purification and characterization of extracellular ... · resultsobtainedfromstarch hydrolysis...
TRANSCRIPT

Vol. 50, No. 6APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 1985, p. 1474-14820099-2240/85/121474-09$02.00/0Copyright © 1985, American Society for Microbiology
Purification and Characterization of Extracellular AmylolyticEnzymes from the Yeast Filobasidium capsuligenum
RENE DE MOT AND HUBERT VERACHTERT*
Laboratory of Industrial Microbiology and Biochemistry, University of Leiven, B-3030 Leuven, Belgium
Received 25 March 1985/Accepted 25 August 1985
The extracellular amylolytic system of Filobasidium capsuligenum consisted of an a-amylase (1,4-a-D-glucanglucanhydrolase, EC 3.2.1.1) and two forms of glucoamylase (1,4-a-D-glucan glucohydrolase, EC 3.2.1.3). Theenzymes were purified by ammonium sulfate fractionation, repeated ion-exchange chromatography (DEAE-Sephadex A-50), and gel filtration (Sephadex G-25, Sephadex G-100 sf). cL-Amylase had an optimum pH of 5.6and an optimum temperature of 50°C but was rapidly inactivated at higher temperature. The molecular weightwas estimated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis to be 64,000. An acarboseconcentration of 20 ,ug/ml was required for 50% inhibition of the a-amylase. Both glucoamylases are
glycoproteins of identical molecular weight (60,000) and produce only glucose by exohydrolysis. Thedebranching activity of the glucoamylases was evidenced with substrates containing at-1,6 linkages. The pHoptima were 5.0 to 5.6 for glucoamylase I and 4.8 to 5.3 for glucoamylase II. Glucoamylase I had a higheroptimum temperature (55°C) than glucoamylase II (50°C) and was also more resistant to thermal inactivation.Only low acarbose concentrations (<0.1 ,ug/ml) were required to reduce the activity of the glucoamylases by50%.
Interest in amylolytic yeasts has increased in recent yearsas their potential value for conversion of starchy biomass tosingle-cell protein or ethanol has been recognized (12, 15, 33,38, 42). The extracellular amylolytic enzymes produced byspecies of Lipomyces (31, 32, 34), Saccharomvcopsis (10,13, 36), and Schwanniomyces (24, 29, 30, 41) are wellcharacterized.A comparative study of a large number of amylolytic
yeasts, conducted in our laboratory, indicated that thesespecies differ strongly with regard to amylase secretion andthe extent of starch hydrolysis (5, 6). Strains of Filobasidiuimcapsuligenum are capable of extensive starch hydrolysis (7,20) and secrete high levels of amylolytic activity in a growthmedium containing corn starch and corn steep liquor (7). Ourresults obtained from starch hydrolysis patterns with a crudeenzyme preparation of F. capsuligentum suggested the pres-ence of both glucoamylase and o-amylase activity (6). How-ever, Ebertova (9) reported the presence of only an ox-amylase in culture fluids of Candida jaiponica. The basidialstate of C. japonica was placed in the new genusLeucosporidium as L. capsuligenum by Fell et al. (11) andreclassified as F. capsuligenum by Rodrigues de Miranda(26). Therefore, the amylolytic system of F. capsuligenumwas investigated in more detail, and in this paper we reporton the purification and characterization of the extracellularamylases from this yeast species.
MATERIALS AND METHODSYeast strains. F. capsuligeniim CBS 4381 was obtained
from the Centraalbureau voor Schimmelcultures, Delft, TheNetherlands. F. capsuligenum CCY 64-2-4, CCY 64-5-1, andCCY 71-1-1 were from the Czechoslovak Collection ofYeasts, Institute of Chemistry, Slovak Academy of Sciences,Bratislava, Czechoslovakia.Media. Cultures were grown at 30°C on slants containing
soluble starch (5) and maintained at 4°C. Amylase produc-
* Corresponding author.
tion medium contained 0.5% corn starch (Union ChemiqueBelge, Brussels, Belgium) and 3% corn steep liquor (SigmaChemical Co., St. Louis, Mo.). The pH of the medium wasadjusted to 6.5 with 1 M NaOH before sterilization. In themedium used for determination of the time course of starchhydrolysis and amylase production, 0.5% soluble starch (E.Merck AG, Darmstadt, Federal Republic of Germany) wassubstituted for corn starch.Enzyme assays. Total amylolytic activity was measured by
the 3,5-dinitrosalicylate (DNS) method as previously de-scribed (5), except that an incubation temperature of 55°Cwas used. At this temperature the highest value for totalamylolytic activity was obtained for the crude enzymepreparation.Glucoamylase was assayed by determining the amount of
glucose liberated from Zulkowsky soluble starch (Merck).The substrate solution consisted of 2% soluble starch inMcllvaine buffer (pH 5.4) (21a). Substrate solution (1 ml) andenzyme solution (0.2 ml) were incubated at 55°C for 30 min.The reaction was stopped by heating the mixture in a boilingwater bath for 5 min. Glucose in the cooled sample wasdetermined by the peroxidase-glucose oxidase method withthe peroxidase-glucose oxidase enzyme mixture (Sigma) and2,2'-azino-di-(3-ethylbenzthiazoline)-6-sulphonate (Boehr-inger GmbH, Mannheim, Federal Republic of Germany) as achromogen (2). One glucoamylase unit is defined as theamount of enzyme producing 1 ,umol of glucose per minunder the conditions specified.
OL-Amylase was determined qualitatively during enzymepurification by a diffusion method, using a solidified mediumcontaining the Phadebas amylase substrate (Pharmacia, Up-psala, Sweden) as previously described (5). The method ofMcCleary (21) with remazol-brilliant blue carboxymethylamylose (Alphachrome; Koch-Light Laboratories,Colnbrook, United Kingdom) as a specific ot-amylase sub-strate was used for all quantitative determinations of theenzyme. Mcllvaine buffer (pH 5.4) was used as substratebuffer. One unit of oL-amylase is defined as the amount ofenzyme liberating an amount of colored, soluble material
1474
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

AMYLASES FROM FILOBASIDIUM CAPSULIGENUM 1475
corresponding to 10 absorbance units at 590 nm from 10 mgof Alphachrome during incubation for 10 min at 40°C and pH5.4.Time course of starch degradation and amylase secretion.
The time course of amylase secretion and of starch degra-dation during shake-flask culture of F. capsuligenum wasdetermined by periodical sampling of the growth medium.Growth was measured by the A650 of a cell suspension,diluted to A650 < 0.4 with sterile medium as a blank (25).Samples for the assay of glucose, starch, glucoamylase, anda-amylase were centrifuged at 10,000 x g for 10 min at 2°C.Part of the supernatant was immediately heated in boilingwater for 5 min to inactivate the amylolytic enzymes andused for the assay of starch and glucose. Starch was deter-mined by the iodine method (5).
Purification of enzymes. F. capsuligenum CCY 64-5-1 wasgrown at 29°C in two 1-liter Erlenmeyer flasks, each con-taining 200 ml of amylase production medium, on a recipro-cal shaker for 48 h. The cultures were then centrifuged at10,000 x g for 15 min at 2°C, and the supernatant (400 ml)containing the extracellular enzymes was collected.
Solid ammonium sulfate was added to the supernatant to40% saturation at 0°C. This solution was stirred for 1 h at 0°Cand then centrifuged for 30 min at 15,000 x g (0°C). Thepellet was discarded, and ammonium sulfate was added tothe supernatant to 80% saturation. The precipitate from thistreatment was collected by centrifugation and dissolved in aminimal volume of 25 mM sodium acetate buffer (pH 6). Theenzyme solution was dialyzed against the same buffer for 12h at 4°C.The concentrated enzyme solution was then applied to a
column (35 by 2.5 cm) of Sephadex G-25 (medium grade;Pharmacia) and eluted at 150C with 50 mM sodium acetatebuffer (pH 6) containing 0.002% chlorhexidine (Sigma). At aflow rate of 15 ml h-', 6.3-ml fractions were collected. Theamylase-positive fractions (DNS method) were combinedand concentrated to 15 ml by ultrafiltration at room temper-ature through a collodion bag (Sartorius GmbH, Gottingen,Federal Republic of Germany).A column (55 by 2.6 cm) packed with DEAE-Sephadex
A-50 ion-exchange resin (Pharmacia) was equilibrated at15°C in 50 mM sodium acetate buffer (pH 6) containing0.002% chlorhexidine. The 15-ml sample from the previousgel filtration step was applied, and elution was carried out ata flow rate of 4.8 ml h-', using an NaCl gradient from 0 to0.25 M developed in 1 liter of elution buffer. Collectedfractions were 4.7 ml. Protein peaks were monitored by UVabsorbance at 280 nm. Three amylase-positive peak frac-tions (100 to 109, 110 to 125, and 134 to 160) were separatelypooled, dialyzed against 50 mM sodium acetate buffer (pH 6)for 6 h at 4°C, and then concentrated to 8, 10, and 7 ml,respectively, by collodion ultrafiltration.The dialyzed samples from the first and second amylase
peaks were applied separately to columns (40 by 2.6 cm) ofDEAE-Sephadex A-50, equilibrated at 15°C with 50 mMsodium acetate buffer (pH 6) containing 0.002%chlorhexidine. In both cases, elution was carried out at aflow rate of 3.2 ml h-1, using an NaCl gradient from 0 to 0.15M NaCl developed in 1 liter of elution buffer. Fractions of4.7 ml were collected. The fractions containing the majoramylase peak from each column were pooled and used as thepurified enzyme preparations for the characterization stud-ies.A column (90 by 1.6 cm) packed with Sephadex G-100 sf
(Pharmacia) was used for further purification of the thirdamylase peak obtained after the first ion-exchange chroma-
tography step. Elution was carried out at 15°C with 50 mMsodium acetate buffer (pH 6) containing 0.002%chlorhexidine. A flow rate of 3.2 ml h-1 was used. Theelution of protein was monitored by UV absorbance. Frac-tions of 3.1 ml were collected and assayed for amylaseactivity by the DNS method. The fractions with the majoramylolytic acitivity (30 to 39) were pooled and used forfurther characterization of the purified enzyme.
Polyacrylamide gel electrophoresis. Electrophoresis undernondenaturing conditions was done in 6.5% polyacrylamidegel rods in buffer (pH 8.5) containing 25 mM Tris and 0.19 Mglycine. Samples (40 ,ul) were applied and run for 2 h at 5 mAper gel rod.
Incubation with 12.5% trichloroacetic acid solution for 45min was used for protein fixation before staining for 4 h with0.05% Coomassie brilliant blue R-250 in a mixture of aceticacid-methanol-water (1:5:5, vol/vol/vol). The gels were thendestained in an aqueous mixture of acetic acid (7.5%) andmethanol (5%).
Amylolytic activity was localized after electrophoresis byimmersing the gels for 15 min in a solution of 2% solublestarch (Merck) in Mcllvaine buffer (pH 5.4) at room temper-ature. The gels were then kept for 10 min in the same bufferwithout soluble starch at 45°C. After this the gels werestained with iodine solution (0.3% 12 in 3% KI) for 5 min.With this procedure some shrinkage of the gels occurred.Enzyme activity was visible as a pale yellow band in thedark-colored gels.The periodic acid-Schiff stain was used for detection of
glycoproteins in the gels. After electrophoresis the gels werekept for 1 h in 7.5% acetic acid at room temperature and thenimmersed in cold (4°C) 0.2% periodic acid solution for 45min. After this the gels were contacted for 45 min with Schiffreagent (Merck) at 4°C and then destained at room temper-ature in several changes of 10% acetic acid. Glycoproteinsproduced pink-colored bands.
Determination of molecular weight. The molecular weightsof the purified enzymes were determined by polyacrylamidegel electrophoresis in the presence of sodium dodecyl sulfate(SDS) (40). Samples (30 ,ul) were run for 6 h in 7.5%polyacrylamide gel rods containing 0.1% SDS, using 0.1 Msodium phosphate buffer (pH 7.2) with 0.1% SDS as electro-phoresis buffer. The enzyme samples were first mixed withan equal volume of sample buffer, heated at 100°C for 5 min,and then cooled. The sample buffer consisted of 1% SDS and1% 2-mercaptoethanol in 0.1 M sodium phosphate (pH 7.2).The calibration proteins (High Molecular Weight CalibrationKit; Pharmacia) were dissolved in sample buffer and kept at60°C for 15 min before electrophoresis. The gel rods werestained for protein as described above.Chromatography of hydrolysis products. The purified
enyzmes were incubated with 2% soluble starch (Merck) or2% pullulan (Sigma) at 40°C. Samples were taken after 0.5, 2,and 4 h for soluble starch and after 4 and 12 h for pullulan.Hydrolysis was stopped by heating the samples at 100°C for5 min.
Hydrolysis products were analyzed by double-ascentchromatogrpahy of 2-,ul samples on precoated Silica Gel 60plates (Merck), using a 7:2:1 (vol/vol/vol) isopropanol-aceticacid-water solvent system. A 5% solution of Maldex 15(Amylum, Aalst, Belgium) and a mixture of 2% glucose and2% maltose were used as standards. The chromatogram wasdeveloped with aniline-diphenylamine reagent, which wasprepared by mixing 20 ml of 85% phosphoric acid with asolution of 4 ml of aniline and 4 g of diphenylamine in 200 mlof acetone.
VOL. 50, 1985
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

1476 DE MOT AND VERACHTERT
15
04-,
0
ca)Vc0V
a)vn0u
aV)
I
10
8-
loF
I
5F
OL
61
41
2
0
.-.u
0
-o
1.5
:D
>1I 4-,
0.5 :
V0
w
U)0
0~
10
9
E
4i'>1
>6 U
o
a)vn0
E0030Da)
Eo0
time: daysFIG. 1. Time course of starch degradation and secretion of amylases by F. capsuligenum CCY 64-5-1 growing at 29°C in a medium
containing 3% corn steep liquor and 0.5% soluble starch (pH 6.5).
Thermal inactivation. Thermal inactivation of enzymeswas studied for temperature values between 40 and 70°Cafter exposure to each temperature for 5 to 30 min. Enzymesamples in Mcllvaine buffer (pH 5.6) were then quicklycooled on ice and assayed for residual activity by the DNSmethod. The results were compared with the activity presentbefore heat treatment.
Total protein. Protein was determined by the method ofLowry et al. (16), using bovine serum albumin to prepare thestandard curve.
RESULTSEnzyme production. Preliminary experiments using vari-
ous concentrations of corn starch (0.5 to 2.5%) and cornsteep liquor (1 to 9%), and different initial pH values (3.5 to8.5) had shown that amylase production by F. capsuligenumwas highest in a medium containing 0.5% corn starch and 3%corn steep liquor with an initial pH of 6.5. Addition of salts[0.5% (NH4)2SO4, 0.1% KH2PO4, 0.05% MgSO4, 0.01%NaCl, and 0.01% CaC12] did not improve amylase secretion.Using the simple corn starch-corn steep liquor medium, theproduction of extracellular a-amylase and glucoamylase wascompared for four F. capsuligenum strains. The supernatantobtained after centrifugation of the cultures at 10,000 x g for15 min at 2°C was used for the enzyme assays. Strain CBS4381 secreted three to four times as much a-amylase (2.3U/ml) as the other strains tested but showed the lowestglucoamylase activity (2.9 U/ml). Strain CCY 64-5-1, whichproduced the highest level of glucoamylase activity (7.2U/ml) together with good ot-amylase activity (0.8 U/ml), wasused for characterization of the enzymes.A typical pattern for the time course of starch degradation
and amylase production (Fig. 1) shows that starch hydrolysisproceeded quickly during the early exponential phase withaccumulation of glucose in the medium. The low levels ofextracellular ot-amylase and glucoamylase probably reflectthe involvement of cell-bound activity in early starch hydro-lysis. The glucose concentration decreased rapidly duringthe exponential growth phase, and the pH was lowered toabout 5.5. Increasing levels of the extracellular amylolyticenyzmes were detected after the exponential phase and gaverise to additional glucose, which was liberated from smallerdextrins not detectable with the iodine reagent. Maximumvalues for both a-amylase and glucoamylase were reachedafter the exponential growth phase (24 to 48 h). While the pHslowly increased to about 8, both a-amylase and glucoam-ylase activity decreased upon further incubation (72 h).Enzyme purification. The culture supernatant was concen-
trated (40-fold) and partially purified (2-fold) with little lossof activity by ammonium sulfate precipitation of theamylolytic enzymes in the range of 40 to 80% saturation at0°C (Table 1). Low-molecular-weight components originat-ing from the corn steep liquor were partially separated fromthe amylolytic enzymes by Sephadex G-25 gel filtration. Thisbrown-colored fraction with strong UV absorbance (280 nm)was removed to avoid interference with UV monitoring ofprotein in column effluents from ion-exchange chromatogra-phy.The elution pattern of the sample after the first DEAE-
Sephadex A-50 chromatography is shown in Fig. 2. Twomajor and one minor peak with amylolytic activity wereseparated. When assayed for a-amylase activity by thePhadebas method, only the fractions from the third peak (134to 160) gave a positive response. The first (fractions 100 to
APPL. ENVIRON. MICROBIOL.
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

AMYLASES FROM FILOBASIDIUM CAPSULIGENUM 1477
TABLE 1. Purification of the extracellular a-amylase, glucoamylase I, and glucoamylase I1 from F. capsuligenum CCY 64-5-1Vola Activity Protein Sp act (U/mg of Total activity Yield Purification
Purification step Enzyme (ml) (U/ml) (mg/ml) protein) (U) (%) (fold)Culture supernatant a-Amylase 400 0.8 1.03 0.8 320.0 100 1
Glucoamylaseb 400 6.7 1.03 6.5 2,688.0 100 1
Ammonium sulfate a-Amylase 10 31.6 20.60 1.5 316.0 98.8 2.0fractionation Glucoamylaseb 10 260.9 20.60 12.7 2,608.8 97.1 1.9
Sephadex G-25 gel a-Amylase 38 7.4 4.45 1.7 279.3 87.3 2.1filtration Glucoamylaseb 38 63.8 4.45 14.3 2,425.5 90.2 2.2
First DEAE-Sephadex a-Amylase 127 1.0 0.02 56.7 129.5 40.5 72.7A-50 chromatography Glucoamylase I 47 4.3 0.06 73.6 200.7 7.4 11.3
Glucoamylase II 75 3.4 0.05 63.3 256.5 9.5 9.7
Sephadex G-100 sf gel a-Amylase 29 4.2 0.03 168.8 122.4 38.3 216.4filtration
Second DEAE-Sephadex Glucoamylase I 80 2.3 0.03 85.9 185.6 6.9 13.2A-50 chromatography
Third DEAE-Sephadex Glucoamylase II 118 2.0 0.02 100.0 236.0 8.7 15.3A-50 chromatographya All volumes were corrected for removal of samples at each step to carry out enzyme assays and protein determinations.b Represents combined activity of glucoamylase I and glucoamylase II.
109) and second (fractions 110 to 125) peaks were devoid ofa-amylase activity and were further purified by DEAE-Sephadex A-50 chromatography with a less-steep gradient (0to 0.15 M NaCI). These enzymes were characterized asglucoamylases (see below) and will be further referred to asglucoamylase I (first peak) and glucoamylase II (secondpeak).The a-amylase was further purified by Sephadex G-100 gel
filtration (Fig. 3). The major activity peak (fractions 30 to 39)contained all a-amylase activity (Phadebas positive) and wasseparated from a minor peak with amylolytic activity (frac-
60 120froction number
tions 40 to 48). The latter was devoid of a-amylase(Phadebas negative) and probably constitutes a small frac-tion of glucoamylase.
Purified samples of a-amylase, glucoamylase I, and gluco-amylase II produced single bands after polyacrylamide gelelectrophoresis when stained for protein and amylolyticactivity (Fig. 4). Beside these three amylases, no otheramylolytic enzymes were detectable in the crude enzymepreparation of F. capsuligenum CCY 64-5-1. With crudeenzyme preparations from the three other F. capsuligenumstrains, similar enzyme patterns were obtained (data not
300 -0.5
.. o0
200 > ._0
-. C.> 0.3c".o a
0U
0
1~~> -0.2S0 0
z0 8 - 0.1
. i~~~~~~. O l_O
FIG. 2. Elution pattern of protein (UV absorbance) and amylolytic activity (DNS method) after thechromatography.
first DEAE-Spehadex A-S0
VOL. SO, 1985
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

1478 DE MOT AND VERACHTERT
0oCN
0
CcaL)u0u
c
2Ca
.t:_U
tiu0
o8
fraction numberFIG. 3. Purification of oa-amylase from ion-exchange chromatography by Sephadex G-100 sf gel filtration. Fractions were assayed for
amylase activity (DNS method) and protein (UV absorbance).
shown). The anodic electrophoretic mobility of the enzymesis consistent with their affinity for the anion-exchange resinDEAE-Sephadex A-50.A summary of results from the purification procedure is
given in Table 1. A 216-fold purification of cx-amylase was
achieved with a yield of 38.3% of the original activity.Purification of glucoamylases I and II was 13- and 15-fold,respectively. Yields of 6.9% for glucoamylase I and 8.8% forglucoamylase II were obtained. The purification and yieldvalues of both glucoamylases are underestimates; both en-
CE CE CE GI GI GlI Gil A A
*(-)
1 2 3 4 5 6 7 8 9FIG. 4. Polyacrylamide gel electrophoresis of the crude concen-
trated enzyme preparation (CE) and of purified glucoamylase I (GI),glucoamylase II (GII), and cx-amylase (A). Gels were stained forprotein (lanes 1, 4, 6, 8), glycoprotein (lane 3), or amylolytic activity(lanes 2, 5, 7, 9).
zymes were present in the crude enzyme preparation andcould not be separately assayed. The specific activity of thepurified glucoamylase II (100 U/mg of protein) was higherthan the value obtained for glucoamylase I (86 U/mg).Enzyme characterization. The glycoprotein nature of both
glucoamylases was demonstrated by staining of the enzymeswith the periodic acid-Schiff reagent after gel electrophore-sis. ox-Amylase was not stained under the conditions used(Fig. 4). The molecular weights, estimated by SDS-polyacrylamide gel electrophoresis, were 64,000 for cx-amylase and 60,000 for both glucoamylases.
Hydrolysis products from soluble starch and pullulan wereseparated by thin-layer chromatography with glucose,maltose, and Maldex 15 as standards (Fig. 5). Maldex 15 is acommercial cx-amylase dextrin containing glucose (minoramount), maltose, and several oligosaccharides. Bothglucoamylases released only glucose from soluble starch andpullulan. However, glucose production from pullulan wasrelatively slow. oL-Amylase did not degrade pullulan. Ini-tially, higher oligosaccharides were obtained from the actionof ox-amylase on soluble starch. As cx-amylase hydrolysisproceeded, lower oligosaccharides, including maltose, wereproduced. Glucose was not detected, but may be producedupon prolonged incubation.The activity of the enzymes at several pH values between
pH 3.2 and 7.2 was determined (data not shown). Amylolyticactivity was assayed by the DNS method with Mcllvainecitrate-phosphate as substrate buffer. The pH optima were5.6 for cx-amylase, 5.0 to 5.6 for glucoamylase I, and 4.8 to5.3 for glucoamylase II. Both glucoamylases were still activeat pH 3.5, whereas these acidic conditions completely inhib-ited oL-amylase. However, at pH 7 the reverse effect wasobtained. Glucoamylase action was negligible while ot-amylase was still active. The effect of pH on the stability ofthe enzymes at 35°C was determined by incubation inMcllvaine citrate-phosphate buffer for 24 h. Residual activ-ity was measured by the DNS method and compared with
APPL. ENVIRON. MICROBIOL.
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

AMYLASES FROM FILOBASIDIUM CAPSULIGENUM 1479
FIG. 5. Thin-layer chromatography of hydrolysis products froma-amylase (A), glucoamylase I (GI), and glucoamylase II (GII),using soluble starch (S) and pullulan (P) as substrates. The corre-sponding hydrolysis times (hours) are indicated at the top of eachlane. Maldex 15 (MX) and a mixture of glucose and maltose (GM)were used as standards. Samples were applied at the origin (0). Thedirection of solvent migration is indicated by the arrow.
the initial activity (data not shown). The three enzymes weremost stable at their optimum pH. Both glucoamylases weremore stable than a-amylase at low pH values (<4).
Activity at different temperatures was determined by theDNS method (data not shown). The optimum values were50°C for oa-amylase and glucoamylase II and 550C for gluco-amylase I. The three enzymes differed with regard to thermalstability (Fig. 6). Glucoamylase I was the most stable en-zyme, retaining about 20% of the initial activity after 5 min at60°C. A similar heat treatment completely inactivated a-
100F080k4C
4-*
4-,
0
-o
a)
50
20
10
amylase and glucoamylase II. Glucoamylase II was mostsubject to thermal inactivation with a 20% loss of activityafter 30 min at 40°C. These conditions did not affect a-amylase and glucoamylase I.The substrate specificity of the purified enzymes was
studied with oligosaccharides (10 mM), artificial substrates(10 mM), and polysaccharides (1%). Activities were assayedby the DNS method. Zulkowsky soluble starch, used in thestandard DNS procedure, was replaced by the respectivesubstrates. Activities on the oligomeric substrates, exceptmaltose, maltotriose, and cyclodextrins, were determinedfor the glucoamylases only, as data from the literatureindicate them to be unsuitable substrates for a-amylases.Relative activities were calculated as multiples of the respec-tive values for maltose (relative activity = 1) and are listed inTable 2.
High-molecular-weight substrates, namely unmodifiedstarch (corn, wheat), soluble starch and dextrin (Maldex 15)preparations, amylose, and amylopectin were the bettersubstrates for ct-amylase. Glycogen was hydrolyzed at alower rate. The a-amylase also hydrolyzed cyclodextrins;3-cyclodextrin was a much better substrate than oa-
cyclodextrin. No reducing sugars were released frompullulan by ox-amylase. Maltose and maltotriose hydrolysisproceeded slowly compared with that of the polysaccharidesubstrates.The glucoamylases differed with regard to substrate spec-
ificity. For both enzymes the highest activities were mea-sured with substrates of higher molecular weight, includingglycogen. Pullulan was degraded slowly by both enzymes,but glucoamylase II had the highest relative activity onpullulan. Maltotriose was a better substrate than maltose,especially for glucoamylase I. a-Glucosides and cyclo-dextrins were not hydrolyzed, except for p-nitrophenyl-a-D-glucoside, which was split at a reasonable rate by glucoam-ylase II. Both enzymes could attack oligomers containing
0 15 30 0 15 30 0 15 30
time: min.FIG. 6. Thermal inactivation of a-amylase (A), glucoamylase I (B), and glucoamylase 11 (C).
0.5 2 4 4 12 0.5 2 4 4 12 0.5 2 4 4 12
@0@ a@0 00
t
GMMXS S SP P S S S P P S S S P PA- -j G ' Gl
VOL. S0, 1985
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from
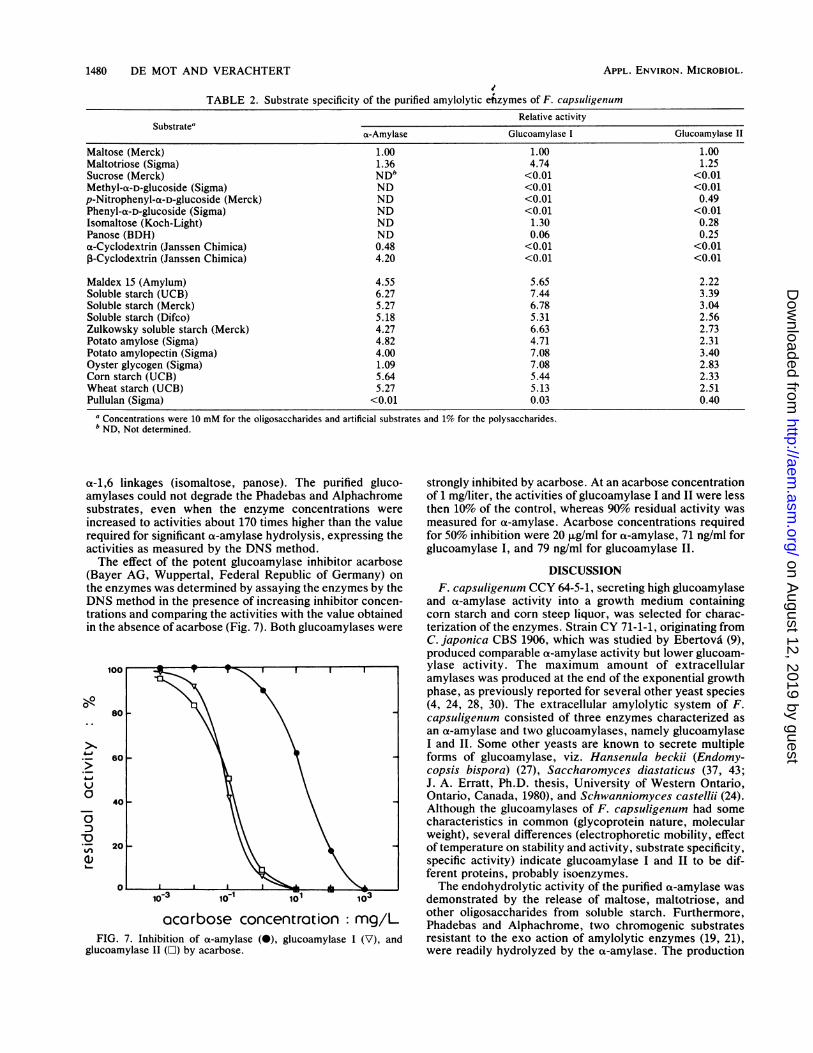
TABLE 2. Substrate specificity of the purified amylolytic enzymes of F. capsuligenumRelative activity
Substrate"cs-Amylase Glucoamylase I Glucoamylase II
Maltose (Merck) 1.00 1.00 1.00Maltotriose (Sigma) 1.36 4.74 1.25Sucrose (Merck) ND" <0.01 <0.01Methyl-a-D-glucoside (Sigma) ND <0.01 <0.01p-Nitrophenyl-a-D-glucoside (Merck) ND <0.01 0.49Phenyl-a-D-glucoside (Sigma) ND <0.01 <0.01Isomaltose (Koch-Light) ND 1.30 0.28Panose (BDH) ND 0.06 0.25a-Cyclodextrin (Janssen Chimica) 0.48 <0.01 <0.01,B-Cyclodextrin (Janssen Chimica) 4.20 <0.01 <0.01
Maldex 15 (Amylum) 4.55 5.65 2.22Soluble starch (UCB) 6.27 7.44 3.39Soluble starch (Merck) 5.27 6.78 3.04Soluble starch (Difco) 5.18 5.31 2.56Zulkowsky soluble starch (Merck) 4.27 6.63 2.73Potato amylose (Sigma) 4.82 4.71 2.31Potato amylopectin (Sigma) 4.00 7.08 3.40Oyster glycogen (Sigma) 1.09 7.08 2.83Corn starch (UCB) 5.64 5.44 2.33Wheat starch (UCB) 5.27 5.13 2.51Pullulan (Sigma) <0.01 0.03 0.40
a Concentrations were 10 mM for the oligosaccharides and artificial substrates and 1% for the polysaccharides.b ND, Not determined.
a-1,6 linkages (isomaltose, panose). The purified gluco-amylases could not degrade the Phadebas and Alphachromesubstrates, even when the enzyme concentrations wereincreased to activities about 170 times higher than the valuerequired for significant ot-amylase hydrolysis, expressing theactivities as measured by the DNS method.The effect of the potent glucoamylase inhibitor acarbose
(Bayer AG, Wuppertal, Federal Republic of Germany) onthe enzymes was determined by assaying the enzymes by theDNS method in the presence of increasing inhibitor concen-trations and comparing the activities with the value obtainedin the absence of acarbose (Fig. 7). Both glucoamylases were
0
-Q
Q1)
ocarbose concentrotion: mg/LFIG. 7. Inhibition of a-amylase (0), glucoamylase I (V), and
glucoamylase II (O) by acarbose.
strongly inhibited by acarbose. At an acarbose concentrationof 1 mg/liter, the activities of glucoamylase I and II were lessthen 10% of the control, whereas 90% residual activity wasmeasured for ox-amylase. Acarbose concentrations requiredfor 50% inhibition were 20 ,ug/ml for ot-amylase, 71 ng/ml forglucoamylase I, and 79 ng/ml for glucoamylase II.
DISCUSSIONF. capsuligenum CCY 64-5-1, secreting high glucoamylase
and ot-amylase activity into a growth medium containingcorn starch and corn steep liquor, was selected for charac-terization of the enzymes. Strain CY 71-1-1, originating fromC. japonica CBS 1906, which was studied by Ebertova (9),produced comparable ot-amylase activity but lower glucoam-ylase activity. The maximum amount of extracellularamylases was produced at the end of the exponential growthphase, as previously reported for several other yeast species(4, 24, 28, 30). The extracellular amylolytic system of F.capsuligenum consisted of three enzymes characterized asan ot-amylase and two glucoamylases, namely glucoamylaseI and II. Some other yeasts are known to secrete multipleforms of glucoamylase, viz. Hansenula beckii (Endomy-copsis bispora) (27), Saccharomyces diastaticus (37, 43;J. A. Erratt, Ph.D. thesis, University of Western Ontario,Ontario, Canada, 1980), and Schwanniomyces castellii (24).Although the glucoamylases of F. capsuligenum had somecharacteristics in common (glycoprotein nature, molecularweight), several differences (electrophoretic mobility, effectof temperature on stability and activity, substrate specificity,specific activity) indicate glucoamylase I and II to be dif-ferent proteins, probably isoenzymes.The endohydrolytic activity of the purified ot-amylase was
demonstrated by the release of maltose, maltotriose, andother oligosaccharides from soluble starch. Furthermore,Phadebas and Alphachrome, two chromogenic substratesresistant to the exo action of amylolytic enzymes (19, 21),were readily hydrolyzed by the ot-amylase. The production
1480 DE MOT AND VERACHTERT APPL. ENVIRON. MICROBIOL.
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

AMYLASES FROM FILOBASIDIUM CAPSULIGENUM 1481
of an endo-acting amylase by F. capsuligenum was previ-ously demonstrated by liquefaction of a growth mediumcontaining insoluble cross-linked amylose (1). As expected,the rate of hydrolysis decreased with increasing degree ofpolymerization of the a-D-glucans. The purified a-amylasecould hydrolyze cyclodextrins. A cyclodextrinase activityhas also been demonstrated for a-amylases from Aspergillusoryzae (35), Bacillus subtilis (22, 23), and Schizophyllumcommune (17). Pullulan was resistant to the action of F.capsuligenum a-amylase, as in previous results obtainedwith purified a-amylases from Schwanniomyces species (29,41). An acarbose concentration of 20 ,ug/ml was required for50% inhibition of the a-amylase. This is close to the valuereported for the bacterial liquefying ac-amylase (41 ,ug/ml) butlower than that for the A. oryzae a-amylase (180 ,ug/ml) (39).Both glucoamylases released only glucose from soluble
starch, indicating an exohydrolytic mechanism. This was
confirmed by the use of chromogenic substrates. Phadebasand Alphachrome were not susceptible to hydrolysis by theglucoamylases. The debranching activity of theglucoamylases was demonstrated with several substratescontaining a-1,6 branch points, namely pullulan, isomaltose,and panose, thus confirming our previous results obtainedwith the crude culture supernatant of F. capsuligenum (8).This activity is a feature of several fungal glucoamylases (18)and has also been demonstrated for purified glucoamylasesof the yeasts Candida tsukubaensis (8a), Saccharomycopsisfibuligera (36), Schwanniomyces alluvius (41), and Schwan-niomyces castellii (29). The lower affinity of the enzymes forlow-molecular-weight substrates and a-glucosides than forhighly polymerized a-D-glucans is consistent with the prop-erties of a true glucoamylase rather than an a-glucosidase(14). The glucoamylases of F. capsuligenum were stronglyinhibited by acarbose. Only 71 and 79 ng/ml were requiredfor 50% inhibition of glucoamylase I and glucoamylase II,respectively. Comparable values have been obtained for theglucoamylase of Aspergillus niger (40 ng/ml [39]) and of C.tsukubaensis (25 ng/ml [8a]). According to Clarke and Svens-son (3), two specific tryptophanyl residues are involved inthe strong but reversible binding of the pseudotetrasac-charide acarbose to the glucoamylases ofA. niger. Similar tomost fungal glucoamylases (18), the F. capsuligenumglucoamylases are glycoproteins. a-Amylase, however, was
not detected with the periodic acid-Schiff reagent for glyco-protein staining. Other yeast amylases with a carbohydratemoiety include the glucoamylases from Saccharomycesdiastaticus (37, 43; J. A. Erratt, Ph. D. thesis) and thea-amylase of Schwanniomyces castellii (29).The three amylases of F. capsuligenum had comparable
molecular weights, namely 64,000 for a-amylase and 60,000for both glucoamylases. Molecular weights of a-amylasesand glucoamylases from most amylolytic yeast species differconsiderably, ranging from 38,000 for the Lipomyceskononenkoae a-amylase (34) to 155,000 for the Schwan-niomyces alluvius glucoamylase (41). The pH and tempera-ture optima for a-amylase (5.6; 50°C), glucoamylase I (5 to5.6; 55°C), and glucoamylase 11 (4.8 to 5.3; 50°C) of F.capsuligenum are within the range of values reported formost other yeast amylases (10, 24, 27, 29, 30, 34, 36, 41).a-Amylase and especially glucoamylase II were less heatstable than glucoamylase I. The a-amylases of Schwan-niomyces species have been reported to be more susceptibleto thermal inactivation than the respective glucoamylases(24, 29, 30, 41).
F. capsuligenum produces the necessary enzymes forextensive starch degradation, namely a-amylase and gluco-
amylase with debranching activity. Therefore, the efficiencyof conversion of starchy biomass into single-cell protein bythis species should be compared with that of knownamylolytic yeasts. However, F. capsuligenum is not suitablefor direct alcoholic fermentation of starchy biomass. Glu-cose is fermented slowly and only partially, and the yeasthas a low ethanol tolerance (6a).
ACKNOWLEDGMENTS
J. Augustfn and M. Kockovd-Kratochvflovd (Slovak Academy ofSciences, Czechoslovakia) are acknowledged for generously supply-ing yeast strains. We thank M. Gruwe (Amylum, Belgium) for kindlyproviding samples of acarbose and Maldex 15.
LITERATURE CITED
1. Augustin, J., J. Zemek. A. Kockova-Kratochvilova, and E.Kuniak. 1978. Production of ax-amylase by yeasts and yeast-likeorganisms. Folia Microbiol. 23:353-361.
2. Bergmeyer, H. U., and E. Bernt. 1974. D-Glucose determinationwith glucose oxidase and peroxidase, p. 1205-1215. In H. U.Bergmeyer (ed.), Methods of enzymatic analysis, 2nd ed.Verlag Chemie, Weinheim.
3. Clarke, A. J., and B. Svensson. 1984. The role of tryptophanylresidues in the function of Aspergillus niger glucoamylase Gland G2. Carlsberg Res. Commun. 49:111-122.
4. Clementi, F., J. Rossi, L. Costamagna, and J. Rosi. 1980.Production of amylase(s) by Schwanniomyces castellii andEndomycopsisfibuligera. Antonie van Leeuwenhoek J. Micro-biol. Serol. 46:399-405.
5. De Mot, R., K. Andries, and H. Verachtert. 1984. Comparativestudy of starch degradation and amylase production byascomycetous yeast species. Syst. Appl. Microbiol. 5:106-118.
6. De Mot, R., M. Demeersman, and H. Verachtert. 1984. Compar-ative study of starch degradation and amylase production bynon-ascomycetous yeast species. Syst. Appl. Microbiol.5:421-432.
6a.De Mot, R., K. Van Dijck, A. Donkers, and H. Verachtert. 1985.Potentialities and limitations of direct alcoholic fermentation ofstarchy material with amylolytic yeasts. Appl. Microbiol.Biotechnol. 22:222-226.
7. De Mot, R., E. Van Oudendijck, S. Hougaerts, and H.Verachtert. 1984. Effect of medium composition on amylaseproduction by some starch-degrading yeasts. FEMS Microbiol.Lett. 25:169-173.
8. De Mot, R., E. Van Oudendijck, and H. Verachtert. 1984.Production of extracellular debranching activity by amylolyticyeasts. Biotechnol. Lett. 6:581-586.
8a.De Mot, R., E Van Oudendijck, and H. Verachtert. 1985.Purification and characterization of an extracellular glucoam-ylase from the yeast Candida tsukybaensis CBS 6389. Antonievan Leeuwenhoek J. Microbiol. Serol. 51:275-287.
9. Ebertova, H. 1963. Study of the formation and properties of theamylolytic system of Candida japonica. Folia Microbiol.8:333-341.
10. Ebertov., H. 1966. Amylolytic enzymes of Endomycopsiscapsularis. II. A study of the properties of isolated oa-amylase,amyloglucosidase and maltase-transglucosidase. Folia Micro-biol. 11:422-438.
11. Fell, J. W., A. C. Statzell, I. L. Hunter, and H. J. Phaff. 1969.Leucosporidium gen. n., the heterobasidiomycetous state ofseveral yeasts of the genus Candida. Antonie van LeeuwenhoekJ. Microbiol. Serol. 35:433-462.
12. Frelot, D., G. Moulin, and P. Galzy. 1982. Strain selection forthe purpose of alcohol production from starch substrates.Biotechnol. Lett. 4:705-708.
13. Kato, K., K. Kuswanto, I. Banno, and T. Harada. 1976. Identi-fication of Endomycopsis fibuligera isolated from Ragi in Indo-nesia and properties of its crystalline glucoamylase. J. Ferment.Technol. 54:831-837.
14. Kelly, C. T., and W. M. Fogarty. 1983. Microbial a-
VOL. 50, 1985
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

1482 DE MOT AND VERACHTERT
glucosidases. Process Biochem. 18(3):6-12.15. Laluce, C., and J. R. Mattoon. 1984. Development of rapidly
fermenting strains of Sacchar-omyces diastaticus for directconversion of starch and dextrins to ethanol. Appl. Environ.Microbiol. 48:17-25.
16. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin phenol reagent. J.Biol. Chem. 193:265-275.
17. Manachini, P. L. 1982. Extracellular amylolytic system of thebasidiomycete Schizophyllum commune. Ann. Microbiol.Enzimol. 32:23-26.
18. Manjunath, P., B. C. Shenoy, and M. R. Raghavendra Rao.1983. Fungal glucoamylases. J. Appl. Biochem. 5:235-260.
19. Marciniak, G. P., and M.-R. Kula. 1982. Vergleichende Unter-suchung der Methoden zur Bestimmung der Aktivitat bakteriel-ler alpha-Amylasen. Starch/Starke 34:422-430.
20. McCann, A. K., and J. A. Barnett. 1984. Starch utilization byyeasts: mutants resistant of carbon catabolite repression. Curr.Genet. 8:525-530.
21. McCleary, B. V. 1980. New chromogenic substrates for theassay of ot-amylase and (1-*4)-13-D-glucanase. Carbohydr. Res.86:97-104.
21a.McKenzie, H. A., and R. M. C. Dawson. 1969. pH, buffers, andphysiological media, p. 475-508. In R. M. C. Dawson, D. C.Elliott, W. H. Elliott, and K. M. Jones (ed.), Data for biochem-ical reserach, 2nd ed. Oxford University Press, London.
22. Moseley, M. H., and L. Keay. 1970. Purification and character-ization of the amylase of B. subtilis NRRL B3411. Biotechnol.Bioeng. 12:251-271.
23. Ohnishi, M. 1971. Studies on the interaction of substrate ana-logues with bacterial liquefying ax-amylase by means of spectro-photometry and steady state kinetics. J. Biochem. 69:181-189.
24. Oteng-Gyang, K., G. Moulin, and P. Galzy. 1981. A study of theamylolytic system of Schwanniomyces castellii. Z. Allg.Mikrobiol. 21:537-544.
25. Pringle, J. R., and J. R. Mor. 1975. Methods for monitoring thegrowth of yeast cultures and for dealing with the clumpingproblem. Methods Cell Biol. 11:131-168.
26. Rodrigues de Miranda, L. 1972. Filobasidium c apsuligenumnov. comb. Antonie van Leeuwenhoek J. Microbiol. Serol.38:91-99.
27. Ruttloff, H., R. Friese, G. Kupke, and A. Taufel. 1969.Differenzierung und Characterisierung von Glucoamylase-Isoenzymen aus Endomycopsis bispora. Z. Allg. Mikrobiol.9:39-47.
28. Ruttloff, H., A. Taufel, R. Friese, and F. Zickler. 1970. ZurAusscheidung von Glucoamylase-Isoenzymen durch einen
Stamm der Gattung Endomyc opsis. Z. Allg. Mikrobiol.10:335-340.
29. Sills, A. M., M. E. Sauder, and G. G. Stewart. 1984. Isolationand characterization of the amylolytic system of Schvan-niomyces castellii. J. Inst. Brew. 90:311-314.
30. Simoes-Mendes, B. 1984. Purification and characterization of theextracellular amylases of the yeast Schwianniomyces alluvius.Can. J. Microbiol. 30:1163-1170.
31. Spencer-Martins, I. 1982. Extracellular isoamylase produced bythe yeast Lipomyces kononenkoae. Appl. Environ. Microbiol.44:1253-1257.
32. Spencer-Martins, I. 1984. Production and properties of an ex-tracellular cyclodextrin hydrolase from the yeast Lipomyceskononenkoae. Int. J. Microbiol. 2:31-38.
33. Spencer-Martins, I., and N. van Uden. 1977. Yields of yeastgrowth on starch. Eur. J. Appl. Microbiol. 4:29-35.
34. Spencer-Martins, I., and N. van Uden. 1979. Extracellularamylolytic system of the yeast Lipomvces kononenkoae. Eur. J.Appl. Microbiol. Biotechnol. 6:241-250.
35. Suetsugu, N., S. Koyama, K. Takeo, and T. Kuge. 1974. Kineticstudies on the hydrolyses of a-. 1-, and y-cyclodextrins byTaka-amylase A. J. Biochem. 76:57-63.
36. Sukhumavasi, J., K. Kato, and T. Harada. 1975. Glucoamylaseof a strain of Endomncopsisfibiuliger-a isolated from mould bran(Look Pang) of Thailand. J. Ferment. Technol. 53:559-565.
37. Tamaki, H. 1980. Purification of glucoamylase isoenzymesproduced by Sac charomyc es diastatic us. Doshisha JoshiDaigaku Gakujutsu Kenkyu Nempo 31:270-286.
38. Touzi, A., J. P. Prebois, G. Moulin, F. Deschamps, and P. Galzy.1982. Production of food yeast from starchy substrates. Eur. J.Appl. Microbiol. Biotechnol. 15:232-236.
39. Truscheit, E., W. Frommer, B. Junge, L. Muller, D. D. Schmidt,and W. Wingender. 1981. Chemie und Biochemie mikrobiellera-Glucosidasen-Inhibitoren. Angew. Chem. 93:738-755.
40. Weber, K., and M. Osborn. 1969. The reliability of molecularweight determinations by dodecyl sulfate-polyacrylamide gelelectrophoresis. J. Biol. Chem. 244:4406-4412.
41. Wilson, J. J., and W. M. Ingledew. 1982. Isolation and charac-terization of Schuvanniomnvees alluvius amylolytic enzymes.AppI. Environ. Microbiol. 44:301-307.
42. Wilson, J. J., G. G. Khachatourians, and W. M. Ingledew. 1982.Schwanniomvces. SCP and ethanol from starch. Biotechnol.Lett. 4:333-338.
43. Yamashita, I., T. Hatano, and S. Fukui. 1984. Subunit structureof glucoamylase of Saccharomyces diastaticus. Agric. Biol.Chem. 48:1611-1616.
APPL. ENVIRON. MICROBIOL.
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from