pregnancy/preeclampsia - semantic scholar filepregnancy/preeclampsia transcriptionally active...
TRANSCRIPT

Pregnancy/Preeclampsia
Transcriptionally Active Syncytial Aggregates in theMaternal Circulation May Contribute to Circulating Soluble
Fms-Like Tyrosine Kinase 1 in PreeclampsiaAugustine Rajakumar, Ana Sofia Cerdeira, Sarosh Rana, Zsuzsanna Zsengeller, Lia Edmunds,
Arun Jeyabalan, Carl A. Hubel, Isaac E. Stillman, Samir M. Parikh, S. Ananth Karumanchi
See Editorial Commentary, pp 191–193
Abstract—The cardinal manifestations of the pregnancy-specific disorder preeclampsia, new-onset hypertension, andproteinuria that resolve with placental delivery have been linked to an extracellular protein made by the placenta, solublefms-like tyrosine kinase 1 (sFlt1), that injures the maternal vasculature. However, the mechanisms by which sFlt1, whichis heavily matrix bound, gain access to the systemic circulation remain unclear. Here we report that the preeclampticplacenta’s outermost layer, the syncytiotrophoblast, forms abundant “knots” that are enriched with sFlt1 protein. Thesesyncytial knots easily detach from the syncytiotrophoblast, resulting in free, multinucleated aggregates (50–150 �mdiameter) that are loaded with sFlt1 protein and mRNA, are metabolically active, and are capable of de novo genetranscription and translation. At least 25% of the measurable sFlt1 in the third-trimester maternal plasma is bound tocirculating placental microparticles. We conclude that detachment of syncytial knots from the placenta results in free,transcriptionally active syncytial aggregates that represent an autonomous source of sFlt1 delivery into the maternal circulation. Theprocess of syncytial knot formation, shedding of syncytial aggregates, and appearance of placental microparticles in the maternalcirculation appears to be greatly accelerated in preeclampsia and may contribute to the maternal vascular injury that characterizesthis disorder. (Hypertension. 2012;59:256-264.) ● Online Data Supplement
Key Words: syncytial knots � syncytial aggregates � microparticles � sFlt1 � soluble VEGFR1 � preeclampsia
Preeclampsia (PE) affects 5% to 7% of all pregnancies andresults in substantial morbidity both to the mother and the
fetus.1,2 Although new-onset maternal hypertension, protein-uria, and edema are the hallmarks of this disorder, PE leftunchecked can progress to seizures, acute liver injury, anddeath. The only known treatment for PE is delivery of theplacenta. Although the pathogenesis of PE remains incom-pletely understood, the rapid and complete resolution of thisdisease after delivery implicates the placenta as a criticalfactor for the pathogenesis of the disease.
We and others have described a marked elevation ofsoluble fms-like tyrosine kinase 1 (sFlt1 or sVEGFR1) in thecirculation of women with PE that is both proportional to theseverity of the disease and is antecedent to the clinicalmanifestations.3–7 Heterologous expression of sFlt1 in preg-nant rodents is sufficient to recapitulate the major features ofPE.6,8,9 sFlt1 may mediate maternal injury by binding andsequestering tropic growth factors, such as vascular endothe-
lial growth factor and placental growth factor, that arenecessary for the maintenance of normal vascular endothelialfunction.10 Northern analysis of tissue arrays and measure-ment of its concentration in the uterine artery versus veinhave implicated the placenta as the major source for circu-lating sFlt1 during pregnancy.6,11
All of the known sFlt1 isoforms contain heparin-bindingdomains in the third and fourth immunoglobulin loops thatfully account for its strong avidity to the extracellularmatrix.12–15 This chemical property has raised questions abouthow sFlt1 made by placental cells can even gain access to thematernal circulation.16 Having observed that the outermostlayer of the placenta, the syncytiotrophoblast, most stronglyexpresses sFlt117 and that placental material has been noted inthe maternal circulation for decades,18 we hypothesized thatsyncytial fragments shed into the maternal circulation are asignificant source of circulating sFlt1 in PE. We performedexperiments in preeclamptic placentas, third-trimester placen-
Received September 7, 2011; first decision October 3, 2011; revision accepted December 5, 2011.From the Department of Medicine (A.R., A.S.C., S.M.P., S.A.K.), Division of Maternal Fetal Medicine, Department of Obstetrics and Gynecology
(S.R., S.A.K.), and Department of Pathology (Z.Z., I.E.S.), Beth Israel Deaconess Medical Center and Harvard Medical School, Boston, MA; HowardHughes Medical Institute (A.R., S.A.K.), Boston, MA; Gulbenkian Programme for Advanced Medical Education (A.S.C.), Lisbon, Portugal;Magee-Womens Research Institute (L.E., A.J., C.A.H.) and Department of Obstetrics, Gynecology and Reproductive Sciences (A.J., C.A.H.), Universityof Pittsburgh School of Medicine, Pittsburgh, PA.
Correspondence to Augustine Rajakumar, RN380G, 99 Brookline Ave, Boston, MA 02215. E-mail [email protected]© 2012 American Heart Association, Inc.
Hypertension is available at http://hyper.ahajournals.org DOI: 10.1161/HYPERTENSIONAHA.111.182170
256
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from

tal organ cultures, and third-trimester maternal plasma to testthis hypothesis. To circumvent the well-described confusionin terms, we have used “syncytial knots” to describe multi-nucleated structures that are loosely attached to the tips ofplacental villi in situ, “syncytial aggregates” to describedetached multinuclear structures of 50 to 150 �m recoveredfrom placental washes and organ cultures, and “micropar-ticles” to describe products isolated by high-speed centrifu-gation of placental washes, culture medium, or maternalplasma.19
Materials and MethodsStudy Population and Sample Collection ProtocolsBiological samples (plasma and placenta) were collected fromnormal and preeclamptic patients. PE was defined as new onset ofhypertension and proteinuria occurring after 20 weeks of gestation.20
Diagnosis of PE was confirmed by an obstetrician after review of themedical charts of the study participants. Approximately 5 mL ofblood were collected from subjects via venipuncture, centrifuged at3500g for 10 minutes, and the plasma was collected and stored at�80°C without thaw before analysis. For placental studies, severalvillous biopsies (2 cm) were excised from the maternal surfacemidway between the chorionic and basal plates, within 30 minutes ofdelivery, and the decidual layer was carefully removed. A portionwas flash frozen in liquid nitrogen for RNA and protein analysis, andthe remaining villous tissue collected was cut to 0.5 cm and rinsedtwice in 50.0 mL of ice-cold PBS for 2 minutes. After rinsing, aportion of the villous tissue was flash frozen, and another portion ofthe villous tissue was used in explant cultures. The washes werecombined (100 mL total) and filtered using a thin layer of gauze. Thematerial in the filtrate was collected by centrifugation (800 g for 10minutes), and then subjected to red blood cell lysis (red blood celllysis solution, Roche Applied Sciences, Mannheim, Germany). Aftercentrifugation and resuspension again in PBS, a portion of the pelletwas used for microscopy and the remaining flash frozen in liquidnitrogen for mRNA and protein analyses. These human studies wereapproved by the institutional review boards at the Beth IsraelDeaconess Medical Center and at the Magee-Womens ResearchInstitute (Pittsburgh, PA), and subjects gave informed consent.
Immunohistochemistry and Electron MicroscopyExpression of sFlt1 protein in formalin-fixed and paraffin-embeddedsections of placenta was evaluated using an antihuman Flt1 antibodythat recognizes the N-terminal region of Flt1/sFlt1 (1:200 dilution,R&D Systems, Minneapolis, MN) and an ImmPRESS antigoatstaining kit (Vector Laboratories, Burlingame, CA) according topublished protocols.21 Evaluation of the hematoxylin-eosin and sFlt1staining was performed by a single pathologist (I.E.S.) in a blindedfashion. Grading of Flt1/sFlt1 staining was done using the semiquan-titative ordinal scale as follows: 1� (focal trophoblast staining), 2�(�50% of the villous trophoblast showing staining), 3� (51% to90% staining), and 4� (�90% staining). Weak staining was con-sidered negative (0).
Contents of the placental washes were fixed in 10% neutral-buffered formalin and then washed with PBS. Samples were centri-fuged at 800 g for 10 minutes at 4°C and were placed into Histogelspecimen medium and then embedded into paraffin. Four-microme-ter sections were cut and stained for sFlt1 expression as describedabove. In addition, pellets of the placental washes were stained withTrypan blue and imaged at 10� and 40� magnifications using anOlympus digital camera.
For electron microscopy studies, placental wash effluents werefixed in 3% formaldehyde 3% glutaraldehyde in 0.1 mol/L ofcacodylate buffer (pH 7.35; Tousismis Research Corporation Rock-ville, MD). The fixed cells/debris was processed in standard fashion(Epon embedded) for transmission electron microscopy, and imageswere acquired using a JEOL 1011 Transmission Electron Micro-
scope, with a Hamamatsu Orca-HR Digital Camera (AdvancedMicroscopy Techniques, Woburn, MA).
Northern Blot AnalysisTotal RNA was isolated from archived placental wash pellets usingRNAwiz (Ambion, Austin, TX), and Northern blot analysis wasperformed using total RNA (20 �g) isolated from the washes asdescribed previously.22 Two regions of the FLT1 mRNA (Gen-eBank Accession No. X51602) spanning the region 250 to 881 (5�ATGGTCAGCTACTGGGACACCGGGGTC and 5� ACTGTT-GCTTCACAGGTCAGAAGC, respectively) and 2300 to 3300 (5�CTAATGGTGTCCCCGAGCCT and 5� CCATTTGTACTCCT-GGGTATGG) were amplified using PCR and were used as probesin the Northern blots using published protocols.
Villous Explant Cultures and Isolation ofSyncytial AggregatesPlacental villous explant preparation and culture were carried outaccording to published protocols.23 Explants were incubated at 37°Cfor 24 hours on an orbital shaker (60 rpm, Belly Dancer, Stovall LifeScience Inc, Greensboro, NC) under standard tissue culture condi-tions in a cell culture incubator (Napco Series 8000 WJ, ThermoScientific, Marietta, OH). At the end of the incubation period, theexplants were removed, blotted with sterile cotton gauze to removeany excess media, and flash frozen in liquid nitrogen and stored at�80°C. To isolate syncytial aggregates, we cultured villous explantson Netwells (15 mm Netwell insert with 500 �m mesh size, Corning)as described.24 To evaluate for shed products, the placental materialcollected in the lower chamber was concentrated by centrifugation at800 g for 5 minutes and used in further experiments.
Ultracentrifugation of Plasma and ExplantCulture MediumWe subjected plasma samples obtained from normal pregnant(n�12) and preeclamptic women (n�16) to ultracentrifugation(100 000 rpm for 90 minutes, �415 000g) after diluting them 10times with ELISA assay calibration buffer (R&D Systems,Minneapolis, MN) or PBS. Similarly, the villous explant condi-tioned medium was centrifuged for 5 minutes at 800 g to removelarger particles and diluted 10 times with PBS and processed forultracentrifugation.
Heparin-Agarose Enrichment of sFlt1 andWestern Blot AnalysessFlt1 in human plasma samples and conditioned medium fromexplant cultures was concentrated by heparin-agarose affinity chro-matography using published protocols25 and Western blots per-formed as described previously.22 Briefly, the 100 000 rpm pelletswere resuspended in 1 mL of PBS and were incubated with 25 �L ofheparin-agarose beads (Sigma Chemical Company, St Louis, MO) at4°C for 1 hour with continuous mixing. The heparin-agarose/sFlt1conjugate was then centrifuged and the pellet washed 3 times withPBS buffer. After the final wash, the beads were resuspended inminimal volume of 1X Laemmli solution and Western blots per-formed using mouse monoclonal vascular endothelial growth factorR1 antibody (V4262; Sigma Chemical Company) that recognizes theamino acid terminus epitope present in both Flt1 and sFlt1.
ELISA and Placental Alkaline PhosphataseColorimetric AssaysFlt1 in culture medium and in maternal plasma, before and after100 000 centrifugation, was measured by ELISA using the humanvascular endothelial growth factor R1 Quantakine kit from R&DSystems (R&D Systems), following the manufacturer’s instructions.Sensitivity of the assay was 5.01 pg/mL, with an intra-assaycoefficient of variation of 2.6% to 3.8% and an interassay coefficientof variation of 7.0% to 8.1%. Placental alkaline phosphatase activityin the 100 000-pellet fraction (same fraction as used in Western blot)was estimated using a kit from Abcam (ab83369-500, Abcam,
Rajakumar et al Syncytial Microparticles and sFlt1 257
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from

Cambridge, MA) according to the manufacturers instructions. Theplacental alkaline phosphatase activity was expressed as micromolesof p-nitrophenol release.
Adenoviral Expression StudiesPlacental explants were incubated for 24 to 48 hours on a Netwell(Corning Life Sciences, Pittston, PA) that facilitates any cells anddebris to move in to the lower chamber.24 The medium was collectedand centrifuged at 800 g to harvest the debris. After red blood celllysis, the concentrated debris were transduced with 2 �L of 1�1010
plaque-forming units per milliliter adenovirus carrying green fluo-rescent protein (Vector BioLabs, Eagleville, PA) and assessed forgreen fluorescent protein expression after 12 to 24 hours followingpublished protocols.24 Briefly, the debris was collected by centrifu-gation, resuspended in 30 to 50 �L of PBS, and spread on amicroscopic slide and let air dry. The slides were dipped in ice-coldacetone for 30 minutes for fixation and PBS, and the nuclei werestained with 4�,6-diamidino-2-phenylindole blue (Molecular ProbesInc, Eugene, OR). The slides were mounted using Gelvetol, fluores-cence microscopy was done using an Olympus digital camera, andthe images were processed using the DP2-BSW program. In anotherexperiment, the debris was transduced with adenovirus expressingtruncated mouse sFlt1 protein26 for 48 hours. The medium wassubjected to heparin-agarose enrichment and probed for sFlt1 ex-pression using Western blots as described above.
ResultssFlt1 Is Highly Expressed in Syncytial KnotsWithin Placentas of Women With PECompared with normal third-trimester placentas (N�9),Flt1&sFlt1 expression was significantly increased in thesyncytial layer of preeclamptic placentas (N�12), with pro-nounced overexpression in syncytial knots (Figure 1A and1B). Furthermore, the abundance of such knots was increasedin preeclamptic placentas. Blinded scoring confirmed thatFlt1&sFlt1 staining was markedly enhanced in the syncytiallayer of preeclamptic placentas (Figure 1C; P�0.001).
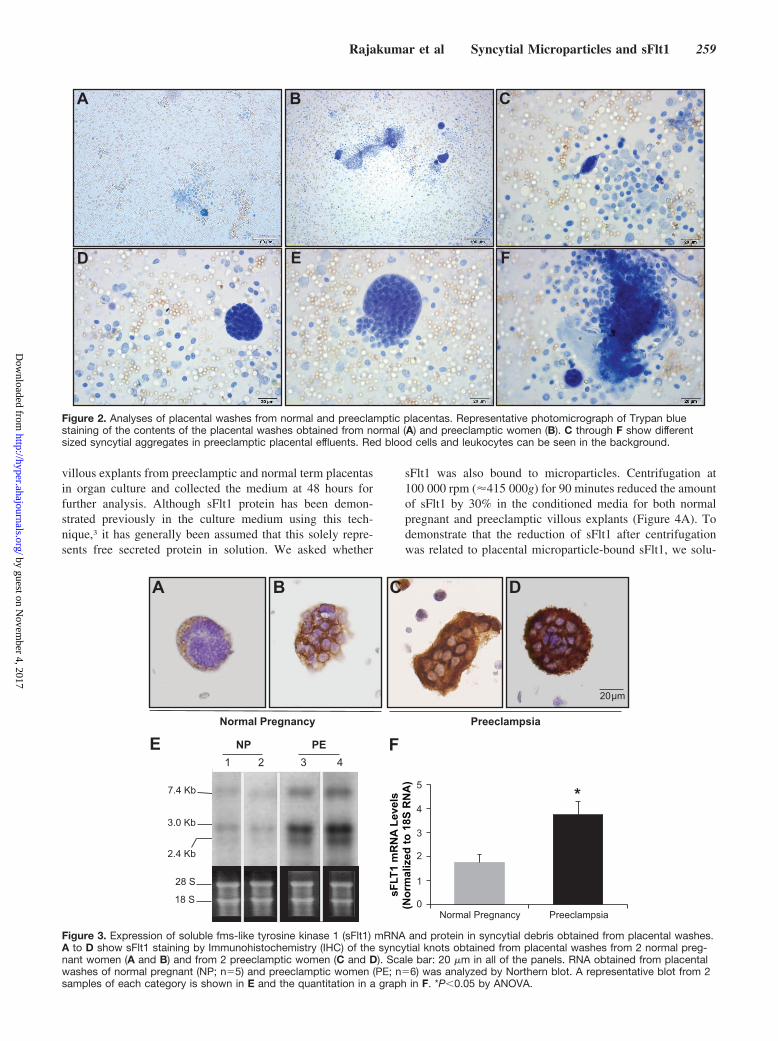
Washing of Preeclamptic Placentas ReleasesSyncytial AggregatesBased on the anatomic orientation of the syncytial knots atthe tips of placental villi, we hypothesized that they mayeasily detach from this outermost layer of the placenta. Totest this, we gently flushed placentas from preeclampticpregnancies with PBS and collected the effluent. The effluentcontained large structures of which the size of 50 to 150 �m
was consistent with detached syncytial knots (Figure 2).Closer examination demonstrated that these structures werealways membrane bound and always multinucleated, againconsistent with a syncytial origin (Figure 2B through 2F). Werepeated this experiment in placentas from normal termpregnancies and observed very few multinucleated structuresin the effluent (Figure 2A). These data showed that theabundant syncytial knots in preeclamptic placentas mighteasily detach from the syncytial layer to become free aggre-gates of syncytial origin.
sFlt1 mRNA and Protein Are Elevated inPreeclamptic Placental EffluentsHaving previously observed that gentle washing of pre-eclamptic placentas reduced the abundance of sFlt1 mRNA,22
we now asked whether the released syncytial aggregates inthe effluent contained sFlt1. To test this, we first performedimmunohistochemistry on syncytial aggregates isolated bylow speed spin (800 g) of placental washes from normal andpreeclamptic third-trimester placentas and observed that thelatter stained much stronger for sFlt1 (Figure 3A through 3D)and the related angiogenic protein endoglin (see Figure S1,available in the online Data Supplement athttp://hyper.ahajournals.org).
Next, we performed Northern analysis to quantify therelative amount of sFlt1 mRNA associated with this liberatedplacental material. Both normal and preeclamptic placentaleffluents demonstrated the 7.4-kb band that corresponds withmembrane-bound Flt1 and the 2 smaller bands (3.0 and 2.4kb) corresponding with alternatively spliced products encod-ing soluble forms of Flt1 (Figure 3E), but the relativeamounts of sFlt1-encoding bands were markedly higher inpreeclamptic placental effluents (Figure 3F). Finally, sFlt1-encoding mRNA was not significantly altered in peripheralblood mononuclear cells obtained from preeclamptic sub-jects, suggesting that loss of peripheral blood cells wasunlikely to account for the reduction in sFlt1 mRNA follow-ing placental washes (see Figure S2).
sFlt1 Is Associated With Microparticles in VillousExplant Culture MediumTo rule out the possibility that the observed syncytial aggre-gates were artifacts of physical manipulation, we next placed
Nor
mal
Pre
gnan
cy
Pree
clam
psia
A B
(400 X)
C
Num
ber o
f Sub
ject
s
Flt1/sFlt1 Staining Intensity
0
1
2
3
4
5
6
7
0 1+ 2+ 3+ 4+
NP (n=9)PE (n=12)
(400 X)
Figure 1. Immunohistochemistry for soluble fms-like tyrosine kinase 1 (Flt1&sFlt1) expression in normal and preeclamptic placentas.Immunohistochemical staining and analysis of placental tissues from normal pregnancy (n�9) and preeclampsia (n�12) for Flt1&sFlt1expression were performed. A and B show a representative staining of normal and preeclamptic placenta, at term, respectively. Thered arrowheads represent syncytial knots. Magnification: �400. C shows a graphical representation of the quantitation of the Flt1&sFlt1staining.
258 Hypertension February 2012
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from
villous explants from preeclamptic and normal term placentasin organ culture and collected the medium at 48 hours forfurther analysis. Although sFlt1 protein has been demon-strated previously in the culture medium using this tech-nique,3 it has generally been assumed that this solely repre-sents free secreted protein in solution. We asked whether
sFlt1 was also bound to microparticles. Centrifugation at100 000 rpm (�415 000g) for 90 minutes reduced the amountof sFlt1 by 30% in the conditioned media for both normalpregnant and preeclamptic villous explants (Figure 4A). Todemonstrate that the reduction of sFlt1 after centrifugationwas related to placental microparticle-bound sFlt1, we solu-
A B C
D E F
Figure 2. Analyses of placental washes from normal and preeclamptic placentas. Representative photomicrograph of Trypan bluestaining of the contents of the placental washes obtained from normal (A) and preeclamptic women (B). C through F show differentsized syncytial aggregates in preeclamptic placental effluents. Red blood cells and leukocytes can be seen in the background.
F
0
1
2
3
4
5
Normal Pregnancy Preeclampsia
*
28 S
18 S
7.4 Kb
3.0 Kb
2.4 Kb
NP PE1 2 3 4
ENormal Pregnancy Preeclampsia
20 μm
A B C D
sFLT
1 m
RN
A Le
vels
(Nor
mal
ized
to 1
8S R
NA
)
Figure 3. Expression of soluble fms-like tyrosine kinase 1 (sFlt1) mRNA and protein in syncytial debris obtained from placental washes.A to D show sFlt1 staining by Immunohistochemistry (IHC) of the syncytial knots obtained from placental washes from 2 normal preg-nant women (A and B) and from 2 preeclamptic women (C and D). Scale bar: 20 �m in all of the panels. RNA obtained from placentalwashes of normal pregnant (NP; n�5) and preeclamptic women (PE; n�6) was analyzed by Northern blot. A representative blot from 2samples of each category is shown in E and the quantitation in a graph in F. *P�0.05 by ANOVA.
Rajakumar et al Syncytial Microparticles and sFlt1 259
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from

bilized the pellet and assayed for sFlt1 and placental alkalinephosphatase. Pellets obtained from culture medium frompreeclamptic women showed significantly higher amounts ofsFlt1 (Figure 4B) and higher placental alkaline phosphatasethan their normal counterparts (Figure 4C). These resultsshow that shedding of the placental microparticle materialcan occur spontaneously rather than being a simple artifact ofaggressive flushing and that this release is exaggerated inpreeclamptic placentas. Moreover, these data suggest thatapproximately one third of the secreted sFlt1 is associatedwith released microparticles.
Ex Vivo Organ Culture Recapitulates SyncytialKnot Formation and Spontaneous SyncytialAggregate ReleaseTo evaluate further the nature of syncytial material beingreleased by the preeclamptic placenta, we performed third-trimester placental organ cultures on Netwell inserts andcollected the culture medium below the mesh for further
characterization (Figure 5A). This technique has previouslyonly been used for the isolation of microparticles fromfirst-trimester placental organ culture.24 Cytological analysisof the culture medium below the mesh revealed membrane-bound knot-like structures containing multiple nuclei thatappear to be “budding” off a main branch (Figure 5B). Wealso observed free membrane-bound particles 50 to 150 �min diameter containing multiple nuclei (Figure 5C and 5D).Both the size and composition of these aggregates exactlymirrored the contents of effluents from placentas (Figure 2)and the spontaneously released material in villous explantcultures of preeclamptic placentas (Figure 3). These results,therefore, suggest that ex vivo culture of third-trimesterplacentas on Netwell results provides an efficient method forstudying the spontaneous release of syncytial aggregates.
Released Syncytial Aggregates Are Viable andMetabolically ActiveWe applied the Netwell method on third-trimester placentalorgan culture to collect syncytial aggregates for further study.Several previous reports have termed these structures “de-bris,”27 implying that they are dead or nonviable material.Transmission electron microscopy of these structures showedthat the plasma membrane was organized into a microvillousstructure and that the cytoplasm contained several nuclei,
A
*
*
Solu
ble
Flt n
g /m
l (EL
ISA
)
0
20000
40000
60000
80000
100000
120000
Pre- Post Pre- Post
Normal Pregnancy Preeclampsia
B
sFlt1 -13
sFlt1 -14150 Kd
100 Kd
Normal Pregnancy Preeclampsia
0
20
40
60
80
100
120
140
Normal Pregnancy Preeclampsia
Plac
enta
l Alk
alin
e Ph
osph
atas
e(μ
mol
of p
-Nitr
ophe
nol
rele
ased
)
*
C
Figure 4. Microparticle associated soluble fms-like tyrosinekinase 1 (sFlt1) in the culture medium of placental villousexplants. Placental villous explants were cultured as describedin the Materials and Methods section, and supernatant was ana-lyzed for sFlt1 expression. High-speed centrifugation reducedsFlt1 by �30% in placental explant conditioned medium. Theprespin and postspin sFlt1 levels measured by ELISA are shownin A. sFlt1 in 100 000 rpm pellets was detected by Western blotanalyses (B). The quantitation of placental alkaline phosphatase(PlAP) by densitometry from the Western blots is shown in C.*P�0.05 by ANOVA.
A
Netwell insert
Villous tissue
Mesh (500 μm)
Syncytial knots/debris
B
C D
→
→
Figure 5. Characterization of placental syncytial aggregatesusing ex vivo organ cultures. A is a schematic of the ex vivoplacental organ cultures on Netwell inserts. B shows the gener-ation of syncytial knots of different sizes from the main villoustissue. Arrow shows the break point of multinucleated aggre-gates from the villous tissue. C and D show representative high-power images of individual multinucleated aggregates that haveseparated from the villous tissue.
260 Hypertension February 2012
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from
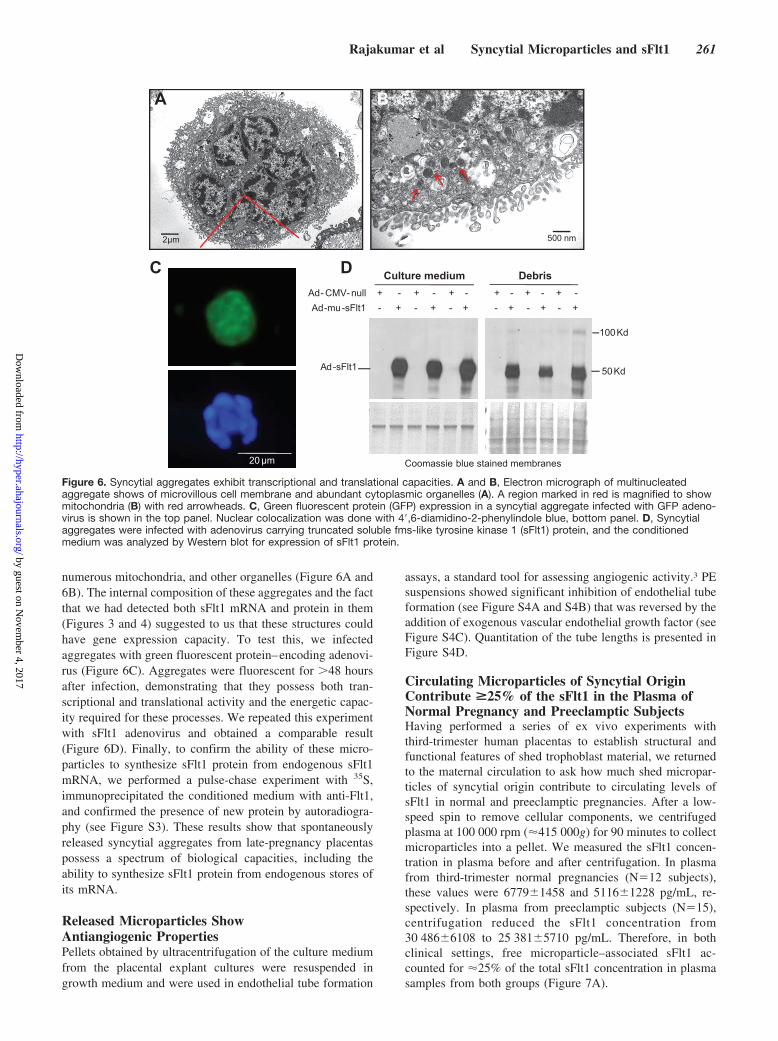
numerous mitochondria, and other organelles (Figure 6A and6B). The internal composition of these aggregates and the factthat we had detected both sFlt1 mRNA and protein in them(Figures 3 and 4) suggested to us that these structures couldhave gene expression capacity. To test this, we infectedaggregates with green fluorescent protein–encoding adenovi-rus (Figure 6C). Aggregates were fluorescent for �48 hoursafter infection, demonstrating that they possess both tran-scriptional and translational activity and the energetic capac-ity required for these processes. We repeated this experimentwith sFlt1 adenovirus and obtained a comparable result(Figure 6D). Finally, to confirm the ability of these micro-particles to synthesize sFlt1 protein from endogenous sFlt1mRNA, we performed a pulse-chase experiment with 35S,immunoprecipitated the conditioned medium with anti-Flt1,and confirmed the presence of new protein by autoradiogra-phy (see Figure S3). These results show that spontaneouslyreleased syncytial aggregates from late-pregnancy placentaspossess a spectrum of biological capacities, including theability to synthesize sFlt1 protein from endogenous stores ofits mRNA.
Released Microparticles ShowAntiangiogenic PropertiesPellets obtained by ultracentrifugation of the culture mediumfrom the placental explant cultures were resuspended ingrowth medium and were used in endothelial tube formation
assays, a standard tool for assessing angiogenic activity.3 PEsuspensions showed significant inhibition of endothelial tubeformation (see Figure S4A and S4B) that was reversed by theaddition of exogenous vascular endothelial growth factor (seeFigure S4C). Quantitation of the tube lengths is presented inFigure S4D.
Circulating Microparticles of Syncytial OriginContribute >25% of the sFlt1 in the Plasma ofNormal Pregnancy and Preeclamptic SubjectsHaving performed a series of ex vivo experiments withthird-trimester human placentas to establish structural andfunctional features of shed trophoblast material, we returnedto the maternal circulation to ask how much shed micropar-ticles of syncytial origin contribute to circulating levels ofsFlt1 in normal and preeclamptic pregnancies. After a low-speed spin to remove cellular components, we centrifugedplasma at 100 000 rpm (�415 000g) for 90 minutes to collectmicroparticles into a pellet. We measured the sFlt1 concen-tration in plasma before and after centrifugation. In plasmafrom third-trimester normal pregnancies (N�12 subjects),these values were 67791458 and 51161228 pg/mL, re-spectively. In plasma from preeclamptic subjects (N�15),centrifugation reduced the sFlt1 concentration from30 4866108 to 25 3815710 pg/mL. Therefore, in bothclinical settings, free microparticle–associated sFlt1 ac-counted for �25% of the total sFlt1 concentration in plasmasamples from both groups (Figure 7A).
A
2μm
B
500 nm
20 μm
Ad-mu-sFlt1 - + - + - + - + - + - +Ad- CMV- null + - + - + - + - + - + -
Ad-sFlt1 50 Kd
Coomassie blue stained membranes
100 Kd
Culture medium DebrisC D
Figure 6. Syncytial aggregates exhibit transcriptional and translational capacities. A and B, Electron micrograph of multinucleatedaggregate shows of microvillous cell membrane and abundant cytoplasmic organelles (A). A region marked in red is magnified to showmitochondria (B) with red arrowheads. C, Green fluorescent protein (GFP) expression in a syncytial aggregate infected with GFP adeno-virus is shown in the top panel. Nuclear colocalization was done with 4�,6-diamidino-2-phenylindole blue, bottom panel. D, Syncytialaggregates were infected with adenovirus carrying truncated soluble fms-like tyrosine kinase 1 (sFlt1) protein, and the conditionedmedium was analyzed by Western blot for expression of sFlt1 protein.
Rajakumar et al Syncytial Microparticles and sFlt1 261
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from

Next, we performed Western analysis on the centrifugedpellets and observed that preeclamptic plasma was enrichedfor sFlt1 bound to these microparticles (Figure 7B). Finally,to confirm that the sFlt1 in these pellets was of syncytiotro-phoblast origin, we performed coimmunoprecipitation bypulling down the syncytiotrophoblast marker syncytin 1 andblotting for sFlt1 (Figure 7C).
DiscussionOur results show that third-trimester placentas from pre-eclamptic women have more syncytial knots that are moreheavily loaded with sFlt1 protein compared with those fromnormal pregnancies. Gentle flushing of these placentas selec-tively releases more trophoblast mass in the form of syncytialaggregates from the preeclamptic placentas than their normalcounterparts. Liberation of syncytial aggregates could not beattributed to aggressive handling, because preeclamptic pla-centas in organ culture spontaneously released aggregates ofidentical size and multinuclear composition into the medium.This shed material contained both sFlt1 protein and mRNA.Placement of third-trimester placental explants over a
500-�m mesh in organ culture not only enabled efficientisolation of syncytial aggregates but also suggested that thisshed material may arise by syncytial sprouting and fissionfrom the underlying syncytium. Isolated aggregates weremultinuclear, rich in cytoplasmic organelles, and capable ofde novo gene expression, demonstrating that these structureswere not only viable but also biologically active. Finally, wereturned to the third-trimester maternal circulation, where wecould demonstrate that �25% of plasma sFlt1 was associatedwith microparticles. Although placental heparinase upregula-tion has been implicated recently as one factor that maycontribute to the release of sFlt1 into systemic circulation,16
our data suggest that release of syncytial microparticles maybe an important additional factor that contributes to theelevated sFlt1 in human PE.
Based on these results, we speculate that third-trimesterplacentas spontaneously form living syncytial sprouts/knots(Figure 1) that detach from placental villi through fission(Figure 5), liberating membrane-bound multinuclear struc-tures (Figures 2 and 3) that we called syncytial aggregatesthat possess critical biological capacities (Figure 6), includingthe ability to synthesize sFlt1 protein from endogenous storesof mRNA. Because each phase of this sequence is exagger-ated in PE—in situ knots (Figure 1) followed by liberatedsFlt1-expressing syncytial aggregates (Figure 3) that thenperhaps further disaggregate to sFlt1-associated micropar-ticles (Figure 7)—we also speculate that accelerated knot/sprout formation within the placenta may be an early event inPE that enhances the delivery of sFlt1 into the maternalcirculation.
Normal pregnancy is characterized by trophoblast turnoverand shedding, as evidenced by the detection of trophoblasticmicroparticles in the maternal circulation throughout preg-nancy28–32 and by the appearance of detached syncytialaggregates with euchromatic nuclei, labeled “syncytialsprouts,” in term human placentas.33 Based on the currentresults, we speculate that third-trimester placentas spontane-ously form syncytial sprouts/knots (Figure 1) that detachfrom placental villi through fission (Figure 5), liberatingmembrane-bound multinuclear structures (Figures 2 and 3),termed syncytial aggregates, that possess critical biologicalcapacities (Figure 6), including the ability to synthesize sFlt1protein from endogenous stores of mRNA. Therefore, al-though apoptotic, necrotic, or “aponecrotic” placental mate-rial34 loaded with sFlt1 protein may well be released into thematernal circulation, our data suggest that term placentasdeport or shed biologically active syncytial aggregates that, inturn, may release smaller microparticles with similar biolog-ical capacities into the maternal circulation, akin to therelease of platelets from megakaryocytes residing in the bonemarrow.
In addition to demonstrating the biological capacity ofthird-trimester shed trophoblast material, the current findingsalso add to the existing literature in other ways. First,deportation of living placental material, followed by de novotranslation of preexisting mRNA, may be a new mechanismby which sFlt1 is delivered into the maternal circulation.Second, although sFlt1 protein has been identified on circu-lating placental particles35 and an increase in circulating
A
B
sFlt1 -13sFlt1 -14
150 Kd100 Kd
Normal Pregnancy Preeclampsia
Preeclampsia Non - pregnant
HA IP HA IP HA IP HA IP
sFlt1-13
sFlt1-14
100 Kd
145 Kd
C
Normal Pregnancy Preeclampsia
*
*
sFlt1
ng/
ml (
ELIS
A)
0
5000
10000
15000
20000
25000
30000
35000
40000
Pre Post
Figure 7. Microparticle associated soluble fms-like tyrosinekinase 1 (sFlt1) in plasma of pregnant women. A, Plasma sam-ples from normal pregnant (n�12) and preeclamptic women(n�15) at term show an �25% reduction in circulating sFlt1 lev-els after ultracentrifugation. B is a representative Western blotanalysis for sFlt1 protein expression in the 100 000 pellets ofthe plasma obtained from normal and preeclamptic women. Cdemonstrates that sFlt1-containing microparticles also expresssyncytin. Plasma samples from preeclamptic patients (n�2) andnonpregnant women (n�2) were subjected to ultracentrifugation.The 100 000 rpm pellet was precipitated using heparin-agarose(HA) or using syncytin antibody (IP) and Western blot performedwith antibody directed against the N terminus of Flt1. *P�0.05by ANOVA.
262 Hypertension February 2012
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from

particles has been associated with PE,18 no explanation hasbeen proposed for how these particles are formed. This isperhaps because late-pregnancy particles in the maternalcirculation have been assumed to be dead material. To ourknowledge, ours are the first data that physically connectcirculating microparticles to syncytial knots by suggestingthat shed syncytial aggregates are the intermediary form.Third, if syncytial knots give rise to circulating sFlt1-expressing microparticles through the shed aggregates, and ifthese phenomena are quantitatively stronger in PE, our datasuggest that accelerated syncytial knot formation is a proxi-mal event in the pathogenesis of PE, in agreement withprevious reports.16 Regardless, it is unlikely that syncytialknots within the intact placenta simply represent an artifact oftangential sectioning, as has also been proposed.36
Important questions remain for future investigation. Met-abolically active microparticles appear to begin forming inthe first trimester.24 Does PE, therefore, represent the sameprocess, but accelerated? If so, what fraction of peripheralsFlt1 protein is contributed by viable sFlt1-expressing micro-particles versus dead/inactive microparticles that are alreadypreloaded with sFlt1 protein? Also, what PE-specific mech-anisms drive the induction of knot formation, deportation ofsyncytial aggregates, and microparticle generation? Con-versely, could enhanced sFlt1 production somehow triggersyncytial knot formation? Second, we present novel evidencethat living syncytial aggregates arise from syncytial sproutingand fission, but the molecular apparatus of nuclear aggrega-tion and cytokinesis remains to be described. Third, differentmechanisms for sFlt1 export from the placenta have beendescribed, including the shedding of dead syncytial material37
and the release of matrix-bound sFlt1 by matrix-dissolvingenzymes, such as heparinase.16 Although upregulation oraccumulation of heparinase has not been shown to occur inthe preeclamptic human placenta,16 it will be of interest todetermine the relative contributions of these processes tomaternal sFlt1 exposure. Fourth, we have established some ofthe biological capacity of syncytial aggregates by demonstrat-ing de novo gene expression, but other functions, includingregulation of inflammation and immunity, may also beimportant.38 It would also be important to determine whetherproinflammatory stimuli and other factors, such as angioten-sin autoantibodies, that have been linked with PE pathogen-esis may induce syncytial knot and microparticle formation.Finally, it has been suggested that shed syncytial aggregatesget trapped in the capillary beds of lung tissue, where theyfurther undergo disaggregation or apoptosis/necrosis to re-lease the smaller microparticles into the systemic circula-tion.39–41 The relative contribution of these processes to theformation of trophoblast microparticles within the maternalcirculation remains to be determined.
PerspectivesThe present studies show that syncytial knots in the third-trimester placentas may give rise to biologically activemicroparticles that can translate packaged sFlt1 mRNA intoprotein. In turn, this process may be a novel means by whichsFlt1 and other toxic proteins, such as soluble endoglin, maybe delivered into the maternal circulation, where they mediate
the major manifestations of PE. Our findings not only havedirect implications for the care of women and unborn childrenwith this disease but may also advance our understanding offundamental cell biological processes. Future work on thebasic biology of syncytialization may shed clues on themolecular defect in PE.42,43
AcknowledgmentsWe thank Dawn McCullough and Saira Sallaludhin for help insample procurement and Saumya Hegde and Poorna Natarajan fortechnical assistance.
Sources of FundingThis work was funded by National Institutes of Health grants RO3HD055219-03 (to A.R.) and P01 HD030367 (to C.A.H. and A.J.).S.R. is supported by a Harvard Diversity and Community PartnershipFaculty Fellowship Award. The Gulbenkian Programme for Ad-vanced Medical Education is sponsored by Fundacao CalousteGulbenkian, Fundacao Champalimaud, and Ministerio da Saude eFundacao para a Ciencia e Tecnologia (Portugal). S.A.K. is aninvestigator of the Howard Hughes Medical Institute.
DisclosuresS.A.K. is a coinventor of multiple patents related to angiogenicproteins for the diagnosis and therapy of preeclampsia. These patentshave been licensed to multiple companies. S.A.K. reports havingserved as a consultant to Roche and Beckman Coulter and hasfinancial interest in Aggamin LLC. The remaining authors report noconflicts.
References1. Redman CW, Sargent IL. Latest advances in understanding preeclampsia.
Science. 2005;308:1592–1594.2. Sibai B, Dekker G, Kupferminc M. Pre-eclampsia. Lancet. 2005;365:
785–799.3. Ahmad S, Ahmed A. Elevated placental soluble vascular endothelial
growth factor receptor-1 inhibits angiogenesis in preeclampsia. Circ Res.2004;95:884–891.
4. Chaiworapongsa T, Romero R, Espinoza J, Bujold E, Mee Kim Y,Goncalves LF, Gomez R, Edwin S. Evidence supporting a role forblockade of the vascular endothelial growth factor system in the patho-physiology of preeclampsia. Am J Obstet Gynecol. 2004;190:1541–1547.
5. Levine RJ, Maynard SE, Qian C, Lim KH, England LJ, Yu KF,Schisterman EF, Thadhani R, Sachs BP, Epstein FH, Sibai BM, SukhatmeVP, Karumanchi SA. Circulating angiogenic factors and the risk ofpreeclampsia. N Engl J Med. 2004;350:672–683.
6. Maynard SE, Min JY, Merchan J, Lim KH, Li J, Mondal S, LibermannTA, Morgan JP, Sellke FW, Stillman IE, Epstein FH, Sukhatme VP,Karumanchi SA. Excess placental soluble fms-like tyrosine kinase 1(sFlt1) may contribute to endothelial dysfunction, hypertension, and pro-teinuria in preeclampsia. J Clin Invest. 2003;111:649–658.
7. Chaiworapongsa T, Romero R, Kim YM, Kim GJ, Kim MR, Espinoza J,Bujold E, Goncalves L, Gomez R, Edwin S, Mazor M. Plasma solublevascular endothelial growth factor receptor-1 concentration is elevatedprior to the clinical diagnosis of pre-eclampsia. J Matern Fetal NeonatalMed. 2005;17:3–18.
8. Bergmann A, Ahmad S, Cudmore M, Gruber AD, Wittschen P, Lin-denmaier W, Christofori G, Gross V, Gonzalves A, Grone HJ, Ahmed A,Weich HA. Reduction of circulating soluble Flt-1 alleviatespreeclampsia-like symptoms in a mouse model. J Cell Mol Med. 2010;14:1857–1867.
9. Lu F, Longo M, Tamayo E, Maner W, Al-Hendy A, Anderson GD,Hankins GD, Saade GR. The effect of over-expression of sFlt-1 on bloodpressure and the occurrence of other manifestations of preeclampsia inunrestrained conscious pregnant mice. Am J Obstet Gynecol. 2007;196:396e1–396e7.
10. Powe CE, Levine RJ, Karumanchi SA. Preeclampsia, a disease of thematernal endothelium: the role of antiangiogenic factors and implicationsfor later cardiovascular disease. Circulation. 2011;123:2856–2869.
Rajakumar et al Syncytial Microparticles and sFlt1 263
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from

11. Bujold E, Romero R, Chaiworapongsa T, Kim YM, Kim GJ, Kim MR,Espinoza J, Goncalves LF, Edwin S, Mazor M. Evidence supporting thatthe excess of the sVEGFR-1 concentration in maternal plasma in pre-eclampsia has a uterine origin. J Matern Fetal Neonatal Med. 2005;18:9–16.
12. Holash J, Davis S, Papadopoulos N, Croll SD, Ho L, Russell M, BolandP, Leidich R, Hylton D, Burova E, Ioffe E, Huang T, Radziejewski C,Bailey K, Fandl JP, Daly T, Wiegand SJ, Yancopoulos GD, Rudge JS.VEGF-Trap: a VEGF blocker with potent antitumor effects. Proc NatlAcad Sci U S A. 2002;99:11393–11398.
13. Kendall RL, Thomas KA. Inhibition of vascular endothelial cell growthfactor activity by an endogenously encoded soluble receptor. Proc NatlAcad Sci U S A. 1993;90:10705–10709.
14. Park M, Lee ST. The fourth immunoglobulin-like loop in the extracellulardomain of FLT-1, a VEGF receptor, includes a major heparin-bindingsite. Biochem Biophys Res Commun. 1999;264:730–734.
15. Sela S, Itin A, Natanson-Yaron S, Greenfield C, Goldman-Wohl D, YagelS, Keshet E. A novel human-specific soluble vascular endothelial growthfactor receptor 1: cell-type-specific splicing and implications to vascularendothelial growth factor homeostasis and preeclampsia. Circ Res. 2008;102:1566–1574.
16. Sela S, Natanson-Yaron S, Zcharia E, Vlodavsky I, Yagel S, Keshet E.Local retention versus systemic release of soluble VEGF receptor-1 aremediated by heparin-binding and regulated by heparanase. Circ Res.2011;108:1063–1070.
17. Nevo O, Soleymanlou N, Wu Y, Xu J, Kingdom J, Many A, Zamudio S,Caniggia I. Increased expression of sFlt-1 in in vivo and in vitro modelsof human placental hypoxia is mediated by HIF-1. Am J Physiol RegulIntegr Comp Physiol. 2006;291:R1085–R1093.
18. Redman CW, Sargent IL. Circulating microparticles in normal pregnancyand pre-eclampsia. Placenta. 2008;29(suppl A):S73–S77.
19. Askelund KJ, Chamley LW. Trophoblast deportation part I: Review ofthe evidence demonstrating trophoblast shedding and deportation duringhuman pregnancy. Placenta. 2011;32:716–723.
20. National Institutes of Health. Report of the National High Blood PressureEducation Program Working Group on High Blood Pressure inPregnancy. Am J Obstet Gynecol. 2000;183:S1–S22.
21. Silasi M, Rana S, Powe C, Cohen B, Lim KH, Zsengeller ZK, Karu-manchi SA, Stillman IE. Placental expression of angiogenic factors inTrisomy 13. Am J Obstet Gynecol. 2011;204:546.
22. Rajakumar A, Michael HM, Rajakumar PA, Shibata E, Hubel CA, Karu-manchi SA, Thadhani R, Wolf M, Harger G, Markovic N. Extra-placentalexpression of vascular endothelial growth factor receptor-1, (Flt-1) andsoluble Flt-1 (sFlt-1), by peripheral blood mononuclear cells (PBMCs) innormotensive and preeclamptic pregnant women. Placenta. 2005;26:563–573.
23. Rajakumar A, Doty K, Daftary A, Harger G, Conrad KP. Impairedoxygen-dependent reduction of HIF-1� and -2� proteins in pre-eclampticplacentae. Placenta. 2003;24:199–208.
24. Abumaree MH, Stone PR, Chamley LW. An in vitro model of humanplacental trophoblast deportation/shedding. Mol Hum Reprod. 2006;12:687–694.
25. Rajakumar A, Powers RW, Hubel CA, Shibata E, von Versen-Hoynck F,Plymire D, Jeyabalan A. Novel soluble Flt-1 isoforms in plasma andcultured placental explants from normotensive pregnant and preeclampticwomen. Placenta. 2009;30:25–34.
26. Kuo CJ, Farnebo F, Yu EY, Christofferson R, Swearingen RA, Carter R,von Recum HA, Yuan J, Kamihara J, Flynn E, D’Amato R, Folkman J,Mulligan RC. Comparative evaluation of the antitumor activity of anti-angiogenic proteins delivered by gene transfer. Proc Natl Acad Sci U S A.2001;98:4605–4610.
27. Redman CW, Sargent IL. Placental debris, oxidative stress and pre-eclampsia. Placenta. 2000;21:597–602.
28. Aharon A, Brenner B. Microparticles and pregnancy complications.Thromb Res. 2011;127(suppl 3):S67–S71.
29. Germain SJ, Sacks GP, Sooranna SR, Sargent IL, Redman CW. Systemicinflammatory priming in normal pregnancy and preeclampsia: the role ofcirculating syncytiotrophoblast microparticles. J Immunol. 2007;178:5949–5956.
30. Kumpel B, King MJ, Sooranna S, Jackson D, Eastlake J, Cheng R,Johnson M. Phenotype and mRNA expression of syncytiotrophoblastmicroparticles isolated from human placenta. Ann N Y Acad Sci. 2008;1137:144–147.
31. Than NG, Abdul Rahman O, Magenheim R, Nagy B, Fule T, Hargitai B,Sammar M, Hupuczi P, Tarca AL, Szabo G, Kovalszky I, Meiri H, SzillerI, Rigo J Jr, Romero R, Papp Z. Placental protein 13 (galectin-13) hasdecreased placental expression but increased shedding and maternalserum concentrations in patients presenting with preterm pre-eclampsiaand HELLP syndrome. Virchows Arch. 2008;453:387–400.
32. van der Post JA, Lok CA, Boer K, Sturk A, Sargent IL, Nieuwland R. Thefunctions of microparticles in pre-eclampsia. Semin Thromb Hemost.2011;37:146–152.
33. Burton GJ. Deportation of syncytial sprouts from the term humanplacenta. Placenta. 2011;32:96–98.
34. Chamley LW, Chen Q, Ding J, Stone PR, Abumaree M. Trophoblastdeportation: just a waste disposal system or antigen sharing? J ReprodImmunol. 2011;88:99–105.
35. Guller S, Tang Z, Ma YY, Di Santo S, Sager R, Schneider H. Proteincomposition of microparticles shed from human placenta during placentalperfusion: potential role in angiogenesis and fibrinolysis in preeclampsia.Placenta. 2011;32:63–69.
36. Huppertz B. IFPA Award in Placentology Lecture: biology of the pla-cental syncytiotrophoblast–myths and facts. Placenta. 2010;31(suppl):S75–S81.
37. Guller S. Role of the syncytium in placenta-mediated complications ofpreeclampsia. Thromb Res. 2009;124:389–392.
38. Redman CW, Sargent IL. Pre-eclampsia, the placenta and the maternalsystemic inflammatory response: a review. Placenta. 2003;24(supplA):S21–S27.
39. Attwood HD, Park WW. Embolism to the lungs by trophoblast. J ObstetGynaecol Br Commonw. 1961;68:611–617.
40. Johansen M, Redman CW, Wilkins T, Sargent IL. Trophoblast depor-tation in human pregnancy: its relevance for pre-eclampsia. Placenta.1999;20:531–539.
41. Lapaire O, Holzgreve W, Oosterwijk JC, Brinkhaus R, Bianchi DW.Georg Schmorl on trophoblasts in the maternal circulation. Placenta.2007;28:1–5.
42. Fox H. The significance of villous syncytial knots in the human placenta.J Obstet Gynaecol Br Commonw. 1965;72:347–355.
43. Huppertz B. Placental origins of preeclampsia: challenging the currenthypothesis. Hypertension. 2008;51:970–975.
264 Hypertension February 2012
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from

Arun Jeyabalan, Carl A. Hubel, Isaac E. Stillman, Samir M. Parikh and S. Ananth KarumanchiAugustine Rajakumar, Ana Sofia Cerdeira, Sarosh Rana, Zsuzsanna Zsengeller, Lia Edmunds,
Contribute to Circulating Soluble Fms-Like Tyrosine Kinase 1 in PreeclampsiaTranscriptionally Active Syncytial Aggregates in the Maternal Circulation May
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 2012 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/HYPERTENSIONAHA.111.1821702012;59:256-264; originally published online January 3, 2012;Hypertension.
http://hyper.ahajournals.org/content/59/2/256World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://hyper.ahajournals.org/content/suppl/2012/01/03/HYPERTENSIONAHA.111.182170.DC1Data Supplement (unedited) at:
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialHypertensionin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on Novem
ber 4, 2017http://hyper.ahajournals.org/
Dow
nloaded from

Online Supplement
Manuscript Title: Transcriptionally active syncytial aggregates in the maternal
circulation may contribute to circulating sFlt1 in preeclampsia
Figure S1: Expression of endoglin in synytial aggregates
Figure S2: Expression of sFlt1 mRNA in peripheral blood mononuclear cells
Figure S3: De novo protein synthesis in cultures of syncytial aggregates
Figure S4: Endothelial tube formation assay using syncytial debris

Supplement Figure S1
A B
C D
E F
20 μm
Figure S1. Expression of endoglin in syncytial aggregatesPlacental washes obtained from normal (Panels A-B) and preeclamptic placentas (C-F) were analyzed for endoglin expression by immunohistochemistry.

Supplement Figure S2
NP PE PE
28 S
18 S
7.4kB
3.0 kB2.4 kB
PBMC Placenta
Figure S2. sFlt1 expression in peripheral mononuclear cells Northern blot analysis of total RNA isolated from peripheral blood mononuclear cells (PBMCs) women with normal pregnancy and preeclampsia. Five microgram of total RNA from 10 patients in each group were pooled and 50 µg of total RNA per lane was analyzed. As a control 15 µg of total placental RNA from a single preeclamptic patient was used.

Supplementary Figure S3
Figure S3. De novo protein synthesis in cultures of syncytial aggregates
Placental syncytial aggregates obtained from 12 Netwells were pooled and incubated with methionine and cysteine free DMEM with 200 µCi of 35S (Easy TagTM Express 35S Protein Labeling Mix, Perkin Elmer,Waltham, MA) for 72 hrs. Total protein was prepared from the debris pellet in 1X Laemmli buffer (10 mM Tris, pH 6.8, 1% SDS, 10% glycerol) with protease inhibitors. The cell extract and the culture supernatant were diluted 1:1 with RIPA buffer (50 mM Tris HCl, pH 7.4, 150 mM NaCl, 1% NP-40, 0.5% sodium Deoxycholate, 0.1% SDS and protease inhibitors) and precleared using protein A/G agarose (Santa Cruz Biotech) for 30 min. The clarified supernatants were incubated with 1.0 µg of human VEGFR1 antibody (mouse monoclonal, V4262, Sigma Chemicals co., St Louis, MO). The immune complexes were captured with protein A/G agarose and after 3 washes with RIPA buffer and the bound proteins were eluted in 1 X Laemmli buffer. Proteins were separated on a SDS containing 8% polyacrylamide gel and transferred on to a PVDF nitrocellulose membrane. After coating the membrane three times with with ENHANCETM(Perkin Elmer, Waltham, MA) the membrane was exposed for autoradiography for 5-7 days.
Immunoprecipitated 35S-labeled sFlt1 from the syncytial aggregrates (lane 1) and supernatant (lane 2) is shown in the figure.
35S-sFlt1
1 2
Autoradiography
100 Kd

0.0
20.0
40.0
60.0
80.0
100.0
120.0
control PE PE+VEGF
Tube
leng
th(%
of c
ontr
ol)
D* *
Control PE Debris PE Debris + VEGF
A B C
Figure S4
Supplementary Figure S4. Endothelial tube formation assay using syncytial debrisTwenty thousand human umbilical vein endothelial cells (HUVEC) were plated onto Matrigel-pre-coated wells of a 48 well plate. These were then incubated at 37 C for 12-16 hours with 200µl of post ultracentrifugation pellets (after brief sonication) from preeclamptic placental explants (n=4) that were resuspended in 200µl of 2.5% serum containing media with or without addition of 200 ng of exogenous VEGF. Tube formation was then quantified using an inverted phase-contrast microscope at X40 (Nikon TE 300, Nikon Corporation). Images were acquired using a Leica DFC350FX camera controlled with LeicaFirecam version 1.5 (Leica Microsystems Imaging Solutions). Total length of tube-like structures per microscopic field was enumerated using Image J freeware (Image J, National Institutes of Health).
A representative micrograph of control and debris treated is shown in Panels A-B. The anti-angiogenic effect of the debris could be reverted by pre-incubating with recombinant VEGF, panel C. Quantitation of the tube formation is shown in Panel D. *p<0.05 by ANOVA.