quantitation of human, choriocarcinoma spheroid … dokumente/john_et_al_1993_in_vitro.pdf ·...
TRANSCRIPT

In Vitro Cell. Dev. Biol. 29A:461-468, June 1993 © 1993 Tissue Culture Association 0883-8364/93 $01.50+0.00
QUANTITATION OF HUMAN, CHORIOCARCINOMA SPHEROID ATTACHMENT TO UTERINE EPITHELIAL CELL MONOLAYERS
NANCY J. JOHN, MANUELA LINKE, AND HANS-WERNER DENKER t
lnstitut fffr Anatomic, Universitfftsklinikum, Hufelandstr. 55, D-4300 Essen 1, Federal Republic of Germany
(Received 12 August 1992; accepted 24 November 1992)
SUMMARY
Adhesive interactions of trophoblast cells with the endometrium are essential for embryo implantation in the uterus. Choriocarcinoma cells, the malignant counterpart of trophoblast, show pronounced invasiveness and are of interest for model studies. We describe here an in vitro model system for the study of adhesion of human JAR choriocarcinoma muhicellular spheroids to different human endometrial epithelial cell lines (RL95-2, HEC-1A, KLE, AN3-CA) grown as monolayers. Cell characterization showed JAR spheroids to secrete the placental hormones human chorionic gonadotropin and progesterone into the culture medium; distinct patterns of keratin, vimentin, and uvomorulin expression were seen in the endometrial cell lines. Spheroid attachment to endometrial monolayers was quantified using a centrifugal force-based adhesion assay, and morphology was examined by light and electron microscopy. Results showed the JAR spheroids to attach to three of the endometrial monolayers (RL95-2, HEC-1A, KLE) progressively over a 24-h period (by which time >80% of the spheroids attached). Significant differences in spheroid attachment were most pronounced at 5 h (RL95-2 > HEC-1A > KLE and poly-D-lysine control, i.e. 90:45:17:17% attached). JAR spheroids did not attach to the endome- trial cell line AN3-CA. Morphology revealed choriocarcinoma cells to begin to intrude between the uterine RL95-2 epithelial cells at 5 h. At 24 h, this intrusive type of penetration continued to be seen only with the RL95-2 monolayer. The assay system thus identifies differences in attachment properties between chorioearcinoma cells and various endometrial cell lines and forms the basis for further studies on the molecular interactions involved.
Key words: adhesion; invasion; choriocarcinoma; endometrial cell lines; in vitro.
INTRODUCTION
The process of embryo implantation involves a complex and dis- tinct series of trophoblast-endometrial interactions. Subsequent to hatching of the blastoeyst from the zona pellueida, the trophoblast establishes contact with the endometrial epithelium and firm attach- ment of these two cell types occurs. In the human, the trophoblast cells may either intrude in between the uterine epithelial cells (6) as they appear to do in monkeys (13), or they may fuse with the cells (30). After this initial interaction, the trophoblast cells come in contact with and invade the basement membrane and the endome- trial stroma. Invasion stops gradually once the trophoblast cells have tapped the maternal vessels and established the hemoehorial placenta, whereby blood from the maternal circulation bathes the embryonal chorionie villi. Of the individual steps that are involved in this process, the initial interaction between trophoblast and uterine epithelium merits particular attention because regulation of implan- tation seems to take place at this level (11).
Information about the cellular, and molecular mechanisms in- volved in the initial adhesion and invasion of the trophoblast into the endometrium during embryo implantation is still rudimentary and is based primarily on species such as the mouse (5,15,35) or rabbit (10,11,42) whereas, understandably, information on the human is
1 To whom correspondence should be addressed.
largely descriptive and experimental data are rare (1,8,27). This gap becomes especially important in light of studies which suggest that considerable morphologic differences exist between the process of implantation in humans and in experimental and domestic ani- mals (31,39).
Choriocarcinoma, malignant cells derived from human tropho- blast of the first trimester placenta, cultured in vitro retain several characteristics of the placental trophoblast (38). Hormone produc- tion and secretion are maintained (3,25,34) and their invasive prop- erties have been exploited to study the interactions of trophoblast with host tissue (19,23,26,41) or with extracellular matrix in in vitro model systems (2,16). In the present communication, we de- scribe an in vitro model system that allows quantitation and examina- tion of the initial adhesion of human choriocarcinoma spheroids to monolayers of uterine endometrial cell lines. Our assay results dem- onstrate that JAR choriocarcinoma spheroids attach to different en- dometrial cell lines with differences in both attachment kinetics and penetration properties. Further studies on the molecular interac- tions underlying these differences are now possible.
MATERIALS AND METHODS
Monolayer cell culture. Human JAR choriocarcinoma cells (American Type Culture Collection HTB 144, Rockville, MD) were maintained in monolayer culture in RPM11640 (GIBCO, Paisley, UK) supplemented with 10% fetal bovine serum (FBS) (Biochrom, Berlin, Germany) and 1% peni-
461

4 6 2 JOHN ET AL.
ENDOMETRIAL CELL MONOLAYER
-trypsin/EDTA
PLATE CELLS ON IPD-LYSINE COATED COVERSLIPS
in 2 cm 2 well (24 well multiwell dish)
CONFRONT WITH CHORIOCARCINOMA SPHEROIDS
-count spheroids/cs -incubate: 1, 5, & 24 his
I 1.5x105 colls/cs (KLE~ 2x10 cells/cs (RL95-2, HEC-1A, AN3-CA)
37o C; 5% CO2/95% air culture 5 dys
37oc; 5% CO2-/95% air
COVERSLIP INTO CONICAL TEST TUBE
-cs removed w/forceps -cell side facing down in tt -cs gently ovedayed w/2 cc
F12 medium
12g, 5 min RCF ----4,,--
CENTRIFUGE (swinging rotor bucket) [ ~ . , , ~
1 QUANTITATE % BOUND ~ - - ~ attached spheroids
F.. J counted in 1 cc PBS -replace cs in multiwell or 2.5% glutaraldehyde
Fm 1. Schematic overview of the centrifugal-force based attachment assay. Endometrial cell lines were grown to confluency on poly-D-lysine coated glass cover shps (cs), and confrontation-cultured with JAR spheroids for the indicated times. Non-attached spheroids were removed by centrifuga- tion, attached spheroids were quantitated as the percentage of the number of spheroids seeded (% bound).
cillin-streptomycin (100 U/ml-100 #g/ml) (GIBCO). The human endome- trial cell lines were routinely cultured as monolayers in: RL95-2 (40) (ATCC, CRL 1671 ), a 1:1 mixture of Dulbecco's modified Eagle;s medium (MEM) (Boehringer Mannbeim, Germany) and Ham's F12 (Biochrom) sup- plemented with 10% FBS, 10 mM HEPES, 5 ttg/ml insuhn (Sigma, Dei- senhofen, Germany); HEC-1A (28) (ATCC, HTB 112) McCoy's 5A me- dium (Biochrom) supplemented with 10% FBS; KLE (ATCC, CRL 1622) a 1:1 mixture of DMEM:F12 supplemented with 10% FBS; and AN3-CA (9) (ATCC, HTB 111) Eagle's MEM with Earle's salts and non-essential amino acids (GIBCO) supplemented with 10% FBS. All endometrial cell line me- dia were additionally supplemented with 1% penicillin-streptomycin (GIBCO). The cells were grown in 250-mt tissue culture flasks (Greiner, Frickenhausen, Germany) at 37 ° C in a humid atmosphere of 5% CO2:95% air. JAR medium was changed every 2 days; endometrial cell medium was changed every 3 days.
100
80
"0 60 t - '
0 .0
o ~ 40
20
0 ~ 0 5 10 15 20 25
time (hrs) FIg. 2. Attachment kinetics of JAR choriocarcinoma spheroids to endo-
metrial cell hnes. JAR spheroids were dehvered onto confluent RL95-2 (open squares), HEC-1A (open circles), KLE (solid circles), or AN3-CA (open triangles) monolayers, or poly-D-lysine-coated control glass cover slips (solid triangles) and at 1, 5, or 24 h, attachment expressed as percent was determined using the centrifugal-force based assay (as described in Mate- rials and Methods and Fig. 1). Each data point represents mean + SEM of triplicate experiments.
Spheroid culture. For chofiocarcinoma spheroid culture, JAR cells were harvested by incubating monolayers in a solution of 0 .05% trypsin/ 0 .02% EDTA (Boehringer Mannheim) for 15 rain at 37 ° C in 5% CO2:95% air. Cells were diluted in JAR medium, and single cell suspen- sions were aliquoted into 25-ml Erlenmeyer tlasks at a concentration of 6
TABLE 1
CHARACTERISTICS OF HUMAN ENDOMETRIAL CELL LINES
Cell Markers, b Doubling
Celt Line time, = h a-Keratin Uvoraorulln Vimentin
RL95-2 47 + 1.7 ~ + - - HEC-1A 45 + 2.1 + + - KLE 78 + 4.5 + - + AN3-CA 45 + 3.1 - - +
= Cells were plated in their respective tissue culture media and the DNA content was analyzed by a diaminobenzoie acid fluorescence assay through a 9 to 10-day time course as described in Materials and Methods.
b a-Keratin, uvomoruhn, or vimentin was detected by indirect immunoflu- orescenee as described in Materials and Methods.
c Values are expressed as means + SEM from triplicate wells.
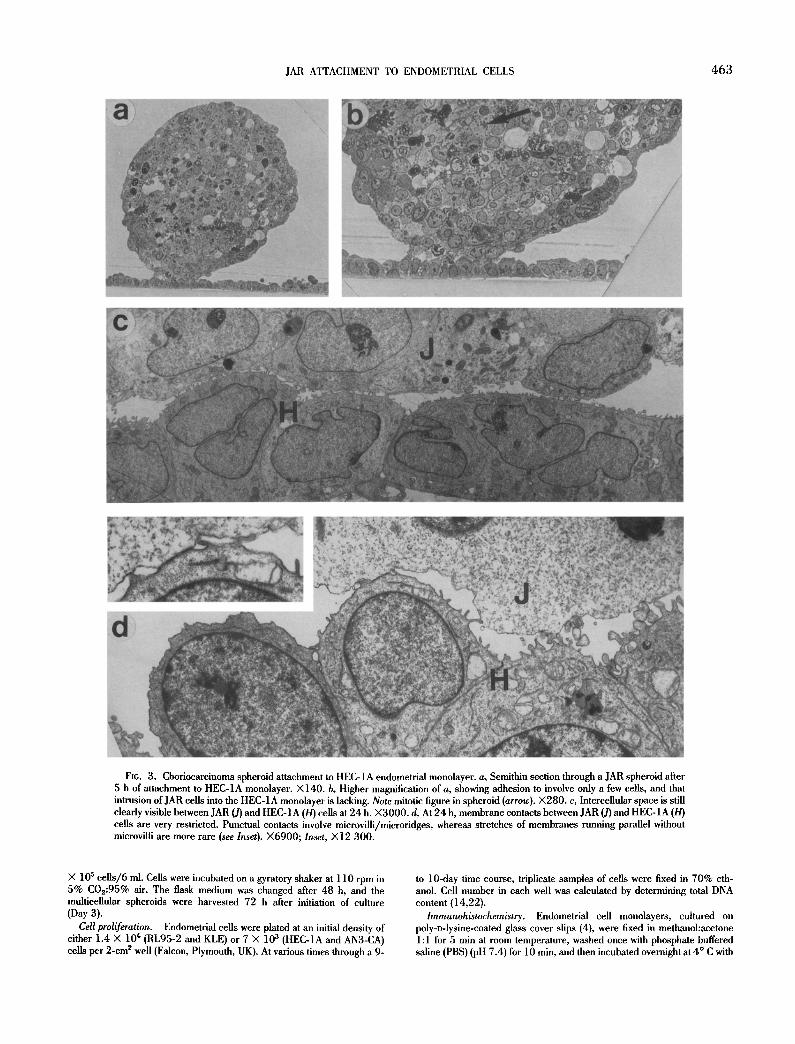
JAR ATTACHMENT TO ENDOMETRIAL CELLS 463
FIG. 3. Choriocarcinoma spheroid attachment to HEC-1A endometrial monolayer, a, Semithin section through a JAR spheroid after 5 h of attachment to HEC-1A monolayer. X140. b, Higher magnification of a, showing adhesion to involve only a few cells, and that intrusion of JAR cells into the HEC-1A monolayer is lacking. Note mitotic figure in spheroid (arrow). ×280. c, Intercellular space is still clearly visible between JAR (J) and HEC-IA (H) cells at 24 h. ×3000 . d, At 24 h, membrane contacts between JAR (J) and HEC-1A (H) cells are very restricted. Punctual contacts involve microvilli/microridges, whereas stretches of membranes running parallel without microvilli are more rare (see Inset). ×6900; Inset, )<12 300.
× 105 ceUs/6 ml. Cells were incubated on a gyratory shaker at 110 rpm in 5% C0~:95% air. The flask medium was changed after 48 h, and the muhieellular spheroids were harvested 72 h after initiation of culture (Day 3).
Cell proliferation. Endometrial cells were plated at an initial density of either 1.4 X 10 ¢ (RL95-2 and KLE) or 7 × l 0 s (HEC-1A and AN3-CA) cells per 2-cm ~ well (Falcon, Plymouth, UK). At various times through a 9-
to 10-day time course, triplicate samples of cells were fixed in 70% eth- anol. Cell number in each well was calculated by determining total DNA content (14,22).
Immunohistochemistry. Endometrial cell monolayers, cultured on poly-o-lysine-coated glass cover slips (4), were fixed in methanol:acetone 1:1 for 5 min at room temperature, washed once with phosphate buffered saline (PBS) (pH 7.4) for 10 min, and then incubated overnight at 4 ° C with

4 6 4 JOHN ET AL.
Fit;. 4. Choriocarcinoma spheroid attachment to KLE endometrial monolayer, a, Phase contrast micrographs of JAR spheroid on confluent monolayer of KLE cells (focused on monolayer), attachment assay zeru-time. Note the morphologically heterogeneous KLE cell population, consisting of small, rounded cells (asterisk) interspersed between large, flat cells (arrows). ×100. b, Semithin section showing focal contact between the superficial JAR spheroid cells and the taller (rounded) subpopulation of KLE cells at 5 h attachment. ×280. c, At 24 h of attachment a reduction of extraceUular space between the JAR and KLE cells was seen but intrusive interaction was not observed. ×280. d, TEM of 5 h attachment between JAR (J) and KLE (K) cells showing punctual contacts and extensive intercellular space. ×6800. Inset shows focal contacts to prevail even at 24 h. X9800.
PBS containing 0.5% bovine serum albumin (Sigma) (PBS/BSA) to block nonspecific binding. The cells were then incubated with mouse anti-a- cytokeratin (Dianova, Hamburg, Germany, recognizing cytokeratin of mo- lecular weight 50-67 kDa; 1:50 in PBS/BSA), mouse monoclonal anti-vi- mentin (DAKO, Hamburg, Germany; 1:15 in PBS/BSA), or mouse anti- uvomorulin (culture supernatant, 1:2, generous gift of W. Birchmeier, Institut fiir Zellbiologie, Universit/itsklinikum, Essen, Germany) (17) for 90 min at room temperature in a moist incubation chamber. After washing twice in PBS/BSA cells were incubated with fluorescein isothiocyanate-con- jugated goat anti-mouse lgG (DAKO) (1:I00 in PBS/BSA) for 60 rain at room temperature, cover slips were washed twice with distilled water and
mounted on slides in glycerin/p-phenylenediamine (24). Slides were exam- ined on a Zeiss photomicroscope equipped for epifluorescence.
Attachment assay. An overview of the assay is shown in Fig. 1. Endo- metrial cell lines RL95-2, HEC-1A, and AN3-CA were plated at an initial cell density of 2 × l0 s and KLE at 1.5 × l0 s in 1 ml of their respective media on poly-~lysine-coated glass cover slips (12 mm diameter), or for subsequent electron microscopy, on thermonox cover slips (Nunc, Naper- ville, IL) in 2--era 2 Falcon multiwells. Medium was changed every 48 h (Days 2 and 4). On Day 5, medium of the confluent endometrial monolayer was replaced with 1 ml JAR medium. After 30 rain, JAR spheroids (average of 20) were gently delivered onto the center of each cover slip in 50 #1 of

JAR ATTACHMENT TO ENDOMETRIAL CELLS 4 6 5
Fro. 5. TEM•fam•n••ayer•ftheend•metria•cell•ineAN3-CAt•whichch•ri•carcin•maspher•idsd•n•tattach.Thesece••ssh•w very little indication of polar organization, with almost no microvilli and poorly developed primitive junctions. Although some cells are considerably flattened against the substratum, others pile up slightly. X3600.
JAR medium. Spheroids were counted (quadrants were drawn under each well to facilitate counting), individually measured using an eyepiece microm- eter, and incubated at 37 ° C in a humid atmosphere of 5% CO2:95% air. At the indicated time points, cover slips containing the confrontation-cul- tures were removed with forceps and placed in 10-ml conical test tubes containing 1 ml RPMI 1640 medium (without supplements). The cover slips were overtayed with 1 ml additional medium, and test tubes were centrifuged (with cell/spheroid surface facing down) in a swinging bucket rotor at room temperature for 5 min, at 12 Xg (modified from the method of 32). Cover slips were then removed and placed into multiwells containing 1 ml PBS with Ca+2/Mg +2. Attached spheroids were counted and expressed as the percentage of the number of spheroids seeded (% bound). In control experiments, cover slips were used that were coated with poly-D-lysine alone rather than with cell monolayers. All experiments were repeated at least 3 times.
Transmission electron microscopy (TEM). Confrontation cultures on thermonox cover slips were fixed in 2.5% glutaraldehyde in 0.1 M cacodyl- ate buffer (pH 7.4) for 30 min. After repeated washing in 0.1 M cacodylate buffer, the specimens were fixed for 1 h in 1% osmium tetroxide in 0.1 cacodylate buffer, dehydrated in graded alcohol and propylene oxide, and embedded in epoxyresin (7). The monolayers were separated from the thermonox cover slip by snap freezing in liquid nitrogen. Semithin (0.4 #m) sections were stained with toluidine blue and uhrathin sections were stained with uranyl acetate and lead citrate.
Hormone production. Culture medium containing secreted hormone products over a 24-h period was collected on ice from the culture flasks after spheroid harvest on Day 3. Samples were centrifuged for 3 min at 600 ×g, and the supernatant was collected and stored at - 2 0 " C until assayed. Human chorionic gonadotropin (hCG) levels were determined using an im- munoenzymatic assay kit (Tandem-E hCG, Hybritech, KiJln, Germany). Progesterone was quantified by a time-resolved fluoroimmunoassay (DEL- FIA, Pharmacia, Turku, Finland). Protein concentrations were determined by the method of Lowry et al. (33) with BSA as a standard.
RESULTS
Attachment. Figure 2 shows the time dependent attachment of JAR spheroids to the various endometrial cell monolayers. JAR spheroids attached rapidly and with high efficiency (83% attached at 1 h) to the RL95-2 monolayer. In contrast, < 3 % of the spheroids had attached to the other endometrial monolayers at 1 h (0% at- tachment to KLE or AN3-CA; 2% attachment to poly-o-lysine; 3% attachment to HEC-1A). At 5 h, 91% of the spheroids had attached to RL95-2; 46% attached to HEC-1A; and 17% to KLE and poly- D-lysine. By 24 h, 98% of the spheroids were attached to the RL95- 2 monolayer, and > 7 2 % had attached to KLE (87%), HEC-1A (84%), and poly-D-lysine (72%). Interestingly, JAR spheroids did
not attach to the AN3-CA monolayer, even after 24 h of confronta- tion.
Endometrial cell line morphology. Parameters such as doubling time and the expressed pattern of epithelial cell markers (a-keratin, uvomorulin, vimentin) differed between the four human endome- trial cell lines (Table 1).
Characteristic expression of a differentiated epithelial phenotype seemed to decrease in the following order: The most differentiated cell lines seemed to be RL95-2 and HEC-1A. RL95-2 (40) exhib- ited well-defined adherens-type junctions, including numerous des- mosomes, intermediate filaments/a-keratin (Table 1), surface mi- crovilli, and estrogen receptors (data not shown). HEC-1A (28) possessed prominent apical surface microvilli and microridges (Fig. 3) and expressed a-keratin and uvomorulin (Table 1). Intermediate in differentiation phenotype, KLE cells possessed well-developed adherens-type junctions and surface microvilli (Fig. 4) and they expressed both a-keratin and vimentin (Table 1). The KLE cell population was morphologically very heterogeneous, with the mono- layer consisting of small, relatively high, rounded cells interspersed between large, fiat cells (Fig. 4 a) which could spread over the surface for extended distances (seen as clear cytoplasmic bridges between cell monolayers) (Fig. 4 b). Cell line AN3-CA (9) seemed most undifferentiated, giving little indication of polar organization and exhibiting only primitive junctions (Fig. 5) and vimentin (Table 1).
JAR choriocarcinoma spheroid morphology. Uniformly shaped JAR multicellular spheroids with an average diameter of 492 __+ 12 #m were produced after 3 days of shaker culture (Figs. 3 a and 4 a). Semithin sections showed the spheroids to be composed of cells that were relatively densely packed and were heterogeneous with respect to size, shape, and nuclear morphology (Figs. 3 a,b and 6 a). Mitotically active cells were seen dispersed throughout the spheroids interspersed with other cells that exhibited various stages of necrosis (Fig. 3 b). However, no accumulation of centralized necrosis was apparent.
JAR cells secreted hCG (20.2 + 1.8 IU, mg -1 cell protein. 24 h - t ) and progesterone (P) (491.6 + 34 pg- mg -1 cell protein- 24 h - l ) into the culture medium.
Attachment morphology. Analysis of semi- and uhrathin sec- tions through the attached spheroids revealed that there were char- acteristic differences between the various confrontation pairs:
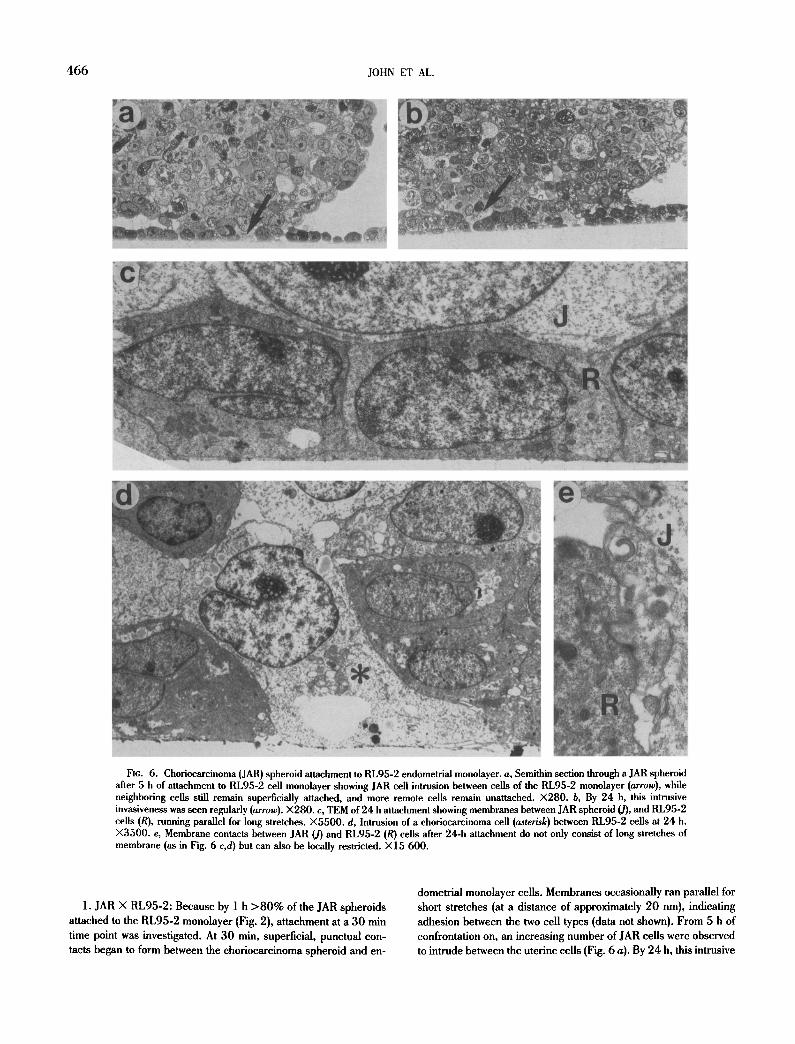
4 6 6 JOHN ET AL.
4 ~
:, - - -
. ' . ,
i
Fig. 6. Chofiocarcinoma (JAR) spheroid attachment to RL95-2 endometrial monolayer, a, Semithin section through a JAR spheroid after 5 h of attachment to RL95-2 cell monolayer showing JAR cell intrusion between cells of the RL95-2 monolayer (arrow), while neighboring cells still remain superficially attached, and more remote cells remain unattached. X280. b, By 24 h, this intrusive invasiveness was seen regularly (arrow).)<280. c, TEM of 24 h attachment showing membranes between JAR spheroid (J), and RL95-2 cells (R), running parallel for long stretches. )<5500. d, Intrusion of a ehoriocarcinoma cell (asterisk) between RL95-2 cells at 24 h. X3500. e, Membrane contacts between JAR (J) and RL95-2 (R) cells after 24-h attachment do not only consist of long stretches of membrane (as in Fig. 6 c,d) but can also be locally restricted. )<15 600.
1. JAR X RL95-2: Because by 1 h > 8 0 % of the JAR spheroids attached to the RL95-2 monolayer (Fig. 2), attachment at a 30 min time point was investigated. At 30 min, superficial, punctual con- tacts began to form between the choriocarcinoma spheroid and en-
dometrial monolayer cells. Membranes occasionally ran parallel for short stretches (at a distance of approximately 20 nm), indicating adhesion between the two cell types (data not shown). From 5 h of confrontation on, an increasing number of JAR cells were observed to intrude between the uterine cells (Fig. 6 a). By 24 h, this intrusive

JAR ATTACHMENT TO ENDOMETRIAL CELLS 467
invasiveness was seen regularly. Membranes ran parallel for long stretches where extracellular space was excluded (Fig. 6 b-e).
2. JAR × HEC-1A: At 5 h, adhesion was limited to focal contact between the JAR and uterine cells (Fig. 3 a,b). On electron micro- graphs, extracellular space was clearly seen to be interspersed be- tween both cell types in most regions. Even after 24 h of confronta- tion-culture, membranes did not run parallel over long stretches, and no intrusion of JAR ceils into the HEC-1A monolayer occurred (Fig. 3 c,d).
3. JAR × KLE: At 5 h, contact was made between the superficial JAR spheroid cells and the small, relatively taller subpopulation of KLE cells. These represented the only sites of attachment; the clear cytoplasmic extensions that interconnected the fiat cell population laterally were entirely free of JAR cell contact (Fig. 4 b). By 24 h, the extracellular space seen between the spheroid and monolayer was reduced, but still clearly seen in most places. Focal contacts were formed between the two cell types, however, intrusive interac- tion was not observed (Fig. 4 c,d).
DISCUSSION
Trophoblast is a cell type that receives interest for mediating attachment and invasion of the mammalian embryo in the uterus. Invasiveness is particularly pronounced and uncontrolled in its ma- lignant counterpart, choriocarcinoma (12,21,36,37). Choriocarci- noma cells retain many of the characteristics of placental tropho- blast (i.e. hormone production and release is maintained). Cultured as spheroids, the cells further develop various types of cell-to-cell contacts that may influence cell differentiation and cell behavior, thereby providing many of the attributes of in vivo tissues that can- not be obtained in monolayer or suspension culture (18,20,29).
With the use of a centrifugal-force-based assay system, we were able to compare and quantitate choriocarcinoma spheroid attach- ment to different uterine monolayers over time after exposing the confrontation-cultures equally to a 12 )<g force. This procedure was adopted after a preliminary series of experiments (using JAR spher- oids and the RL95-2 endometrial monolayer) had shown that after 1 h of confrontation-culture, attachment results were identical whether low or high shear forces (12, 45, or 90 ×g) were used in the attachment assay (data not shown).
In a previous study using a centrifugal-force-based assay to quantitate the attachment strength of single cell suspensions of fibro- blasts and gliomas binding to extracellular matrix proteins (32), 12 )<g was considered a "low detachment force" [equal to approxi- mately 3.6 )< 10 -6 dynes/cell, where dyne force per cell was calcu- lated by F = (specific density of the cell-specific density of the medium) X volume of the cell × relative centrifugal force (RCF)].
The corresponding force (measured in dynes/spheroid) for 12 )<g in our system was considerably greater because we were work- ing with multicellular spheroids with a diameter in excess of 400 #m. Inclusion of this centrifugal step, however, has presumably allowed discrimination between high strength adhesion/specific in- teraction between cells and weak adhesion that may be non-specific. Although manipulation of the cover slip introduced shear forces other than centrifugal, results were reproducible with low experi- mental error (see 4- SEM bars, Fig. 2).
Quantification and comparison of adhesion for the different en- dometrial cell lines over a 24-h period showed remarkable differ- ences with respect to both kinetics and morphology to exist in the
attachment of the JAR spheroids. Of significant interest, even after 24 h of confrontation-culture, JAR spheroids failed to attach to the AN3-CA monolayer.
Morphologic variations in attachment presumably are based on fundamental differences at the cellular and molecular levels. With RL95-2 monolayers, JAR cells established contacts over relatively long stretches of membranes that were running in parallel and at a distance of about 20 nm, thus showing morphologic evidence for a very firm attachment which could also allow for intense direct cell- to-cell signaling. With KLE, contacts were predominately focal; with HEC-1A, only focal. In the case of JAR X RL95-2, initial attachment was followed by an intrusive type of penetration that bears some resemblance to what has been described to occur in vivo in species like the rhesus monkey (13) and in carnivores (39). The in vivo situation in the human is still unclear because adequately fixed and plastic embedded material is unavailable from these initial phases of implantation. Intrusion, however, has been described to be the mode of interaction of the human blastocyst with uterine monolayers in vitro (31). Because our in vitro confrontation system did not allow deep penetration and dispersal of invasive cells, mor- phologic discrimination of trophoblast/choriocarcinoma vs. endo- metrial cells posed no major problems. Therefore, no attempts were made to apply histochemical markers for these cell types.
Our assay system has identified specific human endometrial cell lines that are adhesive vs. non-adhesive for JAR choriocarcinoma cells. The system is useful for further characterization of these cell lines with respect to putative adhesion molecules, thus aiding in the understanding of the factors involved in the early stages of tropho- blast attachment and invasion.
ACKNOWLEDGEMENTS
We thank Dorothea Schiinke for technical assistance with microscopy. This work was supported by a fellowship to N. J. J. from the Alexander yon Humboldt Foundation, Germany. M. L. was supported: by funds from the Mildred Scheel Stiftung to H.-P. Hohn and H.-W. D. (Wl6/90/Ho 1).
REFERENCES
1. Aplin, J. D. Implantation, trophoblast differentiation and haemoehorial placentation: mechanistic evidence in vivo and in vitro. J. Gel] Sei. 96:207-217; 1991.
2. Aphn, J. D.; Charhon, A. K. The role of matrix macromolecules in the invasion of decidua by trophoblast: model studies using BeWo cells. Trophoblast Res. 4:139-158; 1990.
3. Bahn, R. S~; Worsham, A.; Speeg, K. V., et al. Characterization of steroid production in cultured human choriocarcinoma cells. J. Clin. Endocrinol. Metab. 52:447-450; 1981.
4. Bottenstein, J.; Sato, G. Fibronectin and polylysine requirement for proliferation of neuroblastoma cells in defined medium. Exp. Cell Res. 129:361-366; 1980.
5. Carson, D. D.; Wilson, O. F.; Dutt, A. Glycoconjugate expressima and interactions at the cell surface of mouse uterine epithelial cells and periimplantation stage embryos. Trophoblast Res. 4:211-241; 1990.
6. Coutifaris, C.; Babalola, G. O.; Abisogun, A. O., et al. In vitro systems for the study of human trophoblast implantation. In: Bulletti, C.; Gurpide, E., eds. The primate endometrium, v01. 622. New York: New York Academy of Sciences; 1991:191-201.
7. Cross, R. M. M. A reliable epoxyresin mixture and its application in routine biological transmission electron microscopy. Micron Mi- crosc. Acta 20:1-7; 1989.
8. Damsky, C. H.; Fitzgerald, M. L.; Fisher, S. J. Distribution patterns of extracellular matrix components and adhesion receptors are intrica- tely modulated during first trimester eytotrophoblast differentiation

468 JOHN ET AL.
along the invasive pathway, in vivo. J. Clin. Invest. 89:210-222; 1992.
9. Dawe, C. J.; Banfield, W. G.; Morgan, W. D., et al. Growth in continu- ous culture, and in hamsters, of cells from a neoplasm associated with acanthosis nigricans. JNCI 33:441-456; 1964.
10. Denker, H.-W. Basic aspects of ovoimplantation. In: Wynn, R. M., ed. Obstetrics and gynecology annual. Norwalk, CT: Appleton-Century- Crofts; 1983:15-42.
11. Denker, H.-W. Trophoblast-endometrial interactions at embryo im- plantation: a cell biological paradox. Trophoblast Res. 4:3-29; 1990.
12. Elston, C. W. Development and structure of trophoblastic neoplasms. In: Loke, Y. W.; Whyte, A., eds. Biology of trophoblast. New York: Elsevier Science Publishers B. V.; 1983:187-232.
13. Enders, A. C.; Hendrickx, A. G.; Schlafke, S. Implantation in the rhesus monkey: initial penetration of endometrium. Am. J. Anat. 167:275-298; 1983.
14. Firestone, G. L.; John, N. J.; Haffar, O. K., et al. Genetic evidence that the steroid-regulated trafficking of cell surface glycopruteins in rat hepatoma cells is mediated by glucocorticoid-inducible cellular com- ponents. J. Cell. Biochem. 35:271-284; 1987.
15. Fisher, S. J.; Sutherland, A.; Moss, L., et al. Adhesive interactions of murine and human trophoblast cells. Trophoblast Res. 4:115-138; 1990.
16. Foidart, J. M.; Christiane, Y.; Emonard, H. Interactions between the human trophoblast cells and the extraeellular matrix of the endome- trium. Specific expression of a-galactose residues by invasive hu- man trophoblastic cells. Trophoblast Res. 4:159-177; 1990.
17. Frixen, U. H.; Behrens, J.; Sachs, M., et al. E--cadherin-mediated cell-cell adhesion prevents invasiveness of human carcinoma cells. J. Cell Biol. 113:173-185; 1991.
18. Grover, A.; Oshima, R. G.; Adamson, E. D. Epithelial layer formation in differentiating aggregates of F9 embryonal carcinoma cells. J. Cell Biol. 96:1690-1696; 1983.
19. Griimmer, R.; Denker, H.-W. Investigations on the invasion of chorio- carcinoma cells grown as spheroids. Placenta 10:511-512; 1989.
20. Griimmer, R.; Hohn, H.-P.; Denker, H.-W. Choriocarcinoma cell spheroids: an in vitro model for the human trophoblast. Trophoblast Res. 4:97-111; 1990.
21. Hertz, R. Choriocarcinoma and related gestational trophoblastic tu- mors in women. New York: Raven Press; 1978.
22. Hinegardner, R. T. An improved fluorometric assay for DNA. Anal. Biochem. 39:197-201; 1971.
23. John, N. J.; Linke, M.; Denker, H.-W. Retinoic acid decreases attach- ment of JAR choriocarcinoma spheroids to a human endometrial cell monolayer in vitro. Placenta 14:13-24; 1993.
24. Johnson, G. D.; Nogueira Aranjo, M. de C. A simple method of reduc- ing the fading of immunofluorescence during microscopy. J. Im- munol. Methods 43:349-350; 1981.
25. Kato, Y.; Braunstein, G. D. Retinoic acid stimulates placental hormone secretion by choriocarcinoma cell lines in vitro. Endocrinology 128:401-407; 1991.
26. Kliman, H. J.; Feinberg, R. F.; Haimowitz, J. E. Human trophoblast- endometfial interactions in an in vitro suspension culture system. Placenta 11:349-367; 1990.
27. Korhonen, M.; Yliinne, J.; Laitinen, L., et al. Distribution of the oq-ot 6 integrin subunits in human developing and term placenta. Lab. In- vest. 65:347-356; 1991.
28. Kuramoto, H.; Tamura, S.; Notake, Y. Establishment of a cell line of human endometrial adenocarcinoma in vitro. Am. J. Obstet. Gyne- col. 114:1012-1019; 1972.
29. Landry, J.; Freyer, J. P. Regulatory mechanisms in spheroidal aggre- gates of normal and cancerous cells. In: Acker, H.; Carlsson, J.; Durand, R., et al., eds. Spheroids in cancer research. Berlin: Springer-Verlag; 1984:50-60.
30. Larsen, J. F. Electron microscopy of nidation in the rabbit and obser- vations on the human trophoblastic invasion. In: Hubinont, P. O.; Leroy, F.; Robyn, C., el al., eds. Ovo-implantation, human gonado- tropins and prolactin. Basel: S. Karger; 1970:38-51.
31. Lindenberg, S.; Hyttel, P.; Sjogren, A., et al. A comparative study of attachment of human, bovine and mouse blastocysts to uterine epi- thelial monolayer. Human Reprod. 4:446-456; 1989.
32. Lotz, M. M.; Burdsal, C. A.; Erickson, H. P., et al. Cell adhesion to fibronectin and tenascin: quantitative measurements of initial bind- ing and subsequent strengthening response. J. Cell Biol. 109:1795-1805; 1989.
33. Lowry, O. H.; Rosebrough, N. J.; Farr, A. L., et al. Protein measure- ment with the folin phenol reagent. J. Biol. Chem. 193:265-275; 1951.
34. Marth, C.; Brattia, M.; Mueller-Holzner, E., et al. Modulation of secre- tion of human chorionic gonadotropin by biologic response modi- tiers on term placenta and chofiocarcinoma cells. Mol. Biother. 1:140-144.
35. Nose, A.; Takeichi, M. A novel cadherin adhesion molecule: its expres- sion patterns associated with implantation and organogenesls of mouse embryos. J. Cell Biol. 103:2649-2658; 1986.
36. Ober, W. B. Pathology of trophoblastic diseases. Hum. Reprod. 2:143-152; 1987.
37. Pattillo, R. A.; Hussa, R. 0., eds. Human trophoblast neoplasms. New York: Plenum Press; 1984.
38. Pattillo, R. A.; Ruckert, A.; Hussa, R., et al. The JAR cell line-con- tinuous human multihormone production and controls. In Vitro 6:398; 1971.
39. Schlatke, J.; Enders, A. C. Cellular basis of interaction between tropho- blast and uterus implantation. Biol. Reprod. 12:41-65; 1975.
40. Way, D. L.; Grosso, D. S.; Davis, J. R., et al. Characterization of a new human endometrial carcinoma (RL95-2) established in tissue cul- ture. In Vitro 19:147-158; 1983.
41. White, T. E.; Saltzman, R. A.; Di Sant'Aguese, P. A., et al. Human choriocarcinoma (JAR) cells grown as multicellular spheroids. Pla- centa 9:583-598; 1988.
42. Winterhager, E.; Briimmer, F.; Dermietzel, R., et al. Gap junction formation in rabbit uterine epithelium in response to embryo recogni- tion. Dev. Biol. 126:203-211; 1988.