Blanchard-Fillion, B., Servy, C., and Ducrocq, C. (2001) Free Rad. Res. 35, 857-866
41. Challis, B. C., and Lawson, A. J. (1972) J. Chem. Soc. Perkin Trans. 2 , 918-925
42. Turjanski, A. G., Leonik, F., Estrin, D. A., Rosenstein, R. E., and Doctorovich, F. (2000) J. Am.
Chem. Soc. 122, 10468-10469
43. Feelisch, M., Rassaf, T., Mnaimneh, S., Singh, N., Bryan, N. S., Jourd'heuil, D., and Kelm, M.
(2002) FASEB J. 16, 1775-1785
44. Berman, H. M., Westbrook, J., Feng, Z., Gilliland, G., Bhat, T. N., Weissig, H., Shindyalov, I.
N., and Bourne, P. E. (2000) Nucleic Acids Res. 28, 235 - 242
45. Myers, P. R., Minor, R. L., Guerra, R., Bates, J. N., and Harrison, D. G. (1990) Nature 345,
161-163
46. Stamler, J. S., Simon, D. I., Jaraki, O., Osborne, J. A., Francis, S., Mullins, M., Singel, D., and
Loscalzo, J. (1992) Proc. Natl. Acad. Sci. USA 89, 8087-8091
47. Nguyen, T., Brunson, D., Crespi, C. L., Penman, B. W., Wishnok, J. S., and Tannenbaum, S.
R. (1992) Proc. Natl. Acad. Sci. USA 89, 3030 - 3034
48. Arnelle, D. R., and Stamler, J. S. (1995) Arch. Biochem. Biophys. 318, 279 - 285
49. Kirsch, M., and de Groot, H. (2002) J. Biol. Chem. 277, 13379 - 13388
50. Lai, T. S., Hausladen, A., Slaughter, T. F., Eu, J. P., Stamler, J. S., and Greenberg, C. S.
(2001) Biochemistry 40, 4904-4910
51. Dimmeler, S., Haendeler, J., Nehls, M., and Zeiher, A. M. (1997) J. Exp. Med. 185, 601-607
52. DeMaster, E. G., Redfern, B., Quast, B. J., Dahlseid, T., and Nagasawa, H. T. (1997) Alcohol
14, 181-189
53. Arstall, M. A., Bailey, C., Gross, W. L., Bak, M., Balligand, J. L., and Kelly, R. A. (1998) J. Mol.
Cell. Cardiol. 30, 979-988
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 17 -
N2O3-induced tryptophan nitrosation
54. Jaffrey, S. R., Erdjument-Bromage, H., Ferris, C. D., Tempst, P., and Snyder, S. H. (2001)
Nat. Cell. Biol. 3, 193-197
55. Konorev, E. A., Kalyanaraman, B., and Hogg, N. (2000) Free Radical Biol. Med. 28, 1671-
1678
56. Wolosker, H., Panizzutti, R., and Engelender, S. (1996) FEBS Lett. 392, 274-276
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 18 -
N2O3-induced tryptophan nitrosation
FIGURE LEGENDS
Fig. 1. UV-Vis absorption spectra of N-nitroso-tryptophan residues generated from the
reactions of both N-acetyl tryptophan and lysine-tryptophan-lysine with various NO-sources.
A, spectra of N-acetyl-tryptophan (trace a) and of authentic N-acetyl-N-nitrosotryptophan (trace b) in
air-saturated potassium phosphate buffer/NaHCO3 at pH 7.4. Spectra observed after reaction of N-
acetyl-tryptophan (1 mM) with either spermine NONOate (2 mM) (trace c), S-nitrosocysteine (3.4 mM)
(trace d, dilution 1:4) in the same buffer solution or preformed •NO-gas in oxygenated 250 mM
potassium phosphate buffer pH 8.0/25 mM triglycine (trace e, dilution 1:10). B, spectrum of lysine-
tryptophan-lysine (5mM) (trace a) in air-saturated potassium phosphate/NaHCO3/CO2 buffer (50 mM/25
mM/5 %, pH 7.4, 37°C). Spectrum observed after reaction of lysine-tryptophan-lysine with NaNO2 (5
mM each) at pH 3.5 (trace b). Spectra observed after reaction of lysine-tryptophan-lysine (5 mM) with
either S-nitrosocysteine (3.4 mM) (trace c, dilution 1:2), spermine NONOate (2 mM) (trace d) in air-
saturated potassium phosphate/NaHCO3/CO2 buffer (50 mM/25 mM/5 %, pH 7.4, 37°C) or preformed
•NO-gas in oxygenated potassium phosphate/triglycine buffer (250 mM/25 mM, pH 8.0, 37°C) (trace e,
dilution 1:10). In the latter buffer the pH decreased to 7.4 during experiment.
Fig. 2. 15N-NMR spectra of 15N-labeled N-nitrosotryptophan derivatives.
A, 15N-labeled N-acetyl-N-nitrosotryptophan prepared from reaction of N-acetyl-tryptophan with NaNO2
(20 mM each) in acidified aqueous solution at pH 3.5 (after completion of the reaction, the reaction
mixture was adjusted to pH 7.4 by addition of 1N NaOH). B, 15N-labeled N-acetyl-N-nitrosotryptophan
and C, 15N-labeled lysine-15N-nitrosotryptophan-lysine were produced from reaction of either 100 mM
N-acetyl-tryptophan or 200 mM lysine-tryptophan-lysine with preformed 1 5•NO in oxygenated
potassium phosphate/triglycine buffer (250 mM/25 mM, pH 8.0, 37°C). Under the conditions of B and C
the pH decreased to 7.4 during experiment.
Fig. 3. Influence of morpholine and piperazine on MAMA NONOate-induced nitrosation of N-
acetyl-tryptophan.
MAMA NONOate (0.5 mM) was incubated for 30 min with 2 mM N-acetyl-tryptophan in potassium
phosphate buffer (50 mM, pH 7.5, 37°C) in the presence of various concentrations of A morpholine or
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 19 -
N2O3-induced tryptophan nitrosation
B piperazine, respectively. N-acetyl-N-nitrosotryptophan formation was monitored at 335 nm. Each
value represents the mean +/- SD of 3 determinations.
Fig. 4. Kinetic data of the first-order rate decay of N-acetyl-N-nitrosotryptophan.
A, decay kinetic of 100 µM N-acetyl-N-nitrosotryptophan in potassium phosphate buffer (50 mM, pH
7.4, 37°C). B, Arrhenius plot of the first-order rate constants of decay of 100 µM N-acetyl-N-
nitrosotryptophan in the same buffer. Each value represents the mean +/- SD of 3 determinations.
Fig. 5. Time course of ascorbic acid-induced •NO-liberation from N-acetyl-N-nitrosotryptophan
and S-nitrosoglutathione.
•NO-release from N-acetyl-N-nitrosotryptophan and S-nitrosoglutathione (100 µM each) in potassium
phosphate buffer (50 mM, pH 7.4, 37°C) in the presence of 400 µM ascorbate as electrochemically
monitored with the •NO-electrode.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 20 -
N2O3-induced tryptophan nitrosation
Table I
Rate data for the reaction of tryptophan derivatives with N2O3
N2O3-target competitor T IC50 IC50 k x 107
(apparent) (correcteda) (apparent)
2 mM °C mM mM M-1s-1
N-acetyl-tryptophan morpholine 37 13.4 2.1 6.4
N-acetyl-tryptophan piperazine 37 2.7 2.7 n.c.b
tripeptide morpholine 37 14.8 2.3 6.4
N-acetyl-tryptophan morpholine 25 15.0 1.4 4.4
melatonin morpholine 37 23.4 3.0 9.2
a, corrected for the concentration of the operating amine.
b, n.c.: not calculable.
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 21 -
N2O3-induced tryptophan nitrosation
Table II
Tryptophan and cysteine residues in proteins which may be subject to S-nitrosation in
physiological regulation mechanisms
proteina tryptophan cysteine references
tissue transglutaminase 30 17 (50)
caspase 3 8 21 (51)
aldehyde dehydrogenase 5 5 (52)
tissue plasminogen activator 10 24 (46)
creatine kinase 4 4 (53-56)
ryanodine receptor 56 89 (13)
Ca 2+ ATPase 12 25 (14)
mouse adenylyl cyclase type I 15 28 (18)
c-jun N-terminal kinase 4 8 (15)
a, randomly selected from the RCSB Protein Data Bank (44).
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 22 -
N2O3-induced tryptophan nitrosation
FIGURE 1A
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from
- 23 -
N2O3-induced tryptophan nitrosation
FIGURE 1B
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 24 -
N2O3-induced tryptophan nitrosation
Figure 2
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 25 -
N2O3-induced tryptophan nitrosation
Figure 3
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 26 -
N2O3-induced tryptophan nitrosation
Figure 4A
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 27 -
N2O3-induced tryptophan nitrosation
Figure 4B
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

- 28 -
N2O3-induced tryptophan nitrosation
Figure 5
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Michael Kirsch, Anke Fuchs and Herbert de Grootat physiological pH
Regiospecific nitrosation of N-(terminal) blocked tryptophan derivatives by N2O3
published online January 27, 2003J. Biol. Chem.
10.1074/jbc.M300237200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on October 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from