purification and characterization of recombinant caulobacter crescentus cu,zn superoxide dismutase
TRANSCRIPT

.elsevier.com/locate/bba
Biochimica et Biophysica Acta
Purification and characterization of recombinant Caulobacter crescentus
Cu,Zn superoxide dismutase
Ivana De Domenico a, Amalia Lania a, Maria Carmela Bonaccorsi di Patti b, Andrea Battistoni c,
Giovanni Musci a,*, Alessandro Desideri c
a Department of Microbiological, Genetic and Molecular Sciences, University of Messina, Salita Sperone, 31, 98166 Messina, Italyb Department of Biochemical Sciences, University of Rome La Sapienza, Rome, Italy
c Department of Biology, University of Rome Tor Vergata, Rome, Italy
Received 13 July 2005; received in revised form 20 August 2005; accepted 24 August 2005
Available online 13 September 2005
Abstract
Recombinant Cu,Zn Superoxide Dismutase from Caulobacter crescentus has been expressed in Escherichia coli and characterized. The
corresponding recombinant protein has a molecular weight typical of a homodimeric Cu,ZnSODs and an activity comparable to that of other
prokaryotic enzymes. The copper active site is characterized by a peculiar axial geometry as evidenced by its electron paramagnetic resonance
spectrum, moreover, the copper atom displays a low accessibility toward external chelating agents indicating a lower solvent accessibility when
compared to other prokaryotic enzymes. Investigation of the enzyme thermal stability through differential scanning calorimetry indicates the
occurrence of two transitions at low and higher temperature that are found to be due to the apo and holo protein, respectively, confirming that the
metals have a crucial role in the stabilization of this class of enzymes.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Superoxide dismutase; Caulobacter crescentus; Copper
1. Introduction
Cu,Zn superoxide dismutase (Cu,ZnSOD) is a major
component of the antioxidant defense system that protects
cells from oxidative damage by converting superoxide radicals
into hydrogen peroxide and oxygen [1]. For a long time,
Cu,ZnSOD has been considered almost exclusively an eukary-
otic enzyme, but it is now firmly established that Cu,ZnSOD is
present in a large number of bacteria [2]. While in eukaryotes,
Cu,ZnSODs are mainly found in the cytoplasm, in bacteria,
they are exported to the periplasm or to the outer membrane.
Such extracytoplasmic location of the enzyme suggests that its
role is to protect bacteria from superoxide generated outside the
bacterium or within the periplasmic space [2].
1570-9639/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.bbapap.2005.08.021
Abbreviations: SOD, superoxide dismutase; DTT, dithiothreitol; EDTA,
ethylenediaminetetraacetic acid; EPR, Electron Paramagnetic Resonance; DSC,
Differential Scanning Calorimetry
* Corresponding author. Tel.: +390906765194; fax: +39090392733.
E-mail address: [email protected] (G. Musci).
Bacterial and eukaryotic Cu,ZnSODs share a similar
monomer fold and a conserved arrangement of the redox
center, where, in the oxidized form of the enzyme, Cu2+ is
coordinated by four histidine residues and Zn2+ is coordinated
by three histidine and one aspartate residues [3]. Despite these
similarities, which indicate that they likely derive from a
common ancestor gene, prokaryotic and eukaryotic Cu,Zn-
SODs show remarkable differences in the organization of the
active site channel and in the way subunits are assembled [2–
4]. Moreover, several prokaryotic Cu,ZnSODs show unique
sequence features, which are suggestive of significant structural
differences not only with respect to eukaryotic Cu,ZnSODs,
but also with respect to the already characterized prokaryotic
enzymes. Examples of species-specific variations include the
presence of insertions and/or deletions in the loops connecting
the elements forming the beta-barrel [3,4], mutations in
residues involved in the metal coordination [5,6] and the
presence of additional protein domains [7]. Such differences
between bacterial Cu,ZnSODs are expected to significantly
influence the structural and functional properties of individual
enzyme variants. Recent studies have also revealed that
1764 (2006) 105 – 109
http://www

I. De Domenico et al. / Biochimica et Biophysica Acta 1764 (2006) 105–109106
bacterial Cu,ZnSOD activity and stability can be finely
modulated even by small differences in the quaternary structure
of different enzymes [8–10].
An example of bacterial Cu,ZnSOD exhibiting an unusual
primary sequence is the enzyme from Caulobacter crescentus,
a free living bacterium originally cultured from fresh water
ponds [11]. This enzyme, which has been shown to be active
and able to bind either copper and zinc [11], is characterized by
a specific pattern of insertion and deletions that could modify
either the structure of the active site or the modality of
interactions between subunits, and by the substitution of a
conserved zinc ligand [12]. These features make C. crescentus
Cu,ZnSOD a very interesting candidate for structure/function
investigations on this class of enzymes. Here, we describe the
purification and a preliminary characterization of the enzyme
produced in E. coli by recombinant DNA techniques.
2. Materials and methods
C. crescentus CB15 ATCC strain 19089 was obtained from American Type
Culture Collection. E. coli strain DH5a was a kind gift of Dr. C. Falcone
(University of Rome La Sapienza). C. crescentus was grown in nutrient broth
(peptone 1 g/l, yeast extract 0.2 g/l and MgSO4 0.2 g/l). The cells were
collected by centrifugation and genomic DNA extraction was performed
according to Chen and Kuo [13]. Oligonucleotides complementary to the amino
and the carboxyl terminals of the C. crescentus Cu,ZnSOD sequence were
synthesized and used to PCR amplify the coding region of the gene. The
amplification product was cloned into the vector pUC18 using EcoRI and XbaI.
The recombinant plasmid was transformed into E. coli DH5a. DNA sequence
analysis at Biogen-ENEA facilities was performed to verify the identity of the
cloned sequence with that of the C. crescentus Cu,ZnSOD gene present in the
GeneBank.
Recombinant clones were grown in LB medium containing 50 mg/ml
ampicillin and 1 mM CuSO4 for 18 h at 37 -C. The periplasmic fraction was
isolated as described previously [14] and dialyzed against 50 mM potassium
phosphate buffer pH 6.5 to remove sucrose. Proteins contained in the
periplasmic fraction were concentrated and subjected to (NH4)2SO4 fraction-
ation at 4 -C. At variance with what reported on the non-recombinant native
Fig. 1. Alignment of C. caulobacter Cu,ZnSOD amino acid sequence to that of o
pleuropneumoniae and E. coli. The global alignment has been deduced from optim
described [4]. The h-strand elements are indicated above amino acid sequences with
shown in bold and are indicated by C and Z, respectively. Residues involved in the d
of A. pleuropneumoniae Cu,ZnSOD, encoding a His-rich metal binding domain [7
protein expressed by C. crescentus, which was found to precipitate already at
50% (NH4)2SO4 [11], all the recombinant enzyme expressed in E. coli has
been found in the supernatant of 85% (NH4)2SO4 precipitation. The
supernatant containing Cu,ZnSOD was dialyzed against several changes of
50 mM potassium phosphate buffer pH 7.4 for 18 h, followed by 20 mM
potassium phosphate buffer pH 7.4 for 24 h. At this stage, the enzyme was
more than 90% pure and was used for subsequent studies without further
purification. Protein concentration was quantified with the bicinchoninic acid
(BCA) protein assay kit (Pierce, Rockford IL). The final yield of recombinant
Cu,ZnSOD was 8 mg of protein per liter of bacterial culture. Copper content,
evaluated by double integration of the EPR spectrum vs. a Cu–EDTA
standard complex, indicated the presence of only 1 Cu/dimer. This result was
confirmed by atomic absorption spectroscopy, carried out with a Perkin Elmer
3030 instrument equipped with a graphite furnace, that indicated the presence
of 0.9 Cu atom/protein and of 1.75 Zn atom/protein, in line with a previous
report [11].
SOD activity was visualized in non-denaturing polyacrilamide gels as an
achromatic zone that did not stain upon photochemical reduction of nitroblue
tetrazolium to formazan blue [15]. SOD activity was estimated by the inhibition
of pyrogallol autoxidation, as described previously [16]. The stability of
superoxide dismutase activity in the presence of EDTA was studied following
described procedures [10,17].
Low temperature EPR spectra were recorded on a Bruker ESP 300
spectrometer operating at 9 GHz with 100-kHz field modulation. The EPR
spectra were manipulated by using the EPR package provided by Stelar (Mede,
Italy) and simulated by using software kindly provided by Dr. Bencini
(Florence, Italy). DSC experiments were performed with a MicroCal VP-DSC
microcalorimeter (MicroCal Inc., Northampton, MA). Proteins were extensive-
ly dialyzed against 100 mM potassium phosphate buffer pH 7.8 and gently
degassed before scanning. The protein concentration was 0.2–0.3 mg/ml, and
the scan speed was 60 -C/h. No reversibility of thermal unfolding was found at
the end of the scan, therefore, the second scan was used as baseline to correct
the thermograms. Heat capacity versus temperature profiles for the thermally
induced transition were analyzed, using the Origin version 5.0 software
provided by MicroCal, to obtain values for Tm (temperature of maximum heat
capacity) and DH (heat of reaction).
Apo-Cu,Zn SOD was obtained by extensive dialysis of the enzyme (15–30
mg/ml) in 50 mM sodium acetate buffer pH 3.8 containing 1 mM EDTA for
three days at room temperature. The enzyme was then dialyzed overnight at 4
-C in the same buffer containing 100 mM NaCl to remove excess EDTA, and
finally against 50 mM acetate buffer pH 5.5 at 4 -C for 12 h. Copper removal
was monitored spectrophotometrically at 680 nm. Distilled water and glassware
ther enzymes from Gram-negative bacteria: P. leiognathi, S. typhimurium, A.
al superimposition of available bacterial Cu,ZnSOD structures, as previously
the6 symbol. The histidine residues coordinating the active site metal ions are
imer interface of P. leiognathi SOD [18] are underlined. The N-terminal region
] has been omitted.
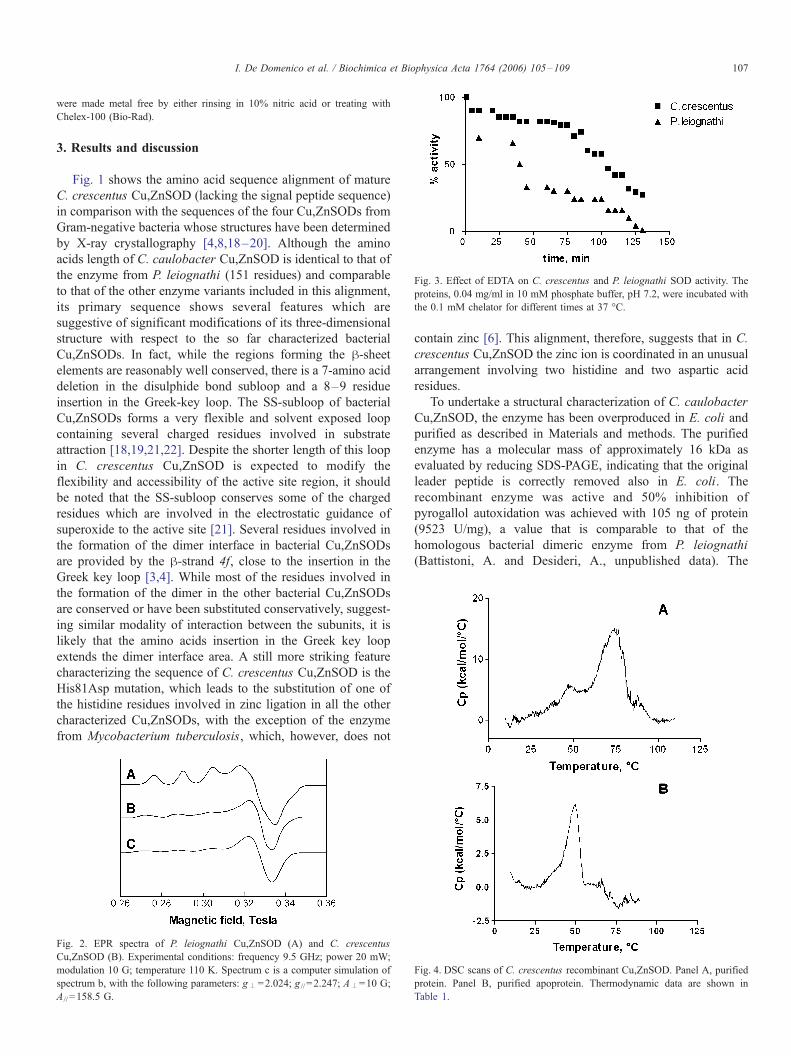
Fig. 3. Effect of EDTA on C. crescentus and P. leiognathi SOD activity. The
proteins, 0.04 mg/ml in 10 mM phosphate buffer, pH 7.2, were incubated with
the 0.1 mM chelator for different times at 37 -C.
I. De Domenico et al. / Biochimica et Biophysica Acta 1764 (2006) 105–109 107
were made metal free by either rinsing in 10% nitric acid or treating with
Chelex-100 (Bio-Rad).
3. Results and discussion
Fig. 1 shows the amino acid sequence alignment of mature
C. crescentus Cu,ZnSOD (lacking the signal peptide sequence)
in comparison with the sequences of the four Cu,ZnSODs from
Gram-negative bacteria whose structures have been determined
by X-ray crystallography [4,8,18–20]. Although the amino
acids length of C. caulobacter Cu,ZnSOD is identical to that of
the enzyme from P. leiognathi (151 residues) and comparable
to that of the other enzyme variants included in this alignment,
its primary sequence shows several features which are
suggestive of significant modifications of its three-dimensional
structure with respect to the so far characterized bacterial
Cu,ZnSODs. In fact, while the regions forming the h-sheetelements are reasonably well conserved, there is a 7-amino acid
deletion in the disulphide bond subloop and a 8–9 residue
insertion in the Greek-key loop. The SS-subloop of bacterial
Cu,ZnSODs forms a very flexible and solvent exposed loop
containing several charged residues involved in substrate
attraction [18,19,21,22]. Despite the shorter length of this loop
in C. crescentus Cu,ZnSOD is expected to modify the
flexibility and accessibility of the active site region, it should
be noted that the SS-subloop conserves some of the charged
residues which are involved in the electrostatic guidance of
superoxide to the active site [21]. Several residues involved in
the formation of the dimer interface in bacterial Cu,ZnSODs
are provided by the h-strand 4f, close to the insertion in the
Greek key loop [3,4]. While most of the residues involved in
the formation of the dimer in the other bacterial Cu,ZnSODs
are conserved or have been substituted conservatively, suggest-
ing similar modality of interaction between the subunits, it is
likely that the amino acids insertion in the Greek key loop
extends the dimer interface area. A still more striking feature
characterizing the sequence of C. crescentus Cu,ZnSOD is the
His81Asp mutation, which leads to the substitution of one of
the histidine residues involved in zinc ligation in all the other
characterized Cu,ZnSODs, with the exception of the enzyme
from Mycobacterium tuberculosis, which, however, does not
Fig. 2. EPR spectra of P. leiognathi Cu,ZnSOD (A) and C. crescentus
Cu,ZnSOD (B). Experimental conditions: frequency 9.5 GHz; power 20 mW;
modulation 10 G; temperature 110 K. Spectrum c is a computer simulation of
spectrum b, with the following parameters: g–=2.024; g // =2.247; A–=10 G;
A// =158.5 G.
contain zinc [6]. This alignment, therefore, suggests that in C.
crescentus Cu,ZnSOD the zinc ion is coordinated in an unusual
arrangement involving two histidine and two aspartic acid
residues.
To undertake a structural characterization of C. caulobacter
Cu,ZnSOD, the enzyme has been overproduced in E. coli and
purified as described in Materials and methods. The purified
enzyme has a molecular mass of approximately 16 kDa as
evaluated by reducing SDS-PAGE, indicating that the original
leader peptide is correctly removed also in E. coli. The
recombinant enzyme was active and 50% inhibition of
pyrogallol autoxidation was achieved with 105 ng of protein
(9523 U/mg), a value that is comparable to that of the
homologous bacterial dimeric enzyme from P. leiognathi
(Battistoni, A. and Desideri, A., unpublished data). The
Fig. 4. DSC scans of C. crescentus recombinant Cu,ZnSOD. Panel A, purified
protein. Panel B, purified apoprotein. Thermodynamic data are shown in
Table 1.

Table 1
Thermodynamic parameters for recombinant C. crescentus holo- and apo
Cu,ZnSOD
Enzyme Tm(-C) DH (kcal/mol
Holo 75.8 132
Apo 50.0 67.8
I. De Domenico et al. / Biochimica et Biophysica Acta 1764 (2006) 105–109108
molecular mass of the native enzyme is 32 kDa when examined
by non-reducing SDS-PAGE, confirming that the enzyme has a
dimeric structure consisting of two identical subunits (data not
shown).
Fig. 2 shows the EPR spectrum of recombinant C.
crescentus compared to that of P. leiognathi Cu,ZnSOD [23].
The line shape, typical of a copper atom in the oxidized state, is
different in the two proteins, the spectrum of recombinant C.
crescentus Cu,Zn SOD showing an axial shape, with values of
the hyperfine splitting and of the g parameter in the parallel
region (as calculated from the simulated spectrum c) higher and
lower than that of P. leiognathi respectively (A// =158.5 G vs.
137.2 G and g// =2.247 vs. 2.267 for C. crescentus and P.
leiognathi, respectively). These parameters indicate that copper
has significantly different coordination environments in C.
crescentus vs. P. leiognathi Cu,ZnSOD, despite the fact that
the copper ligands are identical in the two proteins as evaluated
by sequence alignment. Such an effect can then be due to the
difference in the ligand coordination of the proximal zinc atom,
that in C. crescentus has a histidine in place of the conserved
aspartate, and/or to three dimensional constraints that cannot be
envisaged without the solution of the 3D structure of the
protein.
The accessibility of the copper site of Cu,ZnSOD has been
evaluated in a comparative way for C. crescentus and P.
leiognathi Cu,ZnSOD measuring the dismutase activity as a
function of time, upon incubation of the protein with 0.1 mM
EDTA in 10 mM phosphate buffer, pH 7.2. As shown in Fig. 3,
EDTA induces a loss of the enzymatic activity that can be
related to the active site copper chelation. The loss of activity is
faster in P. leiognathi than in C. crescentus enzyme, being,
after 1 h of incubation, more than 75% for P. leiognathi and
only 20% for C. crescentus. Both SODs were totally
inactivated after longer incubation times in presence of EDTA.
Previous studies have shown that the EDTA-mediated inacti-
vation is faster for the monomeric Cu,ZnSOD from E. coli than
for most dimeric bacterial enzymes of this class [24] and that
mutations at the dimer interface of P. leiognathi Cu,ZnSOD
strongly enhance the rate of copper loss in presence of EDTA
[9,10]. Such studies have suggested that differences in the rates
of inactivation by EDTA are indicative of modifications at the
dimer interface between individual bacterial Cu,ZnSODs,
which affect the conformational flexibility of the active site
region [8,10,24]. On this ground, it is possible to suggest that
C. crescentus Cu,ZnSOD is characterized by a more stable
interaction between the subunits compared to P. leiognathi
Cu,ZnSOD. Stabilization of the subunit interaction in C.
crescentus Cu,ZnSOD is likely favored by an extension of
the subunit interface area due to the insertion in the Greek key
loop.
The thermal stability of C. crescentus Cu,Zn SOD has been
assessed by differential scanning calorimetry. The DSC profile,
shown in Fig. 4A, reveals the presence of two distinct peaks
that can be analyzed in terms of two independent transitions
with melting temperature around 50 -C and 75 -C, respectively.The presence of two peaks in the DSC scan can be explained
either as the occurrence of two different denaturing steps or by
-
)
the presence of two distinct species with different stability. The
DSC scan carried out on a solution containing more than 95%
of apo Cu,ZnSOD (as estimated by atomic absorption
spectrometry) is also reported in Fig. 4B. The DSC profile of
this sample is characterized by an intense peak with melting
temperature of 50 -C. This result clearly indicates that the
peaks at low and high temperature are diagnostic for the
melting of the apo and holo form, respectively. Table 1 reports
the calculated thermodynamic parameters. The Tm of the apo
enzyme is identical to that of the apo-enzyme from E. coli and
Bos taurus [17], whereas the Tm of the metallated enzyme is
5 -C higher than that reported for the dimeric enzyme from P.
leiognathi [18]. These observations confirm that both the
metallation state and the dimeric structure contribute to the
stabilization of Cu,ZnSODs. It is likely that an extended
dimerization area in C. crescentus Cu,ZnSOD could be
responsible of the higher Tm of this enzyme compared to P.
leiognathi Cu,ZnSOD. Interestingly, as most of the contribu-
tion of metals to Cu,ZnSOD stabilization should be ascribed to
zinc [25], the high melting temperatures of both the holo
enzymes from C. crescentus and P. leiognathi suggests that the
differences in the zinc coordination environment in the two
enzymes have little influence on the stability of the zinc site.
In conclusion, the data here reported indicate that the
Cu,ZnSOD from C. crescentus displays specific characteristics
when compared to other bacterial enzymes of this class, such as
an active site with a copper atom characterized by an almost
axial geometry, a low accessibility to external chelating agents
and a high thermal stability. Although only the solution of its
3D structure will permit to formulate clear correlations between
the enzyme structure and its physicochemical properties, it is
likely that most of the specific properties of C. crescentus
Cu,ZnSOD are due to its peculiar active site metal coordination
and to an altered modality of interaction between the subunits.
Acknowledgements
We thank Dr. Anna Giartosio (Universita’ La Sapienza-
Roma) for helping with calorimetric experiments. This work
was partially supported by a Murst Cofin 2004 Project.
References
[1] I. Fridovich, Superoxide radical and superoxide dismutases, Annu. Rev.
Biochem. 64 (1995) 97–112.
[2] A. Battistoni, Role of prokaryotic Cu,Zn superoxide dismutase in
pathogenesis, Biochem. Soc. Trans. 31 (2003) 1326–1329.
[3] D. Bordo, A. Pesce, M. Bolognesi, M.E. Stroppolo, M. Falconi, A.
Desideri, Cu,Zn superoxide dismutase in prokaryotes and eukaryotes, in:
A. Messerschmidt, R. Huber, T. Poulos, K. Wiegardt (Eds.), Handbook of
Metalloproteins, John Wiley and Sons, Chichester, 2001, pp. 1284–1300.

I. De Domenico et al. / Biochimica et Biophysica Acta 1764 (2006) 105–109 109
[4] D. Bordo, D. Matak, K. Djinovic-Carugo, C. Rosano, A. Pesce, M.
Bolognesi, M.E. Stroppolo, M. Falconi, A. Battistoni, A. Desideri,
Evolutionary constraints for dimer formation in prokaryotic Cu,Zn
superoxide dismutase, J. Mol. Biol. 285 (1999) 283–296.
[5] P.R. Langford, B.M. Loynds, J.S. Kroll, Copper-zinc superoxide dis-
mutase in Haemophilus species, J. Gen. Microbiol. 138 (1992) 517–522.
[6] L. Spagnolo, I. Toro, M. D’Orazio, P. O’Neill, J.Z. Pedersen, O. Carugo,
G. Rotilio, A. Battistoni, K. Djinovic-Carugo, Unique features of the
sodC-encoded superoxide dismutase from Mycobacterium tuberculosis, a
fully functional copper-containing enzyme lacking zinc in the active site,
J. Biol. Chem. 279 (2004) 33447–33455.
[7] A. Battistoni, F. Pacello, A.P. Mazzetti, C. Capo, J.S. Kroll, P.R. Langford,
A. Sansone, G. Donnarumma, P. Valenti, G. Rotilio, A histidine-rich metal
binding domain at the N terminus of Cu,Zn-superoxide dismutases from
pathogenic bacteria: a novel strategy for metal chaperoning, J. Biol.
Chem. 276 (2001) 30315–30325.
[8] A. Pesce, A. Battistoni, M.E. Stroppolo, F. Polizio, M. Nardini, J.S. Kroll,
P.R. Langford, P. O’Neill, M. Sette, A. Desideri, M. Bolognesi, Functional
and crystallographic characterization of Salmonella typhimurium Cu,Zn
superoxide dismutase coded by the sodCI virulence gene, J. Mol. Biol.
302 (2000) 465–478.
[9] M.E. Stroppolo, A. Pesce, M. Falconi, P. O’Neill, M. Bolognesi, A.
Desideri, Single mutation at the intersubunit interface confers extra
efficiency to Cu,Zn superoxide dismutase, FEBS Lett. 483 (2000) 17–20.
[10] M.E. Stroppolo, A. Pesce, M. D’Orazio, P. O’Neill, D. Bordo, C.
Rosano, M. Milani, A. Battistoni, M. Bolognesi, A. Desideri, Single
mutations at the subunit interface modulate copper reactivity in
Photobacterium leiognathi Cu,Zn superoxide dismutase, J. Mol. Biol.
308 (2001) 555–563.
[11] H.M. Steinman, Copper-zinc superoxide dismutase from Caulobacter
crescentus CB15. A novel bacteriocuprein form of the enzyme, J. Biol.
Chem. 257 (1982) 10283–10293.
[12] H.M. Steinman, B. Ely, Copper-zinc superoxide dismutase of Caulobacter
crescentus: cloning, sequencing, and mapping of the gene and periplasmic
location of the enzyme, J. Bacteriol. 172 (1990) 2901–2910.
[13] W.P. Chen, T.T. Kuo, A simple and rapid method for the preparation of
gram-negative bacterial genomic DNA, Nucleic Acids Res. 21 (1993)
2260.
[14] L. Britton, I. Fridovich, Intracellular localization of the superoxide
dismutases of Escherichia coli: a reevaluation, J. Bacteriol. 131 (1977)
815–820.
[15] C. Beauchamp, I. Fridovich, Superoxide dismutase: improved assays
and an assay applicable to acrylamide gels, Anal. Biochem. 44 (1971)
276–287.
[16] S. Marklund, G. Marklund, Involvement of the superoxide anion radical in
the autoxidation of pyrogallol and a convenient assay for superoxide
dismutase, Eur. J. Biochem. 47 (1974) 469–474.
[17] A. Battistoni, S. Folcarelli, L. Cervoni, F. Polizio, A. Desideri, A.
Giartosio, G. Rotilio, Role of the dimeric structure in Cu,Zn superoxide
dismutase. pH-dependent, reversible denaturation of the monomeric
enzyme from Escherichia coli, J. Biol. Chem. 273 (1998) 5655–5661.
[18] Y. Bourne, S.M. Redford, H.M. Steinman, J.R. Lepock, J.A. Tainer, E.D.
Getzoff, Novel dimeric interface and electrostatic recognition in bacterial
Cu,Zn superoxide dismutase, Proc. Natl. Acad. Sci. U. S. A. 93 (1996)
12774–12779.
[19] A. Pesce, C. Capasso, A. Battistoni, S. Folcarelli, G. Rotilio, A. Desideri,
M. Bolognesi, Unique structural features of the monomeric Cu,Zn
superoxide dismutase from Escherichia coli, revealed by X-ray crystal-
lography, J. Mol. Biol. 274 (1997) 408–420.
[20] K.T. Forest, P.R. Langford, J.S. Kroll, E.D. Getzoff, Cu,Zn superoxide
dismutase structure from a microbial pathogen establishes a class with a
conserved dimer interface, J. Mol. Biol. 296 (2000) 145–153.
[21] S. Folcarelli, A. Battistoni, M. Falconi, P. O’Neill, G. Rotilio, A. Desideri,
Conserved enzyme–substrate electrostatic attraction in prokaryotic Cu,Zn
superoxide dismutases, Biochem. Biophys. Res. Commun. 244 (1998)
908–911.
[22] M. Falconi, L. Parrilli, A. Battistoni, A. Desideri, Flexibility in monomeric
Cu,Zn superoxide dismutase detected by limited proteolysis and molecular
dynamics simulation, Proteins 47 (2002) 513–520.
[23] D. Foti, C.B. Lo, G. Cuzzocrea, M.E. Stroppolo, F. Polizio, M. Venanzi,
A. Desideri, Spectroscopic characterization of recombinant Cu,Zn
superoxide dismutase from Photobacterium leiognathi expressed in
Escherichia coli: evidence for a novel catalytic copper binding site,
Biochemistry 36 (1997) 7109–7113.
[24] R. Gabbianelli, M. D’Orazio, F. Pacello, P. O’Neill, L. Nicolini, G.
Rotilio, A. Battistoni, Distinctive functional features in prokaryotic and
eukaryotic Cu,Zn superoxide dismutases, Biol. Chem. 385 (2004)
749–754.
[25] J.A. Roe, A. Butler, D.M. Scholler, J.S. Valentine, L. Marky, K.J.
Breslauer, Differential scanning calorimetry of Cu,Zn-superoxide dismu-
tase, the apoprotein, and its zinc-substituted derivatives, Biochemistry 27
(1988) 950–958.