proteins
TRANSCRIPT

12From DNA to Protein: Genotype to Phenotype

OBJECTIVES:
How do we go from DNA to mRNA?
- what happens to an mRNA?
How do we go from mRNA to a protein?
-what happens to a protein?

12.1 What Is the Evidence that Genes Code for Proteins?
The gene-enzyme relationship has been revised to the one-gene, one-polypeptide relationship.
Example: In hemoglobin, each polypeptide chain is specified by a separate gene.
Other genes code for RNA that is not translated to polypeptides; some genes are involved in controlling other genes.

12.2 How Does Information Flow from Genes to Proteins?
Expression of a gene to form a polypeptide:
• Transcription—copies information from gene to a sequence of RNA.
• Translation—converts RNA sequence to amino acid sequence.
12.2 How Does Information Flow from Genes to Proteins?
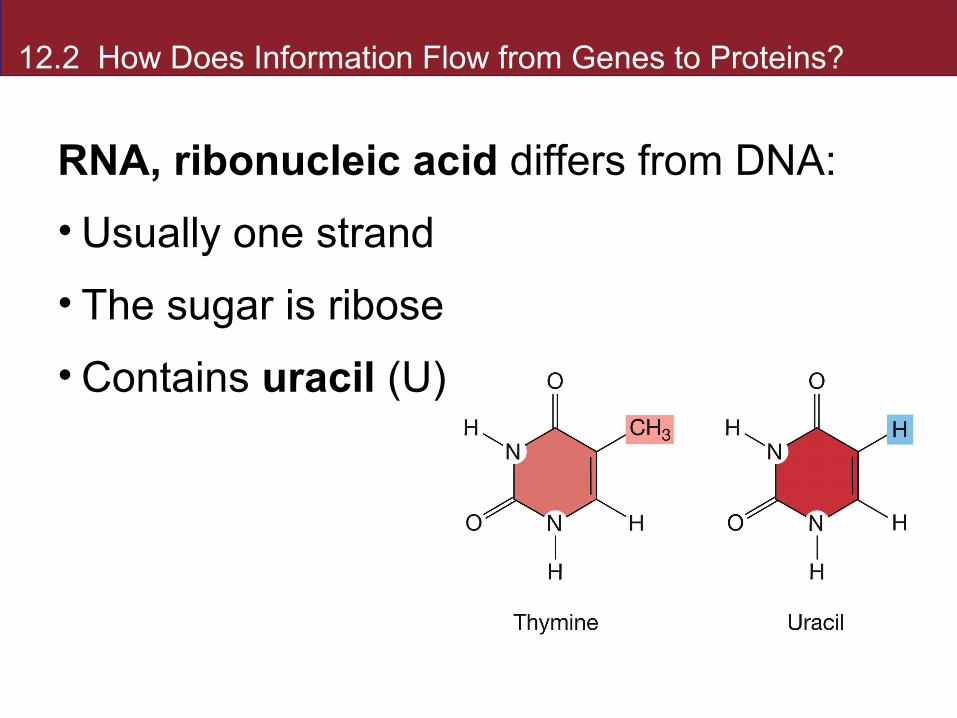
RNA, ribonucleic acid differs from DNA:
• Usually one strand
• The sugar is ribose
• Contains uracil (U) instead of thymine (T)
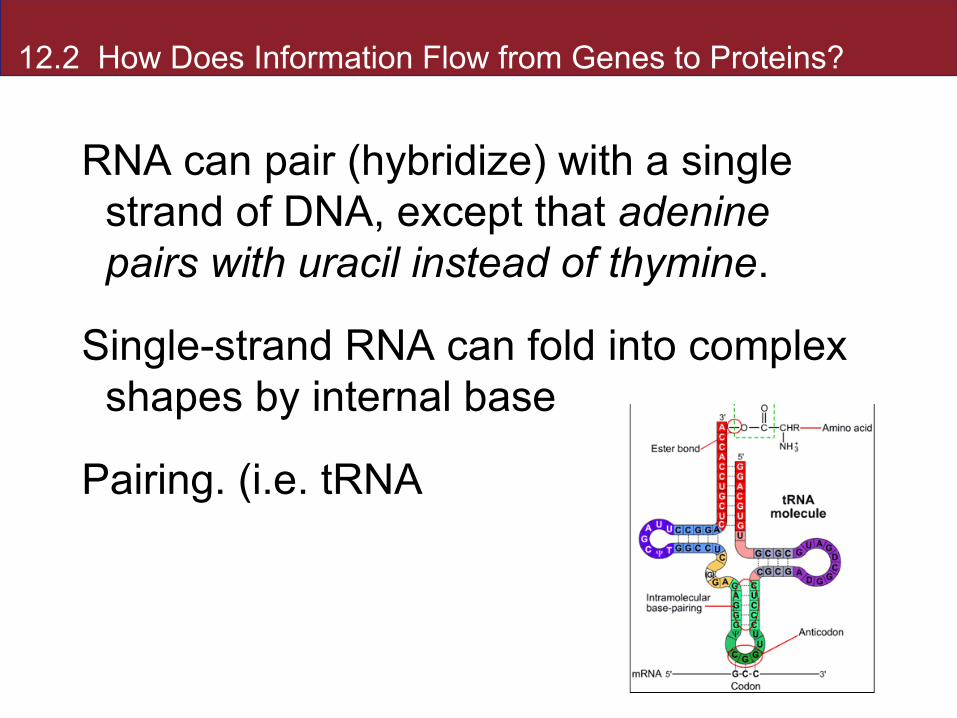
12.2 How Does Information Flow from Genes to Proteins?
RNA can pair (hybridize) with a single strand of DNA, except that adenine pairs with uracil instead of thymine.
Single-strand RNA can fold into complex shapes by internal base
Pairing. (i.e. tRNA

Figure 12.2 The Central Dogma
The central dogma of molecular biology: information flows in one direction when genes
are expressed (Francis Crick).
CENTRAL DOGMA:
reverse transcriptase - HIV

12.2 How Does Information Flow from Genes to Proteins?
The central dogma raised two questions:
• How does genetic information get from the nucleus to the cytoplasm?
• What is the relationship between a DNA sequence and an amino acid sequence?

12.2 How Does Information Flow from Genes to Proteins?
Transcription:
1. Messenger hypothesis—messenger RNA (mRNA) forms as a complementary copy of DNA and carries information to the cytoplasm.

Figure 12.3 From Gene to Protein
Synthesis of DNA from RNA is reverse transcription.
Viruses that do this are retroviruses.

12.2 How Does Information Flow from Genes to Proteins?
2. Adapter hypothesis—an adapter molecule that can bind amino acids, and recognize a nucleotide sequence—transfer RNA (tRNA).
Translation: tRNA molecules carrying amino acids line up on mRNA in proper sequence for the polypeptide chain
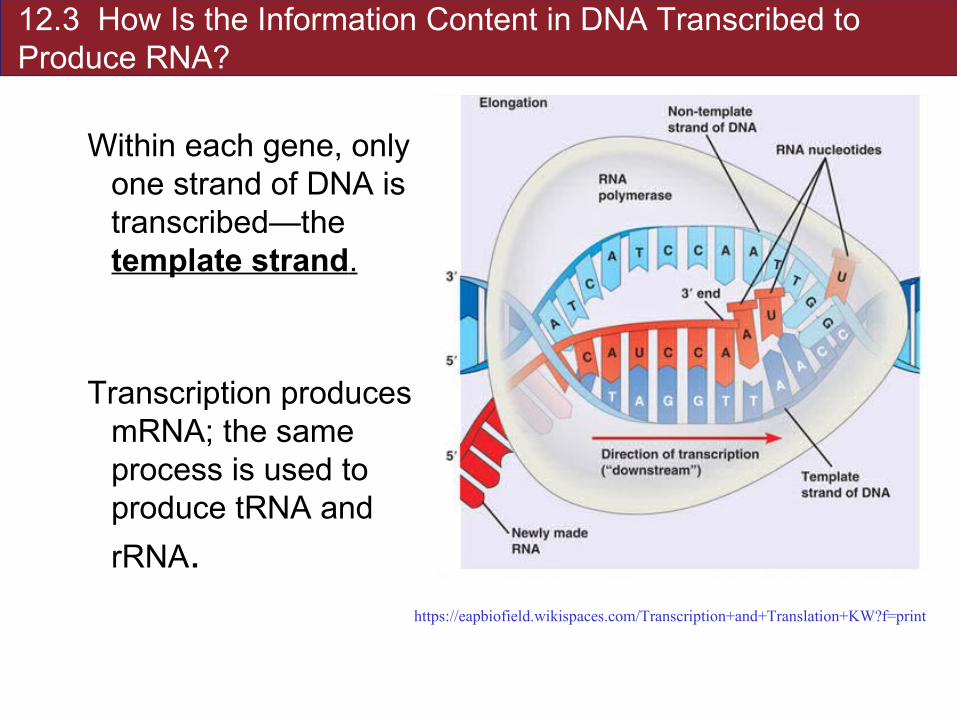
12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
Within each gene, only one strand of DNA is transcribed—the template strand.
Transcription produces mRNA; the same process is used to produce tRNA and
rRNA.https://eapbiofield.wikispaces.com/Transcription+and+Translation+KW?f=print

12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
RNA polymerases catalyze synthesis of RNA.
RNA polymerases are processive—a single enzyme-template binding results in polymerization of hundreds of RNA bases.

Figure 12.4 RNA Polymerase
What are the bonds called thatform between ribose bases?

12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
Transcription occurs in three phases:
• Initiation
• Elongation
• Termination
http://vcell.ndsu.edu/animations/transcription/movie.htm

12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
Initiation requires a promoter—a special sequence of DNA.
RNA polymerase binds to the promoter.
Promoter tells RNA polymerase where to start, which direction to go in, and which strand of DNA to transcribe.
Part of each promoter is the initiation site.
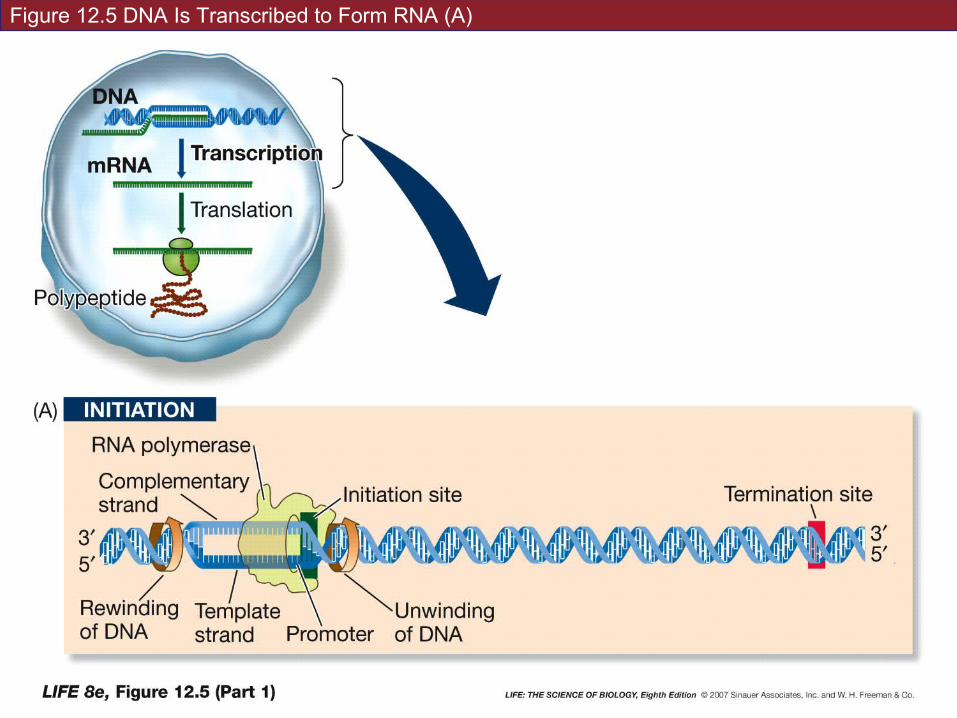
Figure 12.5 DNA Is Transcribed to Form RNA (A)

12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
Elongation: RNA polymerase unwinds DNA about 10 base pairs at a time; reads template in 3' to 5' direction.
The RNA transcript is antiparallel to the DNA template strand.
RNA polymerases do not proofread and correct mistakes.

Figure 12.5 DNA Is Transcribed to Form RNA (B)

12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
Termination: specified by a specific DNA base sequence.
Mechanisms of termination are complex and varied.
Eukaryotes—first product is a pre-mRNA that is longer than the final mRNA and must undergo processing.

Figure 12.5 DNA Is Transcribed to Form RNA (C)

mRNA Splicing
The entire gene is transcripted into a message. Some of the message isJunk (introns) and is removed before exiting the nucleus.
A spliceosome is a complex of specialized RNA and protein that removes introns from a pre-mRNA This process is generally referred to as splicing.
Introns typically have a “GU” nucleotide sequence at the 5' end splice site, and an AG at the 3' end splice site.

Alternative splicing is a source of genetic diversity in eukaryotes. Splicing has been used to account for the relatively small number of genes in the human genome.
Old estimates were for 100,000 genes but due to the Human Genome Project the figure is now roughly 20,000 genes.
One particular Drosophila gene (DSCAM) can be alternatively spliced into 38,000 different mRNAs.
http://vcell.ndsu.edu/animations/mrnasplicing/movie.htm

12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
The genetic code: specifies which amino acids will be used to build a protein
Codon: a sequence of three bases. Each codon specifies a particular amino acid.
Start codon: AUG—initiation signal for translation – Always!! (well some exceptions…)
Stop codons: stops translation and polypeptide is released
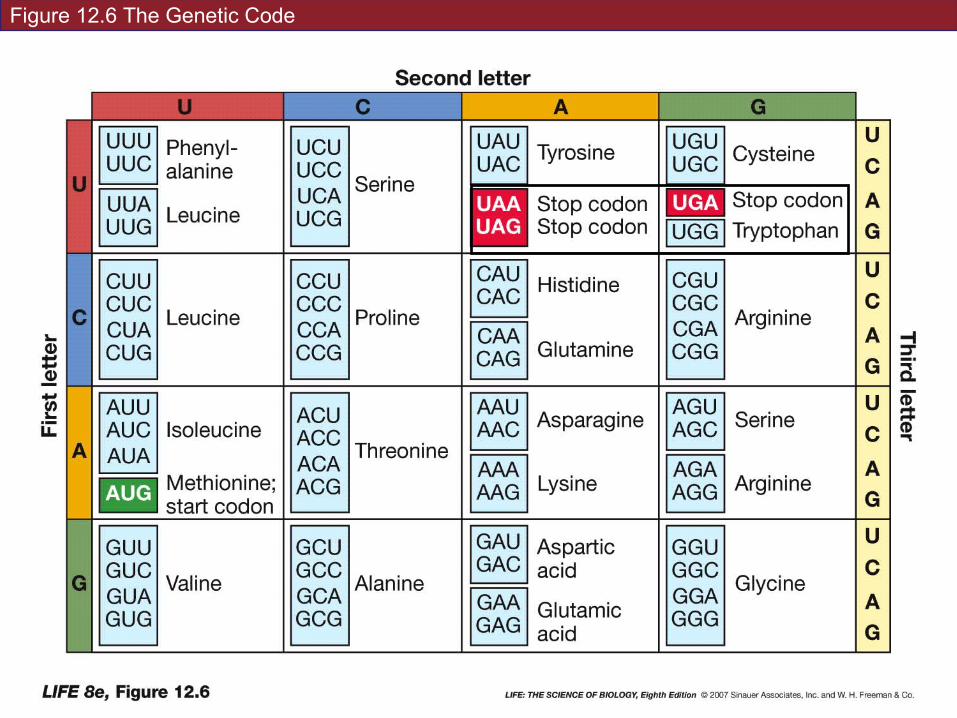
Figure 12.6 The Genetic Code

12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
For most amino acids, there is more than one codon; the genetic code is redundant.
But not ambiguous—each codon specifies only one amino acid.

12.3 How Is the Information Content in DNA Transcribed to Produce RNA?
The genetic code is nearly universal: the codons that specify amino acids are the same in all organisms.
Exceptions: within mitochondria and chloroplasts, and in one group of protists.

12.4 How Is RNA Translated into Proteins?
tRNA, the adaptor molecule: for each amino acid, there is a specific type of tRNA.
Functions of tRNA:
• Carries an amino acid
• Associates with mRNA molecules
• Interacts with ribosomes
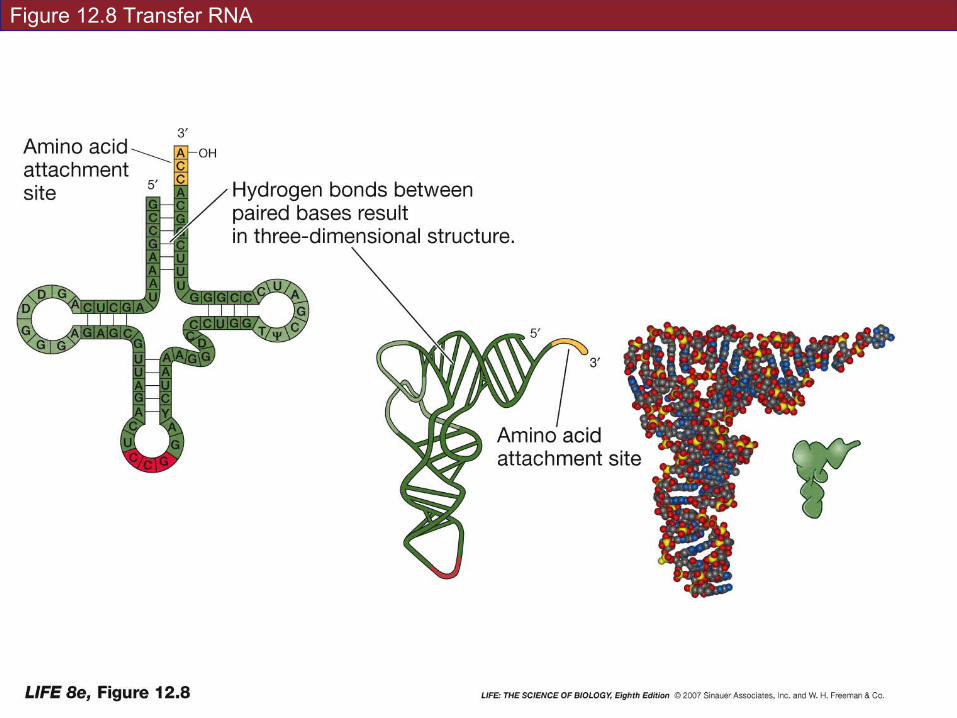
Figure 12.8 Transfer RNA

12.4 How Is RNA Translated into Proteins?
The conformation (three-dimensional shape) of tRNA results from base pairing (H bonds) within the molecule.
3' end is the amino acid attachment site—binds covalently. Always CCA.
Anticodon: site of base pairing with mRNA. Unique for each species of tRNA.

12.4 How Is RNA Translated into Proteins?
Example:
DNA codon for arginine: 3'-GCC-5'
Complementary mRNA: 3'-CGG-5'
Anticodon on the tRNA: 3'-GCC-5' This tRNA is charged with arginine.
TAC - ___ ____ ____ - TAC

12.4 How Is RNA Translated into Proteins?
Wobble: specificity for the base at the 3' end of the codon is not always observed.
Example: codons for alanine—GCA, GCC, and GCU—are recognized by the same tRNA.
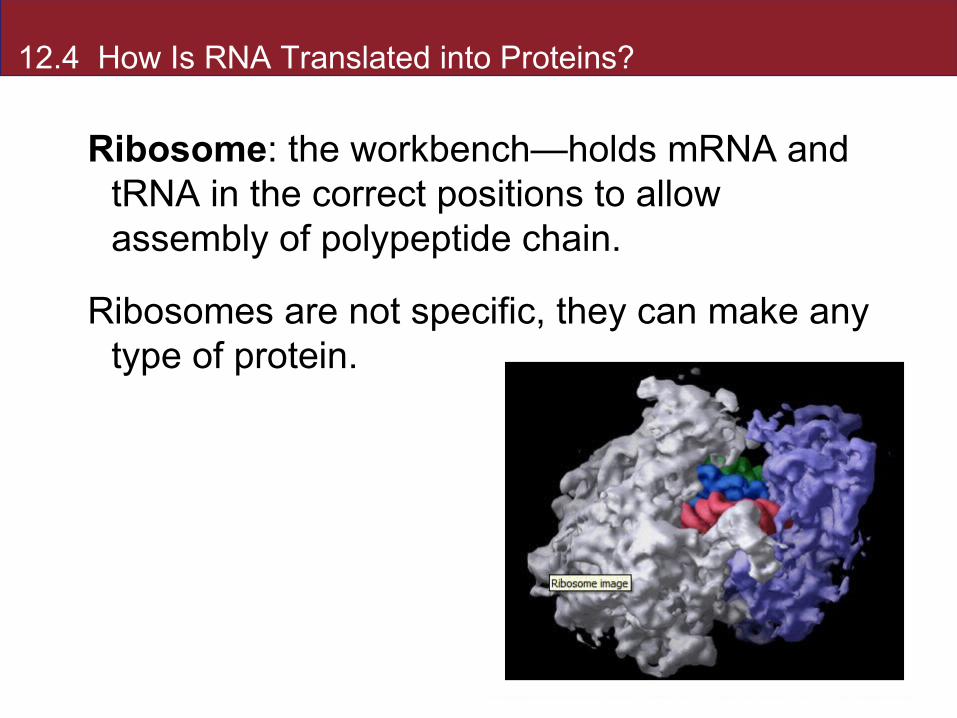
12.4 How Is RNA Translated into Proteins?
Ribosome: the workbench—holds mRNA and tRNA in the correct positions to allow assembly of polypeptide chain.
Ribosomes are not specific, they can make any type of protein.

ProkaryotesSmall Subunit 30s Large subunit 50s
16s 5s21 proteins 23s
34 proteins
Eukaryotes:Small 40S Large 60S
5S18S 28S33 proteins 5.8S
49 proteins
**The numbers are not additive – based on centrifugation rates

12.4 How Is RNA Translated into Proteins?
Subunits are held together by ionic and hydrophobic forces (not covalent bonds).
When not active in translation, the subunits exist separately.

Figure 12.10 Ribosome Structure

12.4 How Is RNA Translated into Proteins?
Large subunit has three tRNA binding sites:
• A site binds with anticodon of charged tRNA.
• P site is where tRNA adds its amino acid to the growing chain.
• E site is where tRNA sits before being released.

12.4 How Is RNA Translated into Proteins?
Hydrogen bonds form between the anticodon of tRNA and the codon of mRNA.
Small subunit rRNA validates the match—if hydrogen bonds have not formed between all three base pairs, it must be an incorrect match, and the tRNA is rejected.

12.4 How Is RNA Translated into Proteins?
Initiation:
An initiation complex forms—charged tRNA, small subunit, both bound to mRNA.
Recount process in paragraph form.Diagrams a bonus!

Figure 12.11 The Initiation of Translation (Part 2)
STEP 1
Step 2
http://www.biostudio.com/demo_freeman_protein_synthesis.htm

12.4 How Is RNA Translated into Proteins?
Start codon is AUG; first amino acid is always methionine, which may be removed after translation.
The large subunit joins the complex, the charged tRNA is now in the P site of the large subunit.

12.4 How Is RNA Translated into Proteins?
Elongation: the second charged tRNA enters the A site.
Large subunit catalyzes two reactions:
1. Breaks bond between tRNA in P site and its amino acid.
2. Peptide bond forms between that amino acid and the amino acid on tRNA in the A site.

Figure 12.12 The Elongation of Translation (Part 1)

Figure 12.12 The Elongation of Translation (Part 2)

12.4 How Is RNA Translated into Proteins?
Termination: translation ends when a stop codon enters the A site.
Stop codon binds a protein release factor—allows hydrolysis of bond between polypeptide chain and tRNA on the P site.
Polypeptide chain—C terminus is the last amino acid added.

Polypeptide chains are always designated N-terminus to C terminus
CORN backwards…
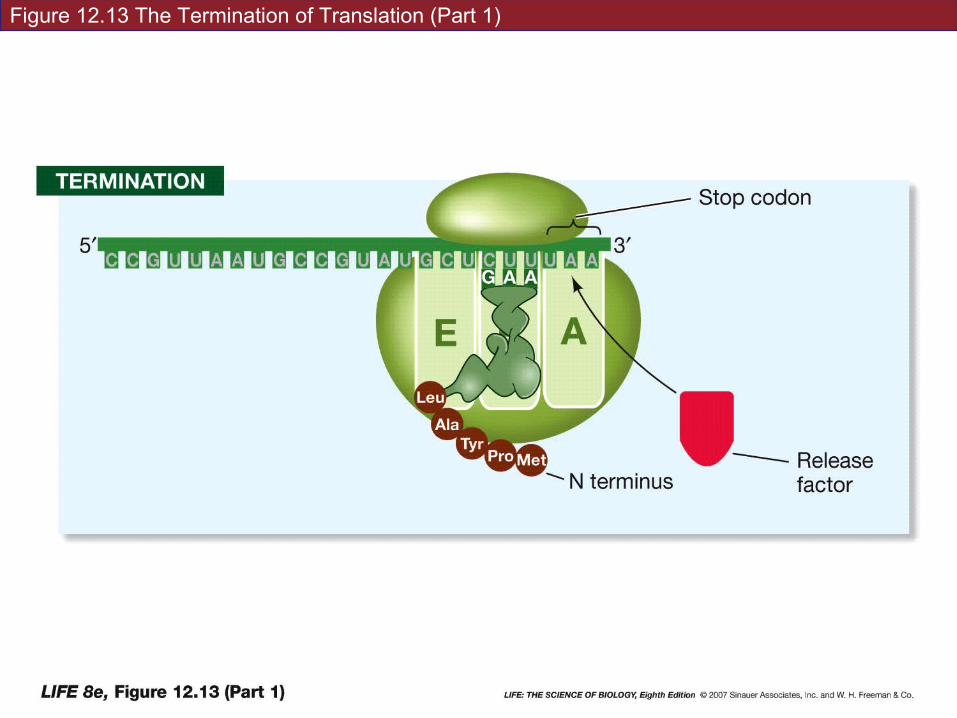
Figure 12.13 The Termination of Translation (Part 1)

Figure 12.13 The Termination of Translation (Part 2)
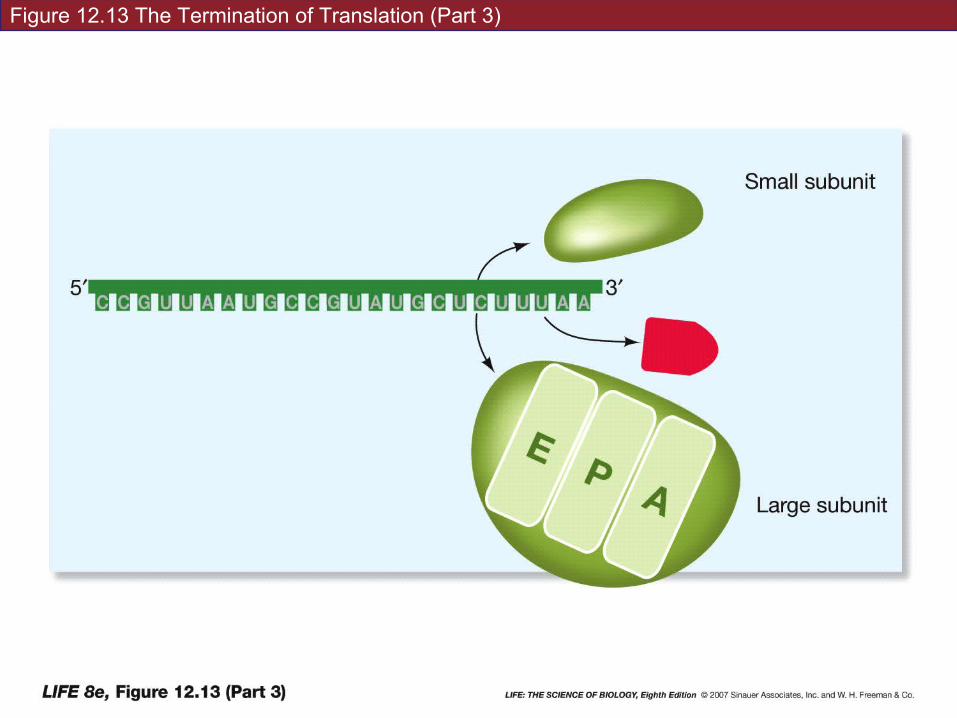
Figure 12.13 The Termination of Translation (Part 3)

Table 12.1

12.4 How Is RNA Translated into Proteins?
Several ribosomes can work together to translate the same mRNA, producing multiple copies of the polypeptide.
A strand of mRNA with associated ribosomes is called a polyribosome or polysome.
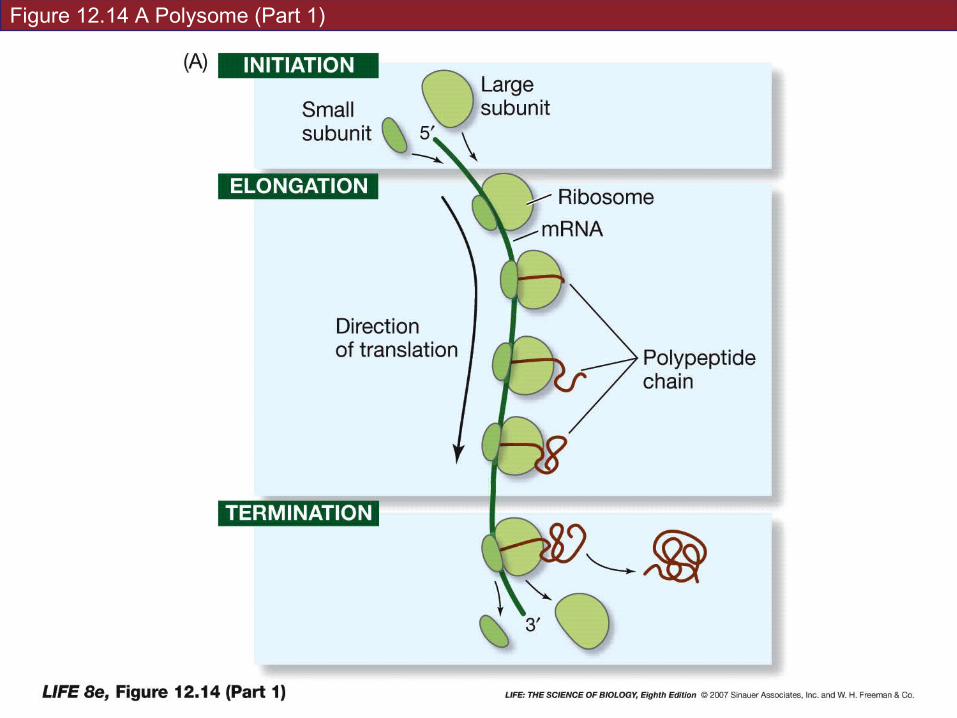
Figure 12.14 A Polysome (Part 1)

Figure 12.14 A Polysome (Part 2)

12.5 What Happens to Polypeptides after Translation?
1. Polypeptide folds as it emerges from the ribosome.
2. The amino acid sequence determines the pattern of folding.

Posttranslational aspects of protein synthesis:
• How is a message (mRNA) stabilized to get enough protein…at the right time?
1. 5’ cap added to N side of new protein
Stabilizing the message
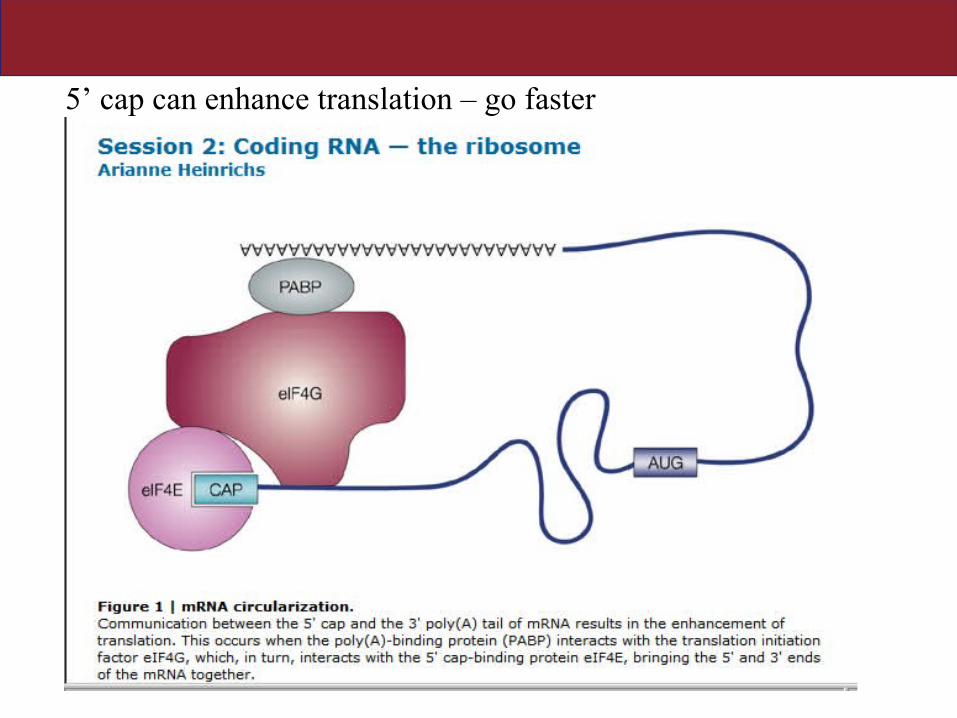
5’ cap can enhance translation – go faster

2. Polyadenylation is the synthesis of a poly(A) tail, a stretch of adenines at the end of the mRNA molecule.
At the end of transcription the last 3’ bit of the newly made RNA is cleaved off by a set aof enzymes. The enzymes then synthesize the poly(A) tail at the RNA's 3' end.
The poly(A) tail is important for the nuclear export, translation and stability of mRNA. The tail is shortened over time and when it is short enough, the mRNA is degraded. In a few cell types, mRNAs with short poly(A) tails are stored for later activation
Stabilizing the message

Processing the protein (product)
3. TARGETING:
Polypeptide may be moved from synthesis site to an organelle, or out of the cell.
Amino acid sequence also contains a signal sequence—an “address label.”
i.e. – proteins targeted to ER 5-10 hydrophobic amino acids on the N-terminus.

Figure 12.15 Destinations for Newly Translated Polypeptides in a Eukaryotic Cell

Figure 12.16 A Signal Sequence Moves a Polypeptide into the ER (Part 1)

Figure 12.16 A Signal Sequence Moves a Polypeptide into the ER (Part 2)
Folding chaperones (proteins) in RER fold proteins appropriately.
Mis-folding diseases:
Altzheimer’s
Creutzfeld–Jakob disease (CJD) (prion disease)
P53 – cancer from misfolded “watchdog”

12.5 What Happens to Polypeptides after Translation?
4. Glycosylation: addition of sugars to form glycoproteins
Sugars may be added in the Golgi apparatus—the resulting glycoproteins end up in the plasma membrane, lysosomes, or vacuoles.
Diseases: incorrect addition of sugars to specific amino acids – shows in infancy-almost always involves nervous system development.

• All proteins inserted into or associated with the cell membrane have sugars attached to them. They aid in recognition of other molecules.
• What would be some consequences of incorrect glycosylation at the cell membrane?

12.5 What Happens to Polypeptides after Translation?
Protein modifications:
5. Proteolysis: cutting the polypeptide chain by proteases. Degradation of protein message.
6. Phosphorylation: addition of phosphate groups by kinases. Charged phosphate groups change the conformation. Generally makes protein into enzymes!
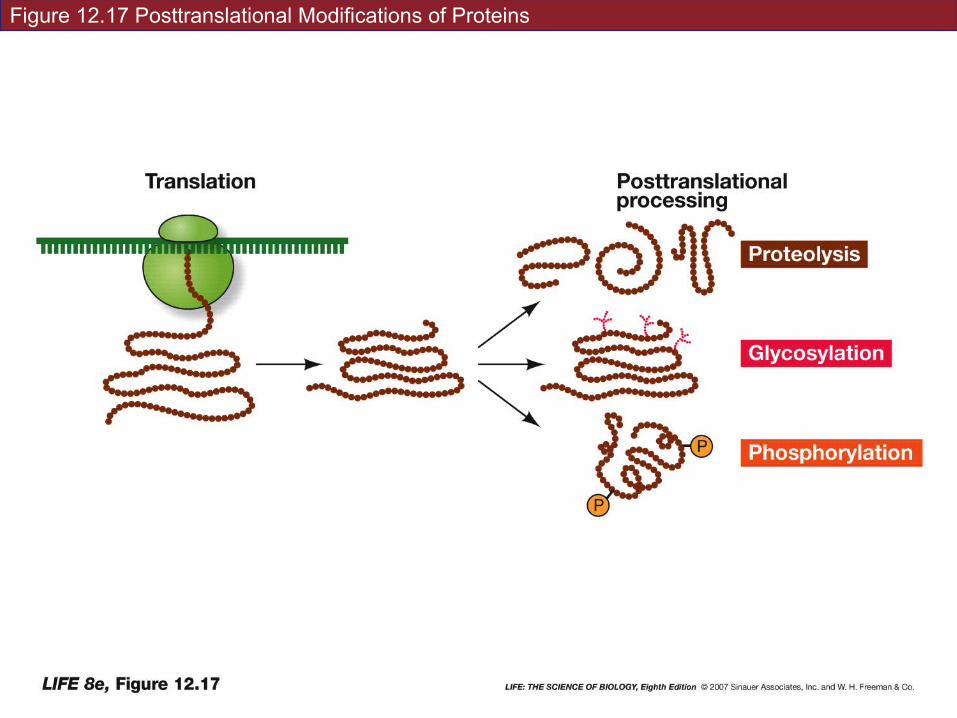
Figure 12.17 Posttranslational Modifications of Proteins


12.6 What Are Mutations?
Somatic mutations occur in somatic (body) cells. Mutation is passed to daughter cells, but not to sexually produced offspring.
Germ line mutations occur in cells that produce gametes. Can be passed to next generation.

12.6 What Are Mutations?
Conditional mutants: express phenotype only under restrictive conditions.
Example: the allele may code for an enzyme that is unstable at certain temperatures.

12.6 What Are Mutations?
All mutations are alterations of the nucleotide sequence.
Point mutations: change in a single base pair—loss, gain, or substitution of a base.
Chromosomal mutations: change in segments of DNA—loss, duplication, or rearrangement.

12.6 What Are Mutations?
Point mutations can result from replication and proofreading errors, or from environmental mutagens.
Silent mutations have no effect on the protein because of the redundancy of the genetic code.
Silent mutations result in genetic diversity not expressed as phenotype differences.
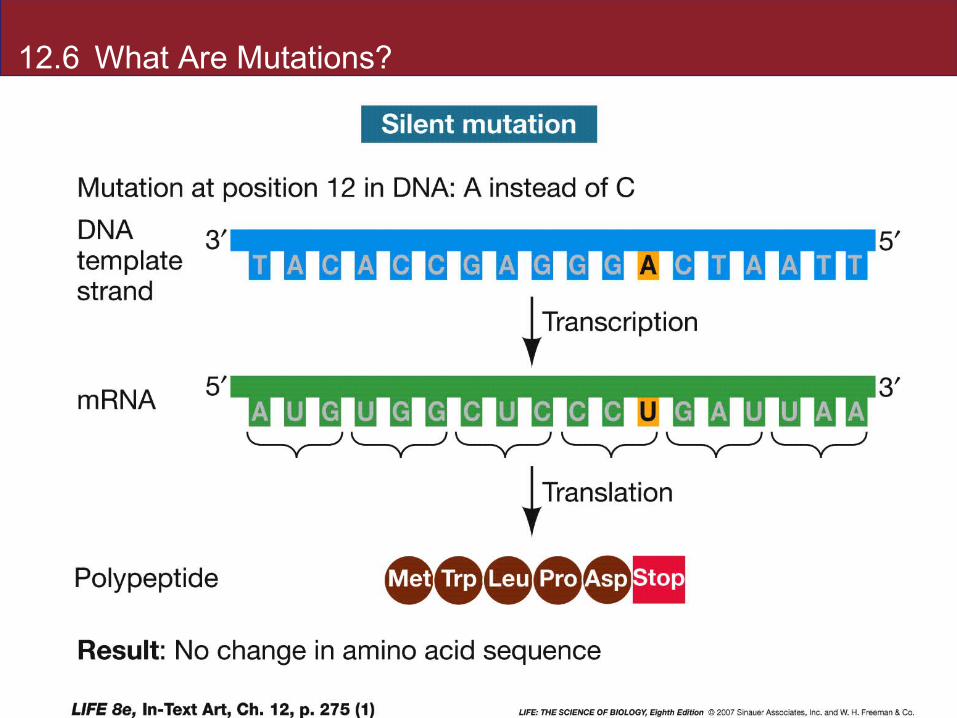
12.6 What Are Mutations?

12.6 What Are Mutations?
Missense mutations: base substitution results in amino acid substitution.

12.6 What Are Mutations?
Sickle allele for human â-globin is a missense mutation.
Sickle allele differs from normal by only one base—the polypeptide differs by only one amino acid.
Individuals that are homozygous have sickle-cell disease.

Figure 12.18 Sickled and Normal Red Blood Cells
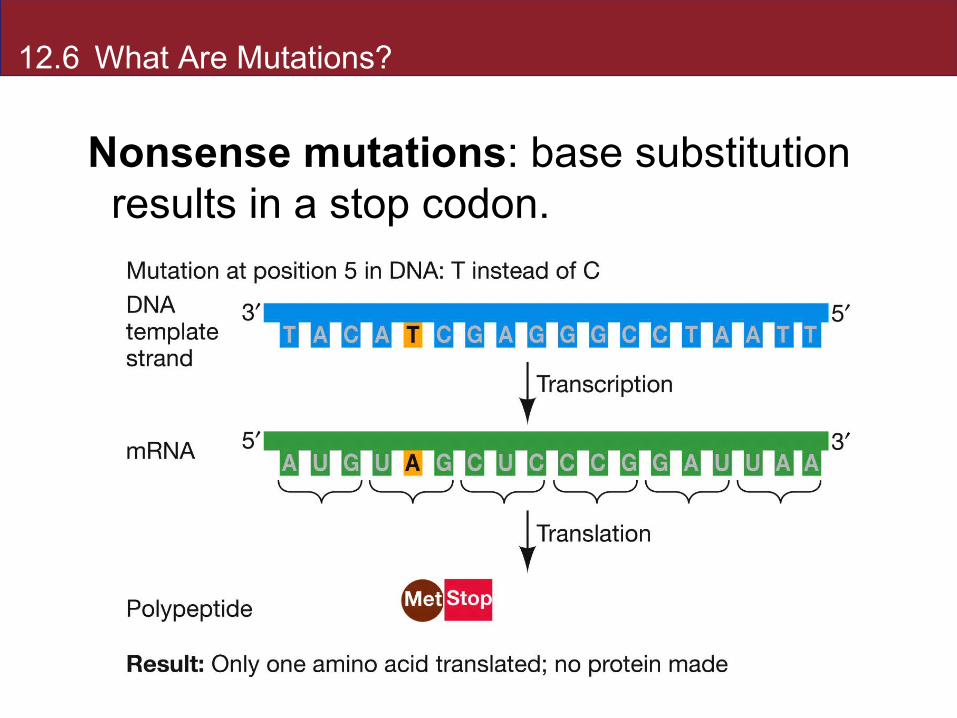
12.6 What Are Mutations?
Nonsense mutations: base substitution results in a stop codon.

12.6 What Are Mutations?
Frame-shift mutations: single bases inserted or deleted—usually leads to nonfunctional proteins.

12.6 What Are Mutations?
Chromosomal mutations:
Deletions—severe consequences unless it affects unnecessary genes or is masked by normal alleles.
Duplications—if homologous chromosomes break in different places and recombine with the wrong partners.

Figure 12.19 Chromosomal Mutations (A, B)

12.6 What Are Mutations?
Chromosomal mutations:
Inversions—breaking and rejoining, but segment is “flipped.”
Translocations—segment of DNA breaks off and is inserted into another chromosome. Can cause duplications and deletions. Meiosis can be prevented if chromosome pairing is impossible.

Figure 12.19 Chromosomal Mutations (C, D)

12.6 What Are Mutations?
Spontaneous mutations—occur with no outside influence. Several mechanisms:
• Bases can form tautomers—different forms; rare tautomer can pair with the wrong base.
• Chemical reactions may change bases (e.g., loss of amino group).

12.6 What Are Mutations?
• Replication errors—some escape detection and repair.
• Nondisjunction in meiosis.

12.6 What Are Mutations?
Induced mutation—due to an outside agent, a mutagen.
Chemicals can alter bases (e.g., nitrous acid can cause deamination).
Some chemicals add other groups to bases (e.g., benzpyrene adds a group to guanine and prevents base pairing). DNA polymerase will then add any base there.

12.6 What Are Mutations?
Ionizing radiation such as X-rays create free radicals—highly reactive—can change bases, break sugar phosphate bonds.
UV radiation is absorbed by thymine, causing it to form covalent bonds with adjacent nucleotides—disrupts DNA replication.

Figure 12.20 Spontaneous and Induced Mutations (Part 1)

Figure 12.20 Spontaneous and Induced Mutations (Part 2)

12.6 What Are Mutations?
Mutation provides the raw material for evolution in the form of genetic diversity.
Mutations can harm the organism, or be neutral.
Occasionally, a mutation can improve an organism’s adaptation to its environment, or become favorable as conditions change.

12.6 What Are Mutations?
Complex organisms tend to have more genes than simple organisms.
If whole genes are duplicated, the new genes would be surplus genetic information.
Extra copies could lead to the production of new proteins.
New genes can also arise from transposable elements (see Chapters 13 and 14).