rapid communication expression of an arabidopsis potassium channel
TRANSCRIPT

Plant Physiol. (1995) 109: 371-374
Rapid Communication
Expression of an Arabidopsis Potassium Channel Gene in Guard Cells'
Robert L. Nakamura, William 1. McKendree, Jr.', Rebecca E. Hirsch, John C. Sedbrook, Richard F. Caber, and Michael R. Sussman*
Program in Cell and Molecular Biology and Department of Horticulture, 1575 Linden Drive (W.L.M., R.E.H., M.R.S.), and Department of Genetics, 445 Henry Mal1 (J.C.S.), University of Wisconsin, Madison,
Wisconsin 53706; and Department of Biochemistry, 21 53 Sheridan Road, Northwestern University, Evanston, lllinois 60208 (R.L.N., R.F.G.)
The Arabidopsis thaliana KATl cDNA encodes a voltage-gated inward-rectifying K+ channel. A KATl genomic DNA clone was isolated and sequenced, and a 5' promoter and coding sequences containing eight introns were identified. Reporter gene analysis of transgenic plants containing the KATl promoter fused to bacterial P-glucuronidase showed robust P-glucuronidase activity primarily in guard cells.
~ ~ _ _ ___
Potassium is utilized by plants and other organisms for regulation of cell membrane potential and turgor. For ex- ample, higher plants require potassium as an osmoticum in the opening of stomata (Schroeder, 1988) and rapid closure of Mimosa leaflets (Kim et al., 1993).
Transport of potassium into and within the plant occurs through membrane proteins, which vary with regard to their affinity for potassium (Kochian et al., 1989; Schroeder and Fang, 1991; Schachtman and Schroeder, 1994) and perhaps their mode of energization (Maathuis and Sanders, 1993). Electrophysiological evidence with protoplasts from leaves and roots has demonstrated that the uptake (Blatt, 1987; Fairley-Grenot and Assmann, 1991; Kourie and Gold- smith, 1992; Maathuis and Sanders, 1993) and efflux (Ketchum and Poole, 1991; Blatt and Armstrong, 1993) of potassium occur through ion channels in the plasma mem- brane. Channel-mediated currents of potassium both into and out of the cell have been detected in guard cells of Vicia faba (Schroeder et al., 1984, 1987; Schroeder, 1988; Fairley- Grenot and Assmann, 1991; Schroeder and Fang, 1991; Blatt and Armstrong, 1993).
Two cDNAs encoding plant potassium channels, KATZ (Anderson et al., 1992) and AKTZ (Sentenac et al., 1992),
' This work was supported by a grant to M.R.S. from the Na- tional Science Foundation (No. DCB-90-04068), by a grant to the University of Wisconsin from the Department of Energy/National Science Foundation/U.S. Department of Agriculture Collaborative Program on Research in Plant Biology (No. BIR 92-20331), and by a grant to R.F.G. from the National Science Foundation (No. MCB- 9406577).
Present address: Florida Genetics Corporation, 12085 Research Drive, Suite 120, Alachua, FL 32615.
* Corresponding author; e-mail mrs8plantpath.wisc.edu; fax 1- 608 -262-4743.
were cloned by their ability to restore potassium uptake to transport-deficient strains of Sacckaromyces cerevisiae (Ramos et al., 1985; Ko and Gaber, 1991). The predicted amino acid sequences of Katl and Aktl are highly related and share sequence identity with members of the voltage- gated potassium channel superfamily (Jan and Jan, 1992). Heterologous expression of KATZ in Xenopus oocytes (Schachtman et al., 1992) and in S. cerevisiae (Bertl et al., 1995) resulted in potassium-selective currents only upon severe polarization of the membrane, demonstrating that the Katl protein functions as a voltage-gated inward-rec- tifying potassium channel.
Voltage-gated potassium channel activity has been cor- related with the opening of guard cells in V. fuba (Schroeder et al., 1987; Schroeder and Fang, 1991). Stimulation of guard cells by light or hormones results in activation of the plasma membrane H+-ATPase, causing a hyperpolariza- tion of the membrane. The resulting potassium influx through voltage-gated channels drives an increase in tur- gor, which causes the guard cells to swell and the stomatal pores to open (reviewed by Assmann, 1993; Kearns and Assmann, 1993).
The need for guard cells to respond to a variety of environmental signals appears to be associated with a highly complex set of regulatory molecules and events. Three types of potassium channels have been demon- strated in guard cells, including an inward-rectifying potassium channel (Schroeder and Fang, 1991), an out- ward-rectifying potassium channel (Blatt, 1988), and a stretch-activated potassium channel (Cosgrove and Hedrich, 1991). The availability of KATZ and AKTZ cDNA clones now permits the analysis of how the genes encoding these proteins are regulated. In this report we show that transgenic Arabidopsis plants containing the KATZ pro- moter fused to GUS express the bacterial reporter gene activity primarily in guard cells, suggesting that this chan- nel is specifically involved in stomatal functioning.
MATERIALS A N D M E T H O D S
Plant Material
Seeds from Arabidopsis thaliana cv RLDl were grown for 2 weeks on germination medium (0.8% [w/v] agar, 1%
371
Dow
nloaded from https://academ
ic.oup.com/plphys/article/109/2/371/6069893 by guest on 25 Septem
ber 2021

372
I I I I I r I I I 1
Nakamura et al. Plant Physiol. Vol. 109, 1995
1 1.0 kb I
xbp I Xba I Hindlll
I 1 MCS a .) 4 - PH1
I I pEl
Figure 1. Restriction map of KAT7 locus and cloning strategy of 5’ regulatory DNA sequence. The pK5 subclone resulted from insertion of the 3.4-kb BamHI/€coRI genomic fragment into the pBluescript multiple cloning site (MCS), and this clone was used to amplify the putative KAT7 promoter. Transiation start i s indicated by the arrow adjacent to the interna1 Hindlll site. Sites of oligonucleotide anneal- ing for PCR amplification of pK5 are shown beneath this subclone. The pH1 and pEl subclones were used to obtain a complete genomic sequence.
[w/v] SUC, 1X Murashige-Skoog medium, 0.5 pg mL-‘ Mes, pH 5.7), and root tissue was harvested and used for transformation according to the method of Valvekens et al. (1988). Each kanamycin-resistant callus was treated as a separate transformed line. Seeds from separate trans- formed lines were incubated on germination medium for 2 d at 4”C, transferred to 24 h of light and 22”C, and grown for 7 to 28 d.
D N A Subcloning and Sequencing
For isolation of a genomic fragment, the cDNA clone of the Arabidopsis potassium channel gene KATZ (Anderson et al., 1992) was used to screen a genomic cosmid library (Olszewski et al., 1988). Seven separate cosmids, which corresponded to three unique cosmid clones based on re- striction digestion analysis, were isolated. Using a 200-bp SpeI-XkoI fragment from the 5’ end of the KATZ cDNA as a radioactive probe, we identified a 4.15-kb BamHI-EcoRI fragment encoding KATl genomic DNA from one cosmid clone. This fragment was isolated and subcloned into pBluescript (Stratagene) for KATl sequence verification and 5’ regulatory DNA isolation. DNA sequence analysis of the subclone, designated pK5, revealed a 3’ EcoRI site at position +530, corresponding to the EcoRI site contained within the open reading frame of the KATZ cDNA (Ander- son et al., 1992). In addition, pK5 contained 3.4 kb of 5’ flanking putative regulatory DNA.
The pK5 subclone was used as a template for PCR am- plification of a promoter fragment, using the T3 primer for forward strand synthesis, and a second primer (5’-GCT- GCGGATCCAGAGATCGACATCTTTTTGA-3’) for re- verse strand synthesis. This primer included a BamHI site for cloning into the BamHI site of the GUS expression vector pBIlOl (Clontech Laboratory, Palo Alto, CA). The fusion included the first four codons of KATZ in frame with the translation initiation of GUS.
Subclones containing an additional 4.0-kb EcoRI frag- ment comprising the remainder of the coding region (pEl) and a 1.0-kb HindIII fragment spanning the EcoRI site at position +530 of the cDNA (pH1) were also isolated from the same cosmid clone to obtain the complete KATZ genomic sequence.
Histochemical Localization of CUS
Seeds from five separate lines of primary transformants were allowed to imbibe for 2 d at 4°C and grown on germination medium containing 100 pg mL-’ kanamycin for 1 to 2 weeks under constant light at 22°C. Seedlings were vacuum infiltrated with GUS staining solution 10.1 M
Na,HPO,/NaH,PO, buffer, pH 7.0, 10 mM Na,EDTA, 1.0 mM K,Fe(CN),, 1.0 mM K,Fe(CN),, 2% (v/v) Triton X-l 00, 0.5 mg mL-’ 5-bromo-4-chloro-3-indolyl-~-glucuronic acid] and incubated overnight at 37°C. Tissue was cleared by treatment with 100% (v/v) ethanol. Pigment resulting from GUS activity was detected by light ~nicroscopy of whole immature seedlings.
RESULTS AND DISCUSSION
A restriction map of the KATZ genomic region is shown in Figure 1. The entire genomic DNA sequence was ob- tained from three overlapping cosmid subclones: pK5, pEl, and pH1. The translation start site is indicated by an arrow following the HindIII site at position -88. The 5’ subclone, pK5, contains a large region of putative 5’ regulatory DNA, in addition to more than 700 nucleotides of the KATZ coding region. The nucleotide sequence of the codirig re- gion within the genomic clone (GenBank accession No. U25088) is identical with the sequence reported for the KATZ cDNA except for the presence of an untranslated 19-bp insert in the cDNA clone (positions -20 to -1). It is likely that this small insert at the extreme 5’ end of the cDNA arose from a cloning artifact during the cDNA clon- ing procedure (Anderson et al., 1992). The location of in- trons in KATZ are shown in Figure 2. KATZ possesses eight introns averaging 100 bp in length.
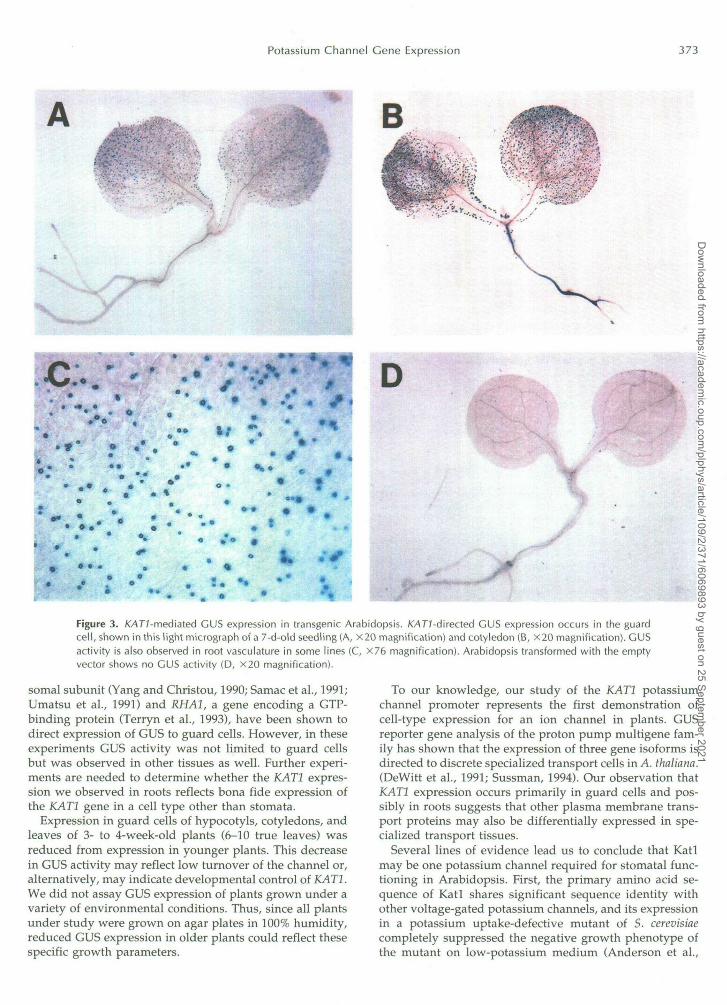
Histochemical localization of GUS activity in KAT1::GUS-transformed plants is shown in Figure 3. In a11 11 KAT1::GUS-transformed lines examined, GUS ez pres- sion occurred abundantly in guard cells in hypocotyls, cotyledons, and leaves of 7- to 12-d-old seedlings (up to two true leaves; Fig. 3, A<). No activity was observed in plants transformed with the vector alone (Fig. 3D). Prelim- inary results indicated that 3- to 4-week-old Nicotiana to- baccum transformed with KAT2::GUS also showed GUS expression in guard cells of leaves (not shown).
In 2 of 11 lines, GUS expression was also observed xn the vascular tissue of the root (Fig. 3B). Staining in the root correlated with strong stomatal expression, suggesting that either excess GUS substrate is transported to and accumu- lates in the root or GUS activity in the root is often below the limit of detection. Promoter sequences from severa1 genes, including the cauliflower mosaic virus 355 ribo-
I I 1 kb :? kb
Figure 2. lntervening sequences and functional domains in Katl . Arrowheads denote intron positions. Black regions indicate trans- membrane domains S1 to S6 (left to right). S4 is the preijumed voltage-sensing domain (asterisk), and the region between S5 and S6 contains the pore-forming region (P).
Dow
nloaded from https://academ
ic.oup.com/plphys/article/109/2/371/6069893 by guest on 25 Septem
ber 2021
Potassium Channel Gene Expression 373
•• - •• ;v-- ,• .. • . -•:. *• /» * t4
*———-X
Figure 3. /0477-mediated GUS expression in transgenic Arabidopsis. KAT/-directed GUS expression occurs in the guardcell, shown in this light micrograph of a 7-d-old seedling (A, X20 magnification) and cotyledon (B, X20 magnification). GUSactivity is also observed in root vasculature in some lines (C, X76 magnification). Arabidopsis transformed with the emptyvector shows no GUS activity (D, X20 magnification).
somal subunit (Yang and Christou, 1990; Samac et al., 1991;Umatsu et al., 1991) and RHA1, a gene encoding a GTP-binding protein (Terryn et al., 1993), have been shown todirect expression of GUS to guard cells. However, in theseexperiments GUS activity was not limited to guard cellsbut was observed in other tissues as well. Further experi-ments are needed to determine whether the KAT1 expres-sion we observed in roots reflects bona fide expression ofthe KAT1 gene in a cell type other than stomata.
Expression in guard cells of hypocotyls, cotyledons, andleaves of 3- to 4-week-old plants (6-10 true leaves) wasreduced from expression in younger plants. This decreasein GUS activity may reflect low turnover of the channel or,alternatively, may indicate developmental control of KAT1.We did not assay GUS expression of plants grown under avariety of environmental conditions. Thus, since all plantsunder study were grown on agar plates in 100% humidity,reduced GUS expression in older plants could reflect thesespecific growth parameters.
To our knowledge, our study of the KAT1 potassiumchannel promoter represents the first demonstration ofcell-type expression for an ion channel in plants. GUSreporter gene analysis of the proton pump multigene fam-ily has shown that the expression of three gene isoforms isdirected to discrete specialized transport cells in A. thaliana.(DeWitt et al., 1991; Sussman, 1994). Our observation thatK.AT1 expression occurs primarily in guard cells and pos-sibly in roots suggests that other plasma membrane trans-port proteins may also be differentially expressed in spe-cialized transport tissues.
Several lines of evidence lead us to conclude that Katlmay be one potassium channel required for stomatal func-tioning in Arabidopsis. First, the primary amino acid se-quence of Katl shares significant sequence identity withother voltage-gated potassium channels, and its expressionin a potassium uptake-defective mutant of S. cerevisiaecompletely suppressed the negative growth phenotype ofthe mutant on low-potassium medium (Anderson et al.,
Dow
nloaded from https://academ
ic.oup.com/plphys/article/109/2/371/6069893 by guest on 25 Septem
ber 2021

3 74 Nakamura et al. Plant Physiol. Vol. 109, I995
1992). Second, heterologous expression of KATZ i n Xenopus oocytes (Schachtman et al., 1992) and more recently in S. cerevisiae (Bertl et al., 1995) showed that the Katl protein confers slowly activating, noninactivating inward currents upon hyperpolarization of the plasma membrane. These same characteristics are observed with the guard cell in- ward rectifier that is associated with stomatal opening and closing i n V. fubu (Schroeder a n d Fang, 1991; Schroeder e t al., 1994). Finally, we show here that KATl-dependent expression of a GUS reporter gene in Arabidopsis results in GUS activity primarily in guard cells i n young seedlings. Experiments are in progress to determine whether environ- mental variables regulating stomatal activity, such as light, water stress, and CO,, will influence patterns or levels of KATl expression.
ACKNOWLEDCMENTS
We thank L. Manney and L. Dickmann for excellent technical assistance and P. Hatfield for the generous donation of pBI101.2 transformed Arabidopsis.
Received May 2, 1995; accepted July 7, 1995. Copyright Clearance Center: 0032-0889/95/ 109/0371/04. The GenBank/EMBL accession number for the sequence reported
in this article is U25088.
LITERATURE ClTED
Anderson JA, Huprikar SS, Kochian LV, Lucas WJ, Gaber RF (1992) Functional expression of a probable Arabidopsis thaliunu potassium channel in Succharomyces cerevisiue. Proc Natl Acad Sci
Assmann SM (1993) Signal transduction in guard cells. Annu Rev Cell Biol 9: 345-375
Bertl A, Anderson JA, Slayman CL, Gaber RF (1995) Use of Sacchuromyces cerevisiue for patch-clamp analysis of heterologous membrane proteins: characterization of Katl, an inward-rectify- ing K+ channel from Arubidopsis thuliana, and comparison with endogenous yeast channels and carriers. Proc Natl Acad Sci USA
Blatt MR (1987) Electrical characteristics of stomatal guard cells: the contribution of ATP-dependent "electrogenic" transport re- vealed by current-voltage and difference-current-voltage analy- sis. J Membr Biol 98: 257-274
Blatt MR (1988) Potassium-dependent bipolar gating of K+ chan- nels in guard cells. J Membr Biol 102: 235-246
Blatt MR, Armstrong F (1993) K+ channels of stomatal guard cells: abscisic acid-evoked control of the outward rectifier mediated by cytoplasmic pH. Planta 191: 330-341
Cosgrove D, Hedrich R (1991) Stretch-activated chloride, potas- sium, and calcium channels coexisting in plasma membranes of guard cells of Vicia fuba L. Planta 186 143-153
DeWitt ND, Harper JF, Sussman MR (1991) Evidence for a plas- ma membrane proton pump in phloem cells of higher plants. Plant J 1: 121-128
Fairley-Grenot K, Assmann SM (1991) Evidence for G-protein regulation of inward K+ channel current in guard cells of fava bean. Plant Cell 1: 1037-1044
Jan LY, Jan YN (1992) Tracing the roots of ion channels. Cell 6 9
Kearns EV, Assmann SM (1993) The guard cell-environment con-
USA 89: 3736-3740
92: 2701-2705
715-718
nection. Plant Physiol 102 711-715
Ketchum KA, Poole RJ (1991) Cytosolic calcium regulates i3 130-
tassium current in corn (Zeu mays) protoplasts. J Membr Biolll9:
Kim HY, Cote GG, Crain RC (1993) Potassium channels in :;ama- neu suman protoplasts controlled by phytochrome and the bio- logical clock. Science 260 960-962
Ko C, Gaber RF (1991) TRKl and TRK2 encode structurally related KC transporters in Saccharomyces cerevisiae. Mo1 Cell Biol 11: 4266-4273
Kochian LV, Schaff JE, Lucas WJ (1989) High affinity KC uptake in maize roots. Plant Physiol 91: 1202-1211
Kourie J, Goldsmith M-H (1992) K' Channels are responsible for an inwardly rectifying current in the plasma membrane 01 rne- sophyll protoplasts of Avenu sutivu. Plant Physiol 98: 1087--1097
Maathuis FJM, Sanders D (1993) Energization of potassium up- take in Arabidopsis thuliana. Planta 191: 302-307
Olszewski N, Martin F, Ausubel FM (1988) Specialized binary vector for plant transformation: expression of the Arabidopsis thaliuna AHAS gene in Nicotiunu tabacum. Nucleic Acids Res 16:
Ramos J, Contreras P, Rodriguez-Navarro A (1985) A potassium transport mutant of Saccharomyces ceuevisiae. Arch Microbioll43:
Samac D, Shah D (1991) Developmental and pathogen-induced activation of the Arabidopsis acidic chitinase promoter. Plant Cell
Schachtman D, Schroeder JI (1994) Structure and transport mech- anism of a high-affinity potassium uptake transporter from higher plants. Nature 370 655458
Schachtman D, Schroeder JI, Lucas WJ, Anderson JA, Gaber RF (1992) Expression of an inward-rectifying potassium channel by the Arubidopsis KATl cDNA. Science 258: 1654-1658
Schroeder JI (1988) K' transport properties of K+ channels in the plasma membrane of Vicia juba guard cells. J Gen Physiol 9 2
Schroeder JI, Fang HH (1991) Inward-rectifying K' channels in guard cells provide a mechanism for low-affinity K+ uptake. Proc Natl Acad Sci USA 88: 11583-11587
Schroeder JI, Hedrich R, Fernandez JM (1984) Potassium-selec- tive single channels in guard cell protoplasts of Vicia faba. Nature 312: 361-362
Schroeder JI, Raschke K, Erwin N (1987) Voltage dependence of K+ channels in guard-cell protoplasts. Proc Natl Acad Sci USA 8 4 4108-4112
Schroeder JI, Ward JM, Gassmann W (1994) Perspectives on the physiology and structure of inward-rectifying K+ channels in higher plants: biophysical implications for K+ uptake. Annu Rev Biophys and Biomol Struct 23: 441471
Sentenac H, Bonneaud N, Minet M, Lacroute F, Salmon J-M, Grignon C (1992) Cloning and expression in yeast of a plant potassium ion transport system. Science 256 663-665
Sussman MR (1994) Molecular analysis of the plant plasma mem- brane. Annu Rev Plant Physiol Plant Mo1 Biol 4 5 211-234
Terryn N, Arias M, Engler G, Tiré C, Villarroel R, Van Moritagu M, Inzé M (1993) rhal, a gene encoding a small GTP binding protein from Arabidopsis, is expressed primarily in developing guard cells. Plant Cell 5: 1761-1769
Umatsu C, Murase M, Ichikawa H, Imamura J (1991) Agrohacte- rium mediated transformation and regeneration of kiwi fruit. Plant Cell Rep 10: 286-290
Valvekens D, Van Montagu M, Van M (1988) Agrobactcrium tumefuciens-mediated transformation of Arubidopsis thaliuna root explants by using kanamycin selection. Proc Natl Acad Sci USA
Yang N, Christou P (1990) Cell type specific expression of a CaMV 35s-GUS gene in transgenic soybean plants. Dev Genet 11:
277-288
10765-10782
88-93
3: 1063-1072
667-683
8 5 5536-5540
289-293
Dow
nloaded from https://academ
ic.oup.com/plphys/article/109/2/371/6069893 by guest on 25 Septem
ber 2021