presence of kdr and s-kdr resistance in musca domestica ... · presence of kdr and s-kdr resistance...
TRANSCRIPT

Bulletin of Insectology 68 (1): 65-72, 2015 ISSN 1721-8861
Presence of kdr and s-kdr resistance in Musca domestica populations collected in Piacenza province (Northern Italy)
Emanuele MAZZONI1, Olga CHIESA
1, Vincenzo PUGGIONI
1, Michela PANINI
1, Gian Carlo MANICARDI
2,
Davide BIZZARO3
1Dipartimento di Scienze delle Produzioni Vegetali Sostenibili - Area Protezione sostenibile delle piante e degli ali-
menti, Università Cattolica del Sacro Cuore, Piacenza, Italy 2Dipartimento di Scienze della Vita, Università di Modena e Reggio Emilia, Reggio Emilia, Italy
3Istituto di Biologia e Genetica, Università Politecnica delle Marche, Ancona, Italy
Abstract
Pyrethroid insecticides combine a high efficacy against insects with a low toxicity towards warm-blooded vertebrates. For this
reason they are largely used to control housefly infestations. The efficacy of these products is affected by insecticide resistance
mechanisms. In particular, some mutations in the sodium channel coding sequence are responsible for target-site resistance to
pyrethroid insecticides. This work presents a new molecular approach based on allele-specific PCRs to point out and to
characterise this resistance mechanism in two M. domestica populations (PNT and TRV) collected near Piacenza (Northern Italy).
The presence of different kdr and s-kdr genotypes was assessed for population PNT whilst kdr only was detected in population
TRV. Dose-response bioassays evidenced quite high resistance factors, expecially for population PNT. This is in line with the kdr
and s-kdr frequencies observed in the assayed populations.
Key words: housefly, insecticide resistance, sodium channel, PASA, haplotypes, Italy.
Introduction
The common housefly Musca domestica L. (Diptera
Muscidae) is a well known pest for health threat as it is
capable to transmit and to distribute many human and
animal pathogens in the environment (Graczyk et al.,
2001; Forster et al., 2007). This insect usually finds in
farms, dairies and also urban dumps suitable environ-
ments for the development and rapid growth of its popu-
lations (Raspi and Belcari, 1989; Trentini et al., 1999;
Cao et al., 2006). For this reason, nuisance to livestocks
and people living in the surroundings cannot be ne-
glected (Howard, 2001; Winpisinger et al., 2005).
Insecticide applications are commonly used to control
housefly populations. In the past, organochloride insec-
ticides like DDT (IRAC MOA 3B, www.irac-online.org)
have been widely and intensively used to limit the
spread of this species. Today, pyrethroid insecticides
(IRAC MOA 3A) represent an important part of the
chemicals adopted for housefly control, as they combine
high efficacy against this pest with a quite low toxicity
towards warm-blooded vertebrates. In addition, this
class of products avoid the long term environmental pol-
lution caused by organochlorine insecticides.
Di-phenyl ethane (like DDT) and pyrethroid insecti-
cides act as modulators of the voltage-gated sodium
channels, membrane proteins that are responsible for the
transmission of nerve impulses. Their interaction with
the channel protein result in electrical signalling altera-
tion that culminates with the paralysis and the conse-
quent dead of the insect.
Unfortunately, insecticide treatments cannot be so ef-
ficacious because of unsatisfactory environment man-
agement strategies that do not consider the high growth
rate of this pest and also because insecticide resistance
mechanisms have been developed and selected in
housefly populations. During the years, the insecticide
abuse have exerted high selection pressure on the popu-
lations selecting different resistant mechanisms, due to
an increased production of detoxifying enzymes or to
structural changes in the pesticide target proteins (Liu
and Pridgeon, 2002). The most important mechanism of
resistance to pyrethroids is target-site insensitivity con-
ferred by mutations which occur in the sodium channel
coding sequence. They cause a reduction of the insecti-
cide binding affinity, compromising the effectiveness of
the insecticide applications and seriously affecting the
integrated management strategies.
Several point mutations have been described in this
target, in M. domestica as well as in many others insect
species (Davies et al., 2007; Soderlund, 2012; Rinke-
vich et al., 2013; Dong et al., 2014). The two most im-
portant non-synonymous mutations found to confer py-
rethroid resistance were first described in housefly and
are known as “knock-down resistance” (kdr) and “su-
per-kdr” (s-kdr) (Williamson et al., 1996). They are re-
sponsible for amino acid substitutions located in the
same functional domain: L1014F for the former and
M918T for the latter (Soderlund and Knipple, 2003).
In houseflies there is an additional mutation located in
the kdr locus and involved in pyrethroid resistance. It is
referred as “kdr-his” and causes a different amino acid
replacement, from Leu to His (L1014H) (Liu and Prid-
geon, 2002; Rinkevich et al., 2012).
Insecticide bioassays do not provide enough informa-
tion about the presence of these mutations and their alle-
lic variants, while a rapid identification is possible by
using a biomolecular approach. Different methods have
been developed in order to detect kdr mutations: PCR-
RFLP, allele-specific PCRs or direct sequencing (Huang
et al., 2004; Rinkevich et al., 2006; Qiu et al., 2012).
On the contrary, till now s-kdr has been detected only

66
by direct sequencing (Rinkevich et al., 2006). In M.
domestica, the analysis of mutation frequencies in volt-
age gated sodium channel can give important informa-
tion about the resistance levels of the analysed popula-
tions and it could be an interesting tool to evaluate the
efficacy of control strategies based on pyrethroid appli-
cations. Recent insecticide bioassays carried on an Ital-
ian population of M. domestica collected from a poultry
farm showed the presence of high resistance levels to-
wards several active ingredients but no evidences about
the resistance mechanisms involved were provided
(Pezzi et al., 2011).
Our study aims to point out the presence and involve-
ment of target-site resistance to pyrethroid insecticides
in two populations of M. domestica collected near
Piacenza (Northern Italy, Po valley), analysed with a
new molecular approach which allows the detection of
the three known amino acidic substitutions in the volt-
age gated sodium channel.
Materials and methods
Insects: origin and rearing Adult houseflies were collected in 2010 in two live-
stock farms located in Piacenza province. The first one, a
modern stable which uses regular insecticide applications
in order to control the housefly populations, was near a
small town named Pontenure (PNT). The second one, a
small traditional stable not applying any control strate-
gies against flies, was near Trevozzo village (TRV).
For each sampling area, a population was originated
from the collected insects and maintained in rearing
cages under controlled conditions (21 °C ± 0.5 °C), with
water and dry mixture of sucrose and milk powder (1:1)
always available. A mixture of bran and milk powder
(4:1) moistened with tap water was used as oviposition
substrate. Eggs were collected every 2 days and trans-
ferred into a rearing substrate (bran: 25% w/w; milk
powder: 5% w/w; water: 70% w/w). Puparia were col-
lected regularly and transferred to another cage for the
next generations.
A susceptible population (S-WHO), used as reference,
was kindly provided by Ralf Nauen (Bayer Crop Sci-
ence, Monheim, Germany) and was maintained in the
same conditions.
Genomic extraction Genomic DNA was extracted from the heads of indi-
vidual adult, following a „salting-out‟ protocol already
described (Panini et al., 2014). Heads were homoge-
nized using a QIAGEN TissueLyser LT for 30 s at 50
Hz in a 2 mL tube containing one stainless steel bead
and 300 μL TNES buffer pH 7.5 (50 mM Tris, 400 mM
NaCl, 20 mM EDTA, 0.5% SDS) with proteinase K
(100 μg mL−1
). Homogenate was heated at 55 °C for 1 h
and then proteins were precipitated with 85 μL NaCl 5
M and pelleted at 16 000 × g for 5 min. DNA was iso-
lated from the supernatant by ethanol precipitation and
resuspended in 50 μL sterile water. The DNA concen-
tration was assessed using a Qubit Fluorimeter 2.0 in-
strument (Quant-iT ds DNA HS Assay kit; Invitrogen,
Carlsbad, CA, USA). The amount of genomic DNA ob-
tained was in the range 2–50 ng µL−1
.
PASA-PCR The presence of kdr (L1014F and L1014H) and s-kdr
(M918T) single point mutations was assessed with al-
lele specific polymerase chain reaction amplification
(PASA-PCR). A set of primers was designed (table 1)
using a cDNA M. domestica sodium channel gene cod-
ing sequence (Gene Bank accession number:
NM_001286885.1) and primer sequences from Huang
et al. (2004). A schematic representation of the primer
positions is reported in figure 1. The gene structure was
analysed by using Spidey (Wheelan et al., 2001), which
provide tools for the alignment between mRNA and ge-
nomic DNA sequences.
PASA-PCR mixtures contained 0.4 μM of each
primer, 1 μL of genomic DNA and 12.5 μL Dream Taq
Green PCR Master Mix (Thermo Scientific, Milan, It-
aly) in a total reaction volume of 25 μL. For both targets
amplification started with 2 min at 94 °C, followed by
30 cycles of 94 °C for 30 s, 60 °C for 30 s and 72 °C for
30 s, with a final elongation at 72 °C for 10 min.
Results of PASA-PCRs were validated by sequencing
the control fragments directly amplified with the exter-
nal primers, encompassing the mutations under investi-
gation, obtained from some samples randomly selected.
The obtained sequences were then aligned with the ge-
nomic reference sequence (Gene Bank accession num-
bers: NW_004774263.1 and NW_004765908.1) using
ClustalW (Thomson et al., 1994).
Table 1. Primers (5‟-3‟) used for kdr and s-kdr characterisation. Nucleotides reported in bold uppercase indicate their
specificity for the susceptible or the resistant alleles. The position of the primers is reported as the nucleotide num-
ber of the cDNA sequence (Gene Bank accession number: X96668).
Target Primer Direction Sequence 5'-3' Position References
kdr K1 FW tcgcttcaaggaccatgaat 2881-2901
kdr K2 RE ttacgtttcacccagttctta 3178-3199
kdr K3 FW acggtcgtgatcggcaatC 3035-3054 Huang et al., 2004
kdr K4 FW acggtcgtgatcggcaatT 3035-3054 Huang et al., 2004
kdr HF_KsH_F FW cggtcgtgatcggcaatcT 3036-3055
kdr HF_KrH_F FW cggtcgtgatcggcaatcA 3036-3055
s-kdr SK1 FW ttcgtgtattcaaattggcaaa 2709-2731
s-kdr SK2 RE cgaaaagttgcattcccatc 2836-2856
s-kdr SK3 RE acccattgtccggcccA 2767-2783
s-kdr SK4 RE acccattgtccggcccG 2767-2783
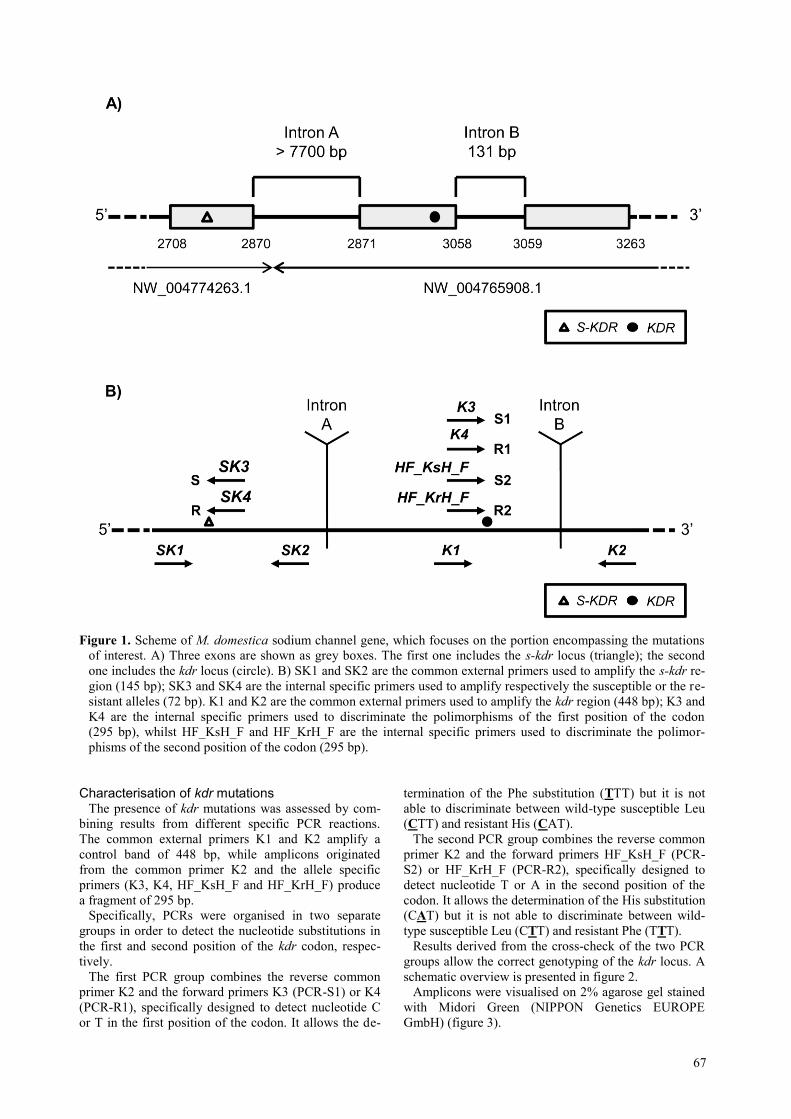
67
Figure 1. Scheme of M. domestica sodium channel gene, which focuses on the portion encompassing the mutations
of interest. A) Three exons are shown as grey boxes. The first one includes the s-kdr locus (triangle); the second
one includes the kdr locus (circle). B) SK1 and SK2 are the common external primers used to amplify the s-kdr re-
gion (145 bp); SK3 and SK4 are the internal specific primers used to amplify respectively the susceptible or the re-
sistant alleles (72 bp). K1 and K2 are the common external primers used to amplify the kdr region (448 bp); K3 and
K4 are the internal specific primers used to discriminate the polimorphisms of the first position of the codon
(295 bp), whilst HF_KsH_F and HF_KrH_F are the internal specific primers used to discriminate the polimor-
phisms of the second position of the codon (295 bp).
Characterisation of kdr mutations The presence of kdr mutations was assessed by com-
bining results from different specific PCR reactions.
The common external primers K1 and K2 amplify a
control band of 448 bp, while amplicons originated
from the common primer K2 and the allele specific
primers (K3, K4, HF_KsH_F and HF_KrH_F) produce
a fragment of 295 bp.
Specifically, PCRs were organised in two separate
groups in order to detect the nucleotide substitutions in
the first and second position of the kdr codon, respec-
tively.
The first PCR group combines the reverse common
primer K2 and the forward primers K3 (PCR-S1) or K4
(PCR-R1), specifically designed to detect nucleotide C
or T in the first position of the codon. It allows the de-
termination of the Phe substitution (TTT) but it is not
able to discriminate between wild-type susceptible Leu
(CTT) and resistant His (CAT).
The second PCR group combines the reverse common
primer K2 and the forward primers HF_KsH_F (PCR-
S2) or HF_KrH_F (PCR-R2), specifically designed to
detect nucleotide T or A in the second position of the
codon. It allows the determination of the His substitution
(CAT) but it is not able to discriminate between wild-
type susceptible Leu (CTT) and resistant Phe (TTT).
Results derived from the cross-check of the two PCR
groups allow the correct genotyping of the kdr locus. A
schematic overview is presented in figure 2.
Amplicons were visualised on 2% agarose gel stained
with Midori Green (NIPPON Genetics EUROPE
GmbH) (figure 3).

68
Figure 2. Schematic representation of the amplification
obtained with the PCRs used in this study. Black
boxes indicate positive amplification for the corre-
sponding genotype. PCRs S1 and R1 are used to dis-
criminate nucleotide C/T in the first position of the
codon; PCRs S2 and R2 are used to discriminate nu-
cleotide T/A in the second position of the codon.
Characterisation of s-kdr mutations The presence of the s-kdr mutation was assessed by
using 3 different PCR reactions. The combination of the
common external primers SK1 and SK2 amplifies a 145
bp control band . The combination of SK1 with SK3 or
SK4 amplifies respectively the susceptible or the resis-
tant allele, both producing 72 bp bands. Amplicons were
visualised on 3.5% agarose gel stained with Midori
Green (figure 4).
Insecticides bioassays Susceptibility toward technical grade methoxychlor
(Sigma-Aldrich) was investigated by using topical ap-
plication bioassays, according to Huang et al. (2004).
Insecticide solution in acetone (1 µL) was applied on
the notum of three days old females with an Hamilton
micro-syringe. For each experiment, 5-6 different con-
centrations of insecticide were prepared and used to
treat pools of 20 houseflies; a control group treated with
acetone only was included in each replicates. Each
group was maintained inside a vented plastic cylinder,
with water and food (dry milk and sucrose, 1:1 ratio)
supplied ad libitum and mortalities were assessed 24
hours after the treatment.
The experiment was replicated at least 4 different
times for each population (PNT, TRV and S-WHO).
Data obtained from the bioassays were pooled together,
corrected for control mortality by using Abbott‟s for-
mula (Abbott, 1925) and concentration-mortality rela-
tionships were estimated by probit analysis using
POLO-Plus software (LeOra Software, Berkeley, CA)
(Finney, 1971).
Results and discussion
Genotyping of kdr and s-kdr mutations Three different alleles for the kdr locus are known in
M. domestica: the wild-type codon „CTT‟ (Leu) and two
different resistant codons „TTT‟ (Phe) and „CAT‟ (His)
(Rinkevich et al., 2006).
In the TRV population (n = 30 specimens) only two
kdr alleles were found: 40% of the analysed individuals
carried the susceptible allele L1014, whilst 60% of them
showed the presence of the resistant allele F1014. In
particular, the majority of the flies was heterozygous
resistant (53%) and a small percentage carried the ho-
mozygous resistant genotype (7%) (table 2).
On the contrary, all the three possible kdr alleles were
detected in the PNT population (n = 40 specimens), with
different genotype combinations. The most diffuse
genotypes were L1014 + F1014 (35%) and F1014 +
H1014 (32.5%). In addition, homozygous resistant flies
for both amino acidic substitutions were found: 17.5%
for the F1014 allele and 7% for the H1014 allele (table
2).
The same flies were analysed for the presence of the
s-kdr mutation (M918T). It was found only in the PNT
population: 50% of the individuals showed a heterozy-
gous genotype and 2.5% a homozygous resistant geno-
type (table 3).
In addition, the mutation was found only in the pres-
ence of kdr F1014. No specimens bearing s-kdr muta-
tion without kdr (F1014) have been found. This is in
Table 2. Percentage of specimens with different kdr
genotype combinations in the two analysed popula-
tions, TRV (n = 30) and PNT (n = 40).
kdr genotype flies (%)
TRV PNT
L/L 40 2.5
L/F 53.3 35
L/H 0 5
F/F 6.7 17.5
F/H 0 32.5
H/H 0 7.5
Table 3. Percentage of specimens with different s-kdr
genotype combinations in the two analysed popula-
tions, TRV (n = 30) and PNT (n = 40).
s-kdr genotype flies (%)
TRV PNT
M/M 100 47.5
M/T 0 50
T/T 0 2.5
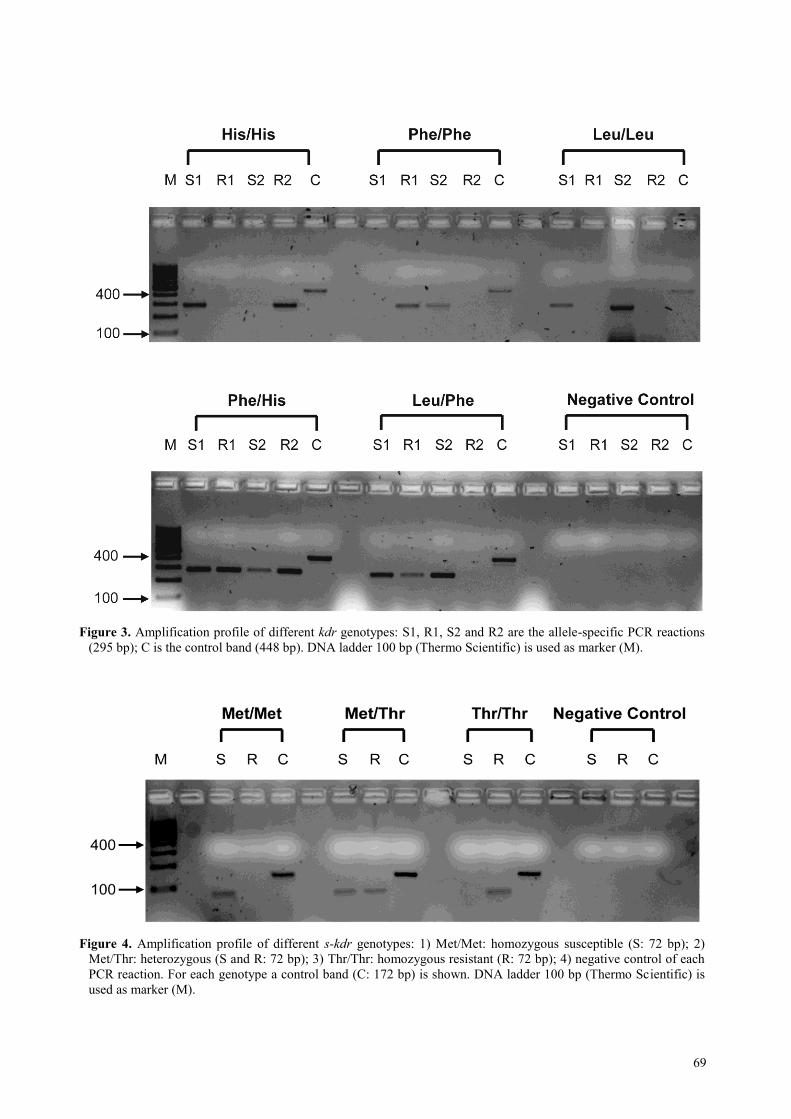
69
Figure 3. Amplification profile of different kdr genotypes: S1, R1, S2 and R2 are the allele-specific PCR reactions
(295 bp); C is the control band (448 bp). DNA ladder 100 bp (Thermo Scientific) is used as marker (M).
Figure 4. Amplification profile of different s-kdr genotypes: 1) Met/Met: homozygous susceptible (S: 72 bp); 2)
Met/Thr: heterozygous (S and R: 72 bp); 3) Thr/Thr: homozygous resistant (R: 72 bp); 4) negative control of each
PCR reaction. For each genotype a control band (C: 172 bp) is shown. DNA ladder 100 bp (Thermo Scientific) is
used as marker (M).

70
agreement with literature data and support the hypothe-
sis of the secondary origin of s-kdr mutation in respect
with kdr mutation and also that the presence of kdr is
compulsory to fix s-kdr mutation in the same allele (Lee
et al., 1999).
Haplotypes Some of the results obtained with allele-specific PCRs
were confirmed by direct sequencing of the control band
amplified with the external primers. Primers K1 and K2,
encompassing the kdr locus, amplify a region of 448 bp
and it is comprehensive of an intron of about 131 bp (in-
tron B) (figure 1) that has been described in the litera-
ture as a very high variable region. More than 120 hap-
lotypes, in several cases showing also insertions as well
as delections, have been reported till now (Rinkevich et
al., 2012).
Sequences derived from Italian samples carrying dif-
ferent kdr genotypes were aligned with all the housefly
sequences now available in GeneBank. The alignment
of intronic region B showed that the Italian specimens
with the susceptible homozigous genotype (Leu/Leu,
e.g. TRV 8) present the same intronic region of a Turk-
ish sample with same genotype (v11, accession number:
AY850269.2). Furthemore, the Italian samples with the
resistant homozigous (Phe/Phe, e.g. PNT 5) and het-
erozygous genotype (Phe/His, e.g. PNT 9) shoved the
same intronic region of samples derived from Turkey,
China and USA (kdr2, accession number: AY850261.2)
(Rinkevich et al., 2012) (figure 5).
Samples carrying genotypes Leu/Leu, Phe/Phe and
Phe/His showed chromatograms with single peaks; on
the contrary, other samples with different genotypes
(Leu/Phe and His/His) showed the presence of double
peaks which suggest the presence of two different in-
tronic region in the same specimen (data not shown).
Figure 5. Alignment of intronic region B of all the sequences available in GeneBank. Intronic region variations are
in grey boxes. The symbols identify two different haplotypes: samples PNT 5 (Phe/Phe) and PNT 9 (Phe/His) have
the same intronic sequence of AY805261.2 (kdr2, triangle) whilst TRV 8 (Leu/Leu) has 100% identity with
AY850269.2 (v11, circle).
71
Table 4. Log-dose probit-mortality data derived from methoxychlor topical application bioassays against adult flies.
LC50: lethal concentration that is expected to cause 50% of mortality; CI 95%: confidence interval limits at 95%;
R.F.: resistance factor calculated as the ratio between LC50 of resistant strain and LC50 of susceptible strain S-WHO.
Population LC50 (g L-1
) CI 95% Slope d.f. χ2 R.F.
S-WHO 0.43 0.30 - 0.61 1.36 ± 0.15 21 14.1 =
TRV 86.4 39.3 - 286 0.71 ± 0.12 14 18.6 201
PNT 1240 346 - 50600 0.67 ± 0.16 14 14.2 2876
This data suggest that at least two different alleles as-
sociated with mutation L1014H are present in population
PNT. Possibly, one of the allele is the same found in the
intronic region of kdr2 (see above), which correspond to
the Italian specimens with Phe/His genotype.
These results are in agreement with already published
data: the intronic sequence of kdr2 haplotype was
found to be the same for different kdr alleles (Leu, Phe,
His), whilst the intronic region of v11 haplotype was
found to be associated only with Leu (Rinkevich et al.,
2012).
Bioassays Preliminary bioassays were performed to estimate a
valid insecticide dose range. In the case of the suscepti-
ble population (S-WHO) the optimal range of technical
active ingredient was from 0.001 g L-1
to 10 g L-1,
while
for populations PNT and TRV the optimal range was
from 1 g L-1
to 200 g L-1
. Parameters of the estimated
“log-probit” mortality curves are reported in table 4.
The LC50 of population TRV, for which no control
strategies have been adopted, was lower than the LC50
of population PNT, derived from a farm that regularly
uses insecticide applications. Both of them were signifi-
cantly higher than the LC50 of the susceptible strain S-
WHO. Estimated resistance factors (RFs) were about
200 for population TRV and more than 2800 for popula-
tion PNT (table 4).
Conclusions
Pyrethroids are commonly used insecticides to control
houseflies. Target-site mutations responsible for pyre-
throid resistance have been documented in samples col-
lected in different locations worldwide (Ahmed and
Wilkins, 2002; Shono et al., 2002; Rinkevich et al.,
2007; 2012) but just a few information are available re-
garding the Italian situation (Pezzi, 2011) and no data
confirm the presence of the most common mutations
which are known to confer pyrethroid insensitivity.
This work evaluated the presence of kdr, kdr-his and
s-kdr mutations in Italian samples collected in livestock
farms of the Po valley. Even if only two populations
have been considered, the presence of target-site muta-
tions was reported, demonstrating for the first time their
involvement in pyrethroid resistance also in Italy. Inter-
estingly, one of those population was collected in a farm
where no insecticide treatments have been used, sug-
gesting the spread of target-site resistance around the
territory.
The molecular approach here reported represents a
possible alternative to the classic technologies till now
adopted to investigate the above reported mutations. In
fact, the allele-specific PCRs allow the rapid genotyping
of the kdr and s-kdr alleles with the reduction of the
costs required for the direct sequencing or for a PCR-
RFLP analysis. Moreover, our set of primers can detect
not only the classic kdr mutation (L1014F) but also the
other described amino acid replacement (L1014H) in
the same locus, thus avoiding the overestimation of the
susceptible allele and overcoming PCR-RFLP limitation
(Rinkevich et al., 2006).
The allele frequencies of kdr and s-kdr reported in
these two populations support the level of resistance that
have been found in dose-response insecticide bioassays
with methoxychlor, a chlorinated insecticide affecting
the same target of pyrethroids.
Significant resistant factors were estimated for both
the assayed populations but the highest resistance factor
was detected in population PNT, collected from a farm
which usually uses insecticide to control housefly popu-
lations. These results are in agreement with the calcu-
lated kdr and s-kdr frequencies: in population TRV only
kdr mutation was detected, and the resistant allele was
present in 60% of the analysed specimens, whilst popu-
lation PNT showed a consistent presence of kdr and s-
kdr mutations, with more than 90% of the individuals
carrying different kdr resistant genotypes and more than
50% the s-kdr mutation.
The study demonstrated the association of the resis-
tance-attributable mutations in the sodium channel gene
with insensitivity to methoxychlor insecticide. As it
shares the same mode of action of pyrethroids (IRAC 3
- sodium channel modulators), our results suggest the
possible involvement of the same mutations also in py-
rethroid resistance. We cannot exclude the involvement
of different resistance mechanisms like detoxification
systems based on P450 enzymes. Further investigations
with different insecticide products would be necessary,
including pre-treatment with PBO in order to better ex-
plore the influence of metabolic resistance.
Even if this study is based on a little number of popula-
tions, it confirms the presence of different genotype
combinations of target-site mutations that can influence
the efficacy of pyrethroids applications. Furthermore, the
intronic analysis of the region closed to the kdr locus
confirms that the haplotype variability, already detected
worldwide, is also present in our samples. Those results
support the multiple origin of target-site resistance al-
ready discussed by other authors (Rinkevich et al.,
2012). Moreover, the s-kdr mutation was not found to be
associated with a specific intronic region, supporting the
theory that it evolved from different kdr alleles.

72
The establishment of different combinations of resis-
tant alleles could be due to crosses among housefly
populations that are capable to spread around different
geographical areas and it is enhanced by their high re-
productive potential.
Additional analysis of a higher number of housefly
populations collected around Italy are then required to
better explain these aspects, to provide more details
about the current insecticide resistance situation in this
pest and to adopt and implement more efficacious insec-
ticide resistance management strategies.
Acknowledgements
The authors are grateful to Ralf Nauen (Bayer CropS-
cience AG, Global Biology Insecticides, Monheim,
Germany) for providing susceptible population S-WHO.
References ABBOTT W. S., 1925.- A method of computing the effectiveness of
an insecticide.- Journal of Economic Entomology, 18: 265-267.
AHMED S., WILKINS R., 2002.- Studies on some enzymes in-
volved in insecticide resistance in fenitrothion-resistant and -
susceptible strains of Musca domestica L. (Dipt, Muscidae).-
Journal of Applied Entomology, 126: 510-516.
CAO X. M., SONG F. L, ZHAO T. Y., DONG Y. D., SUN CH. X., LU
B. L., 2006.- Survey of deltamethrin resistance in house flies
(Musca domestica) from urban garbage dumps in Northern Chi-
na.- Environmental Entomology, 35: 1-9.
DAVIES T. G. E., FIELD L. M., USHERWOOD P. N. R., WILLIAM-
SON M. S., 2007.- DDT, pyrethrins, pyrethroids and insect so-
dium channels.- IUBMB Life, 59: 151-162.
DONG K., DU Y., RINKEVICH F., NOMURA Y., XU P., WANG L.,
SILVER K., ZHOROV B. S., 2014.- Molecular biology of insect
sodium channels and pyrethroid resistance.- Insect Biochemistry
and Molecular Biology, 50: 1-17.
FINNEY D. J., 1971.- Probit analysis.- Cambridge University
Press, Cambridge, UK.
FORSTER M., KLIMPEL S., MEHLHORN H., SIEVERT K., MESSLER
S., PFEFFER K., 2007.- Pilot studies on synantropic flies (e.g.
Musca, Sarcophaga, Calliphora, Fania, Lucilia, Stomoxys) as
vectors of pathogenic microorganisms.- Parasitology Research,
101: 243-246.
GRACZYK T. K., KNIGHT R., GILMAN R. H., CRANFIELD M. R.,
2001.- The role of non-biting flies in the epidemiology of hu-
man infectious diseases.- Microbes and Infection, 3: 231-235.
HOWARD J., 2001.- Nuisance flies around a landfill: patterns of
abundance and distribution.- Waste Management and Research,
19: 308-313.
HUANG J., KRISTENSEN M., QIAO C. L., JESPERSEN J. B., 2004.-
Frequency of kdr gene in house fly field populations: correla-
tion of pyrethroid resistance and kdr frequency.- Journal of
Economic Entomology, 97: 1036-1041.
LEE S. H., SMITH T. J., KNIPPLE D. C., SODERLUND D. M., 1999.-
Mutations in the house fly Vssc1 sodium channel gene associated
with super-kdr resistance abolish the pyrethroid sensitivity of
Vssc1/tipE sodium channels expressed in Xenopus oocytes.- In-
sect Biochemistry and Molecular Biology, 29: 185-194.
LIU N., PRIDGEON J. W., 2002.- Metabolic detoxication and the
kdr mutation in pyrethroid resistant house flies, Musca domes-
tica (L.).- Pesticide Biochemistry and Physiology, 73: 157-163.
PANINI M., DRADI D., MARANI G., BUTTURINI A., MAZZONI E.,
2014.- Detecting the presence of target site resistance to neoni-
cotinoids and pyrethroids in Italian populations of Myzus persi-
cae.- Pest Management Science, 70: 931-938.
PEZZI M., LANFREDI M., CHICCA M., TEDESCHI P., BRANDOLINI
V., LEIS M., 2011.- Preliminary evaluation of insecticide resis-
tance in a strain of Musca domestica (Diptera: Muscidae) from
an intensive chicken farm of Northern Italy.- Journal of Envi-
ronmental Science and Health B, 46: 480-485.
QIU X., PAN J., LI M., LI Y, 2012.- PCR-RFLP methods for detec-
tion of insecticide resistance-associated mutations in the house
fly (Musca domestica).- Pesticide Biochemistry and Physiology,
104: 201-205.
RASPI A., BELCARI A., 1989.- Alcune osservazioni sulla composi-
zione quali-quantativa della ditterofauna presente in stalle di bo-
vini per la produzione del latte.- Frustula Entomologica, 12: 1-13.
RINKEVICH F. D., ZHANG L., HAMM R. L., BRADY S. G., LAZZARO
B. P., SCOTT J. G., 2006.- Frequencies of the pyrethroid resistance
alleles of Vssc1 and CYP6D1 in house flies from the eastern
United States.- Insect Molecular Biology, 15 (2): 157-167.
RINKEVICH F. D., HAMM R. L., GEDEN C. J., SCOTT J. G., 2007.-
Dynamics of insecticide resistance alleles in house fly popula-
tions from New York and Florida.- Insect Biochemistry and Mo-
lecular Biology, 37: 550-558.
RINKEVICH F. D., HEDTKE S. M., LEICHTER C. A., HARRIS S. A.,
SU C., BRADY S. G, TASKIN V., QIU X., SCOTT J. G., 2012.-
Multiple origins of kdr-type resistance in the house fly, Musca
domestica.- PLoS ONE, 7: e52761.
RINKEVICH F. D., DU Y., DONG K., 2013.- Diversity and conver-
gence of sodium channel mutations involved in resistance to py-
rethroids.- Pesticide Biochemistry and Physiology, 106: 93-100.
SHONO T., KASAI S., KAMIYA E., KONO Y., SCOTT J. G., 2002.-
Genetics and mechanisms of permethrin resistance in the YPER
strain of house fly.- Pesticide Biochemistry and Phisiology, 73:
27-36.
SODERLUND D. M., 2012.- Molecular mechanisms of pyrethroid
insecticide neurotoxicity: recent advances.- Archives of Toxi-
cology, 86: 165-181.
SODERLUND D. M., KNIPPLE D. C., 2003.- The molecular biology
of knockdown resistance to pyrethroid insecticides.- Insect Bio-
chemistry and Molecular Biology, 33: 563-577.
THOMPSON J. D., HIGGINS D. G., GIBSON T. J., 1994.- CLUSTAL
W: improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position‐specific gap
penalties and weight matrix choice.- Nucleic Acids Research,
22: 4673‐4680.
TRENTINI M., GOVERNATORI M., POCATERRA C., RUGGERI L.,
1999.- Monitoraggio delle mosche in tre caseifici della provin-
cia di Bologna.- Industrie alimentari, 38: 261-282.
WHEELAN S. J., CHURCH D. M., OSTELL J. M., 2001.- Spidey: a
tool for mRNA-to-genomic alignments.- Genome Research, 11:
1952-1957.
WILLIAMSON M. S., MARTINEZTORRES D., HICK C. A., DEVON-
SHIRE A. L., 1996.- Identification of mutations in the housefly
para-type sodium channel gene associated with knockdown re-
sistance (kdr) to pyrethroid insecticides.- Molecular and Gener-
al Genetics, 252: 51-60.
WINPISINGER K. A., FERKETICH A. K., BERRY R. L., MOESCH-
BERGER M. L., 2005.- Spread of Musca domestica (Diptera: Mus-
cidae), from two caged layer facilities to neighboring residences
in rural Ohio.- Journal of Medical Entomology, 42: 732-738.
Authors’ addresses: Emanuele MAZZONI (corresponding
author: [email protected]), Olga CHIESA, Vincenzo
PUGGIONI, Michela PANINI, Dipartimento di Scienze delle
Produzioni Vegetali Sostenibili - Area Protezione sostenibile
delle piante e degli alimenti, Università Cattolica del Sacro
Cuore, via Emilia parmense 84, 29122 Piacenza, Italy; Gian
Carlo MANICARDI, Dipartimento di Scienze della Vita, Uni-
versità di Modena e Reggio Emilia, Reggio Emilia, Italy;
Davide BIZZARO, Istituto di Biologia e Genetica, Università
Politecnica delle Marche, Ancona, Italy.
Received July 18, 2013. Accepted March 13, 2015.