proton motive force, energy recycling by endproduct excretion
TRANSCRIPT

Vol. 172, No. 12
Proton Motive Force, Energy Recycling by End Product Excretion,and Metabolic Uncoupling during Anaerobic Growth of
Pseudomonas mendocinaNICOLE VERDONI,' MIGUEL A. AON,2.3t* JEAN M. LEBEAULT,' AND DANIEL THOMAS2
Laboratoire de Technologie Enzymatique URA 41 du Centre National de la Recherche Scientifique2 and Division desProcedes Biotechnologiques,l Universite de Technologie de Compiegne, 60206 Compiegne, France, and E. C. Slater
Institute for Biochemical Research, University ofAmsterdam, 1018 TV Amsterdam, The Netherlands3
Received 21 May 1990/Accepted 11 September 1990
Batch cultures of Pseudomonas mendocina, grown in rich medium with glucose excess, showed metabolicdifferences dependent upon whether the growth conditions were aerobic or anaerobic, with or without addedelectron acceptor. Under anaerobic conditions in the absence of nitrate, P. mendocina reached the stationaryphase of growth after 2 or 3 days, followed by a stationary phase of 4 to 5 days. Under these conditions, a
mixed-type fermentative metabolism (formic, lactic, and acetic acids) appeared. A fivefold-higher specific rateof glucose consumption and eightfold-higher production of organic acids, compared with aerobic cultures, wereshown by this microorganism growing anaerobically in the absence of exogenous electron acceptors. Thegradients of organic acid produced by P. mendocina under these conditions reached a maximum (lactate, 180mV; formate, 150 mV; acetate, 215 mV) between days 2 and 3 of culture. The proton motive force (Ap)decreased during growth from -254 to -71 mV. The intracellular pH remained alkaline during the culture,reaching a steady-state value of 7.9. The gradients of organic acids apparently contributed to the generation ofa Ap, which, according to the Energy Recycling Model (P. A. M. Michels, J. P. J. Michels, J. Boonstra, andW. N. Konings, FEMS Mictobiol. Lett. 5:357-364, 1979), would produce an average energy gain of 1 or 1.5mol of ATP equivalents per mol of glucose consumed with H+/ATP stoichiometry of 3 or 2, respectively. LowYATP and Ygucose values were observed, suggesting that an uncoupled metabolism exists; i.e., ATP producedby catabolic processes is not directly used for biomass synthesis. This metabolic uncoupling could be inducedat least in part by organic acids and the ATP wastage could be induced by a membrane-bound ATPase involvedin intracellular pH regulation.
A well-studied example of metabolic aerobic-anaerobictransitions is the Pasteur effect (19, 23), defined as theincrease in glucose uptake caused by an increased rate ofglycolysis in the absence of 02. Less attention has been paidto the understanding of metabolic aerobic-anaerobic transi-tions through the coupling of cytoplasmic ATP synthesis tothose membrane-associated processes requiring ATP (e.g.,ion and solute transport). When cytoplasmic and membranefunctions are uncoupled, e.g., upon the collapse of mem-brane electrochemical gradients and consequent hydrolysisof available ATP, the cells necessarily become sensitive tohypoxia (9). The coupling between metabolic and membranetransport functions may be viewed as an important andunifying principle in cellular physiology (2, 9).Pseudomonads are well-known microorganisms for their
unusual catabolic potential and nutritional versatility (5, 14).In particular, Pseudomonas mendocina is a potentially use-
ful microorganism as a denitrifier and because of its ability toproduce alginate (18). The coupling of cytoplasmic ATPsynthesis to ion and solute transport processes requiringATP and the physiological response of pseudomonads underoxygen limitation and absence of electron acceptors, i.e.,energetic stress, are poorly understood. Therefore, the mainobjectives of the present work are addressed in the followingquestions: Does a strict aerobe like P. mendocina growunder anaerobic conditions and in the absence of added
* Corresponding author.t Present address: EC Slater Institute for Biochemical Research,
University of Amsterdam, 1018 TV-Amsterdam, The Netherlands.
electron acceptors? Does it ferment glucose? Does it use a
particular energetic strategy, e.g., energy recycling, underthose conditions? Does this microorganism show physiolog-ical and energetic differences when growing under aerobic or
anaerobic plus nitrate conditions with respect to anaerobio-sis and absence of electron acceptors?
In bacterial metabolism the energy supplied by catabolicsubstrates is translated into phosphorylation, redox, andtransmembrane (electro)chemical potentials. The presenceof an electrochemical proton gradient appears to be a generalprerequisite for the transmembrane movement of substratesand ions (13). Metabolite gradients represent a form ofenergy which can be used in certain cases as metabolicenergy. For instance, the efflux of end products of metabo-lism (organic acids) can occur via specific transport proteinsin symport with protons, and this process can lead to thegeneration of a proton motive force (Ap) which subsequentlycan drive energy-requiring processes. In this way, energyrecycling can occur (15, 22). It has been shown that energyrecycling by metabolic end products could occur anaerobi-cally with an energy gain of at least 20 to 45% by lactateexcretion in Streptococcus cremoris (22). Therefore, theexcretion of metabolic end products (organic acids) in fac-ultative anaerobes is an alternative mechanism, other thanATP hydrolysis, of creating an electrochemical gradient.Our results suggest that the aerobe P. mendocina, under
anaerobic conditions and in the absence of exogenous elec-tron acceptor, shows a mixed acid fermentation metabolism.The dissipation of lactic, formic, and acetic acid gradientsappears to contribute to an average energy gain of 1 to 1.5
6673
JOURNAL OF BACTERIOLOGY, Dec. 1990, p. 6673-66810021-9193/90/126673-09$02.00/0Copyright C 1990, American Society for Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.

6674 VERDONI ET AL.
mol of ATP equivalents per mol of glucose consumed ineither the exponential or the stationary phase of growth ofthe organism. The growth arrest of P. mendocina is appar-ently due to the highly uncoupled metabolism, i.e., the weakrelation between ATP synthesis (by catabolism) and growth(anabolism), shown by the microorganism under these con-ditions.
MATERIALS AND METHODS
Culture conditions. P. mendocina (collection no. 7521,Institut Pasteur, Paris, France) was a kind gift of 0. Men-doza, Universitd de Compifgne, Compiegne, France. Themicroorganism was aerobically maintained at 30'C in acomplex medium at pH 7.4, containing, in a final volume of1 liter, 5 g of meat extract, 10 g of microbiological peptone,5 g of NaCl, 15 g of glucose, and 1 g of KNO3. Storage wasin 25% glycerol at -80'C.
Anaerobic and aerobic growth. P. mendocina was growneither aerobically or anaerobically at 30'C in a complexmedium at pH 7.0 to 7.1, containing, in a final volume of 1liter, 16 g of meat extract, 33 g of microbiological peptone, 16g of NaCl, 50 g of glucose, and, when added, 3.3 g of KNO3.
Aerobic (100 ml) and anaerobic (200 ml) cultures weregrown in 250-ml Erlenmeyer flasks at 30'C and shaken at 120and 75 rpm, respectively. Anaerobic conditions were main-tained by nitrogen bubbling (<5 volumes of 02 per million)for 60 min in the culture vessel. Care was also taken tosample under anaerobic conditions.
Anaerobic growth of P. mendocina was monitoredthrough counts of viable cells on media lacking electronacceptors; three parallel cultures were run, and each point ofthe growth curve was obtained from triplicate viable cellcountings (CFU) in each Erlenmeyer flask. When P. men-docina batch cultures were grown under anaerobic condi-tions and in the absence of electron acceptors for 7 dayswithout sampling, the same stationary values of biomass (asCFU per milliliter) were recovered as in cultures in whichsamples were taken each 24 h. The latter observation rulesout the possibility of increasing 02 contamination by tracesof this gas present in nitrogen, since at each sampling theanaerobic conditions were reestablished by nitrogen bub-bling.At various stages of growth, 20-ml samples were taken and
external pH and biomass were measured. Biomass concen-tration was estimated from agar plate colony counts. The dryweight was determined as follows: cell suspensions of dif-ferent density were centrifuged twice at 10,000 x g for 30min (before the second time, the pellet was resuspended indistilled water); after the second centrifugation, the pelletwas recovered by resuspending the cells in water and thewhole volume was dried for 24 h at 100'C. The dry weight ofthe sample was obtained by weight difference, before andafter drying. The relationship between bacterial dry weightand CFU per milliliter was determined by dilution of a singlecell suspension obtained from exponentially growing aerobiccultures. Bacterial dry weight was linearly correlated (r =0.99; double logarithmic plots) to CFU per milliliter, for 107< n < 1010 bacteria per ml suspension (107 CFU/ml = 21.5mg of cell dry weight per liter). When <107 CFU/ml werecounted, dry-weight values were calculated by extrapolationfrom the double logarithmic plot. Cells were concentratedtwofold by centrifugation (10,000 x g, 10 min) and resus-pended in one-half of the initial culture medium volume. Thiscell suspension was used for subsequent measurements (seebelow).
Growth of P. mendocina in the presence of inhibitors.Cultures, 200 ml, of P. mendocina were grown as describedabove, and inhibitors or uncouplers were added at concen-trations specified for each experiment. N,N'-Dicyclohexyl-carbodiimide (DCCD) was dissolved in absolute ethanolwhich represented a 1% (vol/vol) or less final concentrationin the cultures. Todoacetate was dissolved in water withoutneutralization which induced an acidification of 0.2 pH unitwhen added to the cultures.Membrane preparation for assay of ATPase activity. P.
mendocina was grown in the presence of 6 g of glucose perliter under aerobic conditions as described above. Cells wereharvested in the exponential phase of growth (5 x 108 to 1 x109 cells per ml; 1.07 to 2.15 g of dry weight) by centrifuga-tion at 10,000 x g for 10 min, and the pellet was resuspendedin 2 ml of 1 mM Tris chloride, pH 7.6, containing 2 mMMgCl2 and sonicated on ice (30 15-s pulses). Particlessedimenting at between 80,000 x g (30 min) and 150,000 x g(2 h) were resuspended in the same buffer at concentrationsof 3 to 10 mg of protein per ml and immediately assayed orkept frozen at -800C. Further centrifugation (150,000 X g, 2h), after mixing the particles sedimenting at 80,000 x g and150,000 x g at similar protein concentrations, did not showa significant solubilization of the ATPase activity. ATPaseactivities of 1.5 and 0.81 pimol of ATP hydrolyzed min-' mgof protein-' were measured in the 80,000 x g (65% of thetotal activity recovered) and 150,000 x g (29% of the totalactivity recovered) pellets, respectively. The 80,000 x gpellet was used for the ATPase activity assays.The ATPase assay was performed at 30'C with an ATP-
regenerating system containing the following, in a finalvolume of 2 ml: 83 mM sucrose, 6 mM MgCl2, 33 mM Trischloride (pH 8.0), 5 mM ATP, 10 mM KHCO3, 0.5 mMphosphoenolpyruvate, 165 U of lactic dehydrogenase (825U/mg), 50 U of pyruvate kinase (200 U/mg), and NADH inexcess. The ATPase inhibition studies with DCCD wereperformed by preincubation of the membranes (60 to 90 ,ugof protein) at 30°C in a final volume of 100 [lI containing 1mM Tris chloride (pH 7.6), 2 mM MgCl2, and amounts of theinhibitor ranging from 0.05 to 1 mM. Controls withoutDCCD were run in parallel. The percentage of inhibition byDCCD was calculated by taking as 100% of activity thevalues obtained from controls in the absence of DCCD.After 1 h of incubation, the whole volume was diluted into atotal volume of 2 ml of ATPase assay mixture and the rate ofNADH disappearance was followed at 340 nm. The slope ofNADH disappearance was constant during the course of thereaction. The amount of membranes (in protein) used corre-sponded to the linear range with respect to enzymaticactivity.Measurement of organic acid gradients. Extracellular or-
ganic acid concentrations were determined by high-perfor-mance liquid chromatography with a cationic exchangecolumn (OA 1000; Alltech, Paris) at room temperature with5 mM H2SO4 as the mobile phase (Alltech Chromatography,1988). Samples of extracellular culture medium were previ-ously purified by passage through Cartridge packs (extractclean; Alltech). The following organic acids were monitoredby A210: citric, gluconic, propionic, isobutyric, butyric,malic, formic, acetic, and lactic. The amounts of organicacids detected at zero time (sterile culture medium) werediscounted as a background level. The only organic acidswhich evolved during culture were lactic, acetic, and formic(sensitivity threshold, 0.3 to 0.6 ,ug). Results similar to thoseobtained by high-performance liquid chromatography wereobserved when the culture medium was tested for lactic acid
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.

METABOLIC UNCOUPLING DURING P. MENDOCINA GROWTH
with a lactate oxidase enzymatic electrode. Only traces ofethanol were detected which did not increase during theculture when analyzed by gas chromatography (ShimadzuGC-8A; sensitivity threshold, 20 to 50 ng) in a column(WAW 80/100; 10% polyethylene glycol 20M) at a tempera-ture of 100'C.The intracellular concentration of organic acids was de-
termined by enzymatic test. Previous separation of cellsfrom the external medium by the silicone oil centrifugationtechnique was performed (22). Samples of twofold-concen-trated cellular suspension (0.1 to 0.3 ml) were centrifuged(12,000 x g, 10 min). The perchloric acid (PCA) and siliconeoil were preflushed with nitrogen and the Eppendorf tubeswere sealed. While loading the sample, a new nitrogenflushing was done before centrifugation. The supernatant ofthe PCA fraction at the bottom of the tubes was carefullyrecovered after removing the silicone oil phase. Resultsidentical to those of the first centrifugation were obtainedwhen the resuspended pellet was recentrifuged for 3 min at12,000 x g. The PCA fraction was neutralized with 65 ,ul of6 N NaOH, and a final pH of 8.5 + 0.3 was obtained. Theenzymatic assays (Boehringer kits) of formic, lactic, andacetic acids were performed in duplicate in a final volume of1.6 ml and left for 1 h at room temperature before reading at340 nm. The response (A340) is linear up to lactic, formate,and acetic acid concentrations of 0.5 mM. The absorbancewas determined against blanks (without enzyme) whichcame from cellular suspensions following the same process-ing, because after a certain period the A340 also increased inblanks. To rule out an increase in absorbance coming from"parasite" reactions, i.e., other than formate dehydroge-nase, lactate dehydrogenase, or acetyl-coenzyme A syn-thase which reduce NAD, the following control was madefor lactic acid: the sample was preincubated with immobi-lized lactate oxidase to consume lactic acid. Then theenzymatic assay with lactate dehydrogenase was performed.A 10% increase in absorbance was still observed which wasnot attributed to lactate dehydrogenase and thus was dis-counted.
Gradient calculations. From the data of intracellular watervolume (see below) and the amounts of intra- and extracel-lular lactate, formate, and acetate, the corresponding gradi-ents were calculated by the Nernst equation:
AP.OA = 2.3RT/F(log [OA]in/[OAJex) (1)with 2.3 RTIF = 60.44 mV (at 30°C), where OA is organicacids.Measurement of ApH, A*, and internal cell volume. The
pH gradient (ApH) and the transmembrane electric potential(A*4) values were calculated from the accumulation of di-methyloxazolidinedione (DMO) and the lipophilic cationtetraphenylphosphonium (TPP+), respectively (22). The useof benzoic acid as a ApH probe is precluded in Pseudomonassp. since in our previous experience this probe gave higherpH gradients, possibly due to degradation of benzoate (5).Samples of cellular suspensions (0.5 to 1 ml; 0.01 to 0.22 mgof dry weight) were incubated anaerobically (continuousnitrogen flushing) for 10 min at 30°C in the presence of either5 puM [14C]DMO or 0.153 p.M [3H]TPP+, transferred toMicrofuge tubes (previously flushed with nitrogen), andprocessed as described above. Aliquots of the supernatantand the PCA fraction were collected, dissolved in 2.5 ml of2,5-diphenyloxazole-naphthalene-dioxane scintillation mix-ture, and counted in a Beckman LS 8000 liquid scintillationspectrometer. Corrections by quenching and spillover ef-fects introduced by PCA and the culture medium were made.
Quenching and spillover effects were studied with 3H20 and["4C]taurine in the presence of equal volumes of culturemedium (5 ,ul) or PCA (50 p.1) to ensure comparable quench-ing as in the case of [14C]DMO or [3H]TPP+. For ApH andAkP calculations, the radioactivity attributable to nonspecificbinding of the probes to cell components was subtracted(22). The amounts of either [3H]TPP+ or [14C]DMO nonspe-cific binding (-10%) were estimated after deenergization ofthe cells by incubation in the presence of 1% (vol/vol) etherand ionophores (4 ,uM valinomycin plus 4 p.M nigericin) atroom temperature or 30°C for 15 or 60 min, respectively. Nosignificant differences in the amounts of probe binding wereobserved between the different treatments.The intracellular water volume was determined by the
method of Ten Brink and Konings (22). Samples (0.5 ml)were incubated with 2 ,uCi of either 3H20 or ['4Cjtaurine andhandled as for ApH and Aqp determinations. Similar values ofintracellular volumes were obtained for either aerobic oranaerobic (plus nitrate) conditions or growth in the presenceof 15 or 50 g of glucose per liter, giving values of 3.08 + 0.37(n = 3) and 2.93 + 0.3 (n = 3) p.1/mg of cell dry weight,respectively. No significant differences in intracellular vol-ume (3.0 + 0.4) were detected between cells growing expo-nentially (n = 3) and in the stationary phase (n = 3) ofgrowth. A value of 3 p.l/mg of cell dry weight was used in allcalculations.From the data of intracellular water volume and the
amounts of radioactivity in the supernatants and PCA frac-tions, the accumulation ratios of [3H]TPP-' and [14C]DMOwere calculated. The corresponding values of Aiq and ApH(in millivolts) were calculated with equation 1.
Materials. [carboxyl-14C]DMO (60 ,uCi/,umol) was fromNew England Nuclear, and TTP+ (26 mCi/,umol), 3H20 (250,uCi/ml), and [U-14C]taurine (891 ,uCi/mg) were obtainedfrom the Radiochemical Centre, Amersham, United King-dom. Silicone oil (density, 1.05 g/ml), DCCD. valinomycin,and nigericin were purchased from Sigma Chemical Co. Theenzyme kits for testing lactic, formic, and acetic acids werefrom Boehringer, Mannheim, Federal Republic of Germany.The glucose enzymatic test (glucose oxidase-peroxidase)was from Biotrol, Paris, France. All other chemicals werereagent grade and commercially available.
RESULTS AND DISCUSSION
Growth and mixed acid fermentation of P. mendocina underanaerobic conditions in the absence of exogenous electronacceptors. Growth of P. mendocina in batch culture in acomplex medium containing 50 g of glucose per liter, underanaerobic conditions and in the absence of electron accep-tors, proceeded for 2 or 3 days. After this initial phase, themicroorganism attained a stationary state of growth in whichit stayed at least until day 7 of culture (Fig. 1). Thefermentation carried out by this microorganism under anaer-obic conditions was characterized by the production oforganic acids. Only formate, lactate, and acetate were de-tected either intra- or extracellularly during the course of theculture. Figure 1 also shows the time course of organic acidgradients during culture. The gradients of the three organicacids increased steeply after 24 h, reaching a maximumbetween days 2 and 3 of culture. After day 3, while biomassapproached a stationary value, the organic acid gradientsbegan to decrease. A great decrease in the rate of glucoseconsumption under anaerobic conditions was observed after96 h of culture (not shown). The decrease in glucose con-sumption coincided with a pronounced decrease in the
VOL. 172, 1990 6675
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.
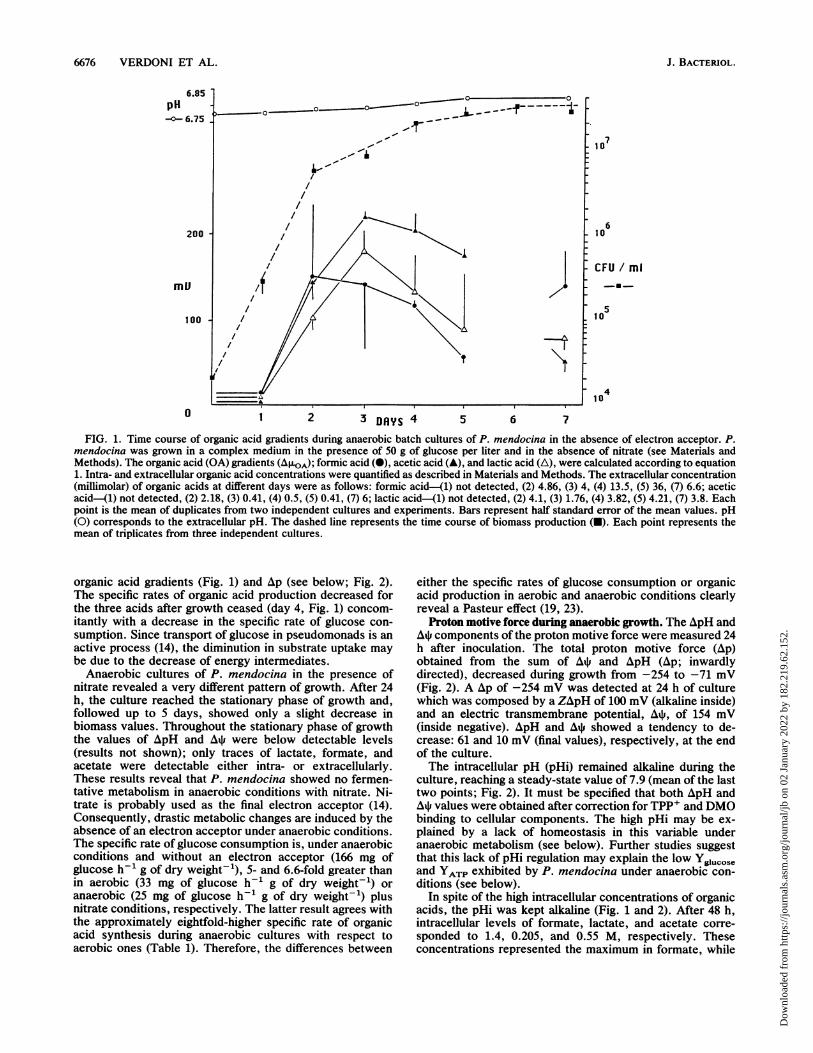
6676 VERDONI ET AL.
6.85pH-0- 6.75
200
mU
100
0
107
610
CFU / ml-_-
510
410
1 2 3 DO9YS 4 5 6 7
FIG. 1. Time course of organic acid gradients during anaerobic batch cultures of P. mendocina in the absence of electron acceptor. P.mendocina was grown in a complex medium in the presence of 50 g of glucose per liter and in the absence of nitrate (see Materials andMethods). The organic acid (OA) gradients (AIXOA); formic acid (0), acetic acid (A), and lactic acid (A), were calculated according to equation1. Intra- and extracellular organic acid concentrations were quantified as described in Materials and Methods. The extracellular concentration(millimolar) of organic acids at different days were as follows: formic acid-(1) not detected, (2) 4.86, (3) 4, (4) 13.5, (5) 36, (7) 6.6; aceticacid-41) not detected, (2) 2.18, (3) 0.41, (4) 0.5, (5) 0.41, (7) 6; lactic acid-(1) not detected, (2) 4.1, (3) 1.76, (4) 3.82, (5) 4.21, (7) 3.8. Eachpoint is the mean of duplicates from two independent cultures and experiments. Bars represent half standard error of the mean values. pH(0) corresponds to the extracellular pH. The dashed line represents the time course of biomass production (-). Each point represents themean of triplicates from three independent cultures.
organic acid gradients (Fig. 1) and Ap (see below; Fig. 2).The specific rates of organic acid production decreased forthe three acids after growth ceased (day 4, Fig. 1) concom-itantly with a decrease in the specific rate of glucose con-sumption. Since transport of glucose in pseudomonads is anactive process (14), the diminution in substrate uptake maybe due to the decrease of energy intermediates.Anaerobic cultures of P. mendocina in the presence of
nitrate revealed a very different pattern of growth. After 24h, the culture reached the stationary phase of growth and,followed up to 5 days, showed only a slight decrease inbiomass values. Throughout the stationary phase of growththe values of ApH and A* were below detectable levels(results not shown); only traces of lactate, formate, andacetate were detectable either intra- or extracellularly.These results reveal that P. mendocina showed no fermen-tative metabolism in anaerobic conditions with nitrate. Ni-trate is probably used as the final electron acceptor (14).Consequently, drastic metabolic changes are induced by theabsence of an electron acceptor under anaerobic conditions.The specific rate of glucose consumption is, under anaerobicconditions and without an electron acceptor (166 mg ofglucose h-1 g of dry weight-'), 5- and 6.6-fold greater thanin aerobic (33 mg of glucose h-1 g of dry weight-') oranaerobic (25 mg of glucose h-1 g of dry weight-') plusnitrate conditions, respectively. The latter result agrees withthe approximately eightfold-higher specific rate of organicacid synthesis during anaerobic cultures with respect toaerobic ones (Table 1). Therefore, the differences between
either the specific rates of glucose consumption or organicacid production in aerobic and anaerobic conditions clearlyreveal a Pasteur effect (19, 23).
Proton motive force during anaerobic growth. The ApH andAd components of the proton motive force were measured 24h after inoculation. The total proton motive force (Ap)obtained from the sum of Ali and ApH (Ap; inwardlydirected), decreased during growth from -254 to -71 mV(Fig. 2). A Ap of -254 mV was detected at 24 h of culturewhich was composed by a ZApH of 100 mV (alkaline inside)and an electric transmembrane potential, A+, of 154 mV(inside negative). ApH and Aq showed a tendency to de-crease: 61 and 10 mV (final values), respectively, at the endof the culture.The intracellular pH (pHi) remained alkaline during the
culture, reaching a steady-state value of 7.9 (mean of the lasttwo points; Fig. 2). It must be specified that both ApH andAdj values were obtained after correction for TPP+ and DMObinding to cellular components. The high pHi may be ex-plained by a lack of homeostasis in this variable underanaerobic metabolism (see below). Further studies suggestthat this lack of pHi regulation may explain the low Yglucoseand YATP exhibited by P. mendocina under anaerobic con-ditions (see below).
In spite of the high intracellular concentrations of organicacids, the pHi was kept alkaline (Fig. 1 and 2). After 48 h,intracellular levels of formate, lactate, and acetate corre-sponded to 1.4, 0.205, and 0.55 M, respectively. Theseconcentrations represented the maximum in formate, while
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.

METABOLIC UNCOUPLING DURING P. MENDOCINA GROWTH
pH7.0 - 0 0 0
A_
I--0- 6.8 -
300 -
200 -
mU
100
0
* Ap
A ZtpH* As
v Biomass
//
//
//
-.-E
l 2 3 4 5 6 7
-107
61 0
- CFU / ml
_-.
151 0
10
DRYSFIG. 2. Proton motive force in anaerobic culture of P. mendocina in the absence of electron acceptor. Cultures were performed as
described in the legend to Fig. 1. Samples were taken at 24-h intervals, and the components of Ap were determined (see Materials andMethods). pH (W) corresponds to the extracellular pH. The dashed line represents the time course of biomass production. Each point is themean of duplicates from two independent experiments and the attached bars represent half standard error of the mean values. Theexperiments are the same as those shown in Fig. 1.
the maxima for acetate (1.78 M) and lactate (1.73 M) wereattained at 72 h. The high cytoplasmic concentration oforganic acids and the alkaline pHi exhibited by the cellssuggest an active (ATP-fueled) proton extrusion by theH'-ATPase (3) or proton-driven active transport systems asfor lactic acid export in streptococci to alkalinize the cell'sinterior (15). Acidification of cytoplasm is expected to occuras a result of the accumulation of acidic end products offermentation (3, 12). However, it has recently been arguedthat only at low (<6) pH would glycolysis lead to theclassical proton stoichiometry (2 lactate- + 2H+) (10). Infermentative metabolism at high (>8) pH, protons need to beneither accumulated nor depleted, although acidification byATP hydrolysis (one proton per ATP) may be expected.Thus, to maintain an alkaline pHi would allow a bettercontrol of internal H' concentrations for P. mendocinafermenting glucose.
Proton motive force during aerobic growth. Growth of P.
TABLE 1. Specific rates of organic acid productiona
% with given specific rate of productionOrganic Anaerobic, -NO3 Aerobic, -NO3acid (17 g h-l g of dry (1.92gh-of
wt-1) dry wt- 1)
Lactic 5 10Formic 65 88Acetic 30 2
a The intra- and extracellular amounts of organic acids and biomass dryweights were determined as described in Materials and Methods. The calcu-lated rates correspond to the initial ones. Each value represents the mean ofduplicates from two independent cultures and experiments.
mendocina under aerobic conditions is shown in Fig. 3. Thetime course of tsp was determined: qualitative and quantita-tive differences with respect to anaerobic conditions wereobserved. During the exponential phase of growth, Apchanged from inwardly to outwardly directed, which impliesa more acid pH and a more positive An* inside the cell.Depolarization of the Ats by electrogenic K+ uptake hasbeen reported (1). Ap seems to reach a stationary value (137mV) at the end of the culture with a pH difference of 0.45unit (pHi = 6.98). It is remarkable that 5 h after inoculationa high pHi (8.82) was determined which decreased during theexponential phase of growth, reaching steady-state values of7.3 and 6.98 (last two points). These apparent wide varia-tions in internal H+ concentrations are intriguing. It seemsthat P. mendocina is unable to regulate its pHi duringaerobic metabolism (see also above). In fact, during the first(4 to 5) hours of aerobic growth, high extracellular concen-trations of formate (0.1 M) were detected (two independentexperiments), while only traces (<1 mM) of lactate and noacetate were found. Only traces (<1 mM) of these organicacids were detected at the intracellular level. Thus, duringthe lag phase an apparent anaerobic metabolism and formicacid excretion were observed and a consequent alkaliniza-tion revealed by ApH values was obtained (Fig. 3). Similarresults have been observed in continuous cultures at highdilution rate (0.5 has) and high dissolved 02 tensions (N.Verdoni and M. A. Aon, unpublished results).Energy production by organic acid excretion in anaerobic
conditions. The results presented above (Fig. 1 and 2)suggest that the excretion of lactic, formic, and acetic acidscould contribute to the energy requirements of survival andgrowth of the aerobe P. mendocina under anaerobic condi-
VOL. 172, 1990 6677
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.

6678 VERDONI ET AL.
8.0 -
plJ , , o Go oG-0-7.0 L
5-I 0
C-)
200 109
I 0100 /
Bloinss0_ 10
-100
-150
0a 5 10 15 20 25
HOURSFIG. 3. Proton motive force in aerobic batch cultures of P.
mendocina. Cultures were performed as described in the legend toFig. 1 but in the presence of oxygen. Samples were taken at 4-hintervals and treated as described in Materials and Methods forApH, Aq/, and Ap determinations.
tions in the absence of electron acceptors. The energy-recycling model postulates that efflux or translocation ofsolutes (fermentation products) according to the electro-chemical solute gradient (ARQOA) can result in the generationof Ap (15). The energy produced by glucose fermentationand end product excretion can be calculated with the follow-ing equation, which in its general form follows that of TenBrink and Konings (22):ATP equivalents produced/mole of glucose consumed =
2+3(n-1)/p (2)where n represents the number of protons translocated insymport with one molecule of either lactate, formate, oracetate, which are assumed to be independently transportedsince their gradients were summed; and p represents thenumber of protons translocated per molecule of ATP syn-thesized. Two molecules ofATP per molecule of glucose aresynthesized by substrate-level phosphorylation, and threemolecules of organic acids comprising acetate, lactate, andformate are assumed to be produced (8). Since one protonper molecule of lactate, formate, or acetate is producedintracellularly and the three organic acids are monovalentand negatively charged, 3 (n - 1) represents the number oftranslocated protons which contribute to the generation ofthe proton motive force.The calculated number of ATP equivalents produced per
a)
0';0)
4
3-
2-
1 -
/I
1e
0
P=2 7
.3
5 A"'
2-An/
//,//1 2 3
n ( it/ OR)FIG. 4. Energy recycling in anaerobic batch cultures of P. men-
docina. The stoichiometry n (H+/organic acids [OA]) was calculatedaccording to equation 3 (see text) after experimental determinationsof ApH, A+, and Ap. p is the H+/ATP stoichiometry. The ATPequivalents produced per mole of glucose consumed were obtainedby applying equation 2 (see text). Each point corresponds to theresults of the experiments described in the legends to Fig. 1 and 2.The number at each point indicates the day of culture.
mole of glucose consumed (see equation 2) is shown in Fig.4 for p = 2 and 3. The n values were calculated from thefollowing expression (22):
n = (AM) - AI.OA)/AP (3)An energy gain of 1.5 (p = 2) to 1 (p = 3) mol of ATPequivalents per mol of glucose on average seemed to haveoccurred, reaching maximum values in the stationary phaseof growth. Since Ap and AROA are opposing forces andenergy recycling appears to occur, it may be stated thatAPOA is always more important than Ap after the first day ofculture (see equation 3). At 24 h of culture, negative valuesof n were obtained (Fig. 4). Ten Brink and Konings (22)postulated that n values significantly lower than 1 indicatethat energy loss had occurred; however, this could not beascertained since no organic acids were detected either intra-or extracellularly at 24 h.One of the main postulates of the energy-recycling model
is that end product excretion is carrier mediated and thatpositive charges or protons or both are translocated with theend product, from the cytoplasm to the external medium.However, some experimental evidence shows the presenceof organic acid carriers, a prerequisite for the energy gain byenergy recycling during fermentation (22). The stoichiom-etry of protons/lactate (n) was assumed to be only dependenton the pH.Ut (15). A change in the stoichiometry wasobserved (Fig. 4), while the extracellular pH remainednearly constant during anaerobic culture of P. mendocina(Fig. 1). On the other hand, since a diminution of 0.86 U inpHi was observed (Fig. 2), it seems more likely that, if acarrier exists, the pH dependence of putative ionizablegroups would be sensitive to the pHi.Growth arrest and metabolic uncoupling in P. mendocina
under anaerobic conditions. The possible involvement ofintracellular pH regulation in growth arrest was also inves-tigated.
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.
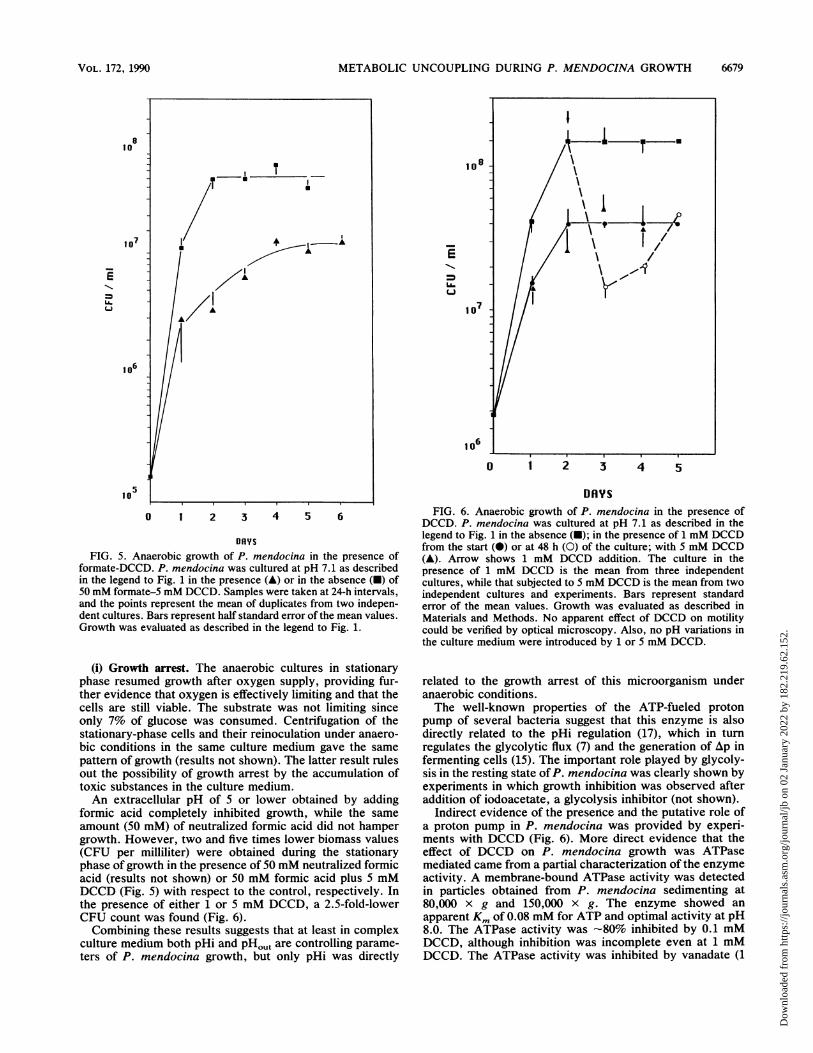
METABOLIC UNCOUPLING DURING P. MENDOCINA GROWTH
EA
A"'AVA
105
0 1 2 3 4 5 6
DAYS
FIG. 5. Anaerobic growth of P. mendocina in the presence offormate-DCCD. P. mendocina was cultured at pH 7.1 as describedin the legend to Fig. 1 in the presence (A) or in the absence (-) of50 mM formate-5 mM DCCD. Samples were taken at 24-h intervals,and the points represent the mean of duplicates from two indepen-dent cultures. Bars represent half standard error of the mean values.Growth was evaluated as described in the legend to Fig. 1.
(i) Growth arrest. The anaerobic cultures in stationaryphase resumed growth after oxygen supply, providing fur-ther evidence that oxygen is effectively limiting and that thecells are still viable. The substrate was not limiting sinceonly 7% of glucose was consumed. Centrifugation of thestationary-phase cells and their reinoculation under anaero-bic conditions in the same culture medium gave the samepattern of growth (results not shown). The latter result rulesout the possibility of growth arrest by the accumulation oftoxic substances in the culture medium.An extracellular pH of 5 or lower obtained by adding
formic acid completely inhibited growth, while the sameamount (50 mM) of neutralized formic acid did not hampergrowth. However, two and five times lower biomass values(CFU per milliliter) were obtained during the stationaryphase of growth in the presence of 50 mM neutralized formicacid (results not shown) or 50 mM formic acid plus 5 mMDCCD (Fig. 5) with respect to the control, respectively. Inthe presence of either 1 or 5 mM DCCD, a 2.5-fold-lowerCFU count was found (Fig. 6).Combining these results suggests that at least in complex
culture medium both pHi and pH0ut are controlling parame-ters of P. mendocina growth, but only pHi was directly
108
I-
Lo
107
6679
0 1 2 3 4 5
DRYSFIG. 6. Anaerobic growth of P. mendocina in the presence of
DCCD. P. mendocina was cultured at pH 7.1 as described in thelegend to Fig. 1 in the absence (-); in the presence of 1 mM DCCDfrom the start (0) or at 48 h (0) of the culture; with 5 mM DCCD(A). Arrow shows 1 mM DCCD addition. The culture in thepresence of 1 mM DCCD is the mean from three independentcultures, while that subjected to 5 mM DCCD is the mean from twoindependent cultures and experiments. Bars represent standarderror of the mean values. Growth was evaluated as described inMaterials and Methods. No apparent effect of DCCD on motilitycould be verified by optical microscopy. Also, no pH variations inthe culture medium were introduced by 1 or 5 mM DCCD.
related to the growth arrest of this microorganism underanaerobic conditions.The well-known properties of the ATP-fueled proton
pump of several bacteria suggest that this enzyme is alsodirectly related to the pHi regulation (17), which in turnregulates the glycolytic flux (7) and the generation of Ap infermenting cells (15). The important role played by glycoly-sis in the resting state of P. mendocina was clearly shown byexperiments in which growth inhibition was observed afteraddition of iodoacetate, a glycolysis inhibitor (not shown).
Indirect evidence of the presence and the putative role ofa proton pump in P. mendocina was provided by experi-ments with DCCD (Fig. 6). More direct evidence that theeffect of DCCD on P. mendocina growth was ATPasemediated came from a partial characterization of the enzymeactivity. A membrane-bound ATPase activity was detectedin particles obtained from P. mendocina sedimenting at80,000 x g and 150,000 X g. The enzyme showed an
apparent Km of 0.08 mM for ATP and optimal activity at pH8.0. The ATPase activity was -80% inhibited by 0.1 mMDCCD, although inhibition was incomplete even at 1 mMDCCD. The ATPase activity was inhibited by vanadate (1
VOL. 172, 1990
810
to7
16
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.

6680 VERDONI ET AL.
mM vanadate, 10-min preincubation: 82% inhibitionwithrespect to the control) and sodium azide (1 mM sodiumazide, 10-min preincubation: 46% inhibition with respect tothe control). Therefore, the present partial characterizationsuggests that at least one of the membrane-bound ATPasesof P. mendocina is of the E1-E2 type (P-ATPases) (16),although at present a combination of several ATPases withdifferent inhibition sensitivities cannot be excluded. Theseresults are consistent with the effect of DCCD on P. men-docina growth being ATPase mediated.
Anaerobic culture in the presence of exogenous neutral-ized formic acid and DCCD (Fig. 5) suggests that pHiregulation not only is a function of H+-ATPase inhibition,but also depends on passive membrane permeation (3) ororganic acid excretion (15). Also in the presence of formicacid and DCCD, the culture reached stationary biomassvalues fivefold lower than the control, suggesting that anincrease in the inward leakage of the exogenous formic acidwithout pHi regulation (ATPase inhibited) provokes a moredrastic decrease in viability. It must be pointed out that, atpH near 7, according to the Henderson-Hasselbach equationonly a small fraction of formate would be undissociated.Then, a continuous displacement of the equilibrium bycellular uptake of the undissociated acid might exist. More-over, the uncoupling effect of acetate has been reported inCandida utilis continuous cultures (11).
If DCCD is added after 48 h of culture, a strong decreasein viability in the following 24 h was observed, whichrecovered at the same level of CFU per milliliter withrespect to cultures that were started in the presence of 1 mMDCCD (Fig. 6). The subsequent increase in biomass afteraddition of 1 mM DCCD at 48 h suggests that alternatemechanisms of regulation of pHi with different sensitivity toDCCD are induced (Fig. 6) (4, 17).
(ii) Metabolic uncoupling and energy recycling. For S.cremoris the extra production of energy or ATP equivalentshas been associated with an increase in yield (22). Accordingto the biomass produced in P. mendocina anaerobic cul-tures, 81 ± 8 mg of dry weight per liter (n = 2, independentexperiments), and glucose consumed (3.4 ± 0.4 g/liter; n =2, independent experiments), a low Yglucose (4.26 g ofbiomass synthesized per mol of glucose consumed) wasobtained. The same yields of biomass with respect to glucosecould be calculated when P. mendocina was grown anaero-bically in batch cultures in the presence of [14C]glucose andin the absence of exogenous electron acceptors (Verdoni andAon, unpublished results).By considering an ATP demand of 40.73 mmol/g of dry
weight (21) and a stoichiometry of 2 ATP per mol of glucoseconsumed, a YATP = 2.13 g of biomass synthesized per molof generated ATP was calculated. Thus, the ATP needed toproduce 1 g of biomass is 469 mmol. In the presence of 1 mMDCCD (Fig. 6), the ATP needed to produce 1 g of biomassunder anaerobic conditions was increased further by 30%(654 mmol). Considering YATP values of 14 for anaerobicbacterial cultures growing in rich medium with a singlecarbon-substrate energy source (21), 71 mmol of ATP isrequired for the synthesis of 1 g of biomass. These observa-tions show an uncoupling between anabolic and catabolicprocesses; i.e., the ATP produced in catabolism is not usedfor biomass production (21). Most of the ATP produced(85%) and the glucose consumed were not associated withgrowth. The uncoupled metabolism may be due to theactivity of the membrane energy-consuming processes,which through the uncoupling effect of organic acids willcause a Ap dissipation. Therefore, a continuous ATP-fueled
H' extrusion by the proton pump will be operating toregulate and maintain a Ap and pHi compatible with growth.It has been reported that 51 to 58% of the total ATPproduced by Escherichia coli was apparently used for mem-brane energization by the membrane H+-ATPase (20). Thelatter effect would be at least in part the cause of the lowyields observed and the fact that 85% of the ATP formed byP. mendocina was not associated with growth. The lowYATP values observed may also have resulted from anoverestimation of the ATP stoichiometry because of anoperative methylglyoxal bypass (6, 21). The methylglyoxalshunt was observed in P. saccharophila cells poisoned withiodoacetate (6), although the existence of this pathway hasyet to be demonstrated in P. mendocina.
According to the energy-recycling model, for continuousorganic acid excretion to occur, the Ap must be lower thanthe A0A (15). The ATPase inhibition by DCCD provokes acollapse of ApH (3); then Ap is lower, which favors acontinuous efflux of end products. In the experiment shownin Fig. 6, both the control and DCCD-treated cultures (1mM) consumed the same amount of glucose, although 30%more ATP was needed by the latter to synthesize 1 g ofbiomass. These considerations lead us to suggest that energyrecycling contributes to greater yields of at least 30% inanaerobiosis, even under conditions of metabolic uncou-pling. Therefore, according to the results obtained in batchcultures, the energy-recycling mechanism appears to bepartly associated with growth.
In summary, the results obtained suggest that energizationof the membrane in P. mendocina by end product excretionunder electron acceptor-limiting conditions may be envis-aged as a strategy of providing additional energy for eithergrowth or maintenance. The growth of this aerobe underanaerobic conditions without exogenous electron acceptorsproceeds with very low yields due to the highly uncoupledmetabolism exhibited under these conditions; i.e., most ofthe ATP produced by catabolism is not associated withgrowth.
ACKNOWLEDGMENTS
We thank W. N. Konings for helpful suggestions and criticallyreading the manuscript and S. Cortassa for her continuous interestand enlightening criticism throughout this work. Also, thanks aregiven to Karel van Dam, EC Slater Institute, University of Amster-dam, for helpful suggestions and kindly allowing us to petform in hislaboratory some of the experiments reported in this work. Discus-sions with H. V. Westerhoff and J. A. Berden, EC Slater Institute,University of Amsterdam, and advice on the ATPase measurementsby L. Hartog, EC Slater Institute, University of Amsterdam, aregratefully acknowledged. We thank M. L. Dutrieux for improvingthe edition of the manuscript. This work has benefitted from thethoughtful criticism of the anonymous reviewers which we alsogratefully acknowledge.
LITERATURE CITED1. Abee, T., K. J. Hellingwerf, and W. N. Konings. 1988. Effects of
potassium ions on proton motive force in Rhodobactersphaeroides. J. Bacteriol. 170:5647-5653.
2. Aon, M. A., and S. Cortassa. 1989. Plant cell growth regulation:a bioelectromechanochemical model. J. Theor. Biol. 138:429-456.
3. Baronofsky, J. J., W. J. A. Schreurs, and E. R. Kashket. 1984.Uncoupling by acetic acid limits growth of and acetogenesis byClostridium thermoaceticum. Appl. Environ. Microbiol. 48:1134-1139.
4. Booth, I. R. 1985. Regulation of cytoplasmic pH in bacteria.Microbiol. Rev. 49:359-378.
5. Clarke, P. H., and L. N. Ornston. 1975. Metabolic pathways and
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.

METABOLIC UNCOUPLING DURING P. MENDOCINA GkOWTH
regulation: I, p. 253. In P. H. Clarke and L. N. Ornston (ed.),Genetics and biochemistry of pseudomonas. John Wiley &Sons, London.
6. Cooper, R. A. 1984. Metabolism of methylglyoxal in microor-ganisms. Annu. Rev. Microbiol. 38:49-68.
7. den Hollander, J. A., K. Ugurbil, T. R. Brown, and R. G.Shulman. 1981. Phosphorous-31 nuclear magnetic resonancestudies of the effect of oxygen upon glycolysis in yeast. Bio-chemistry 20:5871-5880.
8. Gottschalko G. 1979. Bacterial metabolism. Springer-Verlag,Berlin.
9. Hochachka, P. W. 1986. Defense strategies against hypoxia andhypothermia. Science 231:234-241.
10. Hochachka, P. W., and T. P. Mommsen. 1983. Protons andanaerobiosis. Science 219:1391-1397.
11. Hueting, S., and D. W. Tempest. 1977. Influence of acetate in thegrowth of Candida utilis in continous culture. Arch. Microbiol.115:73-78.
12. Kashket, E. R. 1987. Bioenergetics of lactic acid bacteria:cytoplasmic pH and osmotolerance. FEMS Microbiol. Rev.46:233-244.
13. Konings, W. N., and P. A. M. Michels. 1980. Electron-transfer-driven solute translocation across bacterial membranes, p.33-86. In C. Knowles (ed.), Diversity of bacterial respiratorysystems. CRC Press, Inc., Boca Raton, Fla.
14. Lessie, J. G., and P. V. Phibbs, Jr. 1984. Alternative pathwaysof carbohydrate utilization in pseudomonads. Annu. Rev. Mi-
crobiol. 38:359-387.15. Michels, P. A. M., J. P. J. Michels, J. Boonstra, and W. N.
Konings. 1979. Generation of an electrochemical proton gradi-ent in bacteria by the excretion of metabolic end products.FEMS Microbiol. Lett. 5:357-364.
16. Nelson, N., and L. Taiz. 1989. The evolution of H+-ATPases.Trends Biochem. Sci. 14:113-116.
17. Padan, E., D. Zilberstein, and S. Schuldiner. 1981. pH homeo-stasis in bacteria. Biochim. Biophys. Acta 650:151-166.
18. Sengha, S. S., A. J. Anderson, A. J. Hacking, and E. A. Dawes.1989. The production of alginate by Pseudomonas mendocina inbatch and continuous culture. J. Gen. Microbiol. 135:795-804.
19. Sols, A. 1975. The Pasteur effect in the allosteric era, p. 199-206.In A. Kornberg, B. L. Horecker, L. Cornudella, and J. Oro(ed.), Reflections in biochemistry. Pergamon Press, New York.
20. Stouthamer, A. H., and C. W. Bettenhaussen. 1977. A continousculture study of an ATPase-negative mutant of Escherichia coli.Arch. Microbiol. 113:185-189.
21. Tempest, D. W., and 0. M. Neljssel. 1984. The status of YATPand maintenance energy as biologically interpretable phenom-ena. Annu. Rev. Microbiol. 38:459-486.
22. Ten Brink, B., and W. N. Konings. 1982. Electrochemicalproton gradient and lactate concentration gradient in Strepto-coccus cremoris cells grown in batch culture. J. Bacteriol.152:682-686.
23. Wenner, C. E. 1979. Pasteur and Crabtree effects-assay incells. Methods Enzymol. 55:289-297.
VOL. 172, 1990 6681
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
02
Janu
ary
2022
by
182.
219.
62.1
52.