purification of a thermostable alkaline laccase from papaya (carica papaya) using affinity...
TRANSCRIPT

Pp
ND
a
ARRAA
KAPT
1
cvrfbltfimphlfcThia
h0
International Journal of Biological Macromolecules 72 (2015) 326–332
Contents lists available at ScienceDirect
International Journal of Biological Macromolecules
j ourna l ho me pa g e: www.elsev ier .com/ locate / i jb iomac
urification of a thermostable alkaline laccase from papaya (Caricaapaya) using affinity chromatography
ivedita Jaiswal, Veda P. Pandey, Upendra N. Dwivedi ∗
epartment of Biochemistry, University of Lucknow, Lucknow 226007, U.P., India
r t i c l e i n f o
rticle history:eceived 5 May 2014eceived in revised form 11 August 2014ccepted 12 August 2014vailable online 2 September 2014
eywords:lkaline laccaseapaya
a b s t r a c t
A laccase from papaya leaves was purified to homogeneity by a two step procedure namely, heat treat-ment (at 70 ◦C) and Con-A affinity chromatography. The procedure resulted in 1386.7-fold purificationof laccase with a specific activity of 41.3 units mg−1 and an overall yield of 61.5%. The native purifiedlaccase was found to be a hexameric protein of ∼260 kDa. The purified enzyme exhibited acidic and alka-line pH optima of 6.0 and 8.0 with the non-phenolic substrate (ABTS) and phenolic substrate (catechol),respectively. The purified laccase was found to be thermostable up to 70 ◦C such that it retained ∼80%activity upon 30 min incubation at 70 ◦C. The Arrhenius energy of activation for purified laccase was foundto be 7.7 kJ mol−1. The enzyme oxidized various phenolic and non-phenolic substrates having catalytic
−1 −1
hermostability efficiency (Kcat/Km) in the order of 7.25 > 0.67 > 0.27 mM min for ABTS, catechol and hydroquinone,respectively. The purified laccase was found to be activated by Mn2+, Cd2+, Ca2+, Na+, Fe2+, Co2+ and Cu2+
while weakly inhibited by Hg2+. The properties such as thermostability, alkaline pH optima and metal tol-erance exhibited by the papaya laccase make it a promising candidate enzyme for industrial exploitation.
. Introduction
Laccase (benzenediol: oxygen oxidoreductase, EC 1.10.3.2) is aopper-containing enzyme that catalyzes the oxidation of a wideariety of organic and inorganic compounds by coupling it to theeduction of oxygen to water. It is widely distributed in bacteria,ungi, plants, and insects [1]. Extensive studies have been made onacterial and fungal laccases as compared to plant laccases. Though
accase was first discovered in exudates of the Japanese lacquerree, Rhus vernicifera [2], only a few plant laccases have been puri-ed and characterized till date. Structurally, laccases can be eitheronomeric, dimeric or multimeric containing four copper atoms
er monomer and varied molecular weight [3–6]. Plant laccasesave been reported to show higher molecular mass than fungal
accases which may be due to a higher extent of glycosylation thanungal ones [7]. The glycosylation has been reported to play a role inopper retention, thermal stability and activity of the enzyme [8].he temperature, pH optima and substrate specificity of laccases
ave been found to vary depending on the source [9–13]. Laccases a metalloenzyme requiring four copper atoms for its catalyticctivity [14]. Other metal ions such as Na+, Ca2+, Mg2+, Mn2+, Fe2+,
∗ Corresponding author. Tel.: +91 522 2740132; fax: +91 522 2740132.E-mail address: [email protected] (U.N. Dwivedi).
ttp://dx.doi.org/10.1016/j.ijbiomac.2014.08.032141-8130/© 2014 Elsevier B.V. All rights reserved.
© 2014 Elsevier B.V. All rights reserved.
Co2+, Hg2+, etc. have been reported to either activate or inhibit thelaccases depending upon the source [1,15].
Laccase has been a focus of attention since it can be usedfor diverse biotechnological applications such as dye decolour-ization, biopulping, biobleaching, degradation of xenobiotics, foodprocessing, biopolymer modification, ethanol production, develop-ment of biosensors, drug synthesis and organic synthesis, etc. [1,6].Most of the laccases isolated so far either exhibit lower yield ofenzyme activity or are found sensitive to extreme conditions oftemperature, pH, metal ions, etc. which result in loss of catalyticactivity limiting their large-scale commercial and industrial appli-cations. Thus, thermostability is one of the general demands ofenzymes which not only offers the enzymes to work at high pro-cess temperatures with higher reaction rates but also reduces therisk of microbial contamination [16]. Thermostable laccases is ofdemand in biobleaching of pulp and treatment of coloured indus-trial effluents [17,18]. Such thermostable laccases have generallybeen reported from thermophilic bacteria and fungi [16]. However,reports on plant laccases as a source of thermostable enzymes arevery scanty.
Besides, other obstacles which impede the practical application
of laccases in biotechnological industries is the lowered laccaseactivity in neutral to alkaline pH as well as in the presence ofvarious metal ions which is generally found in wastewaters dis-charged from textile industries [19,20]. Thus, use of laccases active
Biolog
aiiipci
mapA
2
2
Sgmm
2
mc3ttibai1shia
i
2
lbat4oa
2
c
2
wbcp
N. Jaiswal et al. / International Journal of
t alkaline pH is another ‘green technology alternative’ over chem-cal treatment as many industrial applications such as the textilendustry (for denim bleaching), cosmetic industry (for hair colour-ng), and waste water treatment, etc. occur at neutral to alkalineH [21,22]. Thus, there is a need to explore newer sources of lac-ases with improved properties to facilitate more bio-catalytic andndustrial process applications.
The present study therefore reports purification of a ther-ostable, alkaline and metal-tolerant laccase from the leaves of
n easily available and economically important medicinal plant,apaya (Carica papaya) employing heat treatment followed by Con-
affinity chromatography.
. Experimental
.1. Chemicals and plant material
Unless otherwise stated, all chemicals were purchased fromigma–Aldrich, USA and were of certified reagent grade. Freshreen and young papaya leaves growing in the garden of Depart-ent of Biochemistry, University of Lucknow, were used as plantaterial.
.2. Enzyme assay and protein estimation
Laccase activity was determined by incubating the reactionixture containing a suitable enzyme aliquot and 10 mM cate-
hol (substrate) in 100 mM Tris–HCl buffer (pH 7.5) for 30 min at7 ◦C [23]. The increase in absorbance due to oxidation of catecholo o-benzoquinone was measured at 390 nm using UV–vis spec-rophotometer (Elico SL-177). A parallel control containing all thengredients of the assay system, except the enzyme, was used aslank and those without substrate were used as control. Enzymectivity was expressed in terms of units. One unit of enzyme activ-ty was defined as the amount of enzyme required to produce
�mol of o-benzoquinone (ε = 1260 M−1 cm−1) in 1 min under thepecified conditions. Similarly, enzyme activity using substratesydroquinone and ABTS, were determined by measuring increase
n absorbance at 390 nm (ε for p-benzoquinone = 2240 M−1 cm−1)nd 420 nm (ε for ABTS+• = 36,000 M−1 cm−1), respectively.
Protein concentration was estimated by the Bradford dye bind-ng method using bovine serum albumin as the standard [24].
.3. Enzyme isolation from papaya leaves
A 30% crude extract was prepared by homogenizing 20 g papayaeaves in 60 ml Tris–HCl buffer (100 mM, pH 7.5) using an ice-coldlender. Solid PVP (polyvinylpyrrolidone, insoluble; 0.1% (w/v))nd 7 mM �-mercaptoethanol were added at the time of extrac-ion. The homogenate was centrifuged at 8500 × g for 30 min at◦C using a Sigma 4K15 centrifuge. The clear supernatant that wasbtained (crude extract) was subjected to purification. All the oper-tions were done at 4 ◦C, unless and otherwise specified.
.4. Purification of laccase
The following two quick steps were undertaken to purify therude enzyme preparation.
.4.1. Heat treatmentThe crude extract (50 ml) was heated at 70 ◦C for 30 min in a
ater bath and was immediately chilled in crushed ice followedy centrifugation at 8500 × g for 15 min at 4 ◦C using Sigma 4K15entrifuge. The supernatant was collected and subjected to furtherurification step.
ical Macromolecules 72 (2015) 326–332 327
2.4.2. Concanavalin-A affinity chromatographyThe heat treated enzyme (48 ml) was loaded onto a
Concanavalin-A CL Agarose column (5 cm × 2.5 cm) (Genei,Bangalore, India) pre-equilibrated with 100 mM Tris–HCl buffer(pH 7.5) containing 0.5 M NaCl, 0.1 mM CaCl2 and 0.1 mM MnCl2.The bound proteins were eluted from the column by a linear(50–300 mM) sucrose gradient in the same buffer. The fractionscontaining high laccase activity were pooled and stored at 4 ◦C forfurther use.
2.5. Native PAGE and in-gel activity staining
Native polyacrylamide gel electrophoresis (PAGE; 7.5%) wasperformed in order to check the homogeneity of the preparationusing resolving gel (7.5%), stacking gel (3%) and Tris–glycine run-ning buffer (pH 8.8) as described by Davis [25]. The enzyme sampleswere loaded into the wells of gel and electrophoresis was carriedout at 4 ◦C at constant power of 100 V for 90 min. In-gel activitystaining of the laccase was done by immersing the gel (after 10%native PAGE) in a catechol solution (50 mM) containing Tris–HClbuffer (100 mM, pH 7.5) until brown coloured band appeared.
2.6. Native and subunit molecular weight determination
Native molecular weight of the purified laccase was deter-mined by gel filtration chromatography [26] using Sephadex G-200column chromatography. The void/unoccupied volume (Vo) ofthe sephadex column was determined using blue dextran. Cata-lase (240 kDa), �-amylase (200 kDa), phosphorylase b (97.4 kDa),bovine serum albumin (67 kDa) and lysozyme (14.3 kDa) were usedas standard proteins (1.0 mg/ml) and were applied onto the col-umn and the amount of protein in column eluent was estimated byBradford’s method [24]. The elution volume (Ve) of each standardprotein as well as purified laccase was measured. The molecularweight of the purified laccase was calculated from a calibrationcurve obtained by plotting the log molecular weight of the standardproteins against the ratio of the elution volumes of the standardproteins and the void/unoccupied volume of the column (Ve/Vo).
The subunit molecular mass was determined using SDS PAGEcarried using a 7.5% resolving gel, 3% stacking gel and Tris–glycinerunning buffer (pH 8.8) run at a constant power of 100 V for90 min according to the method of Laemmli [27]. Proteins werevisualized by silver staining. Standard protein markers containingmyosin (205 kDa), phosphorylase b (97.4 kDa), bovine serum albu-min (66 kDa), ovalbumin (43 kDa), and carbonic anhydrase (29 kDa)were used.
2.7. Effect of temperature
Laccase activity at various temperatures (10–90 ◦C) was inves-tigated under standard assay conditions as described earlier inSection 2. The thermostability of the papaya laccase was deter-mined by incubating the enzyme at different temperatures (50, 60,70 and 80 ◦C) and assaying the activity at various time intervals.
2.8. Effect of pH
The effect of pH on the enzyme activity was studied over apH range of 6.5–9.0 with the phenolic substrate, catechol and5.0–9.0 with the non-phenolic substrate, ABTS. The different buffers(100 mM) used were sodium acetate, pH 6.5; Tris–HCl, pH 7.0, 7.5,
8.0 and 8.5; and sodium borate, pH 9.0. The pH stability of thepapaya laccase was determined by estimating the activity afterincubation of the enzyme at different pH values (6.5–9.0) for 24 hunder standard assay conditions.
3 Biological Macromolecules 72 (2015) 326–332
2
p(rr(w
2
taatm
3
3
wloljpalgbityCCsegc
3p
ftP(gpbia
b6lomhp
Fig. 1. (A) Native-PAGE analysis and silver staining of purified papaya laccase dur-
28 N. Jaiswal et al. / International Journal of
.9. Kinetic constants of papaya laccase
Kinetic constants of papaya laccase for the phenolic and non-henolic substrates, namely catechol, ABTS and hydroquinone0–50 mM) were investigated under standard assay conditions,espectively. Km and Vmax values were obtained by non-linearegression of a plot of enzyme activity vs substrate concentrationhyperbolic Michaelis–Menten plot) using GraphPad Prism soft-are.
.10. Effect of various effectors on laccase activity
The effect of varying concentrations (0–10 mM) of several effec-ors, Na+, Mg2+, Ca2+, Mn2+, Cu2+, Co2+, Fe2+, Cd2+, Hg2+, DTT, SDSnd EDTA on laccase activity was determined by performing thectivity assay under standard assay conditions in the absence (con-rol) as well as presence of the individual effectors in the reaction
ixture.
. Results and discussion
.1. Purification of laccase
For purification of laccase, crude homogenate of papaya leavesere initially subjected to heat treatment at 70 ◦C for 30 min which
ed to removal of more than 96% proteins with a recovery of ∼80%f the laccase activity corresponding to ∼22 fold purification ofaccase. This partially purified laccase preparation was further sub-ected to Con-A affinity chromatography which led to ∼1377 foldurification of laccase with overall recovery of ∼62% and a specificctivity of ∼41 units/mg (Table 1). The homogeneity of the purifiedaccase was established by running a native PAGE where a sin-le band was obtained (Fig. 1A). The purified laccase was found toe catalytically active as established through in-gel activity stain-
ng (Fig. 1B). Laccase from Trichoderma harzianum was purifiedo 151.7 fold by Con-A affinity chromatography with only 0.39%ield and 130.5 units/mg specific activity [9]. Similarly, laccase fromhaetomium thermophilum was purified to 10-fold using the sameon-A affinity chromatography with 40% yield and 37 units/mgpecific activity [28]. However, the two-step purification methodmployed in our study resulted in purified papaya laccase withood yield, thus serving for various industrial applications of lac-ases.
.2. Native and subunit molecular weight determination ofapaya laccase
The native molecular weight of the purified papaya laccase wasound to be ∼260 kDa by gel filtration chromatography (Fig. 2). Forhe determination of subunit structure of purified laccase, SDS-AGE was run where four bands were visualized on silver stainingFig. 3A). For determination of size of these bands, a calibrationraph between relative mobility and log molecular weight waslotted (Fig. 3B). Based on this calibration plot, the size of theseands was found to correspond to 52, 45, 38 and 25 kDa, suggest-
ng a hexameric structure (consisting of two subunits each of 52nd 45 while one each of 38 and 25 kDa) for the native laccase.
Most of the plant laccases reported so far was found toe monomeric having subunit molecular within the range of0–100 kDa [11,29]. Recently, a heterodimeric laccase from a tree
egume, Leucaena leucocephala consisting of two subunits, one
f 100 kDa and other of 120 kDa, have been reported [6]. Multi-eric laccases having a native molecular weight of 137 kDa eachave been reported from two xerophytic plant species, Cereusterogonus (CP137) and Opuntia vulgaris (OV137) [12]. Reports of
ing purification. (B) In-gel activity staining of purified papaya laccase. The gel afterNative-PAGE was immersed in 50 mM catechol solution until the brown-colouredband appeared.
fungal and bacterial laccases in homodimeric, heterodimeric andhomotrimeric forms have also been made in literature [30–34].
3.3. Effect of temperature on papaya laccase activity
The effect of temperature on purified laccase was investigatedand it was found that laccase activity increased up to 70 ◦C andafterwards decreased rapidly (Fig. 4A). The optimum temperaturefor laccases from plants like L. leucocephala, C. pterogonus and O.vulgaris have been reported to be 80, 90, and 70 ◦C, respectively[4–6]. Some thermostable bacterial and fungal laccases have alsobeen reported [9,35,36]. The Q10 value of papaya laccase was foundto be >1 up to 70 ◦C similar to L. leucocephala and Cerrena unicolorlaccase suggesting that the reaction rate is temperature-dependent[6,37]. The energy of activation (Ea) as determined from the slopeof the Arrhenius plot was found to be 7.7 kJ mol−1 which was alsoalmost similar to those found from L. leucocephala (6.9 kJ mol−1 [6])and C. unicolor (6.9 kJ mol−1 [37]).
Time-dependent thermostability of papaya laccase was alsoinvestigated by incubating the enzyme at temperatures 50, 60, 70and 80 ◦C for different time intervals and it was found that theenzyme lost 25, 18, 47 and 100% activity within 1 h of incubationat 50, 60 and 70 and 80 ◦C, respectively (Fig. 4B). Similar stability oflaccase from O. vulgaris (OV90) has been reported when incubatedfor 30 min at 70 ◦C [4] while laccase from Pleurotus ostreatus wasalmost completely inactivated after 15 min at 70 ◦C [38]. However,Gonzalez et al. [39] reported 3.7% residual activity of Pycnoporoussanguineus laccase after incubation at 70 ◦C for 1 h. Similar to our
result, Trametes versicolor laccase have also been reported to showcomplete inactivation of laccase after 1 h of incubation at 80 ◦C[15]. In contrast, laccase from a tree legume, L. leucocephala havebeen found to show activation upon pre-incubation at 80 ◦C for up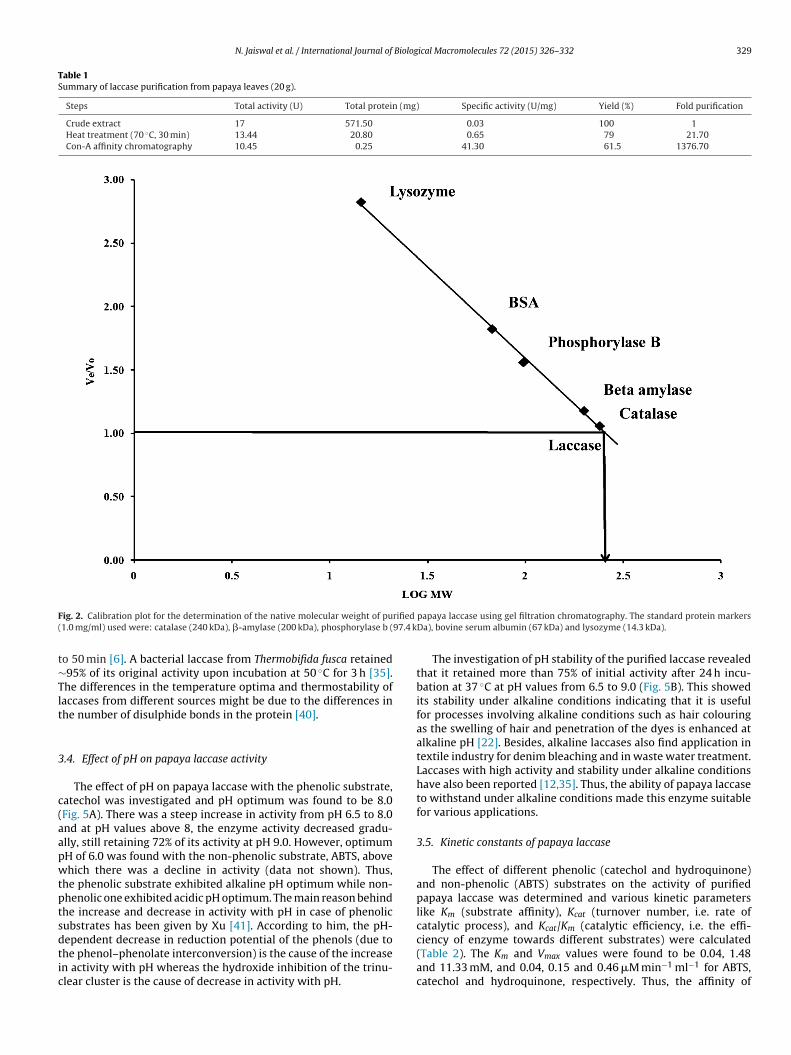
N. Jaiswal et al. / International Journal of Biological Macromolecules 72 (2015) 326–332 329
Table 1Summary of laccase purification from papaya leaves (20 g).
Steps Total activity (U) Total protein (mg) Specific activity (U/mg) Yield (%) Fold purification
Crude extract 17 571.50 0.03 100 1Heat treatment (70 ◦C, 30 min) 13.44 20.80 0.65 79 21.70Con-A affinity chromatography 10.45 0.25 41.30 61.5 1376.70
F rified
( 97.4 k
t∼Tlt
3
c(aapwtptsdtic
ig. 2. Calibration plot for the determination of the native molecular weight of pu1.0 mg/ml) used were: catalase (240 kDa), �-amylase (200 kDa), phosphorylase b (
o 50 min [6]. A bacterial laccase from Thermobifida fusca retained95% of its original activity upon incubation at 50 ◦C for 3 h [35].he differences in the temperature optima and thermostability ofaccases from different sources might be due to the differences inhe number of disulphide bonds in the protein [40].
.4. Effect of pH on papaya laccase activity
The effect of pH on papaya laccase with the phenolic substrate,atechol was investigated and pH optimum was found to be 8.0Fig. 5A). There was a steep increase in activity from pH 6.5 to 8.0nd at pH values above 8, the enzyme activity decreased gradu-lly, still retaining 72% of its activity at pH 9.0. However, optimumH of 6.0 was found with the non-phenolic substrate, ABTS, abovehich there was a decline in activity (data not shown). Thus,
he phenolic substrate exhibited alkaline pH optimum while non-henolic one exhibited acidic pH optimum. The main reason behindhe increase and decrease in activity with pH in case of phenolicubstrates has been given by Xu [41]. According to him, the pH-
ependent decrease in reduction potential of the phenols (due tohe phenol–phenolate interconversion) is the cause of the increasen activity with pH whereas the hydroxide inhibition of the trinu-lear cluster is the cause of decrease in activity with pH.papaya laccase using gel filtration chromatography. The standard protein markersDa), bovine serum albumin (67 kDa) and lysozyme (14.3 kDa).
The investigation of pH stability of the purified laccase revealedthat it retained more than 75% of initial activity after 24 h incu-bation at 37 ◦C at pH values from 6.5 to 9.0 (Fig. 5B). This showedits stability under alkaline conditions indicating that it is usefulfor processes involving alkaline conditions such as hair colouringas the swelling of hair and penetration of the dyes is enhanced atalkaline pH [22]. Besides, alkaline laccases also find application intextile industry for denim bleaching and in waste water treatment.Laccases with high activity and stability under alkaline conditionshave also been reported [12,35]. Thus, the ability of papaya laccaseto withstand under alkaline conditions made this enzyme suitablefor various applications.
3.5. Kinetic constants of papaya laccase
The effect of different phenolic (catechol and hydroquinone)and non-phenolic (ABTS) substrates on the activity of purifiedpapaya laccase was determined and various kinetic parameterslike Km (substrate affinity), Kcat (turnover number, i.e. rate ofcatalytic process), and Kcat/Km (catalytic efficiency, i.e. the effi-
ciency of enzyme towards different substrates) were calculated(Table 2). The Km and Vmax values were found to be 0.04, 1.48and 11.33 mM, and 0.04, 0.15 and 0.46 �M min−1 ml−1 for ABTS,catechol and hydroquinone, respectively. Thus, the affinity of
330 N. Jaiswal et al. / International Journal of Biological Macromolecules 72 (2015) 326–332
Fig. 3. (A) SDS-PAGE and silver staining of purified papaya laccase. Lane 1: purified laccase; lane 2: molecular weight markers. Standard protein markers containingmyosin (205 kDa), phosphorylase b (97.4 kDa), bovine serum albumin (66 kDa), ovalbumin (43 kDa), and carbonic anhydrase (29 kDa) were used. (B) Calibration plot for thedetermination of the subunit molecular weight of purified papaya laccase.
0
10
20
30
40
0 20 40 60 80 100
Act
ivity
(U/m
l) x
10-2
Temperature (oC)
0
20
40
60
80
100
120
0 20 40 60
% R
esid
ual A
ctiv
ity
Time (min)
50oC60oC70oC80oC
A B
F ncubaa . Percw
pteo
TKs
ig. 4. (A) Effect of temperature on the activity of papaya laccase. The enzyme was issay conditions. (B) Thermal stability of papaya laccase at different temperatureshich was set at 100%.
apaya laccase towards various substrates was found to be in
he order: ABTS > catechol > hydroquinone. However, Kcat of thenzyme for these substrates were found to be in just oppositerder: hydroquinone (3.07 min−1) > catechol (0.99 min−1) > ABTSable 2inetic properties of papaya laccase with various phenolic and non-phenolicubstrates.
Substrates Km (mM) Vmax
(�M min−1 ml−1)Kcat
(min−1)Kcat/Km
(mM−1 min−1)
ABTS 0.04 0.04 0.29 7.25Catechol 1.48 0.15 0.99 0.67Hydroquinone 11.33 0.46 3.07 0.27
ted at different temperatures (10–90 ◦C) and activity was measured under standardent relative activity represents the enzyme activity relative to the control (0 min),
(0.29 min−1). The catalytic efficiencies of the purified papayalaccase for various substrates (as presented by Kcat/Km) werefound to be in the order: ABTS (7.25 mM−1 min−1) > catechol(0.67 mM−1 min−1) > hydroquinone (0.27 mM−1 min−1). Thus, thenon-phenolic substrate, ABTS was found to be most efficientlyoxidized by papaya laccase which is evident from the Km aswell as the Kcat/Km values. Catechol was found to be the secondefficient substrate; however, the papaya laccase was less reac-tive towards hydroquinone. Almost similar Km values with ABTShave been reported for laccases from fungi, Ceriolopsis subver-
mispora (0.042 mM [42]), Daedela quercina (0.038 mM [43]) andPhlebia radiata (0.049 mM [44]). Lower Km values for ABTS havealso been reported with laccases from plant, Populus eurameri-cana (0.026 mM [3]), fungal, Pleurotus pulmunarius (0.21 mM [33]),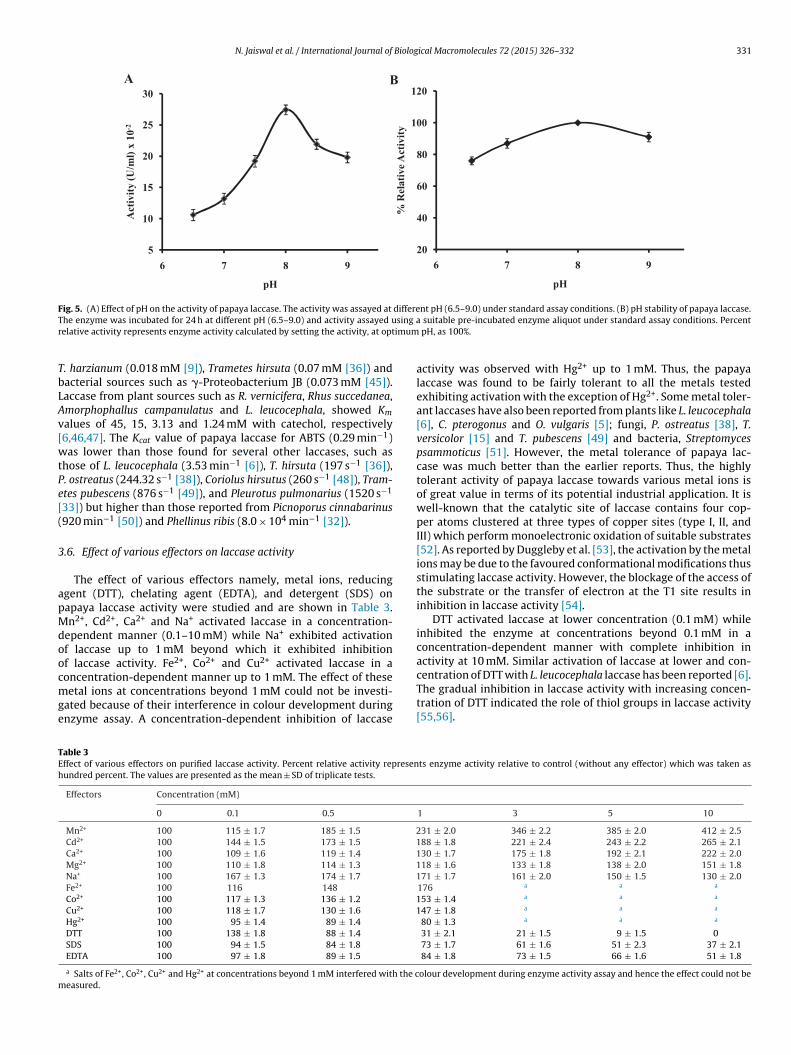
N. Jaiswal et al. / International Journal of Biological Macromolecules 72 (2015) 326–332 331
A B
5
10
15
20
25
30
6 7 8 9
Act
ivity
(U/m
l) x
10-2
pH
20
40
60
80
100
120
6 7 8 9
% R
elat
ive A
ctiv
ity
pH
Fig. 5. (A) Effect of pH on the activity of papaya laccase. The activity was assayed at different pH (6.5–9.0) under standard assay conditions. (B) pH stability of papaya laccase.T sing ar imum
TbLAv[wtPe[(
3
apMdoocmge
TEh
m
he enzyme was incubated for 24 h at different pH (6.5–9.0) and activity assayed uelative activity represents enzyme activity calculated by setting the activity, at opt
. harzianum (0.018 mM [9]), Trametes hirsuta (0.07 mM [36]) andacterial sources such as �-Proteobacterium JB (0.073 mM [45]).accase from plant sources such as R. vernicifera, Rhus succedanea,morphophallus campanulatus and L. leucocephala, showed Km
alues of 45, 15, 3.13 and 1.24 mM with catechol, respectively6,46,47]. The Kcat value of papaya laccase for ABTS (0.29 min−1)as lower than those found for several other laccases, such as
hose of L. leucocephala (3.53 min−1 [6]), T. hirsuta (197 s−1 [36]),. ostreatus (244.32 s−1 [38]), Coriolus hirsutus (260 s−1 [48]), Tram-tes pubescens (876 s−1 [49]), and Pleurotus pulmonarius (1520 s−1
33]) but higher than those reported from Picnoporus cinnabarinus920 min−1 [50]) and Phellinus ribis (8.0 × 104 min−1 [32]).
.6. Effect of various effectors on laccase activity
The effect of various effectors namely, metal ions, reducinggent (DTT), chelating agent (EDTA), and detergent (SDS) onapaya laccase activity were studied and are shown in Table 3.n2+, Cd2+, Ca2+ and Na+ activated laccase in a concentration-
ependent manner (0.1–10 mM) while Na+ exhibited activationf laccase up to 1 mM beyond which it exhibited inhibitionf laccase activity. Fe2+, Co2+ and Cu2+ activated laccase in a
oncentration-dependent manner up to 1 mM. The effect of theseetal ions at concentrations beyond 1 mM could not be investi-ated because of their interference in colour development duringnzyme assay. A concentration-dependent inhibition of laccase
able 3ffect of various effectors on purified laccase activity. Percent relative activity represenundred percent. The values are presented as the mean ± SD of triplicate tests.
Effectors Concentration (mM)
0 0.1 0.5
Mn2+ 100 115 ± 1.7 185 ± 1.5 2Cd2+ 100 144 ± 1.5 173 ± 1.5 1Ca2+ 100 109 ± 1.6 119 ± 1.4 1Mg2+ 100 110 ± 1.8 114 ± 1.3 1Na+ 100 167 ± 1.3 174 ± 1.7 1Fe2+ 100 116 148
Co2+ 100 117 ± 1.3 136 ± 1.2 1Cu2+ 100 118 ± 1.7 130 ± 1.6 1Hg2+ 100 95 ± 1.4 89 ± 1.4
DTT 100 138 ± 1.8 88 ± 1.4
SDS 100 94 ± 1.5 84 ± 1.8
EDTA 100 97 ± 1.8 89 ± 1.5
a Salts of Fe2+, Co2+, Cu2+ and Hg2+ at concentrations beyond 1 mM interfered with the ceasured.
suitable pre-incubated enzyme aliquot under standard assay conditions. Percent pH, as 100%.
activity was observed with Hg2+ up to 1 mM. Thus, the papayalaccase was found to be fairly tolerant to all the metals testedexhibiting activation with the exception of Hg2+. Some metal toler-ant laccases have also been reported from plants like L. leucocephala[6], C. pterogonus and O. vulgaris [5]; fungi, P. ostreatus [38], T.versicolor [15] and T. pubescens [49] and bacteria, Streptomycespsammoticus [51]. However, the metal tolerance of papaya lac-case was much better than the earlier reports. Thus, the highlytolerant activity of papaya laccase towards various metal ions isof great value in terms of its potential industrial application. It iswell-known that the catalytic site of laccase contains four cop-per atoms clustered at three types of copper sites (type I, II, andIII) which perform monoelectronic oxidation of suitable substrates[52]. As reported by Duggleby et al. [53], the activation by the metalions may be due to the favoured conformational modifications thusstimulating laccase activity. However, the blockage of the access ofthe substrate or the transfer of electron at the T1 site results ininhibition in laccase activity [54].
DTT activated laccase at lower concentration (0.1 mM) whileinhibited the enzyme at concentrations beyond 0.1 mM in aconcentration-dependent manner with complete inhibition inactivity at 10 mM. Similar activation of laccase at lower and con-
centration of DTT with L. leucocephala laccase has been reported [6].The gradual inhibition in laccase activity with increasing concen-tration of DTT indicated the role of thiol groups in laccase activity[55,56].ts enzyme activity relative to control (without any effector) which was taken as
1 3 5 10
31 ± 2.0 346 ± 2.2 385 ± 2.0 412 ± 2.588 ± 1.8 221 ± 2.4 243 ± 2.2 265 ± 2.130 ± 1.7 175 ± 1.8 192 ± 2.1 222 ± 2.018 ± 1.6 133 ± 1.8 138 ± 2.0 151 ± 1.871 ± 1.7 161 ± 2.0 150 ± 1.5 130 ± 2.0
176 a a a
53 ± 1.4 a a a
47 ± 1.8 a a a
80 ± 1.3 a a a
31 ± 2.1 21 ± 1.5 9 ± 1.5 073 ± 1.7 61 ± 1.6 51 ± 2.3 37 ± 2.184 ± 1.8 73 ± 1.5 66 ± 1.6 51 ± 1.8
olour development during enzyme activity assay and hence the effect could not be

3 Biolog
rmoCti
4
pwawIttt
A
DtmCoosp
R
[[
[
[
[
[
[[
[[
[
[
[
[
[[[[[[[[
[
[[[
[
[
[
[
[
[[
[[
[[[
[[[
[[
[[
32 N. Jaiswal et al. / International Journal of
SDS and EDTA also inhibited laccase activity similar to thoseeported from peach, O. vulgaris, C. pterogonus, T. versicolor, Strepto-yces cyaneus and S. psammoticus [5,6,15,51,56,57]. The inhibition
f laccase activity by EDTA might be due to the metal chelation ofu in the catalytic site of the enzyme [14]. However, the change inhe conformation of the protein structure might be the cause of thenhibition of activity by SDS.
. Conclusion
A laccase from papaya was purified to homogeneity in a two steprocess, namely heat treatment and Con-A affinity chromatographyith good enzyme yield. It was found to be a hexamer with pH
nd temperature optima of 8.0 and 70 ◦C, respectively. The enzymeas found to be fairly tolerant towards heat, pH and metal ions.
n addition, the papaya laccase showed enhancement in activity inhe presence of most of the metal ions. Such characteristics indicatehe potential suitability of papaya laccase for use in bioremediation,extile, pulp and paper industries.
cknowledgements
Financial support from University Grants Commission, Newelhi, India, in the form of Dr. D.S. Kothari Postdoctoral Fellowship
o NJ is gratefully acknowledged. Financial supports from Depart-ent of Higher Education, Govt. of Uttar Pradesh, India under
entre of Excellence in Bioinformatics, Department of Biotechnol-gy, Govt. of India under BIF Scheme, New Delhi and Departmentf Science and Technology, New Delhi under Promotion of Univer-ity Research and Scientific Excellence (DST-PURSE) programme forroviding infrastructure facilities are also gratefully acknowledged.
eferences
[1] U.N. Dwivedi, P. Singh, V.P. Pandey, A. Kumar, J. Mol. Catal. B: Enzym. 68 (2011)117–128.
[2] H. Yoshida, J. Chem. Soc. 43 (1883) 472–486.[3] P. Ranocha, G. McDougall, S. Hawkins, R. Sterjiades, G. Borderies, D. Stewartz,
M. Cabanes-Macheteau, A.-M. Boudet, D. Goffner, Eur. J. Biochem. 259 (1999)485–495.
[4] G.N. Kumar, K. Srikumar, Biomed. Chromatogr. 25 (2011) 707–711.[5] G.N. Kumar, K. Srikumar, Chem. Nat. Compd. 48 (2012) 451–456.[6] N. Jaiswal, V.P. Pandey, U.N. Dwivedi, Process Biochem. (2014),
http://dx.doi.org/10.1016/j.procbio.2014.04.002.[7] D.S. Arora, R.K. Sharma, Appl. Biochem. Biotechnol. 160 (2010) 1760–1788.[8] F. Xu, in: M.C. Flickinger, S.W. Drew (Eds.), The Encyclopedia of Bioprocessing
Technology: Fermentation, Biocatalysis and Bioseparation, Wiley, New York,1999, pp. 1545–1554.
[9] S. Sadhasivam, S. Savitha, K. Swaminathan, F.-H. Lin, Process Biochem. 43 (2008)736–742.
10] Y.X. Wong, J. Yu, Water Res. 33 (1999) 3512–3520.11] D.S. Kocabas, Z.B. Ogel, U. Bakir, World J. Microbiol. Biotechnol. 27 (2011)
701–707.12] G.N. Kumar, K. Srikumar, Appl. Biochem. Biotechnol. (2013),
http://dx.doi.org/10.1007/s12010-013-0611-6.13] K.K. Sharma, B. Shrivastava, V.R.B. Sastry, N. Sehgal, R.C. Kuhad, Sci. Rep. (2013),
http://dx.doi.org/10.1038/srep01299.
[[
[
ical Macromolecules 72 (2015) 326–332
14] S. Galai, F. Limam, M.N. Marzouki, Appl. Biochem. Biotechnol. 158 (2009)416–431.
15] Z. Youshuang, Z. Haibo, C. Mingle, W. Zhenzhen, H. Feng, G. Peiji, Biotechnol.Bioproc. Eng. 16 (2011) 1027–1035.
16] K. Hilden, T. Hakala, T. Lundell, Biotechnol. Lett. 31 (2009) 1117–1128.17] K.K.Y. Wong, J.D. Richardson, S.D. Mansfield, Biotechnol. Prog. 16 (2000)
1025–1029.18] M. Asgher, H.N. Bhatti, M. Ashraf, R.L. Legge, Biodegradation 19 (2008) 771–783.19] K. Murugesan, Y.M. Kim, J.R. Jeon, Y.S. Chang, J. Hazard. Mater. 168 (2009)
523–529.20] X. Xiao, C.C. Xu, Y.M. Wu, P.J. Cai, W.W. Li, D.L. Du, H.Q. Yu, Bioresour. Technol.
110 (2012) 86–90.21] E. Dube, F. Shareck, Y. Hurtubise, M. Beauregard, C. Daneault, J. Ind. Microbiol.
Biotechnol. 35 (2008) 11–23.22] K. Saito, R. Ikeda, K. Endo, Y. Tsujino, M. Takagi, E. Tamiya, J. Biosci. Bioeng. 113
(2012) 575–579.23] I. Matijosyte, I.W.C.E. Arends, S. de Vries, R.A. Sheldon, J. Mol. Catal. B: Enzym.
62 (2010) 142–148.24] M.M. Bradford, Anal. Biochem. 72 (1976) 248–254.25] B.J. Davis, Ann. N.Y. Acad. Sci. 121 (1964) 404–427.26] P. Andrews, Biochem. J. 91 (1964) 222–233.27] U.K. Laemmli, Nature 227 (1970) 680–685.28] B. Chefetz, Y. Chen, Y. Hadar, Appl. Environ. Microbiol. 64 (1998) 3175–3179.29] C.M. Alba, S.M. de Forchetti, A.T. Horacio, Physiol. Plant. 109 (2000) 382–387.30] S.B. Younes, S. Sayadi, J. Mol. Catal. B: Enzym. 73 (2011) 35–42.31] A.V. Lisov, A.G. Zavarzina, A.A. Zavarzin, A.A. Leontievsky, FEMS Microbiol. Lett.
275 (2007) 46–52.32] K.L. Min, Y.H. Kim, Y.W. Kim, H.S. Jung, Y.C. Hah, Arch. Biochem. Biophys. 392
(2001) 279–286.33] C.G.M. DeSouza, R.M. Peralta, J. Basic Microbiol. 43 (2003) 278–286.34] T.B. Ng, H.X. Wang, Biochem. Biophys. Res. Commun. 313 (2004) 37–41.35] C.-Y. Chen, Y.-C. Huang, C.-M. Wei, M. Meng, W.-H. Liu, C.-H. Yan, AMB Express
3 (2013) 49.36] Z. Haibo, Z. Yinglong, H. Feng, G. Peiji, C. Jiachuan, Biotechnol. Lett. 31 (2009)
837–843.37] D. D’Souza-Ticlo, D. Sharma, C. Raghukumar, Mar. Biotechnol. 11 (2009)
725–737.38] H. Patel, S. Gupte, M. Gahlout, A. Gupte, 3 Biotech (2013),
http://dx.doi.org/10.1007/s13205-013-0129-1.39] E.D. Gonzalez, O.V. Vallejo, C.M. Anaya, M.M. Sanchez, M.C. Gonzalez, L.A. Palo-
mares, J.F. Mallol, Int. Microbiol. 11 (2008) 163–169.40] F. Xu, W. Shin, S.H. Brown, J.A. Wahleithner, U.M. Sundaram, E.I. Solomon,
Biochem. Biophys. Acta 1292 (1996) 303–311.41] F. Xu, J. Biol. Chem. 272 (1997) 924–928.42] L.F. Larrondo, L. Salas, F. Melo, R. Vicuna, D. Cullen, Appl. Environ. Microbiol. 69
(2003) 6257–6263.43] P. Baldrian, FEMS Microbiol. Ecol. 50 (2004) 245–253.44] S. Kaneko, M. Cheng, S. Murai, S. Takenaka, S. Murakami, K. Aoki, Biosci. Bio-
technol. Biochem. 73 (2009) 939–942.45] M. Asgher, S. Kamal, H.M.N. Iqbal, Chem. Cent. J. 6 (2012) 110.46] T. Omura, J. Biochem. 50 (1961) 264–272.47] P.S. Paranjpe, M.S. Karve, S.B. Padhye, Indian J. Biochem. Biophys. 40 (2003)
40–45.48] K.S. Shin, Y.J. Lee, Arch. Biochem. Biophys. 384 (2000) 109–115.49] J. Si, F. Peng, B. Cui, Bioresour. Technol. 128 (2013) 49–57.50] C. Eggert, U. Temp, K.E. Eriksson, Appl. Environ. Microbiol. 62 (1996)
1151–1158.51] K.N. Niladevi, N. Jacob, P. Prema, Process Biochem. 43 (2008) 654–660.52] M. Frasconi, G. Favero, H. Boer, A. Koivula, F. Mazzei, Biochim. Biophys. Acta
1804 (2010) 899–908.53] R.G. Duggleby, J. Theor. Biol. 81 (1979) 671–684.54] Z.M. Fang, T.L. Li, F. Chang, P. Zhou, W. Fang, Y.Z. Hong, X.C. Zhang, H. Peng, Y.Z.
Xiao, Bioresour. Technol. 111 (2012) 36–41.
55] C. Johannes, A. Majcherczyk, J. Biotechnol. 78 (2000) 193–199.56] M. Lorenzo, D. Moldes, S. Rodríguez Couto, M.A. Sanromán, Chemosphere 60(2005) 1124–1128.57] M.E. Arias, M. Arenas, J. Rodriguez, J. Soliveri, A.S. Ball, M. Hernandez, Appl.
Environ. Microbiol. 69 (2003) 1953–1958.