profoundalteration aj3 t-cell polymorphism region 13 - pnas · vol. 88, pp. 10267-10271,...
TRANSCRIPT

Proc. Nati. Acad. Sci. USAVol. 88, pp. 10267-10271, November 1991Immunology
Profound alteration in an aj3 T-cell antigen receptor repertoire dueto polymorphism in the first complementarity-determining regionof the 13 chain
(aflelic polymorphism/T-ceil antigen recognition)
SARA-JO GAHM*t, B. J. FOWLKESt, STEPHEN C. JAMESON§, NICHOLAS R. J. GASCOIGNE§,MELISSA M. COTTERMAN*, OSAMI KANAGAWA¶, RONALD H. SCHWARTZt, AND Louis A. MATIS**Biological Response Modifiers Program, Frederick Cancer Research and Development Center, National Cancer Institute, National Institutes of Health,Frederick, MD 21702; tHoward Hughes Medical Institute-National Institutes of Health Research Scholars Program and tLaboratory of Cellular and MolecularImmunology, National Institute of Allergy and Infectious Disease, National Institutes of Health, Bethesda, MD 20892; §Department of Immunology, ResearchInstitute of Scripps Clinic, La Jolla, CA 92037; and 1Department of Pathology, Washington University School of Medicine, St. Louis, MO 63110
Communicated by William E. Paul, August 13, 1991
ABSTRACT Amino acid residues that are critical in main-taining the framework structure ofimmunoglobulin heavy- andlight-chain variable (V) regions are strongly conserved in theVa and Vfi proteins of the a.8 T-cell antigen receptor(TCRaB). Consequently, it has been proposed thatTCRaB hasa conformation similar to that of an immunoglobulin Fabfragment and that the regions of the TCR homologous to thethree immunoglobulin complementarity-determining regions(CDRs 1, 2, and 3) bind to the peptide antigen-ajor histo-compatibility complex (MHC) molecule ligand. A single aminoacid substitution in the predicted CDR1 of the Vj33 protein ofcertain mouse strains dramatically altered TCRafi usage in anantigen-specific MHC-restricted immune response but did notabrogate Vfi3 specificity for the superantigens minor lympho-cyte stimulatory locus (Mls)C and staphylococcal enterotoxin A(SEA). The results confirm the importance of the Vfi CDR1 inantigen-MHC molecule recognition, supporting the Fab-likestructural model of TCRaet., and provide further evidence thatconventional antigen-MHC recognition and superantigen rec-ognition are mediated by distinct regions of the TCR ,B chain.They also suggest that allelic polymorphism may be a signifi-cant source of diversity in the TCR repertoire.
The T-cell antigen receptor (TCR) ap3 heterodimer recognizescomplexes of peptide antigens in association with majorhistocompatibility complex (MHC) molecules (1-4). Theconformational structure of the TCR has not yet been deter-mined. However, examination of TCR variable a and P (Vaand VP) sequences has revealed extensive homology to thoseimmunoglobulin heavy- and light-chain V residues that arecritical in maintaining the framework structure of the Vdomains (5-10). X-ray crystallographic analysis of immuno-globulin-antigen complexes has shown antigen binding to bemediated primarily by amino acid residues within threehypervariable complementarity-determining regions (CDRs)of heavy- and light-chain V regions (11, 12). Thus, severalgroups have proposed that TCR Va-VB dimers have aconformation similar to that of immunoglobulin Fab frag-ments and that regions of the TCR Va and VB proteins inhomologous positions to the three immunoglobulin CDRs willhave a predominant role in binding to the antigen-MHCmolecule ligand (8-10). Consistent with this model, V, di-versity, joining, and junctionally encoded sequences of theTCR a and p chains, corresponding to the putative CDR3s,can affect TCRa,8 specificity (13-17). Also, mutation of twoamino acids in the predicted CDR1 of the Va chain of an
antigen-specific TCRaI3 resulted in impaired antigen-MHCmolecule recognition (18).
Structural and functional studies in well-defined antigenresponses that have correlated TCR expression with speci-ficity have shown that all the V elements of the TCR may beselected and, therefore, contribute to antigen-MHC mole-cule recognition (1). For example, TCR usage in the murineimmune response to the protein antigen pigeon cytochromec (cyt) has been extensively studied (13, 14, 19, 20). In theC57BL/10 (B10) mouse strain background, most class IIMHC Ek-restricted cyt-specific T cells express Vall pairedwith Vf33 V elements (13, 19, 20). However, in preliminaryanalysis of TCR usage in cyt-specific Ek-restricted T cellsfrom C57BR (H-2k) mice, we did not observe consistentlyhigh levels of expression of both Vall and V/33 (21, 22).
In the current study, we determine that the strain-dependent differences in TCR usage in the C57BR cyt-specific response can be attributed to an allelic polymorphismin the V(33 protein encoding a single amino acid substitutionin the putative CDR1. This sequence polymorphism does notalter VP/3 specificity for the superantigens minor lymphocytestimulatory locus (Mls)c or staphylococcal enterotoxin A(SEA). These results provide independent support for anFab-like structure of the TCR and may have implications formodels of T-cell-mediated autoimmunity.
MATERIALS AND METHODSAnimals. B1O.BR, C57BR, and B10.A mice were pur-
chased from The Jackson Laboratory. (B10.BR x C57BR)F1mice were bred in our facility. To develop the B1O.A(TBL)/AI strain, B10.A/AI (VI3b) mice were bred withC57L/N (V3Ba) mice. The resulting F1 mice were back-crossed to B10.A and the offspring were typed for heterozy-gosity at the VP3 locus using the monoclonal antibody (mAb)KJ16, specific for Vp8.1,2 of B10.A. Heterozygotes wereback-crossed to B10.A for 11 more generations and then werebrother-sister mated to derive the inbred strain homozygousfor the V/3 locus (negative for KJ16). This strain is availableupon request from R.H.S.
Cell Lines and Antibodies. The generation of antigen-specific T-cell lines has been described (13, 19, 20). To purifyantigen-specific T-cell lines for cells expressing VB3 or Vall,flasks were coated with appropriate goat anti-hamster immu-noglobulin, goat anti-rat immunoglobulin, and goat anti-
Abbreviations: TCR, T-cell receptor; CDR, complementarity-determining region; mAb, monoclonal antibody; MHC, major his-tocompatibility complex; Mls, minor lymphocyte stimulatory locus;SEA, staphylococcal enterotoxin A; cyt, pigeon cytochrome c; V,variable; APC, antigen-presenting cell.
10267
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

10268 Immunology: Gahm et al.
mouse immunoglobulin antibodies (Caltag, South San Fran-cisco, CA) at 20 ,gg/ml in phosphate-buffered saline andincubated overnight at 40C. Nonadherent antibody was re-moved and the flasks were washed twice. Vall-specific orV,8-specific mAbs in 5% (vol/vol) fetal calfserum were addedand incubated overnight at 40C. Unbound mAbs were re-moved and the flasks were washed twice. Approximately 5 x106 cultured cyt-specific T cells were added to the flasks andincubated for 30 min at 370C with gentle shaking every 5 min.Nonadherent cells were removed and 1.5 x 107 irradiated(2000 R; 1 R = 0.258 mC/kg) syngeneic spleen cells wereadded. After 24 hr of culture in 10 ml of complete medium,the cells were harvested, washed twice, and resuspended incomplete medium supplemented with recombinant interleu-kin 2 (Cetus; 100 units/ml). Four to five days later the cellswere assayed for Vall and VP3 receptor expression byimmunofluorescence staining and flow cytometry. To exam-ine TCR usage in unimmunized spleen-cell populations,spleen-cell suspensions were enriched for CD4+ T cells byincubation with CD8-specific mAbs and complement andthen V.33 cells were purified by activation in the presenceof interleukin 2 after adherence to flasks coated with VB-specific mAb (KJ25). The percentages of V/33+ CD4+ andVall+ CD4+ cells in each population were determined byflow cytometry. TCR expression was analyzed in this fashionbecause steric inhibition by the mAbs made it difficult tosimultaneously examine Va and V13 expression on individualcells. RR8-1 is a Vall-specific rat mAb derived by immuni-
AVB3 Selected
T Cell Line
B10.BR
C57BR
10u 10
t.A...
n~~~~~~~~~~~~~~~~10V 1V0104 10° 1 102 103 1 4
VB3 Vcll I
B C
12-i
0)
0
0.
BR 1 BR 2 BR 3 BR I BR 2 BR 3 C571 C572 C573<V.33 Selected Vcul1 Selected V33 Selected
Proc. Nadl. Acad. Sci. USA 88 (1991)
zation with a Vall.l+ T-cell hybridoma and is specific forV-element-encoded proteins of the Vall family. It has beendetermined that RR8-1 recognizes Vall.1 and Vall.2 but notVall.3 proteins (S.C.J., O.K., and N.R.J.G., unpublisheddata). KJ25 is a V,83-specific mAb (23).Flow Cytometry. Cells were stained with anti-receptor
antibodies KJ25 or RR8-1, which were detected with anappropriate fluorescein isothiocyanate (FITC)-conjugatedsecondary reagent [for KJ25, goat anti-hamster immunoglob-ulin (Caltag); for RR8-1, goat anti-rat immunoglobulin(Kirkegaard and Perry Laboratories, Gaithersburg, MD)].Control staining was performed with FITC-conjugated anti-body alone. Cells were analyzed on either the FACS 440 orthe FACScan (Becton Dickinson). All the lines were deter-mined to be 100%oTCR a,B+ CD4+. The percentages ofVall+and V(33 cells in each line are shown in the figures. Dataobtained from FACScan are based upon analysis of 10,000events and from the FACS 440 are based upon 50,000 eventsanalyzed.
Proliferation Assays. Antigen-specific Vp3' T cells (2 x l04cells per well) were placed in 96-well flat-bottom plates withT-cell-depleted syngeneic (BlO.BR or C57BR) antigen-presenting cells (APCs) (5 x l0O cells per well) plus mediumalone, cyt, or SEA or with (Mls)c-bearing C3H/He APCs (5x 105 cells per well). The APCs were treated with mitomycinC (50 ,g/ml). Cultures were pulse-labeled with [3H]thymi-dine (1 ,uCi; 1 Ci = 37 GBq) after 48 hr and harvested 18 hrlater. Data (cpm) represent the mean of triplicate cultures.
Va 1 Selected
Vail
D
J.l
, t | 1k :
V;,11 Selected
FIG. 1. Vall and VB3 expression in cyt-specific T-cell lines from B1O.BR, C57BR, and (BlO.BR x C57BR)F1 mice. Independentantigen-specific T-cell lines (e.g., BR1, BR2, and BR3 generated from B1O.BR mice; and C571, C572, and C573 generated from C57BR mice)were generated as described (13, 19, 20). After four rounds of in vitro stimulation with cyt and syngeneic APCs, the T-cell lines consisted entirelyof antigen-specific cells (13, 19-22). Each line was then purified for the expression of either Vall or VB3 and examined for Va and V'S usage.(A) Representative FACScan histograms obtained during the course of mAb (V,83, KJ25; Vall, RR8-1) selection of cyt-specific lines BR2 andC572. TCR expression in mAb-selected cyt-specific T-cell lines from B1O.BR (B), C57BR (C), and (BlO.BR x C57BR)F1 (D) mice. The lineswere propagated and the fluorescence analysis was performed. The percentage of VB3- and Vall-expressing T cells in each line is shown.
- 10'.. 102100 lo, 102 10 104
1 i 0--0- i0 I10° lo, 102 13 14 1( )0 10 102 103 1
,"I. -in 4^1 -0 . _,I 1-1-1

Proc. Natl. Acad. Sci. USA 88 (1991) 10269
DNA Sequencing. To sequence the V,83 alleles from C57BRand B10.A(T3L) mice and the Vall.1 allele from C57BRmice, cDNAs were generated from mRNA derived fromcyt-specific V,83' or Vall' T-cell lines by the PCR usingappropriate upstream 5' and complementary constant region3' oligonucleotide primers. PCR-generated products were
cloned into either the pGEM or the pCR2000 (Invitrogen, SanDiego) vectors and then sequenced using the dideoxynucle-otide chain-termination method.
RESULTS
To examine TCR usage in the cyt-specific Ek-restrictedimmune response, we derived several independent Ek_restricted cyt-specific T-cell lines from H-2k-matchedB10.BR and C57BR mice. All of these lines and clonesderived therefrom by limiting dilution recognized the pre-dominant COOH-terminal epitope on cyt and were Ek_restricted (refs. 21 and 22; S.-J.G. and L.A.M., unpublisheddata). Separate Vall- and V,83-enriched populations were
derived from each line by activation with V-region-specificmAbs and these purified populations were analyzed by flowcytometry for expression of Vall and V,33. Prior to fluores-cence analysis, the mAb-purified populations were retestedand confirmed to be antigen-specific and Ek-restricted (S.-J.G. and L.A.M., unpublished data). Representative fluores-cence histograms are shown in Fig. 1A and the data fromanalysis of all the T-cell lines are summarized in Fig. 1 B-D.KJ25 and RR8-1 stained V/833 T cells and Vall' T cellsrespectively, with equal intensity from B1O.BR and C57BRmice (Fig. 1A). The Vf33-purified cyt-specific B10.BR T cellswere uniformly almost 100%o Vall+ and, conversely, theVall+ T cells were 60-70% Vp3' (Fig. 1). In contrast, whenthe cyt-specific lines from C57BR mice were examined inidentical fashion, both Vall+ and Vp3' T cells were ob-served, but very few of the cells coexpressed Vall and V/3(Fig. 1). Vall-V,83 dimers predominated in a cyt-specificT-cell line from (BlO.BR x C57BR)F1 mice (Fig. 1D), rulingout self-tolerance as the basis for the failure to express
Vall-V,83 pairs in the C57BR cyt-specific response.Moreover, we determined that the failure of C57BR cyt-
specific T cells coordinately to express Vall and Vf3 couldnot be attributed to structural or thymic selectional con-straints on the pairing of Vall and V133. Flow cytometricanalysis of splenic T-cell populations from unimmunizedB10.BR and C57BR mice enriched for VP3+ cells by mAbstimulation showed that Vp3+ CD4+ T cells from both strainswere 9-10% Vall+ (Fig. 2). Thus, Vall-V,33 heterodimerswere observed with equal frequency among non-antigen-specific B1O.BR and C57BR T-cell populations. These dataalso show that RR8-1 detects Vall+-VB3+ heterodimersfrom B1O.BR and C57BR mice with the same efficiency.We next considered the possibility that allelic polymor-
phism in the Vall and/or Vf33 gene families could accountfor the differential receptor usage in the antigen-specificresponse. The Vall families in mice with C57 and B10backgrounds are identical by restriction fragment lengthpolymorphism (22, 24) and serological (ref. 25; S.C.J., O.K.,and N.R.J.G., unpublished data) analyses. Moreover, wehave cloned the Vall gene expressed by Vall+ cyt-specificC57BR T cells (M.M.C., S.-J.G., and L.A.M., unpublisheddata) and found that its sequence is identical to that of theVall.1 allele (13) of B10 background mice. Thus, Vallpolymorphism cannot account for the altered receptorexpression. However, the TCR Vl haplotypes of C57 (Vf.?)and B10 (Vpb) background mice are distinct (24, 26). Recentsequence analysis of the Vf33a gene segment from SWR/Jmice revealed a single base-pair difference in V/ 3a relative tothe VP3b allele, encoding an amino acid substitution ofphenylalanine (VP3a) for valine (V/33b) at residue 31 of the
T Cells
1310.13R l
Kli2
9.4%C57BR
V,33 Va 1FIG. 2. Equal percentages of Va1ll-V,33+TCR dimers are found
among non-antigen-selected peripheral CD4' T-cell populations inB10.BR and C57BR mice. CD4' TCRaI3+ splenic T cells were purifiedfor V,83+ cells and then examined for expression of Vall. Greaterthan 95% of the CD4' T cells are Vf33. Two-color fluorimetry wasperformed for analysis of V(33 (KJ25) vs. CD4 and of Vall (RR8-1)vs. CD4. The percentages of V,833CD4' cells that are Vall' areshown for each mouse strain. The cells were examined on a FACS 440and the percentages are based upon analysis of 50,000 events.
Vf33 protein (26). We have confirmed that the V/33a genesegment from C57BR mice has an identical sequence (S.-J.G.and L.A.M., unpublished data).To determine whether this single amino acid substitution in
the V,83 chain was responsible for the differential receptorusage, we examined cyt-specific TCR expression in a newlyconstructed TCR VP3 congenic inbred mouse strain, theBlO.A(TPL)/AI [BlO.A(TPL)]. In this strain the TCR VI3ahaplotype from the C57L mouse has been transferred ontothe BlO.A background. Thus, with the exception of the TCR,3locus and closely linked genes, B10.A (VP33b) andBlO.A(TQ3L)(Vp3a) mice are genetically identical. Ek_restricted cyt-specific T-cell lines were generated from im-munized BlO.A and BlO.A(TPL) mice. They were selected asbefore with receptor-specific mAbs and examined for thecoexpression of Vall and V83. V,833 cyt-specific T cellsfrom BlO.A mice were virtually all Vall+, and the majority(69%) of the Vall+ subset were VB3+ (Fig. 3A). In contrast,among the BlO.A(TpL) cyt-specific T cells, there was little orno coordinate expression of Vall and VP3 (Fig. 3B). We alsosequenced the V83 allele from Vf33 cyt-specificBlO.A(Tf3L) T cells, and its DNA sequence was identical tothat of the SWR/J and the C57BR (ref. 26; S.-J.G. andL.A.M., unpublished data). Also, by restriction fragmentlength polymorphism and sequence analyses (ref. 24; L. R.Smith, personal communication and R. K. Barth, personalcommunication), there is no polymorphism among the Jf3segments of the Vpa and Vab haplotypes. This includes theJ/31.2 and JP2.5 segments (L. R. Smith, personal communi-cation), the most commonly expressed Jf3 elements in V/33+-Vall+ cyt-specific TCRs (13, 19, 20). Thus, these dataindicate that the Val-31 -* Phe substitution in the V33 proteinabolishes V63-Vall heterodimer usage in the Ek-restrictedcyt-specific T-cell response.We also asked whether the Val-31-+ Phe substitution
would abrogate V.83-mediated superantigen recognition.Most Vf33+ TCRs recognize (Mls)c antigens and SEA (21, 23,27-29). The proliferation data in Table 1 show that both VP33a-
Immunology: Gahm et al.
Proc. Natl. Acad. Sci. USA 88 (1991)
/.i3 elected V 1 1 e: e 1
0'CD
hi
Ad;;:::..:
.:.
.::.
:.::..
::...
..:. .:.
16;ID
I
L:I V[II \ 3
Table 1. Superantigen-specific proliferation of V93+ and Vf33b+pigeon cyt-specific T-cell lines
[3H]Thymidineincorporation, cpm
Antigen B1O.BR C57BR
- 2,389 737Cyt (2 yg/ml) 26,618 12,153SEA (1 pg/ml) 26,646 17,545Mlsc (C3H/He APC) 29,225 9,211
V03' subpopulations from the antigen-specific B1O.BR andC57BR T-cell lines (Fig. 1) were examined for responsiveness topigeon cyt, SEA, and Mlsc.
.,c
-7,
V/)3 selected Vu 11 selectec
FIG. 3. Vall and V,83 expression in mAb-selected cyt-specificT-cell lines from B1O.A (A) and B1O.A(TI3L) (B) mice. Antigen-specific mAb-selected lines were prepared and examined for V,83and Vall expression as described in Fig. 1.
and Vp3b-bearing T-cell populations recognize SEA and(Mls)c antigens expressed on C3H/He spleen cells.To interpret these results in the context of the proposed
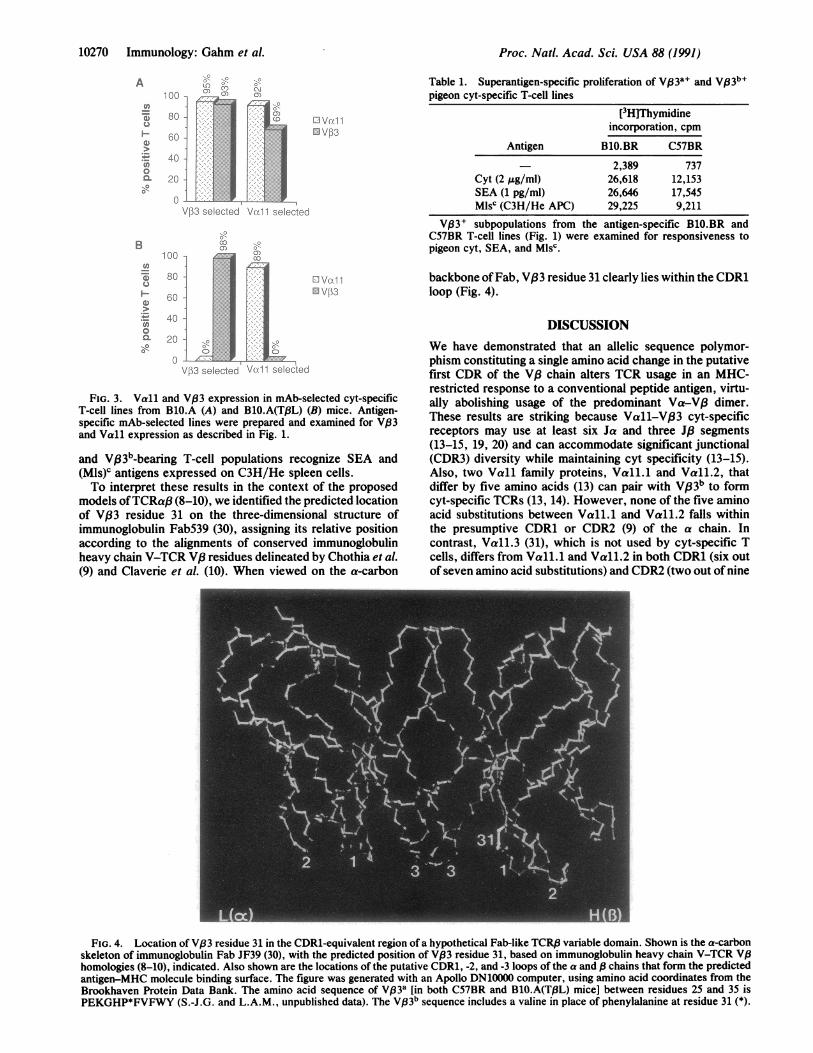
models ofTCRaj3 (8-10), we identified the predicted locationof V,33 residue 31 on the three-dimensional structure ofimmunoglobulin Fab539 (30), assigning its relative positionaccording to the alignments of conserved immunoglobulinheavy chain V-TCR V.8 residues delineated by Chothia et al.(9) and Claverie et al. (10). When viewed on the a-carbon
backbone ofFab, V,83 residue 31 clearly lies within the CDR1loop (Fig. 4).
DISCUSSIONWe have demonstrated that an allelic sequence polymor-phism constituting a single amino acid change in the putativefirst CDR of the V,3 chain alters TCR usage in an MHC-restricted response to a conventional peptide antigen, virtu-ally abolishing usage of the predominant Va-Va dimer.These results are striking because Vall-V133 cyt-specificreceptors may use at least six Ja and three Jf3 segments(13-15, 19, 20) and can accommodate significant junctional(CDR3) diversity while maintaining cyt specificity (13-15).Also, two Vall family proteins, Vall.1 and Vall.2, thatdiffer by five amino acids (13) can pair with V,83b to formcyt-specific TCRs (13, 14). However, none of the five aminoacid substitutions between Vall.1 and Vall.2 falls withinthe presumptive CDR1 or CDR2 (9) of the a chain. Incontrast, Vall.3 (31), which is not used by cyt-specific Tcells, differs from Vall.1 and Vall.2 in both CDR1 (six outof seven amino acid substitutions) and CDR2 (two out of nine
FIG. 4. Location of V,33 residue 31 in the CDR1-equivalent region of a hypothetical Fab-like TCRI3 variable domain. Shown is the a-carbonskeleton of immunoglobulin Fab JF39 (30), with the predicted position of V,33 residue 31, based on immunoglobulin heavy chain V-TCR V,8homologies (8-10), indicated. Also shown are the locations of the putative CDR1, -2, and -3 loops of the a and /3 chains that form the predictedantigen-MHC molecule binding surface. The figure was generated with an Apollo DN10000 computer, using amino acid coordinates from theBrookhaven Protein Data Bank. The amino acid sequence of Vp3a [in both C57BR and BlO.A(T,8L) mice] between residues 25 and 35 isPEKGHP*FVFWY (S.-J.G. and L.A.M., unpublished data). The V,33b sequence includes a valine in place of phenylalanine at residue 31 (*).
A
(I,
C.D
w
0an
1 "DO
80 1
60
40 -
Vo1V1
B1 00
U)
.)
40-U)0s:- 20C-
-a
LCI '-'-:0.C-1 GI)
-1
A
10270 Immunology: Gahm et al.

Proc. Natl. Acad. Sci. USA 88 (1991) 10271
substitutions). Thus, the first and second CDRs of the Vallchain are also likely to be important for cyt-Ek recognition.
In the proposed model of the TCR-ligand complex, theCDR3 regions of Va and V,3 are predicted directly to contactthe peptide within the binding cleft of the MHC molecule,while the regions corresponding to CDR1 and CDR2 overlieand thus interact predominantly with upward-facing residueson the MHC a-helices (8-10). Thus, the larger size of thearomatic-ring-containing side chain of phenylalanine relativeto valine could change the conformation of the CDR1 loopand alter the interaction of VP33 with the Ek molecule.However, because V,63 a can form Ek-restricted cyt-specificTCR when paired with Va chains other than Vall (Figs. 1and 3), the Val-31 -* Phe substitution must also affect theV,83-Vall interaction, either indirectly due to allostericchanges in VP3 or perhaps directly through interaction ofVf33 CDR1 residues with CDR3 residues of the Vall chain(Fig. 4). As a result, the presence of Phe-31 might impairEk-restricted cyt-specific recognition by all possible V,83a-Vall dimers. Alternatively, the subset of V33a-Vall recep-tors with potential cyt specificity might fail to be selectedduring intrathymic development. Transfection analysis ofcyt-specific VB3-Vall TCRs mutated at V,33 residue 31should address these possibilities. The predominant usage ofVall-Vj33 dimers by (BlO.BR x C57BR)F1 cyt-specific Tcells (Fig. 1D) mitigates against the possibility that VB33a pairswith an alternate Va to form higher-affinity TCRs thatpreferentially would be selected over Vall-V,83 dimers.
Recently, amino acid residues on TCR V/3 chains that areimportant in determining superantigen specificity were iden-tified (32, 33). They are predicted to lie on a loop of the V,3protein away from the immunoglobulin-like CDRs (5). Wehave shown that a CDR1 polymorphism that dramaticallyinfluences TCR usage in an MHC-restricted antigen responsedid not abrogate superantigen recognition. Thus, our resultsprovide additional evidence that conventional antigen-MHCrecognition and superantigen recognition are mediated bydistinct regions of the TCR.
Finally, our data could have relevance for T-cell-mediatedautoimmunity. Human TCR V regions exhibit significantpolymorphism, and restriction fragment length polymor-phism analyses have suggested disease associations withTCR alleles (34, 35). Family studies of multiple sclerosis havedemonstrated a disease susceptibility gene in the TCR / locus(36). Here we have shown that allelic polymorphism of TCRV regions can have a major effect on antigen recognition andTCR repertoire selection. Thus, challenge with foreign anti-gen (e.g., viruses or bacteria) could activate distinct popu-lations of receptor-bearing T cells in individuals expressingvarious TCR V region alleles. These populations might varyin their level of cross reactivity to self antigens and, there-fore, in their propensity to mediate autoimmune disease.
We thank J. A. Bluestone, S. M. Hedrick, and F. Ramsdell forinsightful discussions; R. Venable and R. Pastor for their help ingenerating Fig. 4; L. R. Smith and R. K. Barth for communicatingunpublished data regarding J,1 polymorphism; J. Ashwell and R. N.Germain for critical reading of the manuscript; and C. Fogle and L.Ridgell for manuscript preparation. S.C.J. is a fellow of the CancerResearch Institute. N.R.J.G. is a Scholar of the Leukemia Society ofAmerica and is supported by grants from the National Institutes ofHealth (GM-39476) and the Concern Foundation for Cancer Re-search.
1. Matis, L. A. (1990) Annu. Rev. Immunol. 8, 65-82.2. Nikolic-Zugic, J. & Bevan, M. J. (1990) Nature (London) 344,
65-67.
3. Berg, L. J., Frank, G. D. & Davis, M. M. (1990) Cell 60,1043-1053.
4. Sha, W. C., Nelson, C. A., Newberry, R. D., Pullen, J. K.,Pease, L. R., Russell, J. H. & Loh, D. Y. (1990) Proc. Natl.Acad. Sci. USA 87, 6186-6190.
5. Becker, D. M., Patten, P., Chien, Y.-H., Yokota, T., Eshhar,Z., Giedlin, M., Gascoigne, N. R. J., Goodnow, C., Wolf, R.,Arai, K.-I. & Davis, M. M. (1985) Nature (London) 317,430-434.
6. Barth, R. K., Kim, B. S., Lan, N. C., Hunkapiller, T., So-bieck, N., Winoto, A., Gershenfeld, H., Okada, C., Hansburg,D., Weissman, I. L. & Hood, L. (1985) Nature (London) 316,517-523.
7. Novotny, J., Tonegawa, S., Saito, H., Kranz, D. M. & Eisen,H. N. (1986) Proc. Natl. Acad. Sci. USA 84, 742-746.
8. Davis, M. M. & Bjorkman, P. J. (1988) Nature (London) 334,395-402.
9. Chothia, C., Boswell, D. R. & Lesk, A. M. (1988) EMBO J. 7,3745-3755.
10. Claverie, J.-M., Prochnicka-Chalufour, A. & Bougueleret, L.(1989) Immunol. Today 10, 10-14.
11. Amit, A. G., Mariuzza, R. A., Phillips, S. E. V. & Poljak,R. J. (1986) Science 233, 747-753.
12. Sheriff, S., Silverton, E. W., Padlan, E. A., Cohen, G. H.,Smith-Gill, S. J., Finzel, B. C. & Davies, D. R. (1987) Proc.Natl. Acad. Sci. USA 84, 8075-8079.
13. Fink, P. J., Matis, L. A., McElligott, D. L., Bookman, M. &Hedrick, S. M. (1986) Nature (London) 321, 219-225.
14. Winoto, A., Urban, J. L., Lan, N. C., Goverman, J., Hood, L.& Hansburg, D. (1986) Nature (London) 324, 679-682.
15. Hedrick, S. M., Engel, I., McElligott, D. L., Fink, P. J., Hsu,M.-L., Hansburg, D. & Matis, L. A. (1988) Science 239,1541-1544.
16. Engel, I. & Hedrick, S. M. (1988) Cell 54, 473-484.17. Danska, J. S., Livingstone, A. M., Paragas, V., Ishihara, T. &
Fathman, C. G. (1990) J. Exp. Med. 172, 27-33.18. Nalefski, E. A., Wong, J. G. P. & Rao, A. (1990) J. Biol.
Chem. 265, 8842-8846.19. Sorger, S. B., Hedrick, S. M., Fink, P. J., Bookman, M. A. &
Matis, L. A. (1987) J. Exp. Med. 165, 279-301.20. Matis, L. A., Sorger, S. B., McElligott, D. L., Fink, P. J. &
Hedrick, S. M. (1987) Cell 51, 59-69.21. Fry, A. M. & Matis, L. A. (1988) Nature (London) 335, 830-
832.22. Fry, A. M., Cotterman, M. M. & Matis, L. A. (1989) J. Im-
munol. 143, 2723-2729.23. Pullen, A. M., Marrack, P. & Kappler, J. W. (1988) Nature
(London) 335, 796-801.24. Klotz, J. L., Barth, R. K., Kiser, G. L., Hood, L. E. &
Kronenberg, M. (1989) Immunogenetics 29, 191-201.25. Jameson, S. C., Kaye, J. & Gascoigne, N. R. J. (1990) J.
Immunol. 145, 1324-1331.26. Smith, L. R., Plaza, A., Singer, P. A. & Theofilopoulos, A. N.
(1990) J. Immunol. 144, 3234-3237.27. Abe, R., Vacchio, M. S., Fox, B. & Hodes, R. J. (1988) Nature
(London) 335, 827-830.28. White, J., Herman, A., Pullen, A. M., Kubo, R., Kappler,
J. W. & Marrack, P. (1989) Cell 56, 27-35.29. Fry, A. M., Jones, L. A., Kruisbeek, A. M. & Matis, L. A.
(1989) Science 246, 1044-1046.30. Suh, S. W., Bhat, T. N., Navia, M. A., Cohen, G. H., Rao,
D. N., Rudikoff, S. & Davies, D. R. (1986) Proteins 1, 74-80.31. Malissen, M., Trucy, J., Letourneur, F., Rebai, N., Dunn,
D. E., Fitch, F. W., Hood, L. & Malissen, B. (1988) Cell 55,49-59.
32. Pullen, A. M., Wade, T., Marrack, P. & Kappler, J. W. (1990)Cell 61, 1365-1374.
33. Choi, Y., Herman, A., DiGiusto, D., Wade, T., Marrack, P. &Kappler, J. (1990) Nature (London) 346, 471-473.
34. Robinson, M. A. (1989) Proc. Natl. Acad. Sci. USA 86, 9422-9426.
35. Posnett, D. N. (1990) Immunol. Today 11, 368-373.36. Seboun, E., Robinson, M. A., Doolittle, T. H., Ciulla, T. A.,
Kindt, T. J. & Hauser, S. L. (1989) Cell 57, 1095-1100.
Immunology: Gahm et al.