printed liz usa. the preparation and properties of bovine ... · the preparation and properties of...
TRANSCRIPT

THE JOURNAL OF BIOL.OGICAL. CHEMISTRY Vol. 254, No. 5, Issue of March 10, pp. 1657-1683, 1979 Printed LIZ USA.
The Preparation and Properties of Bovine Enterokinase”
(Received for publication, September 5, 1978)
Juris J. Liepnieks and Albert Light
From the Department of Chemistry, Purdue University, West Lafayette, Indiana 47907
Bovine enterokinase was purified from duodenal mu- cosa. The purification included an initial extraction with 2% deoxycholate, ammonium sulfate fractiona- tions, DEAE-cellulose chromatography, and affinity chromatography on basic pancreatic trypsin inhibitor (Kunitz) (PTI)-Sepharose. The purified enzyme con- tained 35% carbohydrate; it had a molecular weight of 150,000, with a heavy (115,000) and light (35,000) chain connected by one or more disulfide bonds. Enteroki- nase hydrolyzed lysine and arginine substrates and slowly reacted with the trypsin active site titrant 4- methylumbelliferyl-p-guanidinobenzoate. The enzyme activated bovine trypsinogen with kinetic parameters similar to those of other preparations of enterokinase. Bovine enterokinase was inhibited by Kunitz pan- creatic trypsin inhibitor with a K,,,,, of 2 x 10’ M-’ and only weakly by other proteinase inhibitors. The amino acid composition differed from bovine enterokinase iso- lated from duodenal contents (Anderson, L. E., Walsh, K. A., and Neurath, H. (1977) Biochemistry 16, 3354- 3360). The mucosal enzyme and the duodenal contents enzymes also differed in the size of the heavy and light chains. The mucosal enterokinase more closely resem- bled the properties of porcine enterokinase (Baratti, J., Maroux, S., Louvard, D., and Desnuelle, P. (1973) Biochim Biophys. Acta 315, 147-161). The amino acid composition and size of the light chain were also similar to bovine trypsin.
Enterokinase (enteropeptidase, EC 3.4.21.9) is one of many hydrolases located in the brush border membrane of the intestinal duodenum (1,2). The enzyme initiates the intestinal digestion of foodstuffs by activating pancreatic trypsinogen, which then activates pancreatic chymotrypsinogen, proelas- tase, procarboxypeptidase, and prolipase (3-5). In the activa- tion process, enterokinase catalyzes the hydrolysis of the Lys 6--Ile 7 bond of bovine trypsinogen with the release of the NHz-terminal hexapeptide (4, 6). The newly formed trypsin catalyzes the hydrolysis of a single bond in the other zymo- gens, converting each to an active enzyme. The activation process has a large amplification since a small amount of enterokinase produces large amounts of active trypsin and other digestive enzymes (7).
The molecular and enzymatic properties of porcine entero-
* This work was supported by Grant GM-22261 from the National Institutes of Health. A portion of this work was taken from a thesis by J. J. L. submitted to Purdue University in partial fulfillment of the requirement for the degree of Doctor of Philosophy. A preliminary report of this work appeared in (1976) Fed. Proc. 35, 1460 and was presented at the 1978 Joint Central Great Lakes Regional Meeting of the American Chemical Society, Indianapolis, Indiana, May 1978. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
kinase have been reported by the Desnuelle group in Marseille (4, 8, 9). The enzyme was purified from intestinal mucosa in a multistep procedure. The purified porcine enzyme has a molecular weight of 195,000 and contains about 40% carbo- hydrate. The protein has a heavy and light chain linked by one or more disulfide bonds. The enzyme is a serine proteinase and components of the active site are only found in the light chain. Enterokinase and bovine trypsin have similar specific- ities since both enzymes are capable of activating trypsinogen (4, 8, 9). However, trypsin is a poor activator of trypsinogen, while enterokinase is very much better (k,.,, is 2000 times larger, K, about 6 times smaller).
Recently, the purification, specificity, and some molecular properties of enterokinase isolated from the intestinal contents of the cow have been reported by Anderson et al. (10). The purified bovine enzyme has a molecular weight of 145,000, which is much less than that of the porcine enzyme, and both the heavy and light chains also differ from the molecular weights of the porcine enzyme.
It is surprising that the molecular weights of bovine and porcine enterokinase differ to such an extent since these species are closely related on the evolutionary scale (11). We purified enterokinase from bovine duodenal mucosa, and in light of the above differences in size, we examined the prop- erties of the purified enzyme and paid particular attention to these comparative aspects. We wish to describe our purifica- tion procedure, which utilizes affinity chromatography, and to report on the size of the enzyme, its amino acid composition, carbohydrate content, and several enzymatic properties to- ward large and small molecules. We will compare these prop- erties with those reported by the Seattle and Marseille labo- ratories.
EXPERIMENTAL PROCEDURES
Materials
Trypsin, trypsinogen, chymotrypsinogen A, ovalbumin, pepsin, and soybean trypsin inhibitor were obtained from Worthington. Bovine serum albumin was obtained from Pentex. Kunitz pancreatic trypsin inhibitor (Trasylol) was a generous gift from Dr. E. Truescheit of the Bayer Werk Co., while other trypsin inhibitors were a gift from Prof. M.-Laskowski, Jr. of Purdue University.
DEAE-cellulose (DE52) was ourchased from Whatman: Seoharose 4B and Con A-Sepharose from Pharmacia; and Ultrogel AcA i2 from LKB. Tos-Arg-OMe,’ Bz-Arg-OEt, sodium deoxycholate, sodium do- decyl sulfate, and Triton X-100 were products of Sigma. Tos-Lys- OMe, Gdn-Bz-ONp, and TLCK were from Cycle. Leucyl ,%naph- thylamide was purchased from Mann. Dithioerythritol and guanidine hydrochloride (Heico) were obtained from Pierce and urea (ultrapure)
I The abbreviations used are: Tos-Arg-OMe, N”-tosyl-L-arginine methyl ester; Bz-Arg-OEt, N”-benzoyl-L-arginine ethyl ester; Tos- Lys-dMe, N”-tosyl-L-lysine methyl ester; TLCK, N”-tosyl-L-lysine chloromethvl ketone; Gdn-Bz-ONp, p-nitrophenvl-p’-guanidinoben- _ _ _ __ zoate; Gdn-Bz-OMum, 4-methylumbelliferyl-p-guanidinobenzoate; STI, soybean trypsin inhibitor (Kunitz); PTI, basic pancreatic trypsin inhibitor (Kunitz); SDS, sodium dodecyl sulfate; Con A, concanavalin A.
1677
by guest on June 9, 2018http://w
ww
.jbc.org/D
ownloaded from

1678 Bovine Enterokinase
from Schwarz/Mann. Dimethylsuberimidate and cyanogen bromide were obtained from Aldrich. Gdn-Bz-OMum was synthesized by the procedure of Chase and Shaw (12) for their synthesis of Gdn-Bz-ONp. 4-Methylumbelliferone was purchased from Eastman. Naphthanil diazo blue was obtained from K & K.
Methods
Assays-Trypsin activity was measured potentiometrically at pH 7.9 and 25’C with 0.01 M solutions of Tos-Arg-OMe, Bz-Arg-OEt, or Tos-Lys-OMe containing 0.05 M calcium chloride (13). A trypsin unit is the amount of enzyme hydrolyzing 1 amol of Tos-Arg-OMe/min. &at values were calculated from the rate of hydrolysis of the substrate and the molarity of the enterokinase solution. The operational mo- larity of enterokinase was calculated from a titration of the enzyme with the active site-directed reagent Gdn-Bz-OMum (14). Typically, 10 to 15 +l of a solution of enterokinase in 3 ml of 0.1 M Veronal, pH 8.3, containing 0.02 M CaC12, were rapidly mixed with 3 ~1 of Gdn-Bz- OMum (0.4 mg/ml of dimethyl formamide). A Perkin-Elmer MPF- 2A spectrofluorometer was used to follow the reaction, with excitation at 360 nm and emission at 450 nm. The time for the “burst” lasted 2 min before a steady state turnover of substrate was reached. The fluorescence burst was standardized with a trypsin solution whose molarity of active sites had been determined by titration with Gdn- Bz-ONp (12).
used for the molecular weight determination of the light chain of enterokinase. The reference proteins and their molecular weights are: the monomer and dimer of bovine serum albumin (67,000 and 134,000), ovalbumin (43,000), pepsin (35,000), chymotrypsinogen A (25,000), cross-linked bovine serum albumin (67,000, 134,000, and 201,000), and cross-linked ovalbumin (43,009, 86,009, 129,OCQ and 172,000). Cross-linked serum albumin and ovalbumin were prepared as described by Carpenter and Harrington (25). The chains of enter- okinase were cross-linked by the method of Davies and Stark (26).
Preparation of Enterokinase-Fresh bovine duodena were ob- tained from a local slaughterhouse and stored at -20°C. Frozen bovine intestinal duodena were thawed overnight at room tempera- ture, excess fat was removed, and the duodena were sliced down the middle. A spatula was used to remove the mucosa lining the intestinal wall by gentle scraping. The mucosal cells were stored at -20°C until needed.
Enterokinase activity was determined from the activation of tryp- sinogen (3). Activation mixtures contained enterokinase, 0.1 M sodium acetate, pH 5.0, 0.05 M calcium chloride, and 0.1 ml of trypsinogen solution (1.0 mg/ml of 1 mrvr HCl) in a total volume of 1 ml. After 30 min at 35”C, the reaction was quenched with 0.1 ml of 1 M HCl, and the tryptic activity was measured with 0.01 M Tos-Arg-OMe. An enterokinase unit is the amount of enzyme producing 1 trypsin unit/ ml of activation mixture. The assay was linear from 0.5 to 7 entero- kinase units.
In a typical preparation, 4 kg of thawed duodenal mucosa were extracted with 12 liters of 2% deoxycholate containing 0.05 M Tris, pH 8.0, for 2 h with continuous stirring. All operations were performed at 4°C. The mixture was centrifuged at 11,000 X g for 1 h in a Beckman J-21 centrifuge with a JA-IO rotor. The supernatant was acidified to pH 6.0 with 1 N acetic acid which precipitated the bulk of the deoxycholic acid and some impurities. The precipitate was re- moved by centrifugation at 11,000 X g for 30 min. Fat particles were removed by filtration through glass wool. The supernatant was ad- justed to pH 8 and brought to 40% saturation with solid ammonium sulfate, stirred 1 h, and centrifuged at 14,000 x g for 1 h. The supernatant was then brought to 70% saturation with ammonium sulfate and centrifuged at 13,000 X g for 30 min. The precipitate was dissolved in 3 liters of water, brought to 30% saturation with ammo- nium sulfate, and centrifuged. The supernatant was made 60% satu- rated with ammonium sulfate and centrifuged. The precipitate was dissolved in 1 liter of water and dialyzed against 0.005 M Tris, pH 6.0, containing 0.02 M NaCl.
The inhibition of enterokinase in the presence of proteinase inhib- itors was determined by incubating 6.4 nM enzyme and 1.3 PM inhibitor in 0.1 M Veronal, 0.02 M CaClz, pH 8.3, for 30 min at room temperature. The amount of enzyme remaining was determined from a titration with Gdn-Bz-OMum. The K,,,,,, for the binding of enterokinase and bovine Kunitz PTI was determined by incubating 6.5 nM enzyme wit,h varying amounts of PTI (from 0 to 19 nM) in the pH 8.3 buffer for 30 min at room temperature. The residual enzyme activity was deter- mined with Gdn-Bz-OMum.
The dialyzed sample was mixed with 700 ml of settled DEAE- cellulose (DE52) equilibrated with 0.01 M Tris, pH 6.0, containing 0.04 M NaCl, and stirred for 2 h. The cellulose was recovered by centrifu- gation at 1500 x g. The cellulose was suspended in the equilibration buffer, poured into a column, and packed by gravity flow. After washing with 4 to 5 column volumes of the pH 6.0 buffer, the enterokinase was eluted with 0.01 M Tris, pH 6.0, containing 0.12 M NaCl (Fig. 1).
Aminopeptidase was detected with a spot test procedure (15). A lo-al sample was mixed with 0.5 ml of 0.3 mM leucyl /j’-naphthylamide in 0.05 M Tris, pH 8.5, containing 0.005 M CaCIZ, and incubated for 30 min at room temperature. After 0.2 ml of naphthanil diazo blue (5 mg/ml of water) was added, the immediate appearance of a pink color was taken as a positive test.
The active fraction was adjusted to pH 8.0 and applied to a PTI- Sepharose column (1.6 X 23 cm) equilibrated with 0.05 M KHcPO,, pH 8.0, containing 0.5 M NaCl. The column was washed with at least 20 column volumes of the equilibration buffer and then with 0.1 M sodium formate, pH 3.0, containing 0.1 M NaCl (Fig. 2). Fifteen- milliliter fractions of the pH 3 eluent were collected in vials containing
Preparation of PTI-Sepharose-Sepharose-bound Kunitz pan- creatic trypsin inhibitor was prepared by the method of March et al. (16). Approximately 25 ml of washed Sepharose 4B were activated with 1 g of cyanogen bromide at 0°C for 2 min. The act,ivated gel was mixed with 60 mg of PTI in 30 ml of 0.2 M NaHCO:,, pH 9.5, for 18 h at 0°C. Ethanolamine was added to 0.1 M and the reaction mixture was kept at room temperature for 4 h for the reaction with the remaining activated sites.
Analyses-Amino acid analyses were performed on a Durrum model D-500 amino acid analyzer. Samples of enterokinase or the S- carboxymethylated heavy and light chains were hydrolyzed in dupli- cate in 6 N HCl at 108°C for 22,48, 72, and 120 h in evacuated, sealed tubes. Norleucine was added to the hydrolysates as an internal standard (17). Half-cystine was determined as cysteic acid after performic acid oxidation (18) or as S-carboxymethyl cysteine. Tryp- tophan was determined spectrophotometrically (19) and after alkaline hydrolysis (20). Glucosamine and galactosamine were estimated from amino acid analyses after extrapolation to zero time to correct for partial destruction. Neutral sugars were determined by the orcinol/ HLSO? procedure (21); galactose was used as a reference standard. Reduced and carboxymethylated proteins were prepared by the method of Hirs (22).
Fraction Number
SDS-Gel Electrophoresis-Polyacrylamide gel electrophoresis, with gels varying in per cent acryiamide, was carried out essentially as described by Weber and Osborn (23). Electrophoresis was per- formed at 6 mA/tube and continued until the tracking dye was approximately 1 cm from the bottom. Gels were stained with Coo- massie brilliant blue G solution and destained in a Bio-Rad diffusion destainer model 172A. The method of Segrest and Jackson (24) was activity (umts/ml X 10 ')
FIG. 1. Purification of enterokinase. Chromatography of the di- alyzed ammonium sulfate precipitate on DEAE-cellulose (5 x 48 cm). The buffer was 10 mM Tris/acetate, pH 5.0, containing 40 mM NaCl, and was changed at 1 to 0.12 M and at 2 to 0.25 M NaCl. The flow rate was 250 ml/h and 20-ml fractions were collected. See text for details on the initial batch adsorption of the sample on DEAE-cellulose. The .solid line is absorbance at 280 nm and the dashed line is enterokinase ..,. . -1
1
by guest on June 9, 2018http://w
ww
.jbc.org/D
ownloaded from

Bovine Enterokinase 1679
Volume (mIxlO
FIG. 2. Affinity chromatography on PTI-Sepharose. The fraction from DEAE-chromatography (Fig. 1) was added to a column (1.6 x 23 cm) equilibrated with 0.05 M sodium phosphate, pH 8.0, containing 0.5 M NaCl. The column was washed with 20 column volumes of buffer, then with 0.1 M formate, pH 3.0, containing 0.1 M NaCl. The low pH fractions (15 ml) were collected in vials containing 3.7 ml of 0.5 M Tris, pH 9.0. The initial flow rate was 40 ml/h and increased to 120 ml/h with the second buffer.
3.7 ml of 0.5 M Tris, pH 9.0, with stirring, or 150 ml of the eluent were collected directly in a beaker containing 37 ml of 0.5 M Tris, pH 9.0. The enterokinase fraction was dialyzed against water, concentrated to 10 ml or less, filtered through a 2-p Polyvic Millipore filter, and lyophilized.
In the purification of the enzyme, protein concentrations were estimated from spectral measurements at 280 nm with an assumed E”’ of 10.0. We determined the E’” to be 8.4 for purified enterokinase. A stock solution of dialyzed enzyme was used to measure the absorb- ance at 280 nm and a fraction was dried to constant weight and corrected for ash.
Polyacrylamide gels of 7% were prepared according to Davis (27), omitting the stacking gel. Glycerol was added to the samples to a final concentiation of 20%.-The reservoir buffers were 5 miTris, 38 mM glycine, at pH 8.5. Electrophoresis was performed at 2 mA/tube.
Electrophoresis on 4 to 30% pore gradients (Pharmacia) used a buffer of 10.75 g of Tris, 5.04 g of boric acid, and 0.93 g of EDTA diluted to 1 liter at pH 8.35. After a prerun for 1 h at 70 V, the sample in diluted buffer (1:2), containing 40% sucrose, was layered on the slabs and run for 20 min at 70 V and 16 to 18 h at 150 V. The samples were fmed with 25% isopropyl alcohol, 10% acetic acid for 2 h, stained with 0.005% Coomassie blue in 7% acetic acid at 36 V for 15 min/side, and destained in 25% isopropyl alcohol, 10% acetic acid at 36 volts for 15 min.
Separation of Chains-The S-carboxymethylated heavy and light chains of enterokinase were separated on a column of Sepharose-4B (1.5 x 88 cm) with 6 M guanidine acetate, pH 6.0, at room temperature, at a flow rate of 2 ml/h. Column fractions were analyzed at 280 nm.
RESULTS
Purification of Enterokinase-Enterokinase is known to be bound to the brush border membrane of the duodenal mucosa (2, 28, 29), and we chose this source from the cow for the purification of the enzyme. Because the enzyme is membrane- bound, extraction of the mucosa with 2% deoxycholate was more effective than with aqueous systems or with Triton X- 100 (1 to 4%). For example, extraction of the mucosa with a pH 8.0 buffer yielded only 37% of the active fraction.
The results of a typical preljaration are shown in Table I. Most of the deoxycholate was conveniently removed from the extract on acid precipitation. The ammonium sulfate fraction- ations served to clarify the extract of dispersed lipids, which eliminated channeling in the chromatography that followed. A batch adsorption on DEAE-cellulose concentrated the ac- tive fraction and bound the protein for gradient elution chro- matography (Fig. 1). Although the active fraction was re- covered in a narrow cut, all of the aminopeptidase activity of the mucosa co-eluted with the enterokinase. Similar proper-
TABLE I Purification of bovine enterokinase from 4 kg of duodenal mucosa
Purification step Protein” Total activ- itvh
Specific Yield Purifica- activitv tion
mg units units/mg % -fold
2% Deoxycholate 1.3 x lo6 1.2 x lo7 9 mm (1.0) extract
pH 6 supernatant 4.0 X IO5 1.0 x 10’ 25 85 2.5 60% saturation 5.9 X lo4 9.6 x lo6 162 82 16
(NH&304 pre- cipitate
DEAE-cellulose 4.7 x lo3 5.3 x lo6 1,130 45 113 chromatogra- phy
PTI-Sepharose 11.5 3.22 x 10“ 280,000 28 28,000
” Protein was determined spectrophotometrically, assuming an E&?i, of 10.
b Trypsinogen (0.1 mg/ml) was activated at pH 5,35”C, for 30 min. Trypsin activity was determined with Tos-Arg-OMe.
A C FIG. 3. Polyacrylamide gel electrophoresis of purified enterokinase
on (A) 7% acrylamide gel, (B) 5% acrylamide gel with 0.1% SDS, (C) 5% acrylamide gel with 0.1% SDS and 1% P-mercaptoethanol.
ties were reported for the porcine preparation (8). In the next step, affinity chromatography on PTI-Sepharose gave a 250- fold purification and yielded a fraction that appeared homo- geneous (Fig. 3). If we use the specific activity value of the purified enzyme and the total enzyme units extracted per gram of mucosal cells, we estimate the content of enterokinase in the brush border membrane as 0.001%. Routinely, we obtained 8 to 12 mg of purified enterokinase.
The stability of enterokinase at pH 3.0 was investigated because a buffer at pH 3 was used to displace the enzyme from PTI-Sepharose. Under the conditions used for affinity chromatography, less than 10% of the molecules were de- stroyed.
Homogeneity of the fraction was based on a constant spe- cific activity across the peak on affinity chromatography (Fig. 2), the absence of aminopeptidase activity, the constant value for the specific activity on repeating the affinity chromatog- raphy or the ion exchange chromatography, a single broad band on electrophoresis on a 7% gel, and a single band on SDS electrophoresis on a 5% gel (Fig. 3B).
by guest on June 9, 2018http://w
ww
.jbc.org/D
ownloaded from
1680 Bovine Enterokinase
A fraction of enterokinase (15 to 40%) did not bind to PTI (Fig. 2) and remained unbound on repeated chromatography on PTI-Sepharose under the above conditions. The fraction was further purified on DEAE-cellulose (5 x 31.5 cm) with a shallow salt gradient at pH 8.0, on Ultrogel AcA 22 (0.9 x 105 cm), and on Con A-Sepharose (1.6 X 8.5 cm). The active fraction was recovered from Ultrogel just before the major protein fraction, and it was unretarded on Con A while a large fraction of the protein remained bound. The partly purified sample now had properties typical of the purified enteroki- nase. The sample could be bound to PTI-Sepharose and displaced with the pH 3 buffer. On a molar basis it had a trypsinogen-activating activity of 46,000 enterokinase units/ nmol, identical with enterokinase. The ratio of the molar specific activity toward lysine and arginine substrates was the same as the homogeneous enzyme. On SDS electrophoresis, with and without mercaptoethanol present, bands were de- tected with the same mobility shown for the homogeneous enzyme (see below). The bound and unbound fractions appear to be identical; only a single species of enterokinase is present in mucosal cells. Difficulties due to heterogeneity of the car- bohydrate structure of enterokinase or from other causes were not observed in the purification of the enzyme.
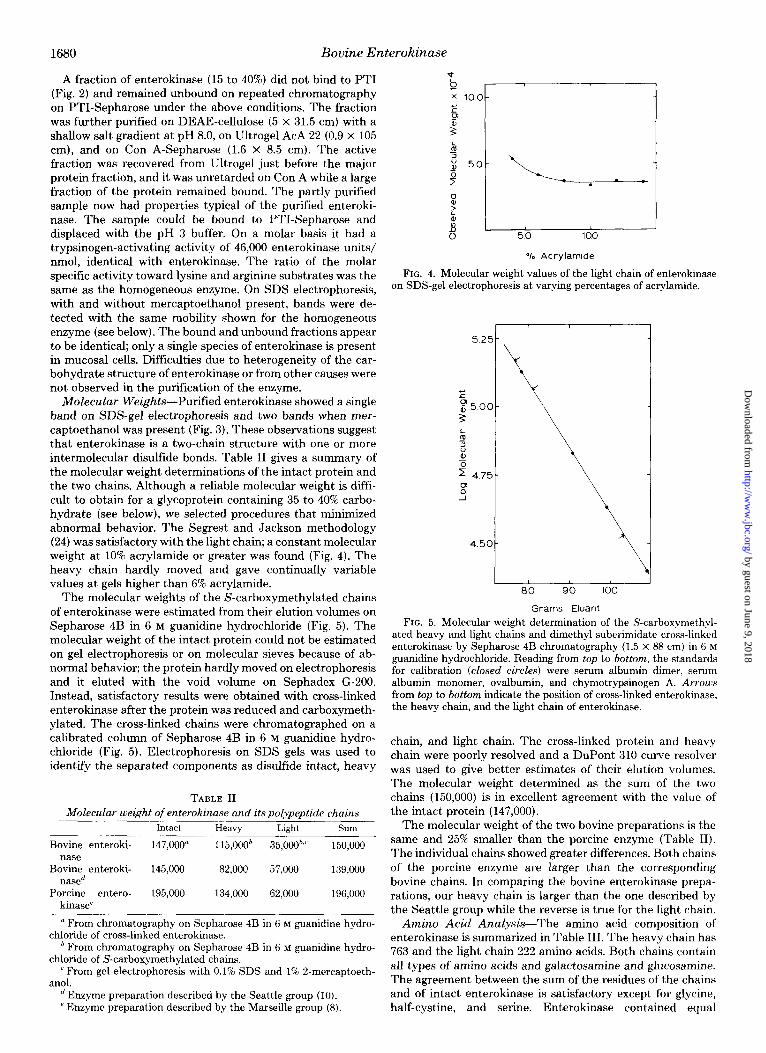
Molecular Weights-Purified enterokinase showed a single band on SDS-gel electrophoresis and two bands when mer- captoethanol was present (Fig. 3). These observations suggest that enterokinase is a two-chain structure with one or more intermolecular disulfide bonds. Table II gives a summary of the molecular weight determinations of the intact protein and the two chains. Although a reliable molecular weight is diffi- cult to obtain for a glycoprotein containing 35 to 40% carbo- hydrate (see below), we selected procedures that minimized abnormal behavior. The Segrest and Jackson methodology (24) was satisfactory with the light chain; a constant molecular weight at 10% acrylamide or greater was found (Fig. 4). The heavy chain hardly moved and gave continually variable values at gels higher than 6% acrylamide.
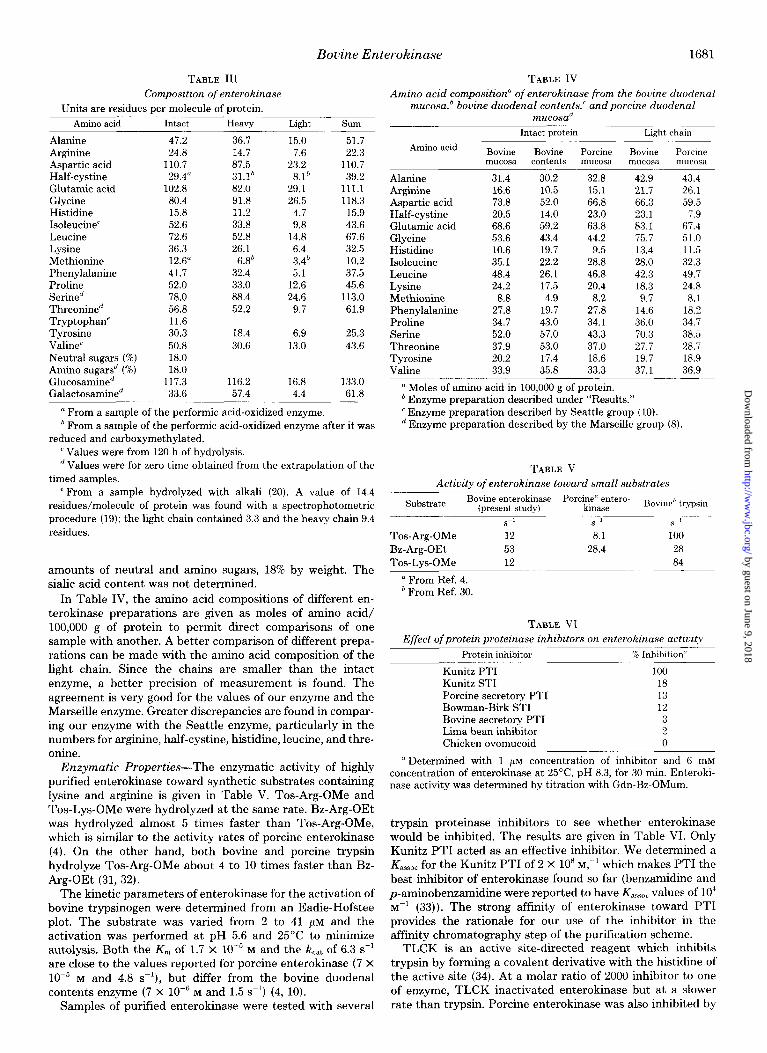
The molecular weights of the S-carboxymethylated chains of enterokinase were estimated from their elution volumes on Sepharose 4B in 6 M guanidine hydrochloride (Fig. 5). The molecular weight of the intact protein could not be estimated on gel electrophoresis or on molecular sieves because of ab- normal behavior; the protein hardly moved on electrophoresis and it eluted with the void volume on Sephadex G-200. Instead, satisfactory results were obtained with cross-linked enterokinase after the protein was reduced and carboxymeth- ylated. The cross-linked chains were chromatographed on a calibrated column of Sepharose 4B in 6 M guanidine hydro- chloride (Fig. 5). Electrophoresis on SDS gels was used to identify the separated components as disulfide intact, heavy
TABLE II Molecular weight of enterokinase and itspolypeptide chains
Intact Heavy Light SUlll
Bovine enteroki- 147,000” 115,oOOh 35,000bJ 150,000 nase
Bovine enteroki- 145,000 82,000 57,000 139,000 nased
Porcine entero- 195,000 134,000 62,000 196,000 kinase’
(1 From chromatography on Sepharose 4B in 6 M guanidine hydro- chloride of cross-linked enterokinase.
’ From chromatography on Sepharose 4B in 6 M guanidine hydro- chloride of S-carboxymethylated chains.
’ From gel electrophoresis with 0.1% SDS and 1% 2-mercaptoeth- anol.
’ Enzyme preparation described by the Seattle group (IO). ’ Enzyme preparation described by the Marseille group (8).
% Acrylamlde
FIG. 4. Molecular weight values of the light chain of enterokinase on SDS-gel electrophoresis at varying percentages of acrylamide.
80 90 100
Grams Eluant
FIG. 5. Molecular weight determination of the S-carboxymethyl- ated heavy and light chains and dimethyl suberimidate cross-linked enterokinase by Sepharose 4B chromatography (1.5 x 88 cm) in 6 M guanidine hydrochloride. Reading from top to bottom, the standards for calibration (closed circles) were serum albumin dimer, serum albumin monomer, ovalbumin, and chymotrypsinogen A. Arrows from top to bottom indicate the position of cross-linked enterokinase, the heavy chain, and the light chain of enterokinase.
chain, and light chain. The cross-linked protein and heavy chain were poorly resolved and a DuPont 310 curve resolver was used to give better estimates of their elution volumes. The molecular weight determined as the sum of the two chains (150,000) is in excellent agreement with the value of the intact protein (147,000).
The molecular weight of the two bovine preparations is the same and 25% smaller than the porcine enzyme (Table II). The individual chains showed greater differences. Both chains of the porcine enzyme are larger than the corresponding bovine chains. In comparing the bovine enterokinase prepa- rations, our heavy chain is larger than the one described by the Seattle group while the reverse is true for the light chain.
Amino Acid Analysis-The amino acid composition of enterokinase is summarized in Table III. The heavy chain has 763 and the light chain 222 amino acids. Both chains contain all types of amino acids and galactosamine and glucosamine. The agreement between the sum of the residues of the chains and of intact enterokinase is satisfactory except for glycine, half-cystine, and serine. Enterokinase contained equal
by guest on June 9, 2018http://w
ww
.jbc.org/D
ownloaded from
Bovine Enterokinase 1681
TABLE III TABLE IV Composition of enterokinase
Units are residues per molecule of protein. Amino acid composition” of enterokinase from the bovine duodenal
mucosa,’ bovine duodenal contents,’ andporcine duodenal muco.sad
Amino acid Intact Heavy - Alanine 47.2 36.7 Arginine 24.8 14.7 Aspartic acid 110.7 87.5 Half-cystine 29.4” 31.1h Glutamic acid 102.8 82.0 Glycine 80.4 91.8 Histidine 15.8 11.2 Isoleucine’ 52.6 33.8 Leucine 72.6 52.8 Lysine 36.3 26.1 Methionine 12.6” 6.8h Phenylalanine 41.7 32.4 Proline 52.0 33.0 Serine” 78.0 88.4 Threonined 56.8 52.2 Tryptophan’ 11.6 Tyrosine 30.3 18.4 Valine’ 50.8 30.6 Neutral sugars (%) 18.0 Amino sugars” (%) 18.0 Glucosamine” 117.3 116.2 Galactosamined 33.6 57.4
n From a sample of the performic acid-oxidized enzyme.
Light
15.0 7.6
23.2 8.1b
29.1 26.5
4.7 9.8
14.8 6.4 3.4* 5.1
12.6 24.6
9.7
6.9 13.0
16.8 4.4
51.7 22.3
110.7 39.2
111.1 118.3
15.9 43.6 67.6 32.5 10.2 37.5 45.6
113.0 61.9
25.3 43.6
133.0 61.8
’ From a sample of the performic acid-oxidized enzyme after it was reduced and carboxymethylated.
’ Values were from 120 h of hydrolysis. ’ Values were for zero time obtained from the extrapolation of the
timed samples. e From a sample hydrolyzed with alkali (20). A value of 14.4
residues/molecule of protein was found with a spectrophotometric procedure (19); the light chain contained 3.3 and the heavy chain 9.4 residues.
amounts of neutral and amino sugars, 18% by weight. The sialic acid content was not determined.
In Table IV, the amino acid compositions of different en- terokinase preparations are given as moles of amino acid/ 100,000 g of protein to permit direct comparisons of one sample with another. A better comparison of different prepa- rations can be made with the amino acid composition of the light chain. Since the chains are smaller than the intact enzyme, a better precision of measurement is found. The agreement is very good for the values of our enzyme and the Marseille enzyme. Greater discrepancies are found in compar- ing our enzyme with the Seattle enzyme, particularly in the numbers for arginine, half-cystine, histidine, leucine, and thre- onine.
Enzymatic Properties-The enzymatic activity of highly purified enterokinase toward synthetic substrates containing lysine and arginine is given in Table V. Tos-Arg-OMe and Tos-Lys-OMe were hydrolyzed at the same rate. Bz-Arg-OEt was hydrolyzed almost 5 times faster than Tos-Arg-OMe, which is similar to the activity rates of porcine enterokinase (4). On the other hand, both bovine and porcine trypsin hydrolyze Tos-Arg-OMe about 4 to 10 times faster than Bz- Arg-OEt (31, 32).
The kinetic parameters of enterokinase for the activation of bovine trypsinogen were determined from an Eadie-Hofstee plot. The substrate was varied from 2 to 41 pM and the activation was performed at pH 5.6 and 25°C to minimize autolysis. Both the K,,, of 1.7 X 10e5 M and the kcat of 6.3 s-l are close to the values reported for porcine enterokinase (7 X 10m5 M and 4.8 s-l), but differ from the bovine duodenal contents enzyme (7 X lo-” M and 1.5 s-‘) (4, 10).
Samples of purified enterokinase were tested with several
Intact protein Light chain
Amino acid Bovine Bovine Porcine Bovine Porcine Ill”COSi3 contents mucosa mucosa mucosa
Alanine 31.4 30.2 32.8 42.9 43.4 Arginine 16.6 10.5 15.1 21.7 26.1 Aspartic acid 73.8 52.0 66.8 66.3 59.5 Half-cystine 20.5 14.0 23.0 23.1 7.9 Glutamic acid 68.6 59.2 63.8 83.1 67.4 Glycine 53.6 43.4 44.2 75.7 51.0 Histidine 10.6 19.7 9.5 13.4 11.5 Isoleucine 35.1 22.2 28.8 28.0 32.3 Leucine 48.4 26.1 46.8 42.3 49.7 Lysine 24.2 17.5 20.4 18.3 24.8 Methionine 8.8 4.9 8.2 9.7 8.1 Phenylalanine 27.8 19.7 27.8 14.6 18.2 Proline 34.7 43.0 34.1 36.0 34.7 Serine 52.0 57.0 43.3 70.3 38.5 Threonine 37.9 53.0 37.0 27.7 28.7 Tyrosine 20.2 17.4 18.6 19.7 18.9 Valine 33.9 35.8 33.3 37.1 36.9
” Moles of amino acid in 100,000 g of protein. b Enzyme preparation described under “Results.” ’ Enzyme preparation described by Seattle group (IO). d Enzyme preparation described by the Marseille group (8).
TABLE V
Activity of enterokinase toward small substrates
Substrate Bovine enterokinase Porcine” entero- Bovi,,e,j trypsin (oresent studv) kinase
Tos-Arg-OMe Bz-Arg-OEt Tos-Lys-OMe
a From Ref. 4. b From Ref. 30.
s-1 s-1 s ’
12 8.1 100 53 28.4 28 12 84
TABLE VI Effect ofproteinproteinase inhibitors on enterokinase activity
Protein inhibitor B Inhibition”
Kunitz PTI 100 Kunitz ST1 18 Porcine secretory PTI 13 Bowman-Birk ST1 12 Bovine secretory PTI 3 Lima bean inhibitor 2 Chicken ovomucoid 0
a Determined with 1 pM concentration of inhibitor and 6 mM
concentration of enterokinase at 25”C, pH 8.3, for 30 min. Enteroki- nase activity was determined by titration with Gdn-Bz-OMum.
trypsin proteinase inhibitors to see whether enterokinase would be inhibited. The results are given in Table VI. Only Kunitz PTI acted as an effective inhibitor. We determined a K,,,,, for the Kunitz PTI of 2 X 10” M,-' which makes PTI the best inhibitor of enterokinase found so far (benzamidine and p-aminobenzamidine were reported to have K,,,,, values of lo4 M-' (33)). The strong affinity of enterokinase toward PTI provides the rationale for our use of the inhibitor in the affinity chromatography step of the purification scheme.
TLCK is an active site-directed reagent which inhibits trypsin by forming a covalent derivative with the histidine of the active site (34). At a molar ratio of 2000 inhibitor to one of enzyme, TLCK inactivated enterokinase but at a slower rate than trypsin. Porcine enterokinase was also inhibited by
by guest on June 9, 2018http://w
ww
.jbc.org/D
ownloaded from

1682 Bovine Enterokinase
TLCK and this is part of the evidence that suggests that enterokinase is a member of the serine proteinase class (4).
DISCUSSION
Enterokinase is a membrane-bound protein located in the brush border of the intestinal mucosa. The enzymatic activity is highest in the tops of the villi and decreases in cells closer to the crypts (2). The tips of the microvilli are continuously desquamated which gives rise to the enterokinase activity found in the duodenal contents (2, 35). In the duodenal contents, the activated forms of the pancreatic zymogens and intestinal hydrolases are also present and these enzymes work- ing in concert are responsible for the terminal steps of protein, carbohydrate, and lipid digestion.
The porcine enterokinase and our bovine mucosal enzyme have many properties in common. The proteins have a heavy and light chain held together by one or more disulfide bonds (Fig. 3). Although the size differs, the amino acid composition of the heavy and light chains is very similar (Table IV). Noteworthy is the close resemblance of the light chains where all amino acids except the half-cystine, glutamic acid, glycine, and serine show good agreement. The similar composition of the light chains is significant since the light chain contains the residues of the active site. The Marseille group labeled the enzyme with [“‘Pldiisopropyl phosphorofluoridate and found the modified serine in the light chain (8). Both enzymes are also inhibited with the active site reagent TLCK. The enzy- matic properties are essentially the same toward trypsinogen (brat, K,), Bz-Arg-OEt, and Tos-Arg-OMe (specific activities) (Table V). However, the bovine enzyme was inhibited by PTI (Table VI), while the porcine enzyme was not inhibited by ST1 or PTI.
In contrast, the Seattle bovine preparation differs in several characteristics from our duodenal mucosa enzyme and also from the porcine preparation. The most significant differences are the size of the heavy and light chains (Table II) and the amino acid compositions of the enzymes (Table IV). Unfor- tunately, the composition of the individual chains was not reported by the Seattle group. The molecular weight differ- ences suggest that the heavy chain of the Seattle enzyme may represent a smaller degraded molecule than the one we found. However, the situation is reversed in comparing the molecular weights of the light chain. Since the light chain is the catalytic chain, and since the porcine enzyme and ours are similar in amino acid composition, it is difficult to imagine that the light chain of our enzyme is a degraded form of the Seattle chain. Furthermore, the size of the light chain of our enzyme is almost the same as bovine trypsin (23,600). It is more reason- able to consider the possibility that the Seattle enzyme and our enzyme are different forms of enterokinase.
We could only detect a single species displaying enteroki- nase activity in extracts of mucosal cells. Our purification scheme gave high recoveries of enzyme units at each step. Only a single active fraction was found following chromatog- raphy on DEAE-cellulose. However, affinity chromatography on PTI-Sepharose separated a bound fraction (major active component) from an unbound active fraction. After purifica- tion of the unbound fraction, it had properties identical with the homogeneous preparations of enterokinase.
In Marseille, porcine enterokinase was isolated initially from duodenal contents (4) and later from duodenal mucosa (8). Unfortunately, the two preparations were not compared but the isolation procedure and enzymatic properties suggest that the two preparations are the same. Therefore, if two forms of bovine enterokinase exist, they must differ to a much greater extent than the porcine preparations. Either bovine enterokinase is elaborated as isoenzymes or differences in
structure exist because of modifications caused by enzymes of the duodenal contents. Unfortunately, the lack of structural information on the two bovine preparations makes it impos- sible at this time to know if either of these explanations is true.
Since the light chain of enterokinase is the catalytic chain, it was of interest to see if the light chain bears any resemblance to bovine trypsin. The molecular weight of the light chain, after a correction is made for the carbohydrate content, is approximately 23,000. The amino acid composition of the light chain will be presented with the corresponding values of bovine trypsin given in parentheses. The light chain contains 15 alanine (14), 52 aspartic and glutamic acids (36), 27 glycine (25), 8 half-cystine (12), 5 histidine (3), 25 isoleucine and leucine (29), 14 lysine and arginine (16), 4 methionine (2), 5 phenylalanine (3), 13 proline (B), 34 serine and threonine (44), 12 tryptophan (4), and 7 tyrosine (10). The overall similarity of the composition of the two polypeptide chains is striking although proline and tryptophan differ to a large extent.
The light chain contains 8 half-cystine residues” while tryp- sin has 12. Of the 8, 1 or 2 would be the minimum number linked in disulfide bridges to the heavy chain. We favor a minimum of two interchain disulfides since one disulfide would require 1 half-cystine remaining as a free sulfhydryl group. We have no evidence for a cysteine residue in the light chain. The remaining 6 half-cystine residues would pair to produce three intramolecular disulfides; trypsin has six disul- fide bonds.’ The fewer disulfides of enterokinase are clearly within the range found with other serine proteinases. Human trypsin has five disulfides (36, 37), porcine pancreatic elastase four (38), and bacterial proteinases three and two (39, 40).
In viewing enterokinase as a two-chain structure, it is clear that the light chain is the catalytic chain and contains the serine and histidine of the active site. The heavy chain, on the other hand, may anchor enterokinase in the brush border membrane. The size of the chain is sufficiently large that a part of the chain could be embedded in the membrane. It is also possible that the heavy chain may be necessary for the light chain to function catalytically; the heavy chain may provide a proper binding region for the biologically active conformation of the light chain. Further structural studies should clarify the mechanism of action of the enzyme.
Acknowledgments-We wish to thank Mr. William Kohr for per- forming the amino acid analyses. We thank Ms. Pam and Audrey Light and Mr. William Yellig for help in scraping bovine intestines.
REFERENCES
1. Eichholz, A., and Crane, R. K. (1965) J. Cell. Biol. 26, 687-691 2. Nordstrom, C., and Dahlqvist, A. (1971) Biochim. Biophys. Acta
242, 209-225 3. Kunitz, M. (1939) J. Gen. Physiol. 22, 429-446 4. Maroux, S., Baratti, J., and Desnuelle, P. (1971) J. Bid. Chem.
246,5031-5039 5. Boyer, P.D., ed (1971) The h’nzymes, 3rd Ed, Vol. 3, Academic
Press, New York 6. Yamashina, I. (1956) Acta Chem. Stand. 10, 739-743 7. Neurath, H. (1975) in Proteases and Biological Control (Reich,
E., Rifkin, D. B., and Shaw, E., eds) Vol. 2, pp. 51-64, Cold Spring Harbor, N. Y.
8. Baratti, J., Maroux, S., Louvard, I~., and Desnuelle, 1’. (1973) Biochim. Biophys. Acta 315, 147-161
9. Baratti, J., and Maroux, S. (1976) Biochim. Biophys. Acta 452,
-
’ The half-cystine content is difficult to determine with high pre- cision. If the recoveries were low and the number of residues is greater than 8, then the number of intramolecular disulfides must be in- creased. Since the light chain appears to be similar to trypsin, which has six disulfides (12 half-cystines), a larger half-cystine content would increase the resemblance still further.
by guest on June 9, 2018http://w
ww
.jbc.org/D
ownloaded from

Bovine Enterokinase I (iH:i
488-496 10. Anderson, L. E., Walsh, K. A., and Neurath, H. (1977) Hiochem-
istry 16, 3354-3360 11. Dayhoff, M. 0. (1972) Aflczs of Protein Sequence and Structure,
Vol. 5, National Biomedical Research Foundation, Silver Spring, Md.
12. Chase, I’., Jr., and Shaw, E. (1970) Methocls &?nzyymol. 19, 20-27 13. Walsh, K. A., and Wilcox, 1’. E. (1970) Met/m& fi::nzynwl. 19, 3-
41 14. Jameson, G. W., Roberts, 1). V., Adams, R. W., Kyle, W. S. A.,
and Elmore, 1). T. (1973) Biochem. J. 131, 107-117 15. Folk. J. TX.. Gladner, J. A.. and Viswanatha. 1‘. (1959) Rzochrm.
Biophys. Actn 36, 256-257 16. March, S. C., Parikh, I., and Cuatrecasas, I’. (1974) Anal. Bio-
them. 60, 149-152 17. Walsh, K. A., and Brown, J. 1% (1962) Biochim. Biophys. Acta
58, 596498 18. Moore, S. (1963) ,/. Biol. Chern. 238, 235-237 19. Beaven, G. H., and Holiday, E. H. (1952) A&,. Protein Chem. ‘7,
319-386 20. Hugli, 1’. E., and Moore, S. (1972) ,I. BloZ. Chem. 247, 2828-2834 21. Francois. C., Marshal, R. I)., and Neuberger. A. (1962) Biochem.
J. 83, X%5-341 22. Hirs, C. H. W. (1970) Methods h’nzymol. 19, 199-203 23. Weber, K., and Osborn, M. (1969) <J. Biol. Chem. 244, 4406-4412 24. Segrest, J. I’., and ,Jackson, K. L. (1972) Methods Enzyme/. 28,
54-63
25.
26.
27. 28.
29.
30.
31. 32. 33. 34.
35. 36.
37.
38. 39.
40.
Carpenter, F. H., and Harrington. K. T. (1971) .J. RLOI (‘h<s/,,. 247 55X0-5586
Daviei, G. E., and Stark. G. H. (1970) Proc. Nntl. ilcud ,‘+I I S. A. 66,651-656
Davis, B. J. (1964) Ann. N. Y. Acud. Sci. 121, 40+4X Louvard, D., Maroux, S., Baratti, J., and Desnuc~llr. 1’ ( 197.i)
Biochim. Biophvs. Acta 309, 127-137 Schmitz. J., Preiser, H., Maestracci, I)., Crane. It. K., l’rorsch. 1’ ,
and Hadorn, B. (1974) Biochim. Biophys. Act0 343, 435-4:111 Hatfield, L. M., Banerjee, S. K.. and Light. A. (1971) .I. Ijrr,/
Chem. 246, 6303-6312 Lazdunski, M. (1965) Bull. Sot. Chin?. Hlol. 47, :101-:11!! Knights, R. J., and Light, A. (1976) ,I. Bd. (‘hem. 251, 222.-22~ Geratz, J. D. (1967) Arch. Biochern. Biophys. 118, 9%9f; Shaw, E., Mares-Guia, M., and Cohen, W. i 1965) Hrwhwrrrstr:~ 4,
2219-2224 Nordstrom, C. (1972) Biochim. Bwphys. .4ctcz 268, 71 i-718 Guy, O., Lombardo. D., Bartelt, I>. C., Amic, J., and Figarella. I ‘,
(1978) Biochemistry 17, 1669-1675 Brodrick, J. W., Largman, C., ,Johnson, J. H.. and (;rokas. M (‘
(1978) J. B&Z. Chem. 253, 2732-2736 Shotton, D. M., and Hart1e.y. B. S. (1970) h’nturc 225, RX-801; Olson, M. 0. J., Nagabhushan. N.. Dzwiniel, M., Smillie, I,. I{
and Whitaker. D. R. (1970) Nnturr 228. 438-442 Jurasek, L., Carpenter, M. R., Smillie, I,. H.. Gertlrr, A I,rvv. S..
and Ericsson. I,. H. (1974) Biochem. Hiophys Kcx (‘ommro~ 61. 1095-l 100
by guest on June 9, 2018http://w
ww
.jbc.org/D
ownloaded from

J J Liepnieks and A LightThe preparation and properties of bovine enterokinase.
1979, 254:1677-1683.J. Biol. Chem.
http://www.jbc.org/content/254/5/1677Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/254/5/1677.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 9, 2018http://w
ww
.jbc.org/D
ownloaded from