production and characterization of pectinase …
TRANSCRIPT

PRODUCTION AND CHARACTERIZATION OF PECTINASE INDUCED
FROM ASPERGILLUS NIGER USING PECTIN EXTRACTED FROM
PINEAPPLE PEELS AS CARBON SOURCE
BY
IBEAWUCHI, ADIELE NORA
PG/M.Sc/10/57599
DEPARTMENT OF BIOCHEMISTRY
UNIVERSITY OF NIGERIA NSUKKA
SEPTEMBER, 2012.

TITLE PAGE
PRODUCTION AND CHARACTERIZATION OF PECTINASE INDUCED FROM
ASPERGILLUS NIGER USING PECTIN EXTRACTED FROM PINEAPPLE PEELS AS
CARBON SOURCE.
A PROJECT WORK SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENT
FOR THE AWARD OF DEGREE OF MASTER OF SCIENCE (M.Sc) IN BIOCHEMISTRY,
UNIVERSITY OF NIGERIA NSUKKA.
BY
IBEAWUCHI, ADIELE NORA
PG/M.Sc/10/57599
DEPARTMENT OF BIOCHEMISTRY
UNIVERSITY OF NIGERIA NSUKKA
SUPERVISORS: PROF. F.C. CHILAKA AND DR. S.O.O. EZE
SEPTEMBER, 2012

CERTIFICATION
Ibeawuchi, Adiele Nora, a post-graduate student with registration number PG/M.Sc/10/57599 in
the Department of Biochemistry has satisfactorily completed the requirement for the course work
and research for the Master Degree of Science (M.Sc) in Biochemistry with Industrial
Biochemistry and Biotechnology as an option. This entire work in this report is original and has
not been submitted in part or full for any other diploma or degree in this or any other university.
__________________________ _______________________
Prof. F.C. Chilaka Dr S.O.O. Eze
(Chief Supervisor) (Co-Supervisor)
____________________________ ________________________
Prof. L.U.S. Ezeanyika External Examiner

DEDICATION
This work is dedicated to the members of my family, for their endless support and prayers. To
my parents Mr & Mrs D.E. Ibeawuchi and siblings; Ijeoma Ibeawuchi, Chinyere Nwanya,
Chinwe Osuagwu, Uloma Ibeawuchi and Victor Ibeawuchi; I say thank you.

ACKNOWLEDGEMENT
I would like to use this means to appreciate my wonderful supervisors; Prof F.C. Chilaka and Dr.
S.O.O. Eze, for their relentless effort in bringing this work to completion. I admire their patience,
hardwork and dedication. To Dr C.U. Anyanwu, I appreciate your generosity by allowing me
carry-out the microbiological aspect of this research in your laboratory.
To the HOD, Prof. L.U.S. Ezeanyika, I thank you for being a man of great honor and living an
exemplary life. To all my lecturers, thank you for imparting knowledge unto me from your
individual wells of experience and understanding. Dr J. Parker, you were always there to correct
and give me guidance whenever I fell short; you are appreciated.
Without a doubt, I thank Dr P. Mounmbegna, Dr F.M Awah and Dr S. Onuoha for being of
assistance when I considered enrolling for this programme. My colleagues; Nsude Nonso, Ezike
Tobechukwu, Ezugwu Arinze, Nwamaka Odu, Agu Chidozie and Okonkwo Chukwudi were
supportive and I thank them for their intelligent input in this work. My friends; Amaka Nwoye,
Onyedikachi Aruma, Preye Wokoro and Olufemi, you gave me a reason to smile every day. The
entire Biochemistry Post-Graduate class of 2010, we sure had an experience that made us better
people today.
I also would love to acknowledge my parents, siblings and their spouses for believing in me and
encouraging me throughout the entire M.Sc programme. My nephews; Chinomso and Chidozie
Osuagwu, Obinna Nwanya and Lewechi Ibeawuchi, you understood when I was always absent
on those special days. I love you all.
I ultimately want to thank God for HIS grace and mercies upon my life. He has been more than
faithful even when tough times prevailed and no silver lining was in sight; You still gladdened
my heart.

ABSTRACT
The peels of pineapple (Ananas cosmosus) generated as agricultural wastes represent about 35%
of the entire fruit. However, these wastes can be converted to various forms or more useful
products such as pectin and dietary fibres. Pectin was extracted from pineapple peels with a
percentage yield of 8.33% at pH 2.2 and temperature of 70ºC. Three pectinolytic fungi:
Aspergillus niger, Aspergillus fumigatus and Aspergillus flavus were isolated from natural
sources and tested for their pectinolytic activity. Aspergillus niger showed relatively higher
pectinolytic activity and was therefore used for further studies. Aspergillus niger was inoculated
into a broth containing pineapple pectin under submerged fermentation system. The enzyme was
harvested after four days of fermentation, the biomass was filtered off and 80% ammonium
sulphate precipitation was carried out on the crude enzyme. The crude enzyme was desalted
through dialysis and yielded a 4- fold increase in specific activity. The pH and temperature
optima were found to be 5.5 and 55ºC respectively. The Vmax and Km were also found to be
5000U and 89.5mg/ml respectively. Thus, pineapple peels can be successfully employed in the
production of pectinase.

TABLE OF CONTENTS
Title Page-------------------------------------------------------------------------------------I
Certification----------------------------------------------------------------------------------II
Dedication------------------------------------------------------------------------------------III
Acknowledgement--------------------------------------------------------------------------IV
Abstract---------------------------------------------------------------------------------------V
Table of Contents---------------------------------------------------------------------------VI
List of Figures-------------------------------------------------------------------------------VII
List of Tables -------------------------------------------------------------------------------XIV
List of Abbreviations-----------------------------------------------------------------------XV
CHAPTER ONE: INTRODUCTION
1.1 Pineapple- scientific classification------------------------------------------------2
1.1.1 History and description of pineapple----------------------------------------------3
1.1.2 Plant cell wall------------------------------------------------------------------------6
1.1.2.1 The middle lamella of the fruit cell------------------------------------------------6
1.2 Pectic substances----------------------------------------------------------------------7
1.2.1 Structure of pectic substances------------------------------------------------------8
1.2.2 Classification of pectic substances-------------------------------------------------9
1.3 Pectin----------------------------------------------------------------------------------10
1.3.1 Structural types of pectin-----------------------------------------------------------11
1.3.2 The primary cell wall pectic network---------------------------------------------13
1.3.3 Types of pectin-----------------------------------------------------------------------14
1.3.4 General properties of pectins-------------------------------------------------------15
1.3.5 Applications of pectin---------------------------------------------------------------15
1.3.5.1 Muco-adhesive polymer-------------------------------------------------------------15
1.3.5.2 Gelling agent, thickener &water binding-----------------------------------------15
1.3.5.3 Pectin in medicine and pharmaceutical industry--------------------------------16
1.4 Role of microbes in pectinase production----------------------------------------16
1.5 Pectinases-----------------------------------------------------------------------------17
1.5.1 Protopectinases----------------------------------------------------------------------18

1.5.2 Polygalacturonases------------------------------------------------------------------21
1.5.3 Lyases---------------------------------------------------------------------------------22
1.5.4 Pectinesterases-----------------------------------------------------------------------24
1.6 Biotechnological application of microbial pectinases--------------------------26
1.6.1 Fruit juice extraction----------------------------------------------------------------27
1.6.2 Coffee and tea fermentation-------------------------------------------------------27
1.6.3 Textile processing and bioscouring of cotton fibers----------------------------27
1.6.4 Degumming of plant bast fibers---------------------------------------------------27
1.6.5 Waste water treatment--------------------------------------------------------------28
1.6.6 Paper and pulp industry------------------------------------------------------------28
1.6.7 Animal feed--------------------------------------------------------------------------28
1.6.8 Purification of plant viruses--------------------------------------------------------28
1.6.9 Improvement of chromaticity and stability of red wines-----------------------28
1.7 Substrates for the production of pectinases--------------------------------------29
1.8 Fermentation conditions------------------------------------------------------------29
1.8.1 Types of fermentation--------------------------------------------------------------30
1.9 Micro-organisms commonly used in submerged and solid--------------------31
state fermentation for pectinases production
1.10 Aim and objectives of the study--------------------------------------------------32
1.10.1 Aim of the study---------------------------------------------------------------------32
1.10.2 Specific objectives of the study----------------------------------------------------32
CHAPTER TWO: MATERIALS AND METHODS
2.1 Materials-------------------------------------------------------------------------------33
2.1.1 Chemicals / Reagents----------------------------------------------------------------33
2.1.2 Apparatus / Equipment--------------------------------------------------------------33
2.1.3 Collection of pineapple samples---------------------------------------------------33
2.1.4 Collection of micro-organisms-----------------------------------------------------34
2.2 Methods-------------------------------------------------------------------------------34
2.2.1 Preparation of reagents--------------------------------------------------------------34

2.2.1.1 Preparation of 3N HCl--------------------------------------------------------------34
2.2.1.2 Preparation of ethanol-HCl solution----------------------------------------------34
2.2.1.3 Preparation of buffers---------------------------------------------------------------35
2.2.1.4 Preparation of 2mg/ml bovine serum albumin (BSA)--------------------------35
standard protein
2.2.1.5 Preparation of dinitrosalicylic acid (DNS) reagent-----------------------------35
2.2.1.6 Preparation of 20mM galacturonic acid------------------------------------------36
2.2.1.7 Preparation of component reagents for protein determination---------------36
2.2.2 Preparation of ground pineapple peels-------------------------------------------36
2.2.3 Extraction of pectin from pineapple peels---------------------------------------36
2.2.4 Isolation of pectinolytic fungi-----------------------------------------------------37
2.2.4.1 Collection of soil samples----------------------------------------------------------37
2.2.4.2 Preparation of soil sample extracts for microbial isolation-------------------37
2.2.4.3 Preparation of the solid medium--------------------------------------------------37
2.2.4.4 Sub-culturing onto solid medium-------------------------------------------------38
2.2.4.5 Storage of micro-organism on potato dextrose agar (PDA)------------------38
2.2.4.6 Microscopic features of the isolated fungi--------------------------------------38
2.2.4.7 Fungal identifications--------------------------------------------------------------38
2.2.5 Fermentation experiments---------------------------------------------------------38
2.2.5.1 Preparation of the fermentation medium----------------------------------------38
2.2.5.2 Inoculating with Aspergillus niger-----------------------------------------------39
2.2.5.3 Harvesting the crude enzyme-----------------------------------------------------39
2.2.6 Galacturonic acid standard curve-------------------------------------------------39
2.2.7 Polygalacturonase assay-----------------------------------------------------------40
2.2.7.1 Procedure for polygalacturonase assay------------------------------------------40
2.2.8 Protein determination--------------------------------------------------------------40
2.2.8.1 Procedure for protein determination---------------------------------------------40
2.2.9 Partial purification of protein-----------------------------------------------------41
2.2.9.1 Ammonium sulphate precipitation profile---------------------------------------41
2.2.9.2 Ammonium sulphate precipitation------------------------------------------------41
2.2.9.3 Dialysis-------------------------------------------------------------------------------41

2.2.10 Studies on partially purified enzymes--------------------------------------------42
2.2.10.1 Effect of pH change on pectinase activity---------------------------------------42
2.2.10.2 Effect of temperature change on pectinase assay-------------------------------42
2.2.10.3 Effect of substrate concentration on pectinase assay---------------------------42
2.2.10.4 Further studies with partially purified enzyme----------------------------------42
CHAPTER THRE: RESULTS
3.1 Pineapple pectin extraction--------------------------------------------------------43
3.1.1 Pineapple pectin extraction yield-------------------------------------------------43
3.1.2 Photograph of pineapple pectin extract------------------------------------------43
3.2 Micro-organisms--------------------------------------------------------------------44
3.2.1 Selection of pectinolytic fungi----------------------------------------------------44
3.2.2 Macroscopic and microscopic examination of fungal isolates----------------45
3.3 Pectinases production undersubmerged fermentation system----------------46
3.4 Ammonium sulphate precipitation-----------------------------------------------47
3.5 Assays carried out on the pectinases obtained----------------------------------48
3.5.1 Protein concentration of the crude, precipitated--------------------------------48
and dialyzed enzymes
3.5.2 Activity of pectinase in the crude, precipitated---------------------------------49
and dialyzed forms
3.5.3 Total protein content of the crude, precipitated---------------------------------50
and dialyzed pectinases
3.5.4 Total activity of the crude, precipitated------------------------------------------51
and dialyzed pectinase
3.5.5 Specific activity of the crude, precipitated--------------------------------------52
and dialyzed pectinases
3.5.6 Purification folds of the partially purified enzymes---------------------------53
3.6 Characterization of pectinase-----------------------------------------------------55
3.6.1 Effect of change in pH on pectinase activity------------------------------------55
3.6.2 Effect of change in temperature on pectinase activity-------------------------56
3.6.3 Kinetic parameters------------------------------------------------------------------57

CHAPTER FOUR: DISCUSSION
4.1 Discussion--------------------------------------------------------------------------------59
4.2 Conclusion-------------------------------------------------------------------------------62
4.3 Suggestions for further studies--------------------------------------------------------63
References------------------------------------------------------------------------------------64
Appendices-----------------------------------------------------------------------------------79

LIST OF FIGURES
Fig.1: Parts of a pineapple fruit-------------------------------------------------------------------4
Fig.2: Structure of the plant cell wall------------------------------------------------------------6
Fig. 3: Primary structure of pectic substances---------------------------------------------------9
Fig. 4: Structure of galacturonic acid------------------------------------------------------------10
Fig. 5: Schematic representative structures of the constituent--------------------------------11
polysaccharides of pectin
Fig. 6: HM pectin formula-------------------------------------------------------------------------14
Fig. 7: LM pectin formula-------------------------------------------------------------------------14
Fig. 8: Amidiated pectin formula-----------------------------------------------------------------14
Fig. 9: Enzymatic mode of action of some of the most frequent------------------------------20
depolymerases on the pectin molecule
Fig. 10: Photograph of pineapple pectin---------------------------------------------------------43
Fig. 11: Pure culture of Aspergillus niger-------------------------------------------------------45
Fig. 12: Ammonium sulphate precipitation profiling of pectinase----------------------------47
obtained from Aspergillus niger
Fig. 13: Comparison in the protein concentration of the crude,-------------------------------48
precipitated and dialyzed pectinases
Fig. 14: Comparison in the activities obtained from the crude,-------------------------------49
precipitated and dialyzed pectinases
Fig. 15: Comparison in the total protein content of the crude,---------------------------------50
precipitated and dialyzed
Fig.16: Comparison in the total activity of the crude,------------------------------------------51
Precipitated and dialyzed pectinases
Fig. 17: Comparison in the specific activities of the crude,------------------------------------52
precipitated and dialyzed pectinases
Fig. 18: Purification folds of the crude, precipitated and dialyzed pectinases--------------53
Fig. 19: Effect of change in pH on pectinase activity------------------------------------------55
Fig. 20: Effect of change in temperature on pectinase activity-------------------------------56
Fig. 21: Lineweaver-Burk plot of pectinases from Aspergillus niger----------------------57

LIST OF TABLES
Table 1: Nutritional values per 100g of pineapple------------------------------------------5
Table 2: Composition of pectic substances in different------------------------------------8
fruits and vegetables
Table 3: Extensive classification of pectinolytic enzymes--------------------------------19
Table 4: Biochemical and physicochemical properties of some-------------------------22
polygalacturonases
Table 5: Biochemical and physicochemical properties of--------------------------------24
some lyases
Table 6: Biochemical and physicochemical properties of --------------------------------26
some pectinesterases
Table 7: Comparison of solid state and submerged-----------------------------------------32
Fermentation for pectinases production
Table 8: Summary of the parameters determined from the crude------------------------54
precipitated and dialyzed pectinases
Table 9: Summary of pectinase characterization-------------------------------------------58

LIST OF ABBREVIATIONS
AG- Arabinogalactan
Ala- Alanine
Api- Apiogalacturonan
Ara- Arabinan
BSA- Bovine Serum Albumin
GRAS- Generally Regarded as Safe
HGA- homogalacturonan
HM- High Methylester
LM- Low Methylester
PDA- Potato Dextrose Agar
PE- Pectinesterase
PG- Polygalacturonase
PGase- Polygalacturonase
PGL- PolygalacturonateLyase
PME- Pectin methylesterase
PMG- polymethylgalacturonase
PMGE- Polymethylgalacturonate Esterase
PMGL- PolymethylgalacturonateLyase
Ppase- Protopectinase

Pro- Proline
RG- Rhamnogalacturonan
Rha- Rhamnose
Ser- Serine
SmF- Submerged fermentation
SSF- Solid state fermentation
Thr- Threonine
UDP-D- Uridinediphosphate
XGA- Xylogalacturonan
Xyl- Xylose
β-Gal- β-Galactosidase

CHAPTER ONE
INTRODUCTION
Pineapple (Ananas cosmosus) is the common name for a tropical plant and its edible fruits which
is actually a multiple fruit consisting of coalesced berries (Purseglove, 1972; Bartholomew et al.,
2003). Pineapple is by far the most economically important plant in the Bromeliaceae family.
Besides being produced for consumption, it can be grown as an ornamental or house plant
(Bartholomew et al., 2003). Pineapple can be consumed fresh, canned or juiced and can be used
in a variety of ways. It is popularly used in salads, jams, yoghurts, ice-cream, etc (Rohrbach et
al., 2003). However, processing and utilization of pineapple into various products leads to
generation of waste in form of peels and pomace. Pineapple waste can be conventionally bio-
transformed anaerobically into humus; although valuable by-products can be produced from the
rich waste. In other words, wealth can be derived from this waste by value addition and products
such as pectin, dietary fibers and predominantly pectinases can be easily harnessed.
Pectin was first isolated and described in 1825 by Henri Braconnot (Braconnot and Keppler,
1825). Pectin is one of the major components of the primary cellular walls in the middle lamella
of plant tissues. The pectic matrix provides an environment for deposition, slippage and
extension of the cellulosic-glycan network, and is the major adhesive materials between cells
(Willats et al., 2001). Pectin degradation leads to disassembly of the cellulose and hemicellulose
network and plays an important role in fruit ripening (Lohani et al., 2004; Sañudo-Barajas et al.,
2009). Wall degrading enzymes were the major factors to initiate disassembly of cellular walls of
harvested fruits (Miller and Fry, 2001). Involvement of pectic enzymes, such as
polygalacturonase (PG), pectin methyl esterase (PME) and β-galactosidase (β-Gal), in enzymatic
disassembly of cellular walls has been widely reported (Prasanna et al., 2007; Nikolić and
Mojovic, 2007; Rugkong et al., 2010; Wei et al., 2010; Almeida and Huber, 2011). In the food
sector, pectins are primarily used as a gelling agent in the production of jam; and as stabilizers in
the production of yoghurt (Koubala et al., 2006). In the pharmaceutical industry, these
polysaccharides are used as drug delivery systems, which can also reduce the toxicity of the
drugs (Morris et al., 2010; Pilnik et al., 1970; Schols et al., 2009; Thakur et al., 1997).

Pectinases are a group of enzymes, which cause degradation of pectin that, are chain molecules
with a rhamnogalacturonan backbone; associated with other polymers and carbohydrates. These
pectinases have wide applications in fruit juice industry and wine industry. In fruit juice industry,
it is used for clarification; reduction in viscosity is caused which ultimately leads to formation of
clear juice. They increase the yield of juices by enzymatic liquefaction of pulps; these pectinases
also helps in formation of pulpy products by macerating the organized tissue into suspension of
intact cells. In wine industry pectinases are mainly used for decreasing astringency by
solubilizing anthocyanins without leaching out procyadin polyphenols, and pectinases also
increase pigmentation by extracting more anthocyanins (Tucker and Woods, 1991). The
increasing energy demand has been focused on the utilization of renewable agricultural and
industrial wastes (Martin et al., 2004).
Pectinases can be produced by both submerged and solid state fermentation (SSF). Submerged
fermentation is cultivation of microorganisms on liquid broth. It requires high volumes of water,
continuous agitation and generates lot of effluents. SSF incorporates microbial growth and
product formation on or within particles of a solid substrate (Mudgett, 1986) under aerobic
conditions, in the absence or near absence of free water, and does not generally require aseptic.
1.1 Pineapple- Scientific Classification
Kingdom Plantae
Phylum Pteridofitae
Class Angiosperm
Sub-class Monocotyledoneal
Order Farinosae
Family Bromaliaceae
Sub-family Bromelioideae
Genus Ananas
Species Cosmosus
Binomial name Ananas cosmosus
Source: Py and Tisseau, 1969)

1.1.1 History and Description of Pineapple
Pineapple (Ananas comosus) is the common name for a tropical plant and its edible fruit, which
is actually a multiple fruit consisting of coalesced berries. It was given the name pineapple due to
its resemblance to a pine cone. The pineapple is by far the most economically important plant in
the Bromeliaceae family. (Coppens d‟Eeckenbrugge and Leal, 2003). Besides being produced for
consumption, it can be grown as an ornamental or houseplant, obtained from the crown of a
supermarket fruit.
Some sources say that the plant will flower after about 24 months and produce a fruit during the
following six months (Purseglove, 1972), while others indicate a 20-month timetable. Pineapple
can be consumed fresh, canned or juiced and can be used in a variety of ways. It is popularly
used in desserts, salads (usually tropical fruit salads, but it can vary), jams, yogurts, ice creams,
various candies, as a complement to meat dishes and in fruit cocktail. (Rohrbach et al., 2003).
The popularity of the pineapple is due to its sweet-sour taste. The core of the pineapple is
continuous with the stem supporting the fruit and with the crown, a feature unique among
cultivated fruits.
Pineapple contains the proteolytic enzyme bromelain which is used as a meat tenderizing agent
and for medicinal purposes. It has been reported to have valuable biological properties such as
interfering with the growth of malignant cells, inhibiting platelet aggregation, fibrinolytic and
anti-inflammatory actions. (Gailhofer et al., 1998). Pineapple leaf juice is used as a purgative
(agent that cleanses the bowel), emmenagogue (agent that induces menstrual bleeding) and
vermifuge (agent that expels intestinal worms). (Leal and Coppens d‟Eeckenbrugge, 1996).
The stems and leaves of the pineapple plant are sources of fiber, which can be processed into
paper and cloth. The cloth made from pineapple fiber is known as „pina cloth‟ and was in use as
early as 1571. Parts of the pineapple plant (Fig. 1) are used as silage and hay for cattle feed such
as the processed wastes in the form of pomace or centrifuged solids from juice production
(Wikipedia, 2011)

Fig. 1: Parts of a pineapple fruit (Elfick, 2007)
The word “pineapple” in English was first recorded in 1398, when it was originally used to
describe the reproductive organs of conifer trees (now termed pine cones). The term pine cone
for the reproductive organ of conifer trees was first recorded in 1694. When European explorers
discovered this tropical fruit, they called them pineapples (term first recorded in that sense in
1664 because of their resemblance to what is now known as the pine cone (Wikipedia, 2011)

Table 1: Nutritional Value per 100g of Pineapple Fruits
Nutrient Nutritional Value per 100g (3.5oz)
Energy 202kJ (48kcal)
Carbohydrate 12.63g
Sugar 9.26g
Dietary Fiber 1.4g
Fat 0.12g
Protein 0.54g
Thiamine (Vit B1) 0.079mg (3%)
Riboflavin (Vit B2) 0.03mg (3%)
Niacin (B3) 0.489mg (3%)
Panthothenic Acid( B5) 0.205
Vitamin B6 0.11mg (8%)
Folate (B9) 15µg (4%)
Vitamin C 36.2mg (44%)
Calcium 13mg (1%)
Iron 0.28mg (2%)
Magnesium 12mg
Source: Wikipedia, 2011

1.1.2 Plant Cell Wall
Plant cell walls consist of plant middle lamella, primary cell wall and secondary cell wall as can
be seen in Fig. 2. The primary walls of enlarging plant cells are composed of approximately 30%
cellulose, 30% hemicellulose and 35% pectin with about 1-5% structural protein (glycoprotein)
on a dry weight basis (Cosgrove, 1997).
Fig. 2: Structure of the Plant Cell Wall (Carpita and Gibeaut, 1993)
1.1.2.1 The Middle Lamella of the Fruit Cell
The middle lamella is the first layer formed during cell division, and can also be seen as the
space between the cell walls, and as the connecting region between adjacent cells, binding cells
together. The highest concentrations of pectin are found in the middle lamella of cell walls, with
a gradual decrease as one passes through the primary wall toward the plasma membrane
(Kertesz, 1951).

In the process of the middle lamella formation, the methyl-esterified homogalacturonan (HGA)-
rich carbon brushes or pectin with virtually no arabinan or galactan are deposited first in the cell
plate of two daughter cells. Subsequently, more layers of pectin molecules are deposited. There
may be de-esterification of HGA hairs by the activity of intact pectin methylesterase (PME). The
unesterified regions of HGAs from different cells become sensitive to Ca2+
and can form gel
(Vicken et al., 2003). Cosgrove (1997) reported the localization of PME and Ca2+
-pectate in the
areas of cell-cell contact such as the middle lamella. Hence, the middle lamella is a Ca2+
-pectate
gel formed by the cross-linking of first layer pectin (HGA-rich molecular brushes) to HGA-
containing molecular brushes of the primary cell wall of two neighboring cells (Vicken et al.,
2003).
1.2 Pectic Substances
Pectic substance is the generic name used for the compounds that are acted upon by the
pectinolytic enzymes. They are high molecular weight, negatively charged, acidic, complex
glycosidic macromolecules (polysaccharides) that are present in the plant kingdom. They are
present as the major components of middle lamella between the cells in the form of calcium
pectate and magnesium pectate (Rastogi, 1998). The synthesis of pectic substances occurs in the
Golgi apparatus from UDP-D-galcturonic acid during early stages of growth in young enlarging
cell walls (Sakai et al., 1993).
Compared with young, actively growing tissues, lignified tissues have a low content of pectic
substances. The content of the pectic substances is very low in higher plants usually less than
1%. They are mainly found in fruits and vegetables, constitute a large part of some algal biomass
(up to 30%) and occur in low concentration in forestry or agricultural residues (Table 2). Pectic
substances account for 0.5–4.0% of the fresh weight of plant material (Kashyap et al., 2001;
Sakai et al., 1993). Contrary to the proteins, lipids and nucleic acids, which are polysaccharides,
pectic substances do not have defined molecular masses.
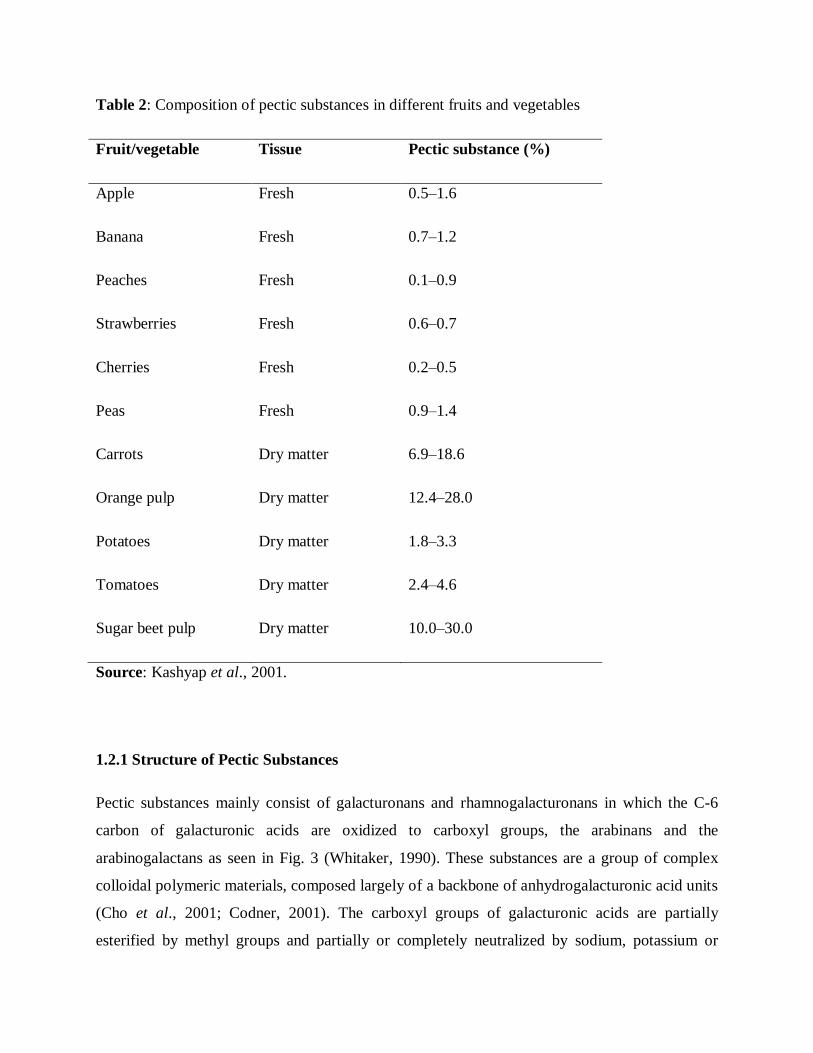
Table 2: Composition of pectic substances in different fruits and vegetables
Fruit/vegetable Tissue Pectic substance (%)
Apple Fresh 0.5–1.6
Banana Fresh 0.7–1.2
Peaches Fresh 0.1–0.9
Strawberries Fresh 0.6–0.7
Cherries Fresh 0.2–0.5
Peas Fresh 0.9–1.4
Carrots Dry matter 6.9–18.6
Orange pulp Dry matter 12.4–28.0
Potatoes Dry matter 1.8–3.3
Tomatoes Dry matter 2.4–4.6
Sugar beet pulp Dry matter 10.0–30.0
Source: Kashyap et al., 2001.
1.2.1 Structure of Pectic Substances
Pectic substances mainly consist of galacturonans and rhamnogalacturonans in which the C-6
carbon of galacturonic acids are oxidized to carboxyl groups, the arabinans and the
arabinogalactans as seen in Fig. 3 (Whitaker, 1990). These substances are a group of complex
colloidal polymeric materials, composed largely of a backbone of anhydrogalacturonic acid units
(Cho et al., 2001; Codner, 2001). The carboxyl groups of galacturonic acids are partially
esterified by methyl groups and partially or completely neutralized by sodium, potassium or

ammonium ions (Kashyap et al., 2001). The primary chain consists of D-galacturonic acid units
linked α-(1-4), with 2–4% of L-rhamnose units linked β-(1-2)and β-(1-4) to the galacturonic acid
units (Whitaker, 1990). The side chains of arabinan, galactan, arabinogalactan, xylose or fructose
are connected to the main chain through their C-1 and C-2 atoms (Blanco et al., 1999;
Sathyanarayama and Panda, 2003; Van der Vlugt-Bergmans et al., 2000). The above description
indicates that the pectic substances are present in various forms in plant cells and this is the
probable reason for the existence of various forms of pectinolytic enzymes.
Fig. 3: Primary Structure of Pectic Substances (Pilnik and Voragen, 1993)
1.2.2 Classification of Pectic Substances
According to Alkorta et al., 1998, the American Chemical Society classified pectic substances as
follows:
i) Protopectin: is the water insoluble pectic substance present in intact tissue. Protopectin on
restricted hydrolysis yields pectin or pectic acids.
ii) Pectic acid: is the soluble polymer of galacturonans that contains negligible amount of
methoxyl groups. Normal or acid salts of pectic acid are called pectates.
iii) Pectinic acids: is the polygalacturonan chain that contains >0 and <75% methylated
galacturonate units. Normal or acid salts of pectinic acid are referred to as pectinates.
iv) Pectin (Polymethyl galacturonate): is the polymeric material in which, at least, 75% of the
carboxyl groups of the galacturonate units are esterified with methanol. It confers rigidity on cell
wall when it is bound to cellulose in the cell wall.

1.3 Pectin
Through various studies, it has been brought in notice that the structure of pectin is difficult to
determine because pectin subunit composition can change during isolation from plants, storage
and processing of plant material (Novosd‟skaya, 2002). Pectin was first isolated and described in
1825 by Henri Braconnot (Braconnot and Keppler., 1825). At present, pectin is thought to
consist mainly of D-galacturonic acid (Gal A) units (Sriamornsak, 2002), joined in chains by
means of α (1-4) glycosidic linkage (Fig. 4). These uronic acids have carboxyl groups which are
naturally present as methyl esters and others which are commercially treated with ammonia to
produce carboxiamide group (Sriamornsak., 1998; Yujaroen et al., 2008).
Fig. 4: Structure of Galacturonic Acid (Pilnik and Voragen, 1993)
Pectin is composed of as many as 17 different monosaccharides (Ridley et al., 2001; Voragen et
al., 2003). These monosaccharides are organized in a number of distinct polysaccharides, the
structures of which are schematically shown in Fig. 5a→g. Together, these polymers form the
pectin network (Visser and Voragen, 1996; Ridley et al., 2001; Voragen et al., 2003).

Fig. 5(a→g): Schematic representative structures of the constituent polysaccharides of pectin.
The symbols for the various monosaccharide building units are explained in the accompanying
legend. (Vincken et al., 2003)
1.3.1 Structural Types of Pectin
Pectins are made up of a backbone of 1,4- linked α-D Galp A residues and are referred to as
galacturonans. Three types of galacturonan can be distinguished in the plant cell wall: the
unsubstituted homogalacturonan (HG), rhamnogalacturonan II (RG-II) and xylogalacturonan
(XGA) (Schols et al., 1995; Visser and Voragen, 1996). Addition of side chains bring about
other polysaccharides such as rhamnogalacturonan I (RG-I), arabinogalactan I and II (AG-I &
AG-II).
The Gal A residues of HG can be methyl-esterified at C-6 and carry acetyl groups on O- 2 and
O-3. This however affects to a large extent, the industrial applicability of pectins. Blocks of more
than 10 unesterified Gal A residues generally yield pectin molecules which are sensitive to Ca 2+
-
cross-linking (Daas et al., 2001). Two unesterified HG chains can engage in a complex
a)
d) e) f)
g)
b) c)

sometimes referred to as “egg boxes”, in which the carboxyl groups of two GalA residues form a
negatively charged pocket that can accommodate a Ca2+
cation (Braccini et al., 1999; Willats et
al., 2001a).
RG-II is not made of a rhamnogalacturonan backbone as the name implies, rather the rhamnose
residues are present in the side chains (Ridley et al., 2001). RG-II are formed when HG contain
clusters of four different side chains with very peculiar sugar residues such as Api, Ace A, Dha
and Kdo). These side chains together with the approximately nine galacturonyl residues to which
they are connected are referred to as RG II (O‟Neil et al., 2001; Ridley et al., 2001). Also the
order in which they follow on the backbone is not known with a certainty (du Penhoat et al.,
1999; Glushka et al., 2003; Perez et al., 2003)
XGA is a branched galacturonan with β-D-xylp-(1→3) side chains (Schols et al., 1995; Visser
and Voragen, 1996).
There is another constituent polysaccharide of pectin known as rhamnogalacturonan I (RG-I). It
is composed of a repeating disaccharide unit [→2),α-L-Rhap-(1→4)-α-D-Galp A-(1→]n, where
n can be larger than 100 (McNeil et al., 1980; Visser and Voragen, 1996). The galacturonyl
residues can carry acetyl groups on O-2 and O-3 while the rhamnosyl residues can be substituted
at O-4 with neutral sugars (McNeil et al., 1980; Lau et al., 1987). These neutral sugars are
mainly D- galactose, L- arabinose and D- xylose (Shembekar and Dohtre, 2009). The side chains
of RG-I can be single units (β-D-Galp-(1→4) or polymeric such as arabinogalactan I (AG-I)
which is composed of 1,4- linked β-D-Galp backbone or arabinan which consists of 1,5- linked
α-L-Araf backbone (Ridley et al., 2001).
Complexes of RG-I, AG-I and arabinan are often referred to as pectic hairy regions (HR) in
which AG-I and arabinan are the “hairs”. However, arabinogalactan II is mainly associated with
proteins (arabinogalactan proteins or AGPs) and it is still unclear whether this polysaccharide is
part of the pectin complex. Pectin and AG-II often seem to co-extract and are subsequently
difficult to separate from each other, suggesting that they can be covalently linked. AG-II is
composed of 1,3- linked β-D-Galp backbone and short side chains of α-L-Araf-(1→6)-[β-D-
Galp-(1→6)]n where n=1, 2 or 3 (Ridley et al., 2001). The major moiety of AGp (> 90%)

consists of polysaccharides while the protein moiety is rich in Pro, Ala, Ser and Thr (Gaspar et
al., 2001).
1.3.2 The primary cell wall pectic network
The covalent cross-linking of the pectic polysaccharides HG, RG-I, and RG-II has been
demonstrated repeatedly in the literature by the EPGase-dependent release of pectic
polysaccharides from the wall (O‟Neil et al., 1996). The available data suggest that the RG-I and
RG-II backbones are continuous with the HG backbone, not that of RG-I side chains, as
suggested by Vincken et al. (2003). If the backbones of the pectins are continuous, the pectic
network may be thought of as a macromolecular structure having specific domains of HG, RG-I,
and RG II, however, the arrangement of these domains in vivo is not known. The linkage of HG,
RG-I, and RG-II through backbone glycosidic linkages is just one possible way in which the
pectins are cross-linked.
The pectic network is based on multiple levels of cross-linking that include, but are not limited
to, backbone glycosidic linkages, calcium cross-linking, borate ester cross-linking, and covalent
linkages to phenolic and possibly other compounds. The HG domains of pectin may self-
associate depending on the degree of methylesterification and thus the affinity of HG for calcium
ions. RG-I has a unique backbone of alternating 2-linked Rhap and 4-linked GalpA residues.
Some rhamnose residues are branched by arabinan, galactan, and/or AG side chains (McNeil et
al., 1980) that may be cross-linked to other wall components such as xylans, xyloglucans,
proteins, and lignins. RG-II domains form cross-links to other RG-II molecules via borate di-
ester linkages, to form RG-II dimmers that contribute to wall strength and that affect pore size
and flexibility of the pectic network (Ishii and Matsunaga, 1996; Fleisher et al., 1999). Greater
than 95% of RG-II molecules participate in dimer complexes of RG-II (O‟Neil et al., 2004).
The linkages that pectic polysaccharides make to other pectins, as well as to other wall
molecules, combine to assemble the pectic network of the plant cell wall. The complexity of the
pectic network structure and the modulation of the pectic cross-links contribute strength,
flexibility, and functionality to the pectic network, and thus, to the primary cell wall.

1.3.3 Types of Pectin
The percentage of ester groups is called degree of esterification. High methyl ester pectins are
classified in groups according to their gelling temperature as rapid set to slow set pectins.
Pectin as extracted normally has more than 50% of the acid units esterified, and is classified as
"high methyl ester (HM) pectin" as shown in Fig. 6.
Fig. 6: HM pectin formula (IPPA, 2001)
Modification of the extraction process, or continued acid treatment, will yield ”low methyl ester
LM pectin" with less than 50% methyl ester groups as observed in Fig. 7.
Fig. 7: LM pectin formula (IPPA, 2001)
Some pectins are treated during manufacture with ammonia to produce amidated pectins, which
have particular advantages in some applications.
Fig. 8: Amidated pectin formula (IPPA, 2001)

1.3.4 General Properties of Pectins
Pectin is soluble in pure water as monovalent cation (alkali metal) salts of pectinic and pectic
acids; are usually soluble in water unlike di- and trivalent cation salts that are weakly soluble or
insoluble (Sriamornsak, 1998). Dilute pectin solutions are Newtonian but at a moderate
concentration, they exhibit the non-Newtonian, pseudo plastic behavior characteristics.
Viscosity, solubility and gelation are generally related. For example, factors that increases gel
strength will increase viscosity and vice versa (Marshal and Chow, 2007). As such, monovalent
cation salts of pectins are highly ionized in solution and the distribution of ionic charges along
the molecule tends to keep it in extended form by reason of coulombic repulsion (Paolettis,
1986). Dissolved pectins are decomposed spontaneously by de-esterification as well as by
depolymerization. Hence the rate of decomposition depends on pH, water activity and
temperature (Sriamornsak, 1998).
1.3.5 Applications of Pectin
1.3.5.1 Mucoadhesive Polymer
Different types of pectin were characterized for gastrointestinal (GI) mucoadhesion
(Sriamornsak, 2002). The mucoadhesive process involved in the formation of bioadhesive bonds
has been described in 3 steps;
i) Wetting and swelling of polymers to permit intimate contact with biological tissues.
ii) Interpenetration of bioadhesive polymer chain and entanglement of polymer and mucin chains
and
iii) Formation of weak chemical bonds between entangled chains (Malviya et al., 2010;
Srivastava et al., 2010).
1.3.5.2 Gelling agent, Thickener and Water Binder
Pectins are mainly used as gelling agents, but can also act as thickner, water binder and
stabilizer. Low methoxyl pectins (< 50% esterified) form thermo-reversible gels in the presence
of calcium ions and at low pH (3 – 4.5) whereas high methoxyl pectins rapidly form thermally

irreversible gels in the presence of sufficient sugars such as sucrose and at low pH (<3.5); the
lower the methoxyl content, the slower the set (Kohn, 1982).
1.3.5.3 Pectin in Medicine and Pharmaceutical Industry
i) As dietary fibers
Ruminant nutritionists recommend that the digestibility and energy concentration in forages can
be improved by increasing pectin concentration in the forage (Helene et al., 2005).
ii) In site specific targeting
Pectin has a promising pharmaceutical use and it is presently considered as a carrier material in
colon-specific drug delivery system (Sriamornsak and Nunthanid, 1998), for the treatment of
diseases such as ulcerative colitis, colon carcinomas. The rationale for this is that pectin and
calcium pectinate will be degraded by colonic pectinolytic enzymes (Malviya et al., 2010), but
will slow down the drug release in the upper gastrointestinal tract due to its insolubility and
because it is not degraded by gastric or intestinal enzymes. Therefore it reduces the toxicity of
the drugs and makes their activity longer lasting without altering their therapeutic effects (Morris
et al., 2010; Pilnik et al., 1970; Schols et al., 2009; Thakur et al., 1997).
1.4 Role of Microbes in Pectinase Production
Pectolysis is one of the most important processes for plant, as it plays a role in cell elongation
and growth as well as in fruit ripening. Pectolytic enzymes are wide spread in nature and are
produced by Bacteria, Fungi, Yeast, Insects, Nematodes and Protozoa. For example Bacteria
such as Bacillus species, Clostridium species; Fungi such as Aspergillus species, Penicillum
species; Yeast such as Saccharomyces and Candida. Microbial pectolysis is important in plant
pathogenesis, symbiosis and decomposition of plant deposits (Lang and Dornenburg, 2000).
Thus by breaking down pectin polymer for nutritional purposes, microbial pectolytic enzymes
play an important role in nature. The enzymes are inducible i.e. produced only when needed and
they contribute to the natural carbon cycle.
Microbial pectinolytic enzymes are not only enzymes available to attack plant polysaccharides.
However, pathogenic attack on plant tissue in normally initiated by pectic enzymes because
pectic substances are most readily accessible. Aspergilli are used by industries for the production

of primary metabolites (organic acids, vitamins, fatty acids and amino acids) and enzymes as
well as bioconversion processes. Two species, Aspergillus niger and Aspergillus oryzae prevail
as enzyme producers (Oxenboll, 1994). While Aspergillus niger is mainly exploited in the area
of food (bakery, starch industry) and beverages (wine and juice, brewery, distilling industry),
Aspergillus oryzae is applied in the fermentation of oriental sauces such as soya sauce. Hence
many of their products obtained the GRAS (Generally Regarded As Safe) status.
1.5 Pectinases
These are a group of related enzymes and may be divided in three broader groups as follows
(Sakai, 1992; Palomaki and Saarilahti, 1997):
i) Protopectinases: degrade the insoluble protopectin and give rise to highly polymerized soluble
pectin.
ii) Esterases: catalyze the de-esterification of pectin by the removal of methoxy esters.
iii) Depolymerases: catalyze the hydrolytic cleavage of the α (1,4) -glycosidic bonds in the D-
galacturonic acid moieties of the pectic substances. Depolymerases act on pectic substances by
two different mechanisms; hydrolysis, in which they catalyze the hydrolytic cleavage with the
introduction of water across the oxygen bridge and trans-elimination lysis, in which they break
the glycosidic bond by a trans-elimination reaction without any participation of water molecule
(Codner, 2001; Albersheim et al., 1960). Depolymerases can be subdivided into four different
categories, depending on the preference of enzyme for the substrate, the mechanism of cleavage
and the splitting of the glycosidic bonds (Rexova-Benkova and Markovic, 1976).
Polygalacturonase and polymethylgalacturonase breakdown pectate and pectin, respectively by
the mechanism of hydrolysis. However, polygalacturonate lyase and polymethylgalacturonate
lyase breakdown pectate and pectin by β-elimination, respectively. Depending upon the pattern
of action, i.e. random or terminal, these enzymes are termed as Endo or Exo enzymes,
respectively.

1.5.1 Protopectinases
This enzyme catalyzes the solubilization of protopectin. Pectinosinase is also synonymous with
protopectinase (PPase). Protopectinase catalyzes the following reaction:
Protopectin + H20 Pectin
(insoluble) (soluble)
Protopectinases are classified into 2 types based on their mechanism of action: A-type
protopectinases react with the inner site, i.e. the polygalaturonic acid region of protopectin
whereas B-type protopectinases react on the outer site i.e. on the polysaccharide chains that may
connect the polygalacturonic acid chain and cell wall constituents (Sakamoto et al., 1994).
The A-type protopectinases are found in the culture filtrates of yeast and yeast-like fungi. Some
have been isolated from Kluyveromyces fragilis IFO 0288, Galactomyces reesei L. and
Trichosporon penicillatum SNO 3 referred to as protopectinase -F, -L and –S respectively
(Whitaker, 1990).
Subsequently, B-type protopectinases have been discovered in Bacillus subtilis IFO 12113,
Bacillus subtilis IFO 3134 and Trametes sp. These are referred to as protopectinase –B, -C and –
T respectively (Sakai and Ozaki, 1988).
All 3 A-type protopectinases are similar in biological properties and have similar molecular
weight of 30 kDa. Protopectinases -F is an acidic protein while protopectinases –L and –S are
basic proteins. The protopectinases -B, -C and –T have molecular weights of 45, 30 and 55kDa
respectively. However, these 3 types act on protopectin from various citrus fruits and other plant
tissues, releasing pectin (Sakai, 1992).

Table 3. Extensive Classification of Pectinolytic Enzyme
Source: Kashyap et al., (2001)
Enzyme E.C No Action
Mechanism
Action
Pattern
Primary
Substrate
Product
ESTERASE 1. Pectin
Methylesterase
3.1.1.11
Hydrolysis
Random
Pectin
Pectic acid +
Methanol
DEPOLYMERASE
a) Hydrolase 1. Protopectinase
Hydrolysis
Random
Protopectin
Pectin
2. Endo-
polygalacturonase
3.2.1.15 Hydrolysis Random Pectic Acid Oligogalacturona
tes
3. Exo-
polygalacturonase
3.2.1.67 Hydrolysis Terminal Pectic Acid Monogalacturon
ates
4. Exo-
polygalacturonan di-
galacturonohydrolase
3.2.1.82 Hydrolysis Penultimate
bonds
Pectic Acid Digalacturonates
5.Oligo-galacturonate
hydrolase
Hydrolysis Terminal Trigalacturonat
e
Monogalacturon
ates
6. Δ4:5 Unsaturated
Oligo-galacturonate
hydrolases
Hydrolysis Terminal Δ4:5
(Galacturonate)
n
Unsaturated
monogalacturona
tes & Saturated
(n-1)
7.Endo-polymethyl-
galacturonase
Hydrolysis Random Highly
Esterified
Pectin
Oligomethylgala
cturonates
8.Endo-polymethyl-
galacturonase
Hydrolysis Terminal Highly
Esterified
Pectin
Oligogalacturona
tes
b) Lyases
1. Endo-
polygalacturonase lyase
4.2.2.2
Trans-
elimination
Random
Pectic Acid
Unsat Oligo-
galacturonates
2. Exo-
polygalacturonase lyase
4.2.2.9 Trans-
elimination
Penultimate
bond
Pectic Acid Unsat Oligo-
galacturonates
3. Oligo-D-
galacturonate lyase
4.2.2.6 Trans-
elimination
Terminal Unsat
digalacturonates
Unsat Mono-
galcturonates
4. Endo-polymethyl-D-
galacturonate lyase
4.2.2.10 Trans-
elimination
Random Unsat poly-
(methyl-D-
galacturonates)
Unsat methyl
Oligogalacturona
tes
5. Exo-polymethyl-D-
galacturonate lyase
Trans-
elimination
Terminal Unsat poly-
(methyl-D-
galacturonates)
Unsat methyl
Monogalacturon
ates.

Fig. 9: Enzymatic mode of action of the most frequent depolymerases on the pectin molecule.
PMGL: Polymethylgalacturonate lyase (pectin lyase); PMG: Polymethylgalacturonase (pectin
hydrolase); PMGE: Polymethylgalacturonate esterase (pectinesterase); PGL: Polygalacturonate
lyase (pectate lyase); PG: Polygalacturonase (pectate hydrolase). (Serra et al., 1992)

1.5.2 Polygalacturonases
These are enzymes that catalyze the hydrolytic cleavage of the polygalacturonic acid chain with
the introduction of water across the oxygen bridge. They can be subdivided into Exo and Endo-
PGase.
Endo-PGases are found in fungi, bacteria and many types of yeast (Luh and Phaff, 1951) also in
some higher plants and some plant parasitic nematodes (Sakai et al., 1993). In contrast, exo-
PGases occur less frequently as they have been reported in Erwina carotovora and Bacillus sp.
(Koboyashi et al., 2001)).
Exo-PGases can exist in 2 forms; as fungal exo-PGases which produces mono-galacturonic acid
as the main end product and as bacterial exo-PGases, which produces di-galacturonic acid as the
main end product (Sakai et al., 1993).
PGases isolated from different microbial sources differ markedly from each other with respect to
their physicochemical and biological properties as well as their mode of action. Table 4
summarizes the biochemical and physicochemical properties of PGases obtained from various
sources. Most of the PGases obtained from different microbial sources have an optimal pH range
of 3.5 - 5.5 and optimal temperature range of 30-50 ºC; but 2 endo-PGases (PG I & PG II)
isolated from Aspergillus niger have optimal pH range of 3.8 - 4.3 and 3.0 - 4.6 respectively
(Singh and Rao, 2002).
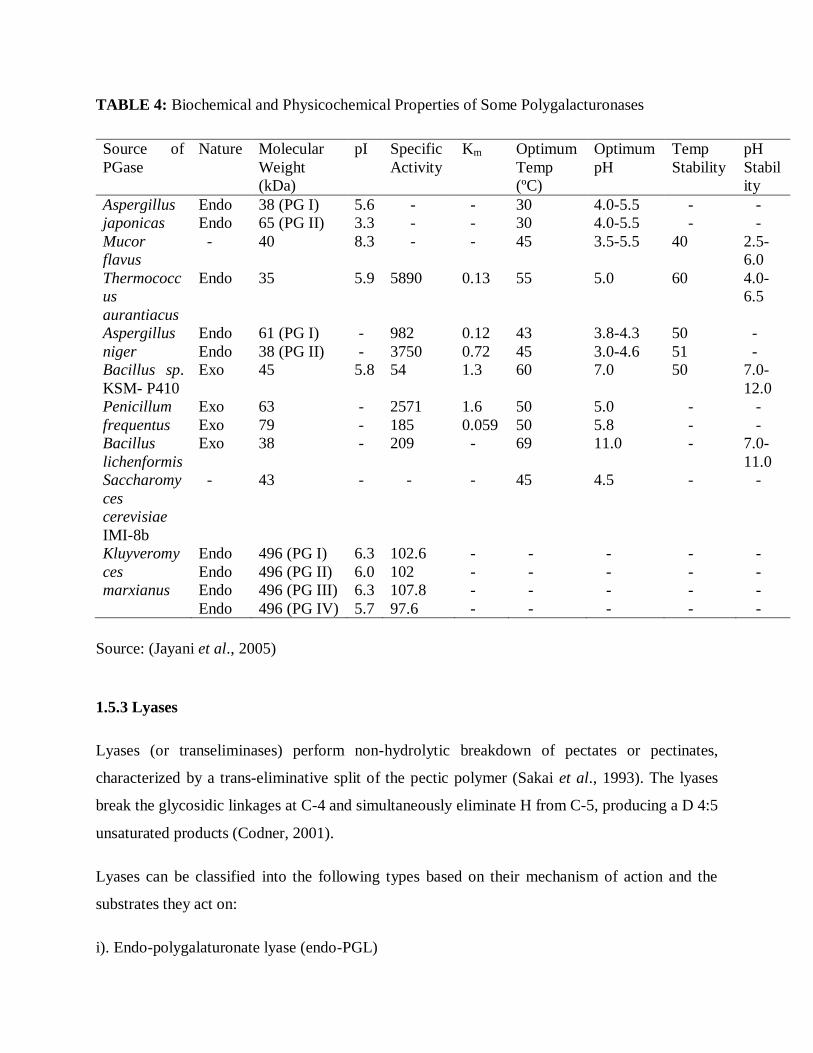
TABLE 4: Biochemical and Physicochemical Properties of Some Polygalacturonases
Source of
PGase
Nature Molecular
Weight
(kDa)
pI Specific
Activity
Km Optimum
Temp
(ºC)
Optimum
pH
Temp
Stability
pH
Stabil
ity
Aspergillus
japonicas
Endo
Endo
38 (PG I)
65 (PG II)
5.6
3.3
-
-
-
-
30
30
4.0-5.5
4.0-5.5
-
-
-
-
Mucor
flavus
- 40 8.3 - - 45 3.5-5.5 40 2.5-
6.0
Thermococc
us
aurantiacus
Endo 35 5.9 5890 0.13 55 5.0 60 4.0-
6.5
Aspergillus
niger
Endo
Endo
61 (PG I)
38 (PG II)
-
-
982
3750
0.12
0.72
43
45
3.8-4.3
3.0-4.6
50
51
-
-
Bacillus sp.
KSM- P410
Exo 45 5.8 54 1.3 60 7.0 50 7.0-
12.0
Penicillum
frequentus
Exo
Exo
63
79
-
-
2571
185
1.6
0.059
50
50
5.0
5.8
-
-
-
-
Bacillus
lichenformis
Exo 38 - 209 - 69 11.0 - 7.0-
11.0
Saccharomy
ces
cerevisiae
IMI-8b
- 43 - - - 45 4.5 - -
Kluyveromy
ces
marxianus
Endo
Endo
Endo
Endo
496 (PG I)
496 (PG II)
496 (PG III)
496 (PG IV)
6.3
6.0
6.3
5.7
102.6
102
107.8
97.6
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Source: (Jayani et al., 2005)
1.5.3 Lyases
Lyases (or transeliminases) perform non-hydrolytic breakdown of pectates or pectinates,
characterized by a trans-eliminative split of the pectic polymer (Sakai et al., 1993). The lyases
break the glycosidic linkages at C-4 and simultaneously eliminate H from C-5, producing a D 4:5
unsaturated products (Codner, 2001).
Lyases can be classified into the following types based on their mechanism of action and the
substrates they act on:
i). Endo-polygalaturonate lyase (endo-PGL)

ii). Exo-polygalacturonate lyase (exo-PGL)
iii). Endo-polymethylgalacturonate lyase (endo-PMGL)
iv).Exo-polymaethylgalacturonate lyases (exo-PMGL)
Polygalacturonate lyases, otherwise known as pectate lyases or PGLs are produced by many
bacteria and some pathogenic fungi with endo-PGLs being more abundant than exo-PGLs. PGLs
have been isolated from bacteria and fungi associated with food spoilage and soft rot.
Erwina carotovora (Kotoujansky, 1987), Collectotrichum lindemuthionum (Wijesundera et al.,
1984) have been reported to produce PGL while PMGLs have been reported to be produced by
Aspergillus japonicas (Ishii and Yokosuka, 1975) and Aspergillus sp (Sunnotel and Nigam,
2002).
In bacteria, lyases are the largest group of pectinolytic enzymes and are directly involved in plant
pathogenicity (Dixit et al., 2004). PGLs have an absolute requirement for Ca2+
ions (Margo et
al., 1994) and hence chelating agents such as EDTA act as their inhibitors whereas PMGLs do
not have an absolute requirement of cations but are stimulated by Ca2+
and other cations (Szajer
and Szajer, 1984).Properties of some of the lyases have been shown on Table 5.
Interestingly, endo-PMGL is the only enzyme known to be able to cleave the α 1,4-glycosidic
bonds of highly esterified pectins without the prior action of other enzymes (Sakai et al., 1993;
Alana et al., 1990).

Table 5: Biochemical and Physicochemical Properties of Some Lyases
Source: (Jayani et al., 2005)
1.5.4 Pectinesterases
Pectinesterase (PE) often referred to as pectin methylesterase, is a carboxylic acid esterase and
belongs to the hydrolase group of enzymes (Whitaker, 1984). It catalyzes the de-esterification of
methyl ester linkages of galacturonan backbone of pectic substances to release acidic pectins and
Source of
Lyases
Nature Molecular
Weight
(kDa)
pI Km Optimum
Temp
(ºC)
Optimum
pH
Temp
Stability
pH
Stability
Erwinia
cantovora
PGL(PL I)
PGL (PL II)
36-38
36-38
10.7
10.1
0.12
1.1
50
60
10.0
10.0
-
Bacteroides
thetaiotaomicr
on
PGL 74 7.5 0.04-
0.07
- 8.7
Aureobasidium
pullulans LV-
10
PMGL (L I)
PMGL(L II)
89
55
-
-
-
-
40
40
5.0
7.5
50
40
3.5
5.9
Penicillum
italicum
PMGL 22 8.6 3.2 40 6.0- 7.0 50 8.0
Aspergillus
japonicas
PMGL - 7.7 0.16 55 6.0 -
Bacillus sp PGL 38 - - 69 11.0 60 7.0-
11.0
Bacillus sp
TS44
PGL 50 5.3 - 70 8.0 70 11.0
Thermoascus
auratniacus
PMGL - - - 65 10.5-11.0 70 4.0

methanol (Cosgrove, 1997). The resulting pectin is then acted upon by polygalacturonases and
lyases (Sakai et al., 1993; Prade et al., 1999).
The mode of action of PE varies according to its origin (Micheli, 2001). Fungal PEs act by a
multi-chain mechanism, removing the methyl groups at random. In contrast, plants PEs tend to
act either at the non-reducing end or next to a free carboxyl group, and proceed along the
molecule by a single chain mechanism. This reaction catalyzed by PE can be represented in the
reaction below
Pectin + nH2O Pectate + nC2H5OH
Pectinesterase activity is implicated in cell wall metabolism including cell growth and ripening
of fruits (Gaffe et al., 1997; Dorokhov et al., 1999). Commercially, PE can be used for protecting
and improving the texture and firmness of several fruits and vegetables as well as in the
extraction and clarification of fruit juices (Gailing et al., 2000). Pectinesterase is found in plants,
pathogenic bacteria and fungi (Hasunuma et al., 2003) such as Saccharomyces cerevisiae
(Gainvors et al., 1994), Aspergillus niger (Maldonaldo et al., 1994; Maldonaldo and Saad, 1998),
E. chrysanthemi 3604 (Laurent et al., 2000). Also there are many reports of occurrence of PE in
plants such as Carica papaya (Fayyaz et al.1993; Innocenzo and Lajalo, 2001), Vitis vinifera
(Corredig et al., 2000), Citrus sp. (Arias and Burns, 2002) and Pouteria sapota (Arenas-Ocampo
et al., 2003). Pectinesterase shows highest activity on 65-75% methylated pectin since the
enzyme is thought to act on methoxyl groups adjacent to free carboxyl groups (Whitaker, 1984).
PE are highly specific enzymes in that, some PE attack only at the reducing chains while others
attack the non-reducing end (Sakai et al., 1993). Most PE are active at a pH range from 4.0-8.0,
however fungal PE have lower pH optimum than those from bacterial sources. Table 6 shows
properties of some pectinesterases.

Table 6: Biochemical and Physicochemical Properties of Some Pectinesterases
Source of
PE
Molecular
Weight
(kDa)
pI Km Optimum
Temp
(ºC)
Optimu
m pH
Temp
Stability
(ºC)
pH
Stability
Rhodotorul
a sp
- - - 40 6.0 50 4.9-9.0
Erwina
chrysanthe
mi 3341
37 9.6-9.9 - 50 5.0-9.0 - -
Aspergillus
niger
- - 1.01 45 5.0 - -
ApplePE 36 9.0 0.12
3
60 7.0 75 -
E.
chrysanthe
mi 3604
37 - 0.03 50 8.0-9.0 - -
Aspergillus
japonicas
46 (PE I)
47 (PE II)
3.8
3.8
-
-
-
-
4.0-5.5
4.0-5.5
50
50
-
-
Source: (Jayani et al., 2005)
1.6 Biotechnological Applications of Microbial Pectinases
Over the years, pectinases have been used in several conventional industrial processes, such as
textile, plant fiber processing, oil extraction, treatment of industrial wastewater, containing
petinacious material, etc. They have also been reported to work on purification of viruses
(Salazar and Jayasinghe, 1999) and in making of paper (Reid and Richard, 2004; Viikari et al.,
2001). However they are yet to be commercialized.

1.6.1 Fruit Juice Extraction
The largest industrial application of pectinases is in fruit juice extraction and clarification. A
mixture of pectinases and amylases is used to clarify fruit juices and thus decreasing the filtration
time up to 50% (Blanco et al., 1999). Treatment of fruit pulps with pectinases also showed an
increase in fruit juice volume from banana, grapes and apples (Kaur et al., 2004). Pectinases in
combination with other enzymes such as cellulases, arabinases and xylanases, have been used to
increase the pressing efficiency of the fruits for juice extraction (Gailing et al., 2000). Vacuum
infusion of pectinases has a commercial application to soften the peel of citrus fruits for removal.
This technique may expand in future to replace hand cutting for the production of canned
segments (Baker and Wicker, 1996).
1.6.2 Coffee and Tea Fermentation
Pectinase treatment accelerates tea fermentation and also destroys the foam forming property of
instant tea powders by destroying the pectins (Carr, 1985). Pectinolytic microorganisms are used
in the fermentation of coffee to remove the mucilaginous coat from the coffee beans.
1.6.3 Textile Processing and Bioscouring of Cotton Fibers
Pectinases have been used in conjunction with amylases, lipases, cellulases and hemicellulases to
remove sizing agents from cotton in a safe and eco-friendly manner, replacing toxic caustic soda
used for the purpose earlier (Hoondal et al., 2000). Bio-scouring is a novel process for removal
of non-cellulosic impurities from the fiber with specific enzymes. According to Hoondal et al.,
(2000) pectinases have been used for this purpose without any negative side effect on cellulose
degradation.
1.6.4 Degumming of Plant Bast Fibers
Bast fibers are the soft fibers formed in groups outside the xylem, phloem or pericycle. For
example Ramie and sunn hemp. The fibers contain gum, which must be removed before its use
for textile making (Hoondal et al., 2000). The chemical degumming treatment is polluting, toxic
and non-biodegradable. Biotechnological degumming using pectinases in combination with

xylanases presents an eco-friendly and economic alternative to the above problem (Kapoor et al.,
2001).
1.6.5 Waste Water Treatment
Vegetable food processing industries release pectin, containing wastewaters as by-product.
Pretreatment of these wastewaters with pectinolytic enzymes facilitates removal of pectinaceous
material and renders it suitable for decomposition by activated sludge treatment (Hoondal et al.,
2000).
1.6.6 Paper and Pulp Industry
During papermaking, pectinase can deploymerize pectins and subsequently lower the cationic
demand of pectin solutions and the filtrate from peroxide bleaching (Reid and Richard, 2004;
Viikari et al., 2001).
1.6.7 Animal Feed
Pectinases are used in the enzyme cocktail, used for the production of animal feeds. This reduces
the feed viscosity, which increases absorption of nutrients, liberates nutrients, either by
hydrolysis of non-biodegradable fibers or by liberating nutrients blocked by these fibers, and
reduces the amount of faeces (Hoondal et al., 2000).
1.6.8 Purification of Plant Viruses
In cases where the virus particle is restricted to phloem, alkaline pectinases and cellulases can be
used to liberate the virus from the tissues to give very pure preparations of the virus (Salazar and
Jayasinghe, 1999).
1.6.9 Improvement of Chromaticity and Stability of Red Wines
Pectinolytic enzymes added to macerated fruits before the addition of wine yeast in the process
of producing red wine resulted in improved visual characteristics (color and turbidity) as
compared to the untreated wines. Enzymatically treated red wines presented chromatic

characteristics, which are considered better than the control wines. These wines also showed
greater stability as compared to the control (Revilla and Ganzalez-Sanjose, 2003).
1.7 Substrates for the Production of Pectinases
Substrates that are employed in the production of enzyme should be solid as solid substrate can
encourage the growing cells. Substrates should provide all needed nutrients to the
microorganisms for its growth.
Other factors like particle size, moisture levels are also to be taken for consideration. Generally
agro-industrial wastes are employed for the pectinase production. Various substrates that are
being used are sugarcane bagasse, wheat bran, rice bran, wheat straw, rice straw, sawdust, corn
cobs, coconut coir pith, banana waste, tea waste, sugar beet pulp, apple pomace, orange peel,
pineapple peels etc (Pilar et al., 1999).
1.8 Fermentation Conditions
Pectinases are constitutive or inducible enzymes that can be produced either by submerged
(Aquilar and Huitron, 1999) or solid state fermentation (Acuna-arguelles et al., 1995).Various
factors related to environment affect the production of pectinase. Some of them are concentration
of nutrients, pH, temperature, moisture content, influence of extraction parameters on recovery
of pectinases and the effects played by the inducers. Both carbon and nitrogen sources show
overall effect on the productivity of pectinases (Catarina et al., 2003; Almeida and Huber, 2011).
Pectin, glucose and sucrose when added to the media in higher concentration have a repression
effect on the studied enzyme activity (Maria et al., 2000) of the various nitrogenous matters that
can be used. Optimum sources are (NH4)2SO4, yeast extract, soya bean pulp powder, soya
peptone.
Temperature and pH are also important parameters to be taken note of. The pH is regulated using
a mixture of sources of nitrogen as when Aspergillus niger is being used, pH turns to be acidic.
Besides the nature of the substance also plays a vital role in the pH maintenance. Moisture
content in the substrate also plays a significant role (Martin et al., 2004). The previous studies

show that it was generally maintained around 50-55% for the production of pectinases by
microbial means (Leda et al., 2000).
Two types of fermentations can be carried out for pectinase production, they are solid state
fermentation and submerged fermentation. The growth of organisms is very high with large
quantities of enzyme being produced in solid- state fermentation (Ramanujam and Saritha,
2008). However in the production of extracellular pectinases, submerged fermentation is
preferable as the extracellular pectinases are easier and cheaper to use in great quantities. Most
important applications of these enzymes are in juice and wine making, and in the processing of
vegetables. Submerged or solid state mediums are used for producing of the pectinolitic enzymes
by fungi (Bali, 2003).
1.8.1 Types of Fermentation
i) Solid State Fermentation (SSF)
ii) Submerged Fermentation (SmF)
Solid state fermentation is defined as the cultivation of microorganisms on moist solid supports,
either on inert carriers or on insoluble substrates that can be used as carbon and energy source.
This process occurs in the absence or near absence of free water in the space between substrate
particles. In this system, water is present in the solid substrate whose capacity for liquid retention
varies with the type of material (Lonsane et al., 1985; Pandey et al., 2001).
Submerged liquid fermentation is the cultivation of microorganisms in liquid nutrient broth.
Industrial enzymes can be produced using this process. This involves growing carefully selected
microorganisms (bacteria and fungi) in closed vessels containing a rich broth of nutrients (the
fermentation medium) and a high concentration of oxygen (Grigelmo-Migeul and Martin-
Belloso, 1998).
SSF has the following advantages over SmF:
i) Low water demand which leads to less waste water production.
ii) Utilization of solid substrates that enhances the concentration of the growth substrates.
iii) Utilization of otherwise unusable carbon sources that is abundant in supply.

However, there are also several disadvantages of SSF which have discouraged the use of this
technique for industrial production and therefore have made SmF more applicable in the
production of enzymes. These include: the buildup of gradients of temperature, pH, moisture,
substrate concentration or CO2 during cultivation which are difficult to control under limited
water availability (Holker et al., 2004).
1.9 Microorganisms Commonly Used in Submerged and Solid State Fermentation for
Pectinases Production
Microorganisms are currently the primary source of industrial enzymes: 50% originate from
fungi and yeast; 35% from bacteria, while the remaining 15% are either of plant or animal origin.
Filamentous microorganisms are most widely used in submerged and solid-state fermentation for
pectinase production. Ability of such microbes to colonize the substrate by apical growth and
penetration gives them a considerable ecological advantage over non-motile bacteria and yeast,
which are less able to multiply and colonize on low moisture substrate (Smith and Aidoo,
1988). Among filamentous fungi three classes have gained the most practical importance in SSF;
the phycomycetes such as genera Mucor, the ascomycetes genera Aspergillus and
Basidiomycetes especially the white and rot fungi (Young et al., 1983). Bacteria and yeasts
usually grow on solid substrates at the 40%to70% moisture levels (Young et al., 1983). Common
bacteria in use are (Bacillus licheniformis, Aeromonas cavi and Lactobacillus) and common
yeasts in use are Saccharomyces and Candida. Pectinase production by Aspergillus strains has
been observed to be higher in solid-state fermentation than in submerged process (Solis et al.,
1996).

TABLE 7: Comparison of Solid State and Submerged Fermentation
Factor Submerged Fermentation
(SmF)
Solid State Fermentation (SSF)
Substrate Soluble Substrate (sugars) Polymer insoluble substrate:
Starch, Cellulose, Pectins
Aseptic Conditions Heat sterilization and aseptic
Control
Vapor treatment, non-sterile
conditions
Water High volumes of water
consumed and effluents
discarded
Limited consumption of water
Metabolic Heating Easy control of temperature Low heat transfer capacity
pH Control pH control Buffered solid substrate
Source: (Raimbault, 1998)
1.10 Aim and Objectives of the Study
1.10.1 Aim of the Study
To extract pectin from pineapple peels
To isolate Aspergillus niger from soil containing decomposing pineapple peels.
To produce extracellular pectinase by inducing Aspergillus niger in submerged fermentation
with pectin extracts from pineapple peels.
To partially purify extracellular pectinase by carrying out ammonium sulphate precipitation
and dialysis.
To characterize the partially purified enzyme
1.10.2 Specific Objectives of the Study
To determine the effect of change in pH on pectinase activity.
To determine the effect of change in temperature on pectinase activity.
To determine the Km and Vmax of the enzyme from the Lineweaver-Burk plot.

CHAPTER TWO
MATERIALS AND METHODS
2.1 MATERIALS
2.1.1 Chemicals/ Reagents
All the chemicals used in this research work were of analytical grade and were obtained from
Sigma Chemical Company, USA; Sigma-Aldrich, USA; Bio Rad Laboratories, India; Merck,
Germany; BDH Chemical Ltd, England; May and Baker Ltd, England; Riedel-De Haen
Hannaves, Germany; Hopkins and Williams Essex, England and Lab Tech. Chemicals Avighkar,
India.
2.1.2 Apparatus/ Equipment
Autoclave: UDAY BURDON‟s Patent Autoclave, made in India.
Centrifuge: Finland Nigeria 80-2B.
Glass wares: Pyrex
Incubator: B and T Trimline incubator.
Magnetic stirrer: AM-3250B Surgi Friend Medicals, England.
Microscope: WESO microscope.
Milling machine: Thomas Willey laboratory Mill Model 4, Anthor H (Thomas
Company, Philadelphia, USA).
Oven: Gallenkamp Hotbox, made in England.
pH meter: Ecosan pH meter, made in Singapore.
UV/ spectrophotometer: Jenway 6405
Water bath: Model DK.
Weighing balance: B2404-5 mettler Toledo, made in Switzerland.
Weighing balance: Ohaus Dial-O-Gram, Ohaus Co-operation, N.J. USA.
2.1.3 Collection of Pineapple Samples
Pineapple (Ananas cosmosus) peels were obtained from Ogige market, Enugu State, Nigeria.

2.1.4 Collection of Micro-organisms
Mixed colonies of microorganisms were obtained from a dump containing decaying pineapple
peels and Aspergillus niger was isolated using morphological characteristics.
2.2 METHODS
2.2.1 Preparation of Reagents
2.2.1.1 Preparation of 3N HCl
Normality of the stock HCl in Winchester bottle was calculated using the formulae outlined
below:
Normality (N) = % assay x 1000 x specific gravity
100 x Equivalent weight
For HCl, % assay = 37, specific gravity = 1.19 and equivalent weight (in this case) = 36.5. The
volume of the stock solution required to prepare 3N HCl was calculated using the formula
N1V1 = N2V2
Where: N1 = Normality of stock HCl,
V1 = volume of stock HCl required for the preparation,
N2 = required normalty (3N),
V2 = working volume (1000ml).
N.B: V1 obtained from the calculation was diluted in 1000ml of diluent (water) to make the
required 3N HCl.
2.2.1.2 Preparation of Ethanol-HCl Solution
The ethanol-HCl solution contained 0.5M HCL prepared using similar formula as that described
above except that molecular weight was used instead of equivalent weight in order to obtain the
molarity of stock HCl. The volume of the stock solution required to prepare 0.5M HCl was
calculated using the formula:
C1V1 = C2V2;
where C1= Molarity of stock HCl,

V1 = volume of stock HCl required for the preparation,
C2 = required molarity (0.5M),
V2 = working volume (1000ml).
N.B: V1 obtained from the calculation was diluted in 1000ml of diluent (ethanol) to make the
required ethanol-HCl solution.
2.2.1.3 Preparation of Buffers
The standard buffers used in this study were pH 4.0 and pH 7.0. These buffers were used to
standardize the pH meter. The working buffers were prepared by this procedure.
Sodium acetate buffer of 0.05M and Tris-HCl buffer of 0.05M were prepared by dissolving
4.10g sodium acetate salt and 6.01g Tris base, respectively in 1000ml of distilled water and
stirred with a magnetic stirrer till a homogenous solution was formed. The solutions were titrated
against acetic acid and HCl, respectively till the required pH was obtained.
Phosphate buffer of 0.05M was prepared by dissolving 7.10g disodium hydrogen phosphate salt
in 1000ml stirred as well with a magnetic stirrer and then titrated against the solution of its
conjugate acid which is sodium di-hydrogen phosphate till the required pH was obtained.
2.2.1.4 Preparation of 2mg/ml Bovine Serum Albumin (BSA) Standard Protein
An amount of 0.2g of BSA was dissolved in 100ml of distilled water and then used as a protein
stock solution.
2.2.1.5 Preparation of Dinitrosalicylic Acid (DNS) Reagent
A modification of DNS reagent method of Miller (1959) as contained in Wang et al. (1997) was
used in the assay. The reagent contains 44mM dinitrosalicylic acid, 4mM sodium sulphite, and
375mM sodium hydroxide.

2.2.1.6 Preparation of 20mM Galacturonic Acid
An amount of 0.42g D-(+)-Galaturonic acidmonohydrate (molecular weight 212.15g/mole) was
dissolved in 100ml 0.05M sodium acetate buffer stirred over a magnetic stirrer until a
homogenous stock solution was obtained.
2.2.1.7 Preparation of the Component Reagents for Protein Determination
Solution A: An alkaline sodium carbonate (Na2CO3) was prepared by dissolving 2g of Na2CO3
in 100ml of 0.1M NaOH (0.4g of sodium hydroxide pellets were dissolved in 100ml of distilled
water).
Solution B: A copper tetraoxosulphate VI - sodium potassium tartarate solution was prepared by
dissolving 0.5g of CuSO4 in 1g of sodium potassium tatarate, all in 100ml of distilled water. It
was prepared fresh by mixing stock solution, and so was done whenever required.
Solution C: Folin-Ciocalteau phenol reagent was made by diluting the commercial reagent with
water in a ratio of 1:1 on the day of use.
Solution D: Standard protein (Bovine Serum Albumin, BSA) solution.
Solution E: Freshly prepared alkaline solution was made by mixing 50ml of solutions A and 1ml
of solution B.
2.2.2 Preparation of Ground Pineapple Peels
The pineapple peels were washed and cut into small bits and then treated with hot 96% ethanol
in order to reduce the microbial load. The ethanol treated peels were washed with water and sun-
dried for 7 days. The dried peels were then ground to powder using a milling machine.
2.2.3 Extraction of Pectin from Pineapple Peels
Pectin was extracted by the method of McCready, (1970). 100g of ground pineapple peels was
poured into a 2000ml beaker containing 800ml of distilled water, then 12g of freshly ground
sodium hexa-metaphosphate was added to the mixture and the initial pH was adjusted with 3N

HCL to 2.2 ± 0. The mixture was heated up in a water bath at 70 ºC for 1 hr and stirred
continuously using a propeller typed stirrer.
The pH was checked at an interval of 15 minutes. The water lost by evaporation was replaced
except in the last 20 minutes of the extraction time. The extract was vacuum-filtered using a
muslin cloth and the residue was washed with 200ml of distilled water. The washings were
however added to the filtrate and then concentrated by evaporation on a hot plate to
approximately one-fifth of its initial volume.
The concentrated pectin mixture was cooled to 50 ºC and a volume of ethanol containing 0.5M
HCL in the ratio of 3:1 to the pectin mixture was added, it was stirred continuously for 30
minutes and allowed to stand for 1 hour. The precipitate was vacuum filtered and washed with
acetone in order to remove traces of HCL and ethanol. The extract was dried in an oven at 40 ºC
for a few hours and ground to powder.
2.2.4 Isolation of Pectinolytic Fungi
2.2.4.1 Collection of Soil Samples
Soil samples were collected from site containing decomposing pineapple peels. This compost
site was located within the school premises of the University of Nigeria Nsukka, Enugu state.
The soil samples were collected in a clean dry plastic container.
2.2.4.2 Preparation of Soil Sample Extracts for Microbial Isolation.
Soil samples from site of decomposing pineapple peels were homogenized in sterile medium
containing 1% pineapple pectin, 0.14% of (NH4)2SO4, 0.2% of K2HPO4, 0.02% of MgSO4.7H2O
& 0.1% of nutrient solution containing 5mg/L of FeSO4.7H2O,1.6mg/L of MnSO4.H2O, 1.4mg/L
of ZnSO4.7H2O, 2.0mg/L of CoCl2.The mixture was incubated at 30ºC for 24hrs.
2.2.4.3 Preparation of the Solid Medium
The mixture contained 1% pineapple pectin, 0.14% of (NH4)2SO4, 0.2% of K2HPO4, 0.02% of
MgSO4.7H2O & 0.1% of nutrient solution containing 5mg/L of FeSO4.7H2O,1.6mg/L of
MnSO4.H2O, 1.4mg/L of ZnSO4.7H2O, 2.0mg/L of CoCl2 and 3% agar-agar (the gelling agent)
(w/v).

The medium was autoclaved at 121oC for 15min. It was allowed to cool to about 45
oC and then
poured into Petri dishes and allowed to gel. The plates were then incubated in a B & T Trimline
incubator at 37oC overnight to check for sterility.
2.2.4.4 Sub-culturing into Solid Medium
A loop of homogenized extract from the sample extracts was streaked onto the solid medium
under the flame of Bunsen burner. The plates were incubated at 35oC (room temperature) till
visible colonies were observed. All morphological contrasting colonies were purified by repeated
streaking and sub-culturing on separate plates. This process was continued till pure fungal
cultures were obtained.
2.2.4.5 Storage of Micro-organisms on Potato Dextrose Agar (PDA)
Pure fungal isolates were maintained on PDA slants as stock cultures. The PDA medium was
prepared according to the manufacturer‟s instructions.
2.2.4.6 Microscopic Features of the Isolated Fungi
Three days old pure cultures were examined; the color, texture, nature of mycelia or spores and
growth patterns were also observed. Photographs of the culture were also taken.
2.2.4.7 Fungal Identification
Three days old cultures were used in preparing microscopic slides. A tuft of the mycelia was
dropped on the slide and a drop of lacto-phenol blue was added to it. A cover slip was placed
over it and viewed under a light microscope at ×400 magnification. Identification was carried out
by relating the microscopic features and the micrographs to „Atlas of Mycology‟ by Barnett and
Hunter (1972).
2.2.5 Fermentation Experiments
2.2.5.1 Preparation of the fermentation medium
The substrate for fermentation consisted of ground pineapple pectin extracts. Submerged
fermentation was carried out using 10 250ml Erlenmeyer flask containing 200 ml of the sterile

cultivation medium. The medium was optimized for pectinase production with 0.1% NH4NO3,
0.1% NH4H2PO4, 0.1% MgSO4.7H2O and 20g of powdered pineapple pectin extracts. The flask
was covered with aluminium foil and autoclaved at 121ºC for 20mins.
2.2.5.2 Innoculating with Aspergillus niger
In every sterile flask, two disc of Aspergillus niger obtained from the freshly prepared plates are
added using a cork borer of diameter 10mm under sterile conditions. The flasks are plugged
firmly and incubated for four days at room temperature.
2.2.5.3 Harvesting the crude enzyme
At the end of day 4 on which the highest enzyme activity has been detected, the mycelia biomass
was filtered using filter paper. 2.0 litres of filtrate recovered was used as the crude enzyme, while
the residue was treated with lime and discarded properly.
2.2.6 Galacturonic Acid Standard Curve
The reaction mixture contained 0.0-1.0ml of galacturonic acid stock solution in test tubes
arranged in triplicates. Each test tube was made up to 1ml using freshly prepared 0.05M sodium
acetate buffer of pH 5.0. 1ml of DNS reagent was added to each of the test tubes and placed in a
boiling water bath for 10min. 1ml of 1.4M Rochelle salt (sodium potassium tartarate) was added
to the test tube immediately after heating and the total volume of the solution was adjusted to
4ml with distilled water. The mixture was cooled to room temperature and the absorbance read at
575nm. The concentration of reducing sugar in each of the tubes was calculated using the
formula
C1 V1 = .C2 V2
Where: C1 = initial concentration of reducing sugar (mM)
C2 = final concentration of reducing sugar (mM)
V1 = initial volume of 20mM galacturonic acid preparation measured into the tube
V2 = final volume of the preparation.

Using the values obtained from above the calculations, the plot of optical density was
constructed and the concentration of galacturonic acid released at a given absorbance was
extrapolated (Appendix Two).
2.2.7 Polygalacturonase Assay
Polygalacturonase activity was determined by measuring the release of reducing sugars from the
pineapple pectin using the 3,5- dinitrosalicyclic acid reagent assay as described by Miller,
(1959).
2.2.7.1 Procedure for Polygalacturonase Assay
This was carried out by the method described by Miller, (1959) as contained in Wang et al.,
(1997) with slight modifications. The reaction mixture contained 0.5ml of 0.5% pineapple pectin
in 0.05M sodium acetate buffer pH 5.0 and 0.5ml of the crude enzyme solution. After one hour
incubation time, 1ml of DNS reagent was added and the reaction was stopped by boiling for
10mins at 70ºC.
The total volume was brought up to 4ml by adding 1ml of rochelles salt and 1ml of distilled
water. The reaction mixture was allowed to cool and then absorbance was read at 575nm. One
unit of enzyme was defined as the amount of enzyme that catalyzes the formation of 1
micromole of galacturonic acid per minute.
2.2.8 Protein Determination
Protein content of the enzyme was determined by the method of Lowry et al. (1951), using
Bovine Serum Albumin as standard.
2.2.8.1 Procedure for Protein Determination
For the reaction mixture, test tubes were arranged in duplicates containing 0.0- 1.0 ml of 0.2mg
of protein stock solution (2mg/ml BSA) and brought up to 1ml with distilled water. For the test
mixture, 0.5ml of sodium acetate buffer pH 5.5 was added to 0.5ml of the crude enzyme.

To both the reaction and test mixture, 5 ml of solution D was added soon after and the mixture
was allowed to stand for 10 mins. 0.5ml of Solution C (dilute Folin-ciocalteau reagent) was
added and then the solution was mixed thoroughly and allowed to stand for 30mins under room
temperature. The absorbance was read at 750nm and the protein concentration was determined
thereafter.
2.2.9 Partial Purification of Protein
2.2.9.1 Ammonium Sulphate Precipitation Profile
This procedure is carried out in order to know the percentage of ammonium sulphate
concentration is suitable to precipitate the most protein from the crude enzyme. Nine test tubes
were used containing 10ml of the crude enzyme and the enzyme was precipitated from 20% -
100% saturation of solid ammonium sulphate at 10% interval in each test tube.
The contents of the tubes were mixed thoroughly to ensure that the salts were dissolved and then
allowed to stand for 30hrs at 4ºC. The test tubes were centrifuged at 3500 rpm for 30 mins and
the filtrates were decanted while the pellets were re-dissolved in equal volume of 0.05M sodium
acetate buffer pH 5.0. Pectinase activity was determined on the contents of each tube.
2.2.9.2 Ammonium Sulphate Precipitation
After determining the percentage saturation of ammonium sulphate salts that gave the highest
activity, the equivalent amount of salt for 1 litre of crude enzyme is added. The salt is allowed to
dissolve completely and the mixture is allowed to stand for 30 hrs at 4ºC. It is then centrifuged at
3500 rpm for 30mins. The pellets are collected and stored in a cool place for further studies.
2.2.9.3 Dialysis
Dialysis tubes stored in 90% ethanol were used. However the tubes were rinsed thoroughly with
distilled water and finally with 0.05M sodium acetate buffer in order to remove traces of ethanol.
An amount of the precipitated enzyme is poured into the dialysis tubes and placed in a beaker
containing 0.05 M sodium acetate buffer. The beaker is placed on a magnetic stirrer which
allows for a homogenous environment. The dialysis is carried out according to Dixon and Webb

(1964) for 12 hours and the buffer is changed after 6 hours which allows for the exchange of low
molecular weight substances and left over ammonium sulphate salts that may interfere with the
activity. After dialysis, the partially purified enzyme is stored frozen at -24ºC.
2.2.10. Studies on Partially Purified Enzyme
2.2.10.1 Effect of pH Change on Pectinase Activity
The effect of pH on enzyme activity was determined using 0.05M sodium acetate buffer pH 3.5
- 5.5, phosphate buffer pH 6.0 - 7.5 and Tris-HCl buffer pH 8.0 - 10.0 at intervals of 0.5. 0.1%
pectin solution was prepared by dissolving 0.1g pectin in 100ml of 0.05M of the respective
buffers. Also 0.5 ml of the partially purified enzymes was added to 0.5 ml of each of the buffers.
Then ultimately, 0.5ml of each of the enzyme-buffer solution was mixed with 0.5ml pectin
solution at the corresponding pH for pectinase assays as described previously.
2.2.10.2 Effect of Temperature Change on Pectinase Activity
The optimum temperature was determined by incubating the enzyme with pectin solution at 25-
70oC interval of 5
oC for 1hour and at the pH with the highest activity. The activity was then
determined as described in previously.
2.2.10.3 Effect of Substrate Concentration on Pectinase Activity.
The effect of substrate concentration on the activity of pectinase was determined by incubating
the enzyme with 0.5 up to 6.0mg/ml of pineapple pectin at an interval of 0.5 using the buffer at
the pH with highest activity and the temperature at which highest activity was determined. The
Vmax and Km values of the enzyme were determined using the double reciprocal plot.
2.2.10.4 Further Studies with Partially Purified Enzyme
Pectinase activity and total protein concentration of partially purified enzymes were determined
using the methods described in sections 2.2.7.1 and 2.2.8.1 respectively.

CHAPTER THREE
RESULTS
3.1 Pineapple Pectin Extraction
3.1.1 Percentage Extraction Yield of Pineapple Pectin
Pectin extraction yield was found to be 8.33% at pH 2.2, temperature of 70oC and extraction time
of 1hr.
3.1.2 Photograph of Pineapple Pectin Extract
Fig. 10 shows a photograph of the pineapple pectin after extraction.
Fig. 10: Photograph of Pineapple Pectin

3.2 Microorganism
3.2.1 Selection of Pectinolytic Fungi
Three species of fungi namely: Aspergillus niger, Aspergillus fumigatus and Aspergillus flavus
were isolated from natural source of soil containing decaying pineapple peels. These organisms
were qualitatively screened for pectinolytic activity on selective media and their isolation was
based on the similarities of their morphological features in both test cultures containing
pineapple pectin and the standard culture containing apple pectin as carbon respectively.

3.2.2 Macroscopic and Microscopic Features of Fungal Isolates
Genus identification was by examining both macroscopic and microscopic features of a three day
old pure culture. Color, texture, nature of mycelia and/or spores produced, growth pattern in
addition to microscopic features such as separation and spore shapes were examined. Based on
these characteristics, Aspergillus niger, Aspergillus fumigatus and Aspergillus flavus were
confirmed as the three pectinolytic fungal isolates, respectively. However, Aspergillus niger
showed relatively higher pectinase activity and was selected for further studies. Fig. 11 shows a
photograph of the pure culture of Aspergillus niger.
Fig. 11: Pure Culture of Aspergillus niger

3.3 Pectinase Production under Submerged Fermentation System
A volume of 2 litres of crude enzyme was harvested after 4 days of submerged fermentation
using Aspergillus niger.

3.4 Ammonium Sulphate Precipitation
Crude enzyme obtained from Aspergillus niger was precipitated within the range of 20-100% at
an interval of 10%. At 80% ammonium sulphate saturation, the highest activity was obtained at
81.62U/ml as shown in Fig. 12; thus, the percentage was used for the precipitation of pectinases.
Fig. 12: Ammonium sulphate precipitation profiling of pectinases obtained from Aspergillus
niger.

3.5 Assays Carried Out on Pectinase Obtained
3.5.1 Protein Concentration of the Crude, Precipitated and Dialyzed Pectinases
In Fig 13, initial protein concentration of pectinase in the crude state was 10.42mg/ml but after
ammonium sulphate precipitation and dialysis, the concentration increased to 15.82mg/ml and
later decreased to 12.84mg/ml.
Figure 13: Comparison in the protein concentration of the crude, precipitated and dialyzed
pectinases

3.5.2 Activity of Pectinase in the Crude, Precipitated and Dialyzed Enzyme
The activities of the enzymes increased progressively from one purification step to the other with
the corresponding values of 116.12U/ml, 272.98U/ml and 634.56U/ml respectively as shown in
Fig. 14.
Figure 14: Comparison in the activities obtained from the crude, precipitated and dialyzed
pectinases
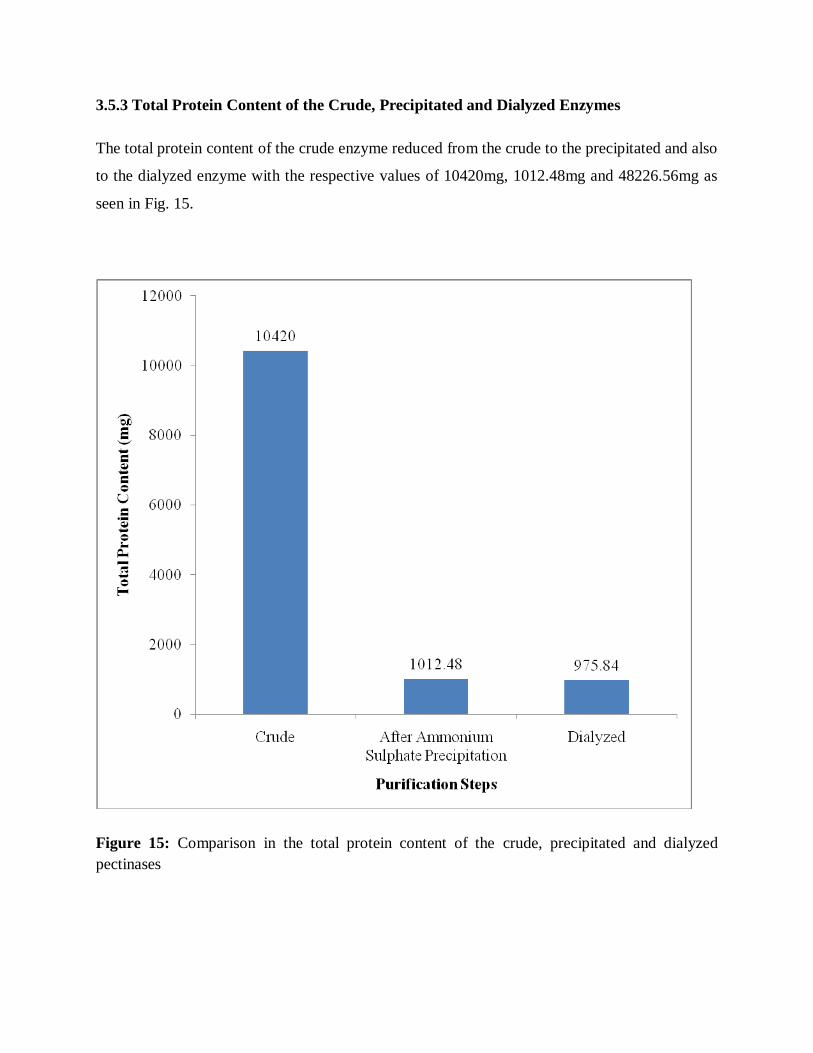
3.5.3 Total Protein Content of the Crude, Precipitated and Dialyzed Enzymes
The total protein content of the crude enzyme reduced from the crude to the precipitated and also
to the dialyzed enzyme with the respective values of 10420mg, 1012.48mg and 48226.56mg as
seen in Fig. 15.
Figure 15: Comparison in the total protein content of the crude, precipitated and dialyzed
pectinases

3.5.4 Total Activity of the Crude, Precipitated and Dialyzed Enzymes
Fig. 16 shows activity-dependent decrease from the crude enzyme across precipitated enzyme to
after dialysed sample. The crude enzyme had the highest total enzyme activity of 116120U with
after dialysed sample having the least enzyme activity of 4822.56U.
Fig. 16: Comparison in the total activity of the crude, precipitated and dialyzed pectinases

3.5.5 Specific Activity Determined on the Crude, Precipitated and Dialyzed Enzymes
Specific activity-dependent increase was observed in Fig. 17 with crude enzymes having the
lowest activity and after, dialyzed sample had the highest enzyme activity with values of
11.14U/mg and 49.42U/mg respectively. This further explains that the enzyme was in a purer
state after dialysis than after ammonium sulphate precipitation.
Fig. 17: Comparison in the Specific Activities of the crude, precipitated and dialyzed pectinases

3.5.6 Purification Folds of the Partially Purified Enzymes
As shown in Fig. 18, after dialysis and ammonium sulphate precipitation, the purification fold
increased from 1.55 after ammonium sulphate precipitation and further increased to 4.43 after
dialysis which infers that the enzyme had undergone a 4.43 purification fold.
Fig. 18: Purification folds of the pectinase after precipitation and dialysis

Table 8: Summary of the Parameters Determined From the Crude, Precipitated and Dialyzed
Pectinases
Purification
Step
Volume
(ml)
Protein
Conc.
(mg/ml)
Activity
(U/ml)
Total
Protein
(mg)
Total
Activity
(U)
Specific
Activity
(U/mg)
Purification
fold
Crude
Enzyme
1000 10.42 116.12 10420 116120 11.14 1
80%
ammonium
sulphate
precipitation
64
15.82
272.98
1012.48
17470.72
17.26
1.55
Dialysed
Enzyme
76 12.84 634.56 975.84 48226.56 49.42 4.43
µmole/min= Unit (U)
According to Table 8, the crude state of pectinase had the least value for activity of 116.12U/ml
unlike the precipitated and dialyzed, which were 272.98U/ml and 634.56U/ml respectively. Also,
the protein concentration increased after ammonium sulphate precipitation due to the precipitated
proteins. However, the protein concentration reduced after dialysis to 12.84mg/ml. Thus it can be
said that protein concentration is inversely relative to activity.

3.6. Characterization of Pectinase
3.6.1 Effect of pH Change on Pectinase Activity
Fig. 19 shows that the highest pectinase activity was recorded at pH 5.5. Also, observed in Fig.
19 was a decline in activity when pH was increased or decreased beyond 5.5.
Figure 19: Showing the effect of change in pH on pectinase activity

3.6.2 Effect of Change in Temperature on Pectinase Activity
Fig. 20 shows gradual decline of the enzyme activity at a temperarture of 40˚C with subsequent
rise at temperature of 45 and 50˚C. The peak of the enzyme activities was observed at a
temperature of 55˚C accompanied with a sharp drop of the enzyme activity with corresponding
rise in temperature.
Figure 20: Effect of Change in Temperature on Pectinase Activity
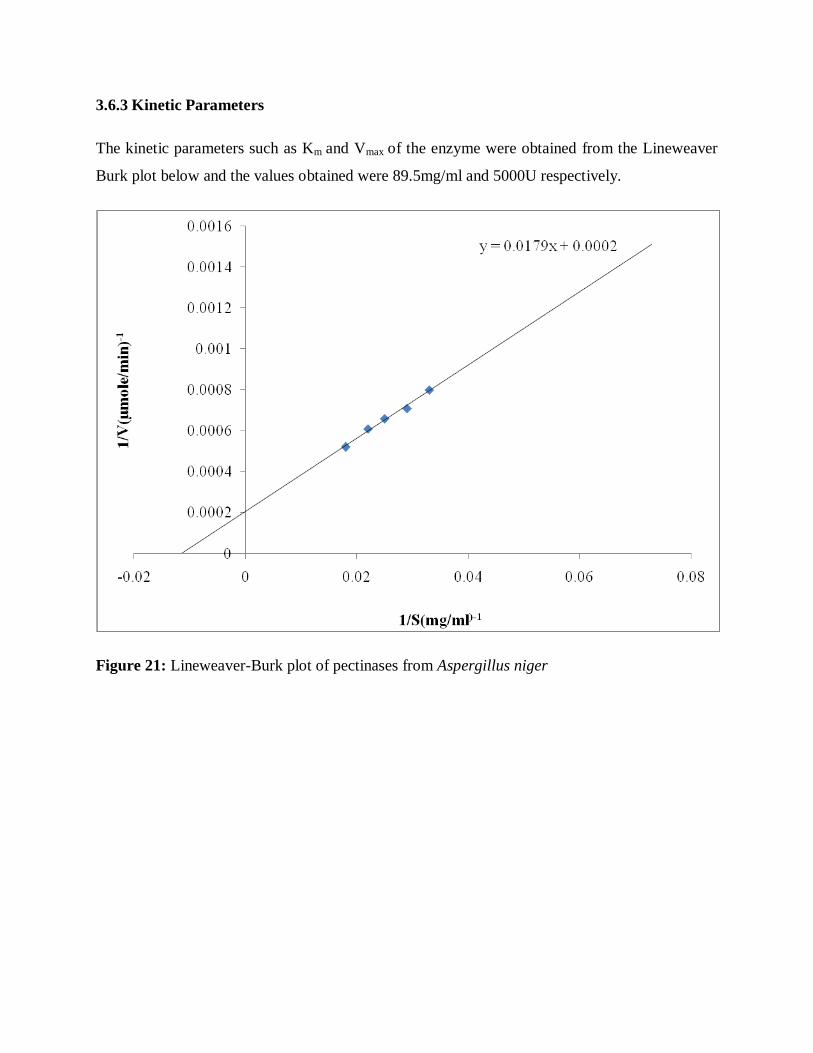
3.6.3 Kinetic Parameters
The kinetic parameters such as Km and Vmax of the enzyme were obtained from the Lineweaver
Burk plot below and the values obtained were 89.5mg/ml and 5000U respectively.
Figure 21: Lineweaver-Burk plot of pectinases from Aspergillus niger

Table 9: Pectinase characterization
Properties Aspergillus niger
pH 5.5
Temperature (°C) 55°C
Vmax(U/ml) 5000U/ml
Km(mg/ml) 89.5mg/ml
Table 9 shows the characterization of the pectinase obtained from Aspergillus niger with the pH
and temperature optima being 5.5 and 55˚C respectively. The Km and Vmax were 89.5mg/ml and
5000U/ml respectively.

CHAPTER FOUR
DISCUSSION
4.1 Discussion
Pineapple (Ananas cosmosus) peels as agricultural wastes represent about 35% of the fruit mass.
During extraction of pectin from pineapple peels, the yield of pectin extracted was 8.33% at pH
2.2, temperature of 70ºC and extraction time of 1hr using the method as described by McCready
(1970). The yield could be affected by the pH of the extraction medium and extraction time.
Three fungal species isolated from natural waste source were selected which include Aspergillus
niger, Aspergillus fumigatus and Aspergillus flavus. However, Aspergillus niger (pure culture as
shown in Fig. 11) was used for the fermentation process because it showed the highest pectinase
activity after 4 days of fermentation after which a decline in activity was observed. This may be
attributed to the depletion of the nutrients and pectin in the fermentation medium which could
result in decrease in enzyme or protein synthesis over time (Haq et al., 2005) since Aspergillus
niger produces extracellular enzymes.
In the present study, accumulation of maximal extracellular pectinase was observed after 96hour
of fermentation. The period of fermentation depends on the nature of medium, fermenting
organisms, concentration of nutrients and the process physiological conditions (Patil and
Dayanand, 2006). Substrates should provide all needed nutrients to the micro-organism for its
growth. Various substrates being used are sugarcane bagasse, rice bran, banana wastes, tea
waste, pineapple peels, apple pomace, etc, (Pilar et al., 1999). Although an increased level in the
production of pectinases was noticed when the agro wastes were supplemented with additional
carbon and nitrogen sources and supplementation of sucrose was more effective than glucose in
solid state fermentation (Patil and Dayanand, 2006). A 96hour incubation time was also obtained
in the poly-galacturonase production in solid state fermentation (Martin et al., 2004).
Ramanujam et al. (2008) recorded an 84hour incubation time for the optimum production of
pectin lyase by Aspergillus niger in solid state fermentation which is comparable to what was
obtained.

In this research, submerged fermentation technique was applied due to the fact that temperature,
pH, moisture, substrate concentration or CO2 during cultivation would have been difficult to
control under limited water availability. (Holker et al., 2004). Submerged liquid fermentation is
the cultivation of microorganisms in liquid nutrient broth. Industrial enzymes can be produced
using this process. Submerged fermentation involves growing carefully selected microorganisms
(bacteria and fungi) in closed vessels containing a rich broth of nutrients (the fermentation
medium) and a high concentration of oxygen (Grigelmo-Migeul andMartin-Belloso, 1998).
Ammonium sulphate precipitation profile was carried out on the crude enzyme and the activities
of the pellets obtained from the different percentages of saturation from 20%-100% was
determined. Fig. 12 shows that 80% had the highest activity and was therefore used for the actual
precipitation process of the enzyme. Buga et al. (2010) reported 70% ammonium sulphate
saturation for pectinase from Aspergillus. niger (SA6) while Adejuwon and Olutiola, 2007
reported 90% ammonium sulphate saturation for pectinase from Lasidioplodia theobromae.
After the protein was precipitated, one way to remove this excess salt is to dialyze the protein.
Also it changes the buffer composition of solutions of biomolecules too large to pass through the
membrane (Rosenberg, 2004). It was observed that there was an increase in volume of the
enzyme after dialysis which was done for 12 hours; this may be due to the buffer that entered
from the dialyzing medium during the process.
The protein concentration increased from 10.42 to 15.82 mg/ml after ammonium sulphate
precipitation and then there was a decrease in the concentration to 12.84mg/ml after dialysis as
seen in Fig. 13. This may be due to the removal of other proteins of lower molecular weight
during dialysis that was not the protein of choice.
Fig. 14 shows the pectinase activity which increased from 116.12 to 272.98 U/ml after
precipitation and further increased to 634.56 U/ml after dialysis. This may be due to the removal
of impurities during dialysis such as other proteins which may have affected the enzyme activity
negatively (Lukong et al., 2007).
The specific activity of the crude, precipitated and dialyzed were determined and it showed that
there was an increase after each purification step as shown in Table 8 from 11.14 U/mg in the

crude to 17.26 U/mg in the precipitated and then to 49.42 U/mg in the dialyzed. This is in
agreement with the report by Lukong et al. (2007) that for a purification procedure to be
successful, the specific activity of the desired enzyme must be greater after the purification
procedure than as it was before. The increase in specific activity is a measure of purification
achieved thus the enzyme had a 4.43 purification fold as purification fold increased from 1.54
after ammonium sulphate precipitation to 4.43 after dialysis as observed in Fig. 18.
The partially purified enzyme was characterized based on the effect of change in pH,
temperature and substrate concentration. The maximal activity was observed at pH 5.5 as seen in
Fig. 19 for pectinase from Aspergillus niger using pineapple peels as substrate which is
comparable to the optimum pH for pectinase activity from thermo-tolerant Aspergillus sp N12
(Freitas et al., 2006). Changes in pH can change the shape of the active site in an enzyme.
Extremely high or low pH concentrations usually result in complete loss of enzyme activity due
to denaturation (Helms et al., 1998). At extremely high and low pH values, the tertiary structure
of the protein (enzyme) may be disrupted and the protein denatured; even at moderate pH values,
where the tertiary structure is not disrupted, enzyme activity may depend on the alteration of
ionisation states of: certain amino acid side chains responsible for substrate binding, certain side
chains involved in catalysis and certain groups on the substrate (Lukong et al., 2007).
Temperature has a complex effect on enzyme activity. The "optimum temperature" of an enzyme
is the temperature in which the enzyme functions most efficient. An increase in temperature
below the optimum results in an increase in the kinetic energy of enzymes. This leads to higher
efficiency of enzyme-substrate complex formation; therefore a higher rate of reaction. It differs
in each enzyme, depending on its nature and structure. At extreme temperatures above the
optimum, the increased kinetic energy disrupts the bonds holding the active site; the enzyme is
unstable and the shape of its active site changes. This means that the enzyme is less efficient and
successful at enzyme-substrate complex formation. When the enzyme is said to be denatured; it
has lost its ability to catalyze reactions. Fig. 20 shows that 55˚C was the temperature at which
pectinase from Aspergillus niger exhibited the highest activity, which falls within the range as
exo-polygalacturonase from Monascus and Aspergillus sp. (Freitas et al., 2006) which exhibited
maximum activity at 60 and 50ºC respectively. It was also reported that polygalacturonase by
Aspergillus kawachii (Esquivel and Voget, 2004) and P. frequentans (Chellagatti et al., 2002)

exhibited maximum activity at 60ºC. At a higher temperature, the activity dropped and this could
be as a result of the denaturation of the enzyme. It could be that enzymes are proteins and are
heat sensitive, once temperatures have reached extreme heat, the enzymes denature, thereby
exposing the protein structure which leads to inactivation. All enzymes have an optimal
temperature at which reaction rates go fastest without denaturing the enzyme (Campbell and
Reece, 2002).
The Km and Vmax were calculated from the double reciprocal plot as shown in Fig. 21 and Km
was 89.5mg/ml while the Vmax was 5000U/ml. According to Anosike (2001), Km values provide
a parameter for comparing enzymes from different organisms. With the knowledge of Michaelis
Menten, it is known that a small Km indicates that the enzyme requires only a small amount of
substrate to become saturated; hence, the maximum velocity is reached at relatively low substrate
concentration while a large Km indicates the need for high substrate concentrations to achieve
reaction velocity. On the other hand, the Vmax or maximum velocity gives information on the
turnover number of an enzyme (Anosike, 2001). The turnover number of an enzyme is the
number of moles of substrate converted into product per active site of the enzyme per second,
when the enzyme is fully saturated with substrate which gives information on how efficient a
given enzyme is as a catalyst.
4.2 Conclusion
From this work, it was discovered that although pineapple peels have low pectin content of about
8.33%, it can be successfully used to induce the production of pectinase under submerged
fermentation system. The enzymes obtained can be industrially used in the production of fruit
juice, paper making, retting of plant fibres, etc. Ultimately, the rationale behind this research was
the conversion of waste to wealth which could increase the revenue base of any establishment or
country obtained and also geared towards a cleaner and safer environment.

4.3 Suggestions for Further Studies
The following suggestions for further studies are made based on the results obtained;
1) The organism used can be modified genetically in order to produce more enzymes and
probably on less number of days.
2) The biotechnological application of this process should be encouraged by individuals and the
governments since the advantages have been addressed.
3) More research works should be done on the production of pectinases from Aspergillus niger
using pineapple peels as carbon source. This inevitably makes their characterization more
available.

REFERENCES
Acuna-Arguelles, M.E., M.Gutierrez-Rojas, G.,Vinigra-Gonz-alez, E. and Favela-Torres. (1995).
Production and properties of three pectinoloytic activities produced by Aspergillus niger in
submerged and solid state fermentation. Applied microbiology and biotechnology, 43: 808-
814.
Adejuwon, A.O. and Olutiola, P.O. (2007). Pectin lyase activity in culture medium of
Lasidioplodia theobramae.Journal of Plant Sciences,2 (3): 334-340.
Alana, A., Alkorta, I., Dominguez, J.B., Llama, M.J. and Serra, J.L. (1990). Pectin lyase activity
in a Penicillium italicum strain. Applied Environmental Microbiology, 56: 3755–3759.
Albersheim, P., Neukom, H. and Deuel, H. (1960). Splitting of pectin chain molecules in neutral
solution. Archeology, Biochemistry and Biophysics, 90: 46–51.
Almeida, D.P.F. and Huber, D.J. (2011). Autolysis of cell walls from polygalacturonase-
antisense tomato fruit in simulated apoplastic solutions, Plant Physiologyof Biochemistry,
10: 1016
Alkorta, I., Garbisu, C., Llama, M.J. and Serra, J.L. (1998). Industrial applications of pectic
enzymes. A review: Process Biochemistry, 33: 21–28.
Anosike, E.O. (2001). Basic Enzymology. University of Port Harcourt Press, Pp 11-87.
Aquiler, G. and C.Huitron. (1999). Constitute exopectinase produced by Aspergillus sps ch-y-
043 on different carbon sources. Biotechnology, 12: 655-660.
Arenas-Ocampo, M.L., Evangelista-Lozano, S., Arana-Errasquin, R., Jime´nez-Aparicio, A. and
Da´vila-Ortíz, G. (2003). Softening and biochemical changes of sapote mamey fruit
(Pouteria sapota) at different development and ripening stages. Journal in Food Chemistry,
27: 91–95.
Arias, C. and Burns, J. (2002). A pectin methylesterase gene associated with a heat stable extract
from citrus. Journal in Agriculture and Food Chemistry, 50: 3465–3472.

Baker, R.A. and Wicker, L. (1996). Current and potential application of enzyme infusion in the
food industry. Trends in Food Science Technology, 7: 279– 284.
Bali, R. (2003). Isolation of extracellular fungal pectinoytic enzymes and extraction of pectin
using kinnow waste as substrate.MSC thesis. Thaper institute of engineering technology
(Deemed University) Punjab, India.
Barnett, H. L., and Hunter, B.B. (1972). Illustrated general of imperfect fungi.3rd
Edn. Burgess,
Minneapolis, Minnesota, USA. pp 20-130.
Barthelomew, D. P., Paul, R. E. and Rohrbach, K. G. (eds). (2003). The Pineapple: Botany,
Production and Uses. CABI Publishing, Wallingford UK. Pp. 1-301.
Blanco, P., Sieiro, C. and Villa, T.G. (1991).Production of pectic enzymes in yeasts.
Microbiological Letters, 175:1–9.
Blanco, P., Sieiro, C. and Villa, T.G. (1999).Production of pectic enzymes in yeasts.
Microbiology Letters, 175:1–9.
Braccini, I., Grasso, R.P. and Pérez, S. (1999). Carbohydrate Resources, 317: 119–130.
Braconnot, H. and Keppler, F. (1825). Methane emission from terrestrial plant under aerobic
conditions.Nature, 439:187-190.
Buga, M.L., Sani, I. and Adrew, J.W. (2010). Partially purified polygalacturonase from
Aspergillus niger (SA6). African Journal of Biotechnology, 9(52): 8944-8954.
Campbell, N, and Reece, I. (2002). Biology, Don O‟Neal (ed.), 6th Edn. Benjamin Cummings
Publishing, Menlo Park, California. Pp. 300-340.
Carpita, N.C. and Gibeaut, D.M. (1993). Structural models of primary cell walls in flowering
plants-consistency of molecular structures with the physical properties of the wall during
growth. Plant Journal, 3:1-30.
Carr, J.G. (1985). Tea, coffee and cocoa. In: Microbiology of fermented foods (ed. Wood,
B.J.B), London: Elsevier Science Ltd, 2:133–154.

Catarina, A., Tomas, B., Pedro, M. and Jose. (2003). Teixeir continuous production of pectinase
by immobilized yeast cell on srent grains. Journal of Bioscience and Bioengineering, 96:
513-551.
Chellegati, M.A.S.C., Fonseca, M.J.V. and Said, S. (2002). Purification and partial
characterization of exo-polygalacturonase from Mucor rouxii NRRL 1894. Enzyme
Microbiology and Technology, 41: 800-805.
Cho, S.W., Lee, S. and Shin, W. (2001). The X-ray structure of Aspergillus aculeatus
polygalacturonases and a modeled structure of the polygalacturonase–octagalacturonate
complex. Journal in Molecular Biology, 314: 863–878.
Codner, R.C. (2001). Pectinolytic and cellulolytic enzymes in the microbial modification of
plant tissues. Journal in Applied Bacteriology, 84: 147–60.
Coppens d'Eeckenbrugge, G; Leal, F. (2003). “Morphology, Anatomy, and Taxonomy". In: D.P
Bartholomew, R.E. Paull, and K.G. Rohrbach. The Pineapple: Botany, Production, and
Uses. Wallingford, UK: CABI Publishing. p. 21.
Corredig, M., Kerr, W. and Wicker, L. (2000). Separation of thermostable pectin methylesterase
from marsh grapefruit pulp. Journal in Agriculture and Food Chemistry, 48: 4918–4923.
Cosgrove, D.J. (1997). Assembly and enlargement of the primary cell wall in plants. Annual
Review on Cell Development and Biology, 13:171–201.
Daas, P.J.H., Boxma, B., Hopman, A.M.C.P., Voragen, A.G.J. and Schols, H.A. (2001).
Biopolymers, 58: 1–8.
Dixit, V.S., Kumar, A.R., Pant, A. and Khan, M.I. (2004). Low molecular mass pectate lyase
from Fusarium moniliforme: similar modes of chemical and thermal denaturation.
Biochemistry, Biophysics Resource Communique, 315: 477–84.
Dixon, M. and Webb, E.C. (1964). Enzymes. 2nd
ed. Longmans, New York. Pp 116-166.

Dorokhov, Y.L., Makinen, K., Frolova, O.Y., Merits, A., Saarinen, J. Kalkkinen, N., Atabekov,
J.G. and Saarma, M. (1999). A novel function for a ubiquitous plant enzyme pectin
methylesterase: the host-cell receptor for the tobacco mosaic virus movement protein.
Federation of European Biochemical Societies, 461: 223–228.
du Penhoat, C.H., Gey, C., Pellerin, P. and Perez, S. (1999). An NMR solution study of the mega
oligosaccharide, rhamnogalacturonan II. Journal of Biomolecules. NMR, 14: 253-271.
Elfick,J. (2007). Pineapple Project.
http://www.uq.ed.au/_School_Science_Lessons/PineProj.html#Lesson2. Retrieved:2-8-2011
Esquivel, J.C.C and Voget, C.E. (2004). Purification and partial characterization of an acidic
polygalacturonase from Aspergillus kawachii. Journal in Biotechnology, 110: 21-28.
Fayyaz, A., Asbi, B.A., Ghazali, H.M., CheMan, T.B. and Jinap, S. (1993). Pectinesterase
extraction from papaya. Food Chemistry, 47: 183–185.
Fleisher, A., O‟Neill, M.A., Ehwald, R. (1999). Plant Physiology, 121: 829-838.
Freitas, P.M., Martin, N., Silva, D., Silva, R. and Gomes, E. (2006). Production and partial
characterization of polygalacturonase production by Thermophilic Monascus sp. N8 and by
Thermotolerant Aspergillus sp. N12 on solid state fermentation. Brazillian Journal in
Microbiology, 37: 32-306.
Gaffe J., Tizando, M.E. and Handa, A.K. (1997). Characterization and functional expression of a
ubiquitously expressed tomato pectin methylesterase. Plant Physiology, 114: 1547–1556.
Gailhofer, G., Wilders-Truschnig, M., Smolle, J., Ludvan, M. (1998). Asthma caused by
bromelain: an occupational allergy. Clinical Allergy 18: 445-450.
Gailing, M.F., Guibert, A. and Combes, D. (2000). Fractional factorial designs applied to
enzymatic sugar beet pulps pressing improvement. Bioprocess Engineering, 22: 69–74.

Gainvors, A., Frezier, V., Lemaresquier, H., Lequart, C., Aigle, M. and Belarbi, A. (1994).
Detection of polygalacturonase, pectin lyase and pectinesterase activities in Saccharomyces
cerevisiae strain. Yeast, 10: 1311–1319.
Gaspar, Y., Johnson, K.L., McKenna, J.A., Bacic, A. and Schultz, C.J. (2001).Plant Molecular
Biology, 47: 161–176.
Glushka, J.N., Terrel, M., York, W.S., O‟Neil, M.A., Gucwa, A., Darvill, A.G., Albersheim, P.
and Prestegard, J.H. (2003). Primary structure of the 2-O-methyl-α-fructose containing side
chain of the pectic polysaccharide, rhamnogalacturonan II. Carbohydrate Residues, 338:
341-352.
Grigelmo-Migeul, N. and Martin-Belloso, O. (1998). Characterization of dietary fiber from
orange juice extraction. Food Resources International, 31: 355-361.
Haq, I., Javed, M.M., Khan, T.S. and Siddiq, Z. (2005). Cotton saccharifying acvtivity of
cellulases produced by co-culture of Aspergillus niger and Trichoderma viride. Research
Journal of Agriculture and Biological Science, 1: 241-245.
Hasunuma, T., Fukusaki, E.I. and Kobayashi, A. (2003). Methanol production is enhanced by
expression of an Aspergillus niger pectin methylesterase in tobacco cells. Journal in
Biotechnology, 106: 45–52.
Helene, M., Canteri-Schenim, Heloísa, C.R.F., Nina, W., Wosiacki, G. (2005).Extraction of
pectin from apple pomace.Brazillian Archeology of Biological Technology. Curitiba, 48: 2-
3.
Helms, D., Helms, C., Kosinski, R. and Cummings, J. (1998). Biology in the Laboratory, 3rd Ed.
Judith Wilson (ed.), Freeman Publishing, New York, New York. Pp. 9: 1-8.
Holker, U., Hofer, M. and Lenz, J.(2004).Biotechnological advantages of laboratory-scale solid
state fermentation with fungi. Applied Microbiology and Biotechnology, 64: 175-186.

Hoondal, G.S., Tiwari, R.P., Tiwari, R., Dahiya, N. and Beg, Q.K. (2000). Microbial alkaline
pectinases and their applications: A review: Applied Microbiology in Biotechnology, 59:
409–418.
Innocenzo, M.D. and Lajalo, F.M. (2001). Effect of gamma irradiation on softening changes and
enzyme activities during ripening of papaya fruit. Journal in Food Biochemistry, 25: 19–27.
IPPA (2001). Types of pectin. Retrieved from http://www.ippa.info/types_of_pectin. htm.
Downloaded on June 5, 2011.
Ishii, S. and Yokotsuka, T. (1975).Purification and properties of pectin lyase from Aspergillus
japonicus.Agriculture, Biology and Chemistry, 39: 313–21.
Ishii, T., Matsunaga, T. (1996). Carbohydrate Residues, 284: 1-9.
Jayani, R.S., Saxena, S. and Gupta, R. (2005). Microbial pectinolytic enzymes: A review.
Process Biochemistry, 40(99): 2931- 2944.
Kapoor, M., Beg, Q.K., Bhushan, B., Singh, K., Dadich, K.S. and Hoondal, G.S. (2001).
Application of alkaline and thermostable polygalacturonase from Bacillus sp. MG-cp-2 in
degumming of ramie (Boehmeria nivea) and sunn hemp (Crotolaria juncia) bast fibers.
Process Biochemistry, 36: 803–807.
Kashyap, D.R., Vohra, P.K., Chopra, S. and Tewari R. (2001). Applications of pectinases in
commercial sector: a review. Bioresource in Technology, 77:215–27.
Kaur, G., Kumar, S. and Satyanarayana, T. (2004). Production, characterization and application
of a thermostable polygalacturonase of a thermophilic mould Sporotrichum thermophile
Apinis. Bioresource in Technology, 94: 239–243.
Kertesz, Z, J. (1951). The Pectic Substance. Interscience Publishers, New York.p 68.
Koboyashi, T., Higaki, N., Yajima, N., Suzumatsu, A., Haghihara, H. and Kawai, S. (2001).
Purification and properties of a galacturonic acid-releasing exo-polygalacturonase from a
strain of Bacillus. Bioscience, Biotechnology and Biochemistry, 65: 842–847.

Kohn, R. (1982). Binding of toxic cations to pectin, it‟s oligomeric fragment and plant tissues,
Carbohydrate Polymers, 2: 273-275.
Kotoujansky, A. (1987). Molecular genetics of pathogenesis by soft-rot Erwinia. Annual Review
in Phytopathology, 25: 405–430.
Koubala, B.B., Kansci, G., Mbome, M.J., Crepeau, J.F., Thibault, J.F. and Ralet, M.C. (2006).
Mango peels as a source of pectin and the effect of extraction method. Food is life- 13th
World Congress of IUFoST (International Union of Food Science and Technology). Nantes
(FRA: 2006/09/17-21).
Lang, C. and Dornenburg, H. (2000).Perspectives in the biological function and the
technological application of polygalacturonases.Applied Microbiology in Biotechnology,
53:366–75.
Lau, J.M., McNeil, M., Darvill, A.G. and Albersheim, P. (1987). Carbohydrate Resources, 168:
245–274.
Laurent, F., Kotoujansky, A. and Bertheau, Y. (2000). Overproduction in Escherichia coli of the
pectin methylesterase A from Erwinia chrysanthemi 3937: one-step purification,
biochemical characterization, and production of polyclonal antibodies. Canadian Journal in
Microbiology, 46: 474–480.
Leal, F., Coppens d'Eeckenbrugge, G. (1996).Fruit Breeding. Leal, F., Coppens d'Eeckenbrugge,
G. (eds.). In. John Wiley and Sons, New York. pp 565-606.
Leda, R., Castilho, A., Ricardo, M. and Tito. L.M. (2000). Alves production and extraction of
pectinase obtained by solid state fermentation of agro industrial residues with Aspergillus
niger. Bioresource Technology, 71: 45-50.
Lohani, S., Trivedi, P.K. and Nath, P. (2004). Changes in activities of cell wall hydrolases during
ethylene-induced ripening in banana: Effect of 1-MCP, ABA and IAA. Postharvest
Biological Technology, 31: 119-126.

Lonsane, B.K., Ghildyal, N.P., Budiatman, S. and Ramakrishna, S.V.(1985). Engineering aspects
of solid state fermentation. Enzyme Microbiology and Technology, 7: 258-265.
Lowry, O.H., Rosenbrough, P.J., Fass, A.L. and Randall, R.J. (1951). Protein measurement with
Folin-phenol reagent. Journal of Biochemistry, 193: 265-275.
Luh, B.S. and Phaff, H.J. (1951). Studies on polygalacturonase of certain yeasts. Archeology,
Biochemistry and Biophysics, 33: 212–227.
Lukong, B.C., Awah, F. M., Nwuke, C.P. and Fobellah, A.D. (2007). Protein Isolation,
Purification and Estimation. In: Introduction to Protein Science. OSPREY Publication
Centre, Owerri, Pp 55-56.
Maldonado, M.C, Saad, A.M.S. and Callieri, D.A.S. (1994). Purification and characterization of
pectinesterase produced by a strain of Aspergillus niger. Curriculum in Microbiology,
24:193–196.
Maldonaldo, M.S. and Saad, A.M.S. (1998). Production of pectinesterase and polygalacturonase
by Aspergillus niger in submerged and solid state systems. Journal of Industrial
Microbiology in Biotechnology, 20: 34–38.
Malviya, R., Srivastava, P., Bansal, M. and Sharma, P.K. (2010). Preparation and evaluation of
disintegrating properties of Cucurbita maxima pulp powder. International Journal in
Pharmaceutical Science, 2(1): 395-399.
Margo, P., Varvaro, L., Chilosi, G., Avanzo, C. and Balestra, G.M. (1994). Pectinolytic
enzymes produced by Pseudomonas syringaepv. Glycinea. Microbiology Letters,117:1–6.
Maria, F.S., Jose, L., Lime, F. and Nelson, D. (2000). Carbon sources effect on pectinase
production from Aspergillus japonicus. Brazillian Journal of Microbiology, 31: 286-290.
Marshal, C. and Chow, K. (2007).Pectin extraction from pectin and sugar beets. Agricultural
Resources, 2: 16-17.

Martin, N., de Souza, S.R., da Silva, R. and Gomes, E. (2004).Pectinase production of fungal
strains in solid-state fermentation using agro-industrial bioproduct. Brazilian Archives of
Biology and Technology, 47(5):813-819.
McCready, R.M. (1970). Methods on Food Analysis. Academic Press, New York and London,
Vol. 2. p 365.
McNeil, M., Darvill, A.G. and Albersheim, P. (1980). Plant Physiology, 66: 1128–1134.
Micheli, F. (2001). Pectin methylesterase: cell wall enzymes with important roles in plant
physiology. Trends in Plant Science, 6: 414–9.
Miller, G.L. (1959). Use of Dinitrosalicylic acid reagent for determination of reducing sugars.
Analytical Chemistry, 31: 426-428.
Miller, J.G. and Fry, S.C. (2001). Characteristics of xyloglucan after attack by hydroxyl radicals.
Carbohydrate Resources, 332: 389-403.
Morris, G., Kӧk, S., Harding, S. and Adams, G. (2010). Polysaccharide drug delivery system
based on pectin and chitosan. Biotechnology & Genetic Engineering Review, 27: 257-284.
Mudgett, A.E. (1986). Solid state fermentations. In: Demain, A.L. and Solomon, N.A.(eds.)
Manual of Industrial Microbiology and Biotechnology,American Society for Microbiology
Washington, D.C., 66-83.
Nikolić, M.V. and Mojovic, L. (2007).Hydrolysis of apple pectin by the coordinated activity of
pectic enzymes. Food Chemistry, 101: 1-9.
Novosel‟skaya, I.L. (2002). Trends in the science and application of pectins.Chemical National
Compilation.36: 1-10.
O‟Neil, M.A., Warrenfiltz, D., Kates, K., Pellerin, P., Doco, T., Darvil, A.G. and Albersheim, P.
(1996). Journal in Biology & Chemistry, 271: 22923-22930.
O‟Neil, M.A., Eberhard, S., Albersheim, P. and Darvill, A.G. (2001). Science 294: 846–849.

O‟Neill, M.A., Ishi, T., Albersheim, P. and Darvill, A. G. (2004). Annual Review of Plant
Physiology and Plant Molecular Biology, 55: 109–139.
Oxenboll, K. (1994). Aspergillus enzymes and industrial uses. In the genus Aspergillus: From
Taxonomy and Genetics to Industrial Application (eds. Powel, K.A., Renwick, A. and
Pederby, J.F). Plenum Press, London. Pp 147-154.
Palomaki, T. and Saarilahti, H.T. (1997). Isolation and characterization of new C-terminal
substitution mutation affecting secretion of polygalacturonases in Erwinia carotovora ssp.
Carotovora, 400: 122–6.
Pandey, A., Soccol, C.R., Rodriguez-Leon, J.A. and Nigam, P.(2001). Solid-State Fermentation
in Biotechnology: Fundamentals and Applications. 1st Edn., Asiatech Publishers Inc., New
Delhi. Pp: 221.
Paoletti, S.(1986). In: Chemistry and function of pectins (eds. Fishman, M.L. and Jen, J.J).
Washington DC: American Chemical Society, pp. 73-87.
Patil, S.R. and Dayanand A. (2006). Production of pectinase from deseeded sunflower head by
Aspergillus niger in submerged and solid-state conditions. Bioresources Technology, 97:
2054-2058.
Perez, S., Rodriguez-Carvajal, M. and Doco, T. (2003). A complex plant cell wall
polysaccharide: rhamnogalacturonan II. A structure in quest of a function. Biochemistry, 85:
109-121.
Pilar, B., Carman, S. and Tmaes, G. (1999). Villa production of pectic enzymes in yeast.
Microbiology Letters.175: 1-9.
Pilnik, N., Voragen, A. G. J. and Hulme A. C. (ed.). (1970). Pectic substances and other
uronides. The biotechnology of Fruits and their products, 1: 53-87.
Pilnik, W. and Voragen, A.G.J. (1993). Pectic enzymes in Fruit and Vegetable Juice
Manufacture. Enzymes in Food Processing,Academic Press Limited, London,Third Edition.
Pp 363-392.

Prade, R.A., Zohan, D., Ayoubi, P. and Mort, A.J. (1999).Pectins, pectinases and plant-microbe
interactions. Biotechnology in Genetic Engineering Review, 16: 361–391.
Prasanna, V., Prabha, T.N. and Tharanatha, R.N. (2007).Fruit ripening phenomena– An
overview. Critical Review in Food Science and Nutrition, 47: 1-19.
Purseglove, J. W. (1972). Tropical crops. Monocotyledons. Longman, London. Pp. 71-91.
Py.C. and Tisseau, M. (1969). La Pina tropical. Ed. Blumé. Barcelona, p. 278.
Raimbault, M. (1998). General and Microbiological Aspects of Solid Substrate
Fermentation.Electronic Journal Of Biotechnology, p 1,3.
Ramanujam, N. and Saritha, P.S. (2008). Production of pectinase by solid-state fermentation of
sugarcane bagasse using Aspergillus niger. Advanced Biotechnology. Pp. 30-33.
Rastogi, G. (1998). Vishal‟s objective botany. Meerut, India: Vishal Publishers. Pp. 23-25.
Reid, I. and Richard, M. (2004). Purified pectinase lowers cationic demand in peroxide-bleached
mechanical pulp. Enzyme in Microbial Technology, 34: 499–504.
Revilla, I. and Ganzalez-san jose, M.L. (2003). Addition of pectolytic enzymes: an enological
practice which improves the chromaticity and stability of red wines. International Journal
on Food Science and Technology, 38: 29–36.
Rexová-Benková, L. and Marković, O. (1976). Pectic enzymes. Advanced Carbohydrate
Chemistry, 33:323–85.
Ridley, B.L., O‟Neil, M.A. and Mohnen, D. (2001). Phytochemistry 57: 929–967.
Rohrbach, K.G., Bathelomew, D. P. and Rohbach, K. G. (2003). Pests, Disease and Weeds.
CABI, New York Pp 203-252.
Rosenberg, I.M. (2004). Protein Analysis and Purification: Benchtop Technique, 2nd
Edn.
Springer, Birkhauser, p 520.

Rugkong, A., Rose, J.K.C., Lee, S.G., Giovannoni, J.J. O‟Neill, M.A. and Watkins, C.B. (2010).
Cell wall metabolism in cold-stored tomato fruit. Postharvest Biological Technology, 57:
106-113.
Sakai, T. and Ozaki, Y. (1988). Protopectin solubilizing enzyme that does not catalyze the
degradation of polygalacturonic acid. Agriculture, Biology and Chemistry, 52: 1091–1093.
Sakai, T. (1992).Degradation of pectins. In: Microbial degradation of natural products. (ed.
Winkelmann, G.). Weinheim. Pp. 57–81.
Sakai, T., Sakamoto, T., Hallaert, J. and Vandamme, E.J. (1993). Pectin, pectinase and
protopectinase: Production, properties and applications. Advanced Application of
Microbiology, 39: 231–294.
Sakamoto, T., Hours, R.A. and Sakai, T. (1994).Purification, characterization and production of
two pectic-transeliminases with protopectinase activity from Bacillus subtilis.Bioscience in
Biotechnology and Biochemistry, 58: 353–358.
Salazar, L. and Jayasinghe, U. (1999).Fundamentals of purification of plant viruses. In:
Techniques in plant, virology, CIP. International Potato Centre, Peru. Pp. 1–10.
Sañudo-Barajas, J.A., Labavitch, J., Greve, C., Osuna-Enciso, T., Muy-Rangel, D. and Siller-
Cepeda, J. (2009). Cell wall disassembly during papaya softening: Role of ethylene in
changes in composition, pectin-derived oligomers (PDOs) production and wall hydrolases.
Postharvest Biological Technology, 51: 158-167.
Sathyanarayana, N.G. and Panda, T. (2003).Purification and biochemical properties of microbial
pectinases. A review: Process Biochemistry, 38: 987–996.
Serra, J.L., Alkorta, I., Llama, M.J. and Alaiia, A.(1992).Aplicacion industrial de 1os enzimas
pecticos.Production, purification, inmovilizacion y algunas propiedades de la pectina liasa
de Penicillium italicum.Alimentacion Equiposy Technology, 11: 127-134.
Schols, H.A., Bakx, E.J., Schipper, D. and Voragen, A.G.J. (1995).Carbohydrate Resources,
279: 265–279.

Schols, H. A. Visser, R. G. F. and Voragen, A. G. J. (2009).Pectins and pectinases. Wegenigen
Academic Publisher, The Netherlands. Pp. 980-990.
Shembekar, V.S. and Dhotre, A. (2009).Studies of pectin degrading microorganisms from
soil.Journal of Microbial World 11(2): 216-222.
Singh, S.A. and Rao, A.G.A. (2002). A simple fractionation protocol for, and a comprehensive
study of the molecular properties of two major endopolygalacturonases from Aspergillus
niger. Biotechnology in Applied Biochemistry, 35: 115–123.
Smith, J.E. and Aidoo, K.E. (1988). Growth of fungi on Solid Substrates. Physiology of
Industrial Fungi, Blackwell, Oxford, England. Pp 249-269.
Solis-Pereyra, S., Favela-Torres, E., Gutierrez–Rojas, M., Roussos, S., Saucedo, C.G. and
Viniegra, G.G.1996. Production ofpectinases by Aspergillus nigerin solid state fermentation
at highglucose concentrations. World Journal of Microbiology and Biotechnology, 12: 275-
260.
Sriamornsak, P. (1998). Chemistry of Pectin and it‟s pharmaceutical uses: A review, Journal of
Pharmacology in Phamaceutical Science, 44: 207-228.
Sriamornsak, P. and Nunthanid, J. (1998). Calcium pectinate gel beads for controlled release
drug delivery: In Preparation and in vitro release studies, International Journal in
Pharmacy, 160: 207-212.
Sriamornsak, P. (2002). Analysis of selected physicochemical properties of pectins and alginate
gels intended for drug delivery. PhD thesis, Charles Sturt University.
Srivastava, P., Malviya, R, Gupta, S. and Sharma, P.K. (2010). Preparation and evaluation of
matrix based tablet using natural polymers as release modifiers, International Journal in
Pharmaceutical Science, 2(1): 411-417.
Sunnotel, O. and Nigam, P. (2002). Pectinolytic activity of bacteria isolated from soil and two
fungal strains during submerged fermentation. World Journal in Microbiology and
Biotechnology, 18: 835–839.

Szajer, I. and Szajer, C. (1984). Pectin lyase of Penicillium paxilli. Biotechnology Letters, 4:
549–52.
Thakur, B. R., Singh, R. K. and Handa A. K. (1997). Chemistry and uses of pectin: a review.
Critical Review in Food science and nutrition, 37: 47-73.
Thakur, A., Pahwa, R., Singh, S. and Gupta, R. (2010). Production, Purification, and
characterization of polygalacturonase from Mucor circinelloides ITCC 6025. Enzyme
Research,(170549), 1-7.
Tucker, G.A. and Woods, L.F.J. (1991). Enzymes in Production of Beverages and Fruit juices.
Enzyme in Food Processing, Blackie, New York, Pp. 201-203.
van der Vlugt-Bergmans, C.J.B., Meeuwsen, P.J.A., Voragen, A.G.J. and van Oogen, A.J.J.
(2000). Endo-xylogalacturonan hydrolase, a novel pectinolytic enzyme. Applied
Environmental Microbiology, 66: 36–41.
Vicken, J.P., Schols, H.A., Oomen, R.J.F.J., McCann, M., Ulvskov, P., Voragen, A.G.J., Visser,
R.G.F. (2003). Plant Physiology, 132: 1781-1789.
Viikari, L., Tenakanen, M. and Suurnakki, A. (2001). Biotechnology in the pulp and paper
industry. In: Biotechnology, Rehm, H.J. (Ed). VCH-Wiley, Pp. 523–546.
Visser, J., Voragen, A.G.J. (1996) Progress in Biotechnology: Pectins and Pectinases. Elsevier,
Amsterdam. Pp. 14-15.
Voragen, A.G.J., Schols, H.A. and Visser, R.G.F. (2003).Advances in Pectin and Pectinase
Research. Kluwer Academic Publishers, Dordrecht, The Netherlands. Pp. 203-205.
Wang, G., Michailides, T.J. and Bostock, R. M. (1997). Improved detection of polygalacturonase
activity due to Mucor piriformis with a modified dinitrosalicylic acid
reagent.Phytopathology, 87: 161-163.

Wei J., Ma,F., Shi, S., Qi, X., Zhu, X. and Yuan, J. (2010). Changes and postharvest regulation
of activity and gene expression of enzymes related to cell wall degradation in ripening apple
fruit. Postharvest Biological Technology, 56: 147-154.
Whitaker, J.R. (1984). Pectic substances, pectic enzymes and haze formation in fruit juices.
Enzyme Microbial Technology, 6: 341–347.
Whitaker, J.R. (1990). Microbial pectinolytic enzymes. In: Microbial enzymes and
biotechnology (eds. Fogarty, W.M. and Kelly, C.T.), 2nd ed. London: Elsevier Science Ltd.
Pp. 133–76.
Wijesundera, P.L.C., Bailey, J.A. and Byrde, R.J.W. (1984). Production of pectic lyase by
Colletotricum lindemuthionum in culture and in infected bean (Phaseolus vulgaris) tissue.
Journal in General Microbiology., 130: 285-290.
Wikpedia.(2011). The Pineapple fruit. Retrieved from http://en.wikipedia.org/wiki/pineapple on
19/06/2011.
Willats, W.G.T., McCartney, L., MacKie, W. and Knox, J.P. (2001a). Plant Molecular Biology
47: 9–27.
Young, M.M.; Moriera, A.R.and Tengerdy, R.P. (1983).Principles of Solid State Fermentation
in Smith J.E.; Berry, D. R. and Kristiansen, B. (eds). Filamentous fungi Fungal
Technology, Arnold, E. London. Pp. 117-144.
Yujaroen, P., Supjaroenkul, U., Rungrodnimitchai, S. (2008). Extraction of pectin from sugar
palm meal. Thamm International Journal in Science and Technology (special edition), 13:
44-47.
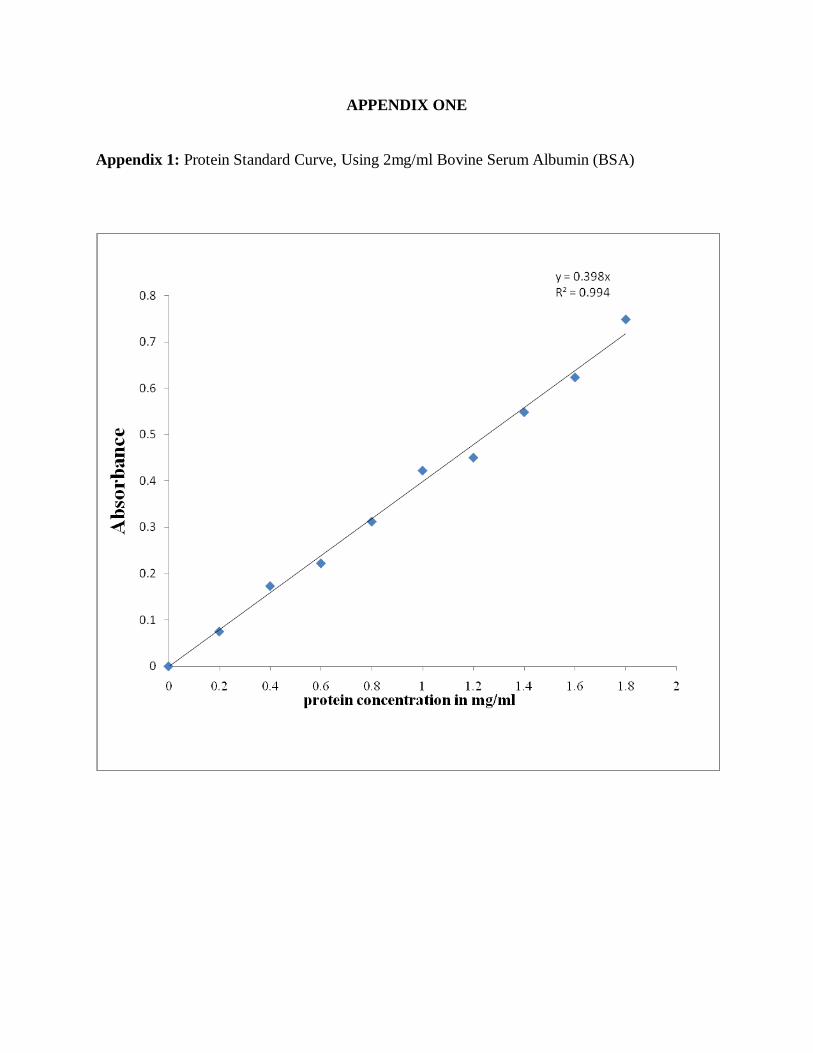
APPENDIX ONE
Appendix 1: Protein Standard Curve, Using 2mg/ml Bovine Serum Albumin (BSA)

APPENDIX TWO
Appendix 2: Galaturonic Acid Standard Curve, Using 20mM D-(+)-Galacturonic Acid
Monohydrate